Abstract
Glutathione transferases (GSTs) are ubiquitous key enzymes with different activities as transferases or isomerases. As key detoxifying enzymes, GSTs are expressed in the chemosensory organs. They fulfill an essential protective role because the chemosensory organs are located in the main entry paths of exogenous compounds within the body. In addition to this protective function, they modulate the perception process by metabolizing exogenous molecules, including tastants and odorants. Chemosensory detection involves the interaction of chemosensory molecules with receptors. GST contributes to signal termination by metabolizing these molecules. By reducing the concentration of chemosensory molecules before receptor binding, GST modulates receptor activation and, therefore, the perception of these molecules. The balance of chemoperception by GSTs has been shown in insects as well as in mammals, although their chemosensory systems are not evolutionarily connected. This review will provide knowledge supporting the involvement of GSTs in chemoperception, describing their localization in these systems as well as their enzymatic capacity toward odorants, sapid molecules, and pheromones in insects and mammals. Their different roles in chemosensory organs will be discussed in light of the evolutionary advantage of the coupling of the detoxification system and chemosensory system through GSTs.
Keywords:
insects; glutathione transferase; olfaction; taste; chemosensory organs; detoxification; evolution; flavor 1. Introduction
Evolution shapes all living organisms to perceive chemicals, enabling them to detect nutritive compounds and avoid toxic compounds. Although chemical detection supports several biological functions related to chemical communication in animals, the primary function of this ancient sense was most likely to find nutritive molecules. According to the Red Queen hypothesis [1], it can be hypothesized that the appearance of detoxification systems results from developments in defensive systems, such as the synthesis of toxic compounds. In response to this new pressure of selection, predators have evolved and developed defensive mechanisms capable of taking in toxic xenobiotics. In this context, the extension of the organism’s detoxifying capacity should have also enabled species to extend their food sources due to their new capacity to metabolize xenobiotic compounds within these food sources. Xenobiotics are molecules that are not used as building blocks for biological macromolecules and also do not provide energy. Glutathione transferases (GSTs; EC 2.5.1.18), originally discovered as detoxification enzymes [2,3], are one of the key enzymes involved in the metabolization of endogenous and exogenous molecules. GSTs are found in most living organisms, including insects and mammals. Xenobiotic molecules include chemicals naturally present in food sources as well as molecules produced industrially as pollutants and pesticides. Most flavor molecules can also be considered xenobiotics, regulated by GSTs and other detoxifying enzymes through the xenobiotic detoxifying network [4,5], such as glutathione transferases (GSTs; EC 2.5.1.18). GSTs, originally discovered as detoxication enzymes [2,3], are found in most living organisms, including insects and mammals. The best known function of GST is to catalyze the conjugation of reduced glutathione (GSH) to xenobiotic electrophilic centers, resulting in an increase in the hydrophilicity of xenobiotic compounds and thereby facilitating their elimination from the body. This function is shared by two entirely distinct superfamilies of enzymes [6], one microsomal (membrane-associated eicosanoid and glutathione metabolism, also called MAPEG), and the other soluble, also called canonical. These two superfamilies are not evolutionarily related and are found both in mammals and insects; moreover, in general, the number of microsomal GSTs is much lower than that of canonical GSTs. A third type of GST can be considered: mitochondrial Kappa GST [7]. Both kappa and canonical GSTs present a thioredoxin-like domain recognizing the GSH motif but are differently organized in the overall fold, suggesting their parallel evolution from a thiol-disulfide oxidoreductase progenitor [8]. Kappa GSTs are found in prokaryotes and eukaryotes, including mammals, but not in insects. However, Kappa GSTs have been identified in other arthropods and crustaceans [7,9]. For example, Acari, Ixodes scapularis, presents two Kappa GSTs involved in glutathione conjugation, and canonical GSTs also catalyze peroxