

Stability Determination of Intact Humanin-G with Characterizations of Oxidation and Dimerization Patterns

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
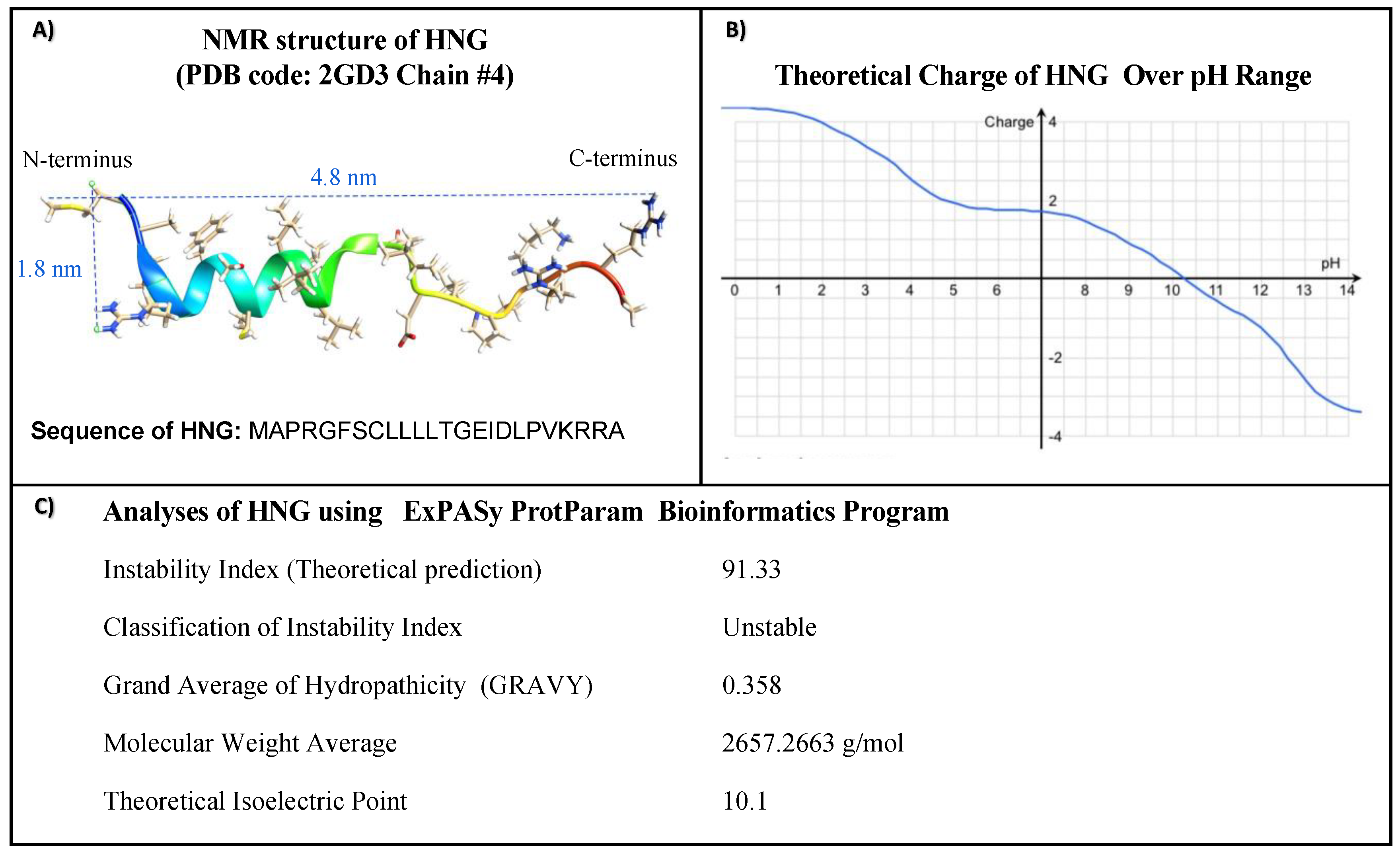
2.2. Physiochemical Properties
2.3. HNG Solution Preparation and Storage
2.4. High-Performance Liquid Chromatography
2.5. High-Resolution Mass Spectrometry
2.6. Data Analysis
3. Results
3.1. Short-Term Stability of HNG Peptide in Different Conditions
3.2. Characterization of Full-Length HNG, Its Oxidized and Dimerized Forms








3.3. Long-Term Stability of HNG Peptide in Different Conditions

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nashine, S.; Cohen, P.; Chwa, M.; Lu, S.; Nesburn, A.B.; Kuppermann, B.D.; Kenney, M.C. Humanin G (HNG) Protects Age-Related Macular Degeneration (AMD) Transmitochondrial ARPE-19 Cybrids from Mitochondrial and Cellular Damage. Cell Death Dis. 2017, 8, e2951. [Google Scholar] [CrossRef] [PubMed]
- Riazi-Esfahani, M.; Kuppermann, B.; Kenney, M. The Role of Mitochondria in AMD: Current Knowledge and Future Applications. J. Ophthalmic Vis. Res. 2017, 12, 424. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Tas, E.; Muzumdar, R. Humanin and Age-Related Diseases: A New Link? Front. Endocrinol. 2014, 5, 210. [Google Scholar] [CrossRef]
- Guo, B.; Zhai, D.; Cabezas, E.; Welsh, K.; Nouraini, S.; Satterthwait, A.C.; Reed, J.C. Humanin Peptide Suppresses Apoptosis by Interfering with Bax Activation. Nature 2003, 423, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Niikura, T.; Tajima, H.; Yasukawa, T.; Sudo, H.; Ito, Y.; Kita, Y.; Kawasumi, M.; Kouyama, K.; Doyu, M.; et al. A Rescue Factor Abolishing Neuronal Cell Death by a Wide Spectrum of Familial Alzheimer’s Disease Genes and Aβ. Proc. Natl. Acad. Sci. USA 2001, 98, 6336–6341. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, I.; Matsuoka, M.; Niikura, T. Unravelling the Role of Humanin. Trends Mol. Med. 2004, 10, 102–105. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Ito, Y.; Niikura, T.; Shao, Z.; Hata, M.; Oyama, F.; Nishimoto, I. Mechanisms of Neuroprotection by a Novel Rescue Factor Humanin from Swedish Mutant Amyloid Precursor Protein. Biochem. Biophys. Res. Commun. 2001, 283, 460–468. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Ishikawa, K.; Spee, C.; Mehta, H.H.; Wan, J.; Yen, K.; Cohen, P.; Kannan, R.; Hinton, D.R. The Mitochondrial-Derived Peptide Humanin Protects RPE Cells From Oxidative Stress, Senescence, and Mitochondrial Dysfunction. Invest. Ophthalmol. Vis. Sci. 2016, 57, 1238. [Google Scholar] [CrossRef]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.-J.; Mehta, H.; Hevener, A.L.; de Cabo, R.; et al. The Mitochondrial-Derived Peptide MOTS-c Promotes Metabolic Homeostasis and Reduces Obesity and Insulin Resistance. Cell Metab. 2015, 21, 443–454. [Google Scholar] [CrossRef]
- Minasyan, L.; Sreekumar, P.G.; Hinton, D.R.; Kannan, R. Protective Mechanisms of the Mitochondrial-Derived Peptide Humanin in Oxidative and Endoplasmic Reticulum Stress in RPE Cells. Oxidative Med. Cell. Longev. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Lee, C.; Wan, J.; Miyazaki, B.; Fang, Y.; Guevara-Aguirre, J.; Yen, K.; Longo, V.; Bartke, A.; Cohen, P. IGF-I Regulates the Age-Dependent Signaling Peptide Humanin. Aging Cell 2014, 13, 958–961. [Google Scholar] [CrossRef] [PubMed]
- Tajima, H.; Niikura, T.; Hashimoto, Y.; Ito, Y.; Kita, Y.; Terashita, K.; Yamazaki, K.; Koto, A.; Aiso, S.; Nishimoto, I. Evidence for in Vivo Production of Humanin Peptide, a Neuroprotective Factor against Alzheimer’s Disease-Related Insults. Neurosci. Lett. 2002, 324, 227–231. [Google Scholar] [CrossRef]
- Men, J.; Zhang, X.; Yang, Y.; Gao, D. An AD-Related Neuroprotector Rescues Transformed Rat Retinal Ganglion Cells from CoCl2-Induced Apoptosis. J. Mol. Neurosci. 2012, 47, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Niikura, T.; Chiba, T.; Aiso, S.; Matsuoka, M.; Nishimoto, I. Humanin: After the Discovery. MN 2004, 30, 327–340. [Google Scholar] [CrossRef]
- Muzumdar, R.H.; Huffman, D.M.; Atzmon, G.; Buettner, C.; Cobb, L.J.; Fishman, S.; Budagov, T.; Cui, L.; Einstein, F.H.; Poduval, A.; et al. Humanin: A Novel Central Regulator of Peripheral Insulin Action. PLoS ONE 2009, 4, e6334. [Google Scholar] [CrossRef] [PubMed]
- Bachar, A.R.; Scheffer, L.; Schroeder, A.S.; Nakamura, H.K.; Cobb, L.J.; Oh, Y.K.; Lerman, L.O.; Pagano, R.E.; Cohen, P.; Lerman, A. Humanin Is Expressed in Human Vascular Walls and Has a Cytoprotective Effect against Oxidized LDL-Induced Oxidative Stress. Cardiovasc. Res. 2010, 88, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Niikura, T.; Ito, Y.; Sudo, H.; Hata, M.; Arakawa, E.; Abe, Y.; Kita, Y.; Nishimoto, I. Detailed Characterization of Neuroprotection by a Rescue Factor Humanin against Various Alzheimer’s Disease-Relevant Insults. J. Neurosci. 2001, 21, 9235–9245. [Google Scholar] [CrossRef]
- Muzumdar, R.H.; Huffman, D.M.; Calvert, J.W.; Jha, S.; Weinberg, Y.; Cui, L.; Nemkal, A.; Atzmon, G.; Klein, L.; Gundewar, S.; et al. Acute Humanin Therapy Attenuates Myocardial Ischemia and Reperfusion Injury in Mice. ATVB 2010, 30, 1940–1948. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chua, C.C.; Gao, J.; Hamdy, R.C.; Chua, B.H.L. Humanin Is a Novel Neuroprotective Agent Against Stroke. Stroke 2006, 37, 2613–2619. [Google Scholar] [CrossRef]
- Oh, Y.K.; Bachar, A.R.; Zacharias, D.G.; Kim, S.G.; Wan, J.; Cobb, L.J.; Lerman, L.O.; Cohen, P.; Lerman, A. Humanin Preserves Endothelial Function and Prevents Atherosclerotic Plaque Progression in Hypercholesterolemic ApoE Deficient Mice. Atherosclerosis 2011, 219, 65–73. [Google Scholar] [CrossRef]
- Takeshita, Y.; Hashimoto, Y.; Nawa, M.; Uchino, H.; Matsuoka, M. SH3-Binding Protein 5 Mediates the Neuroprotective Effect of the Secreted Bioactive Peptide Humanin by Inhibiting c-Jun NH2-Terminal Kinase. J. Biol. Chem. 2013, 288, 24691–24704. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.E.; Cui, L.; Gong, Z.; Su, K.; Muzumdar, R. A Humanin Analog Decreases Oxidative Stress and Preserves Mitochondrial Integrity in Cardiac Myoblasts. Biochem. Biophys. Res. Commun. 2013, 440, 197–203. [Google Scholar] [CrossRef]
- Jin, H.; Liu, T.; Wang, W.-X.; Xu, J.-H.; Yang, P.-B.; Lu, H.-X.; Sun, Q.-R.; Hu, H.-T. Protective Effects of [Gly14]-Humanin on β-Amyloid-Induced PC12 Cell Death by Preventing Mitochondrial Dysfunction. Neurochem. Int. 2010, 56, 417–423. [Google Scholar] [CrossRef]
- Yamagishi, Y.; Hashimoto, Y.; Niikura, T.; Nishimoto, I. Identification of Essential Amino Acids in Humanin, a Neuroprotective Factor against Alzheimer’s Disease-Relevant Insults. Peptides 2003, 24, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, E.; Wickström, M.; Perup, L.S.; Johnsen, J.I.; Eksborg, S.; Kogner, P.; Sävendahl, L. Protective Role of Humanin on Bortezomib-Induced Bone Growth Impairment in Anticancer Treatment. JNCI J. Natl. Cancer Inst. 2014, 106, djt459. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Chang, K.Y.; Yang, J.-R. Analysis and Prediction of Highly Effective Antiviral Peptides Based on Random Forests. PLoS ONE 2013, 8, e70166. [Google Scholar] [CrossRef] [PubMed]
- Enany, S. Structural and Functional Analysis of Hypothetical and Conserved Proteins of Clostridium Tetani. J. Infect. Public Health 2014, 7, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-F.; Lu, Z.-F.; Sun, Y.-N.; Chen, S.-H.; Yi, Y.-J.; Zhang, H.-R.; Yang, S.-Y.; Yu, G.-H.; Huang, L.; Li, C.-N. Molecular Design, Structural Analysis and Antifungal Activity of Derivatives of Peptide CGA-N46. Interdiscip. Sci. Comput. Life Sci. 2016, 8, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, D.; Sánchez, B.; Al-Hassi, H.O.; Mann, E.R.; Urdaci, M.C.; Knight, S.C.; Margolles, A. Microbiota/Host Crosstalk Biomarkers: Regulatory Response of Human Intestinal Dendritic Cells Exposed to Lactobacillus Extracellular Encrypted Peptide. PLoS ONE 2012, 7, e36262. [Google Scholar] [CrossRef]
- Bell, L.N. Peptide Stability in Solids and Solutions. Biotechnol. Prog. 1997, 13, 342–346. [Google Scholar] [CrossRef]
- Mody, R.; Tramontano, A.; Paul, S. Spontaneous Hydrolysis of Vasoactive Intestinal Peptide in Neutral Aqueous Solution. Int. J. Pept. Protein Res. 1994, 44, 441–447. [Google Scholar]
- Taleb, N.; Coriati, A.; Khazzaka, C.; Bayonne, J.; Messier, V.; Rabasa-Lhoret, R. Stability of Commercially Available Glucagon Formulation for Dual-Hormone Artificial Pancreas Clinical Use. Diabetes Technol. Ther. 2017, 19, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Caputo, N.; Castle, J.R.; David, L.L.; Roberts, C.T.; Ward, W.K. Stable Liquid Glucagon Formulations for Rescue Treatment and Bi-Hormonal Closed-Loop Pancreas. Curr. Diabetes Rep. 2012, 12, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Feng, Y.; Zhang, Y.; Kang, W.; Lian, K.; Ai, L. Studies on the Metabolism and Degradation of Vancomycin in Simulated in Vitro and Aquatic Environment by UHPLC-Triple-TOF-MS/MS. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Knoop, A.; Thomas, A.; Thevis, M. Development of a Mass Spectrometry Based Detection Method for the Mitochondrion-Derived Peptide MOTS-c in Plasma Samples for Doping Control Purposes. Rapid Commun. Mass Spectrom. 2019, 33, 371–380. [Google Scholar] [CrossRef]
- Ramkumar, S.; Fan, X.; Wang, B.; Yang, S.; Monnier, V.M. Reactive Cysteine Residues in the Oxidative Dimerization and Cu2+ Induced Aggregation of Human ΓD-Crystallin: Implications for Age-Related Cataract. Biochim. Biophys. Acta-Mol. Basis Dis. 2018, 1864, 3595–3604. [Google Scholar] [CrossRef]
- Schey, K.L.; Finley, E.L. Identification of Peptide Oxidation by Tandem Mass Spectrometry. Acc. Chem. Res. 2000, 33, 299–306. [Google Scholar] [CrossRef]
- Reeg, S.; Grune, T. Protein Oxidation in Aging: Does It Play a Role in Aging Progression? Antioxid. Redox Signal. 2015, 23, 239–255. [Google Scholar] [CrossRef]
- McAuley, A.; Jacob, J.; Kolvenbach, C.G.; Westland, K.; Lee, H.J.; Brych, S.R.; Rehder, D.; Kleemann, G.R.; Brems, D.N.; Matsumura, M. Contributions of a Disulfide Bond to the Structure, Stability, and Dimerization of Human IgG1 Antibody CH3 Domain. Protein Sci. 2007, 17, 95–106. [Google Scholar] [CrossRef]
- Boyson, J.E.; Erskine, R.; Whitman, M.C.; Chiu, M.; Lau, J.M.; Koopman, L.A.; Valter, M.M.; Angelisova, P.; Horejsí, V.; Strominger, J.L. Disulfide Bond-Mediated Dimerization of HLA-G on the Cell Surface. Proc. Natl. Acad. Sci. USA 2002, 99, 16180–16185. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; Durazo, A.; Girotto, S.; Gralla, E.B.; Martinelli, M.; Valentine, J.S.; Vieru, M.; Whitelegge, J.P. Metal-Free Superoxide Dismutase Forms Soluble Oligomers under Physiological Conditions: A Possible General Mechanism for Familial ALS. Proc. Natl. Acad. Sci. USA 2007, 104, 11263–11267. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yamane, H.; Arakawa, T.; Narhi, L.O.; Philo, J. Effect of the Intermolecular Disulfide Bond on the Conformation and Stability of Glial Cell Line-Derived Neurotrophic Factor. Protein Eng. Des. Sel. 2002, 15, 59–64. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos. | Amino Acid | Function | Hydropathicity | Amino Acid Side Chain Properties | Name of Cleaving Enzymes/Chemicals (Theoretical) |
|---|---|---|---|---|---|
| 1 | M | Neuroprotection | 1.900 | Hydrophobic | Chymotrypsin, Cyanogen Bromide |
| 2 | A | Neuroprotection | 1.800 | Hydrophobic | Proteinase K |
| 3 | P* | Neuroprotection | −1.600 | Hydrophobic | N/A |
| 4 | R* | Neuroprotection | −4.500 | Positive Charged (Basic) | Trypsin, Arg-C proteinase, Clostridiopeptidase B |
| 5 | G* | N/D | −0.400 | Hydrophobic | Pepsin, Thermolysin |
| 6 | F* | IGFBP-3 binding, Beta-Amyloid Binding | 2.800 | Hydrophobic | Chymotrypsin, Proteinase K |
| 7 | S* | Beta-Amyloid Binding, Beta-Amyloid Protection, Dimerization | −0.800 | Polar | 2-nitro-5-thiocyanobenzoic acid |
| 8 | C* | Neuroprotection, BAX, BAD, end tBID binding, Disulfide Bond for dimerization | 2.500 | Polar | Pepsin, Thermolysin |
| 9 | L* | Neuroprotection, Secretion, Dimerization | 3.800 | Hydrophobic | Proteinase K, Pepsin, Thermolysin, Chymotrypsin |
| 10 | L* | Secretion | 3.800 | Hydrophobic | Proteinase K, Pepsin, Thermolysin, Chymotrypsin |
| 11 | L* | Secretion | 3.800 | Hydrophobic | Proteinase K, Pepsin, Thermolysin, Chymotrypsin |
| 12 | L* | Neuroprotection | 3.800 | Hydrophobic | Proteinase K, Pepsin, Chymotrypsin |
| 13 | T* | Neuroprotection | −0.700 | Polar | Proteinase K, |
| 14 | G* | Neuroprotection | −0.400 | Hydrophobic | Asp-N Endopeptidase |
| 15 | E* | N/D | −3.500 | Negative Charged (Acidic) | Proteinase K, Glutamyl endopeptidase, Staphylococcal peptidase I |
| 16 | I* | N/D | 4.500 | Hydrophobic | Proteinase K, Asp-N Endopeptidase |
| 17 | D* | N/D | −3.500 | Negative Charged (Acidic) | Formic acid |
| 18 | L* | N/D | 3.800 | Hydrophobic | Proteinase K, Pepsin |
| 19 | P* | Beta-Amyloid Protection, Secretion | −1.600 | Hydrophobic | Thermolysin |
| 20 | V* | Secretion | 4.200 | Hydrophobic | Proteinase K, Peptidyl-Lys metalloendopeptidase |
| 21 | K | IGFBP-3 binding | −3.900 | Positive Charged (Basic) | Lysyl endopeptidase, Trypsin |
| 22 | R | N/D | −4.500 | Positive Charged (Basic) | Arg-C proteinase, Clostridiopeptidase B, Trypsin |
| 23 | R | N/D | −4.500 | Positive Charged (Basic) | Thermolysin, Arg-C proteinase, Clostridiopeptidase B, Trypsin |
| 24 | A | N/D | 1.800 | Hydrophobic | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozgul, M.; Nesburn, A.B.; Nasralla, N.; Katz, B.; Taylan, E.; Kuppermann, B.D.; Kenney, M.C. Stability Determination of Intact Humanin-G with Characterizations of Oxidation and Dimerization Patterns. Biomolecules 2023, 13, 515. https://doi.org/10.3390/biom13030515
Ozgul M, Nesburn AB, Nasralla N, Katz B, Taylan E, Kuppermann BD, Kenney MC. Stability Determination of Intact Humanin-G with Characterizations of Oxidation and Dimerization Patterns. Biomolecules. 2023; 13(3):515. https://doi.org/10.3390/biom13030515
Chicago/Turabian StyleOzgul, Mustafa, Anthony B. Nesburn, Nader Nasralla, Benjamin Katz, Enes Taylan, Baruch D. Kuppermann, and Maria Cristina Kenney. 2023. "Stability Determination of Intact Humanin-G with Characterizations of Oxidation and Dimerization Patterns" Biomolecules 13, no. 3: 515. https://doi.org/10.3390/biom13030515
APA StyleOzgul, M., Nesburn, A. B., Nasralla, N., Katz, B., Taylan, E., Kuppermann, B. D., & Kenney, M. C. (2023). Stability Determination of Intact Humanin-G with Characterizations of Oxidation and Dimerization Patterns. Biomolecules, 13(3), 515. https://doi.org/10.3390/biom13030515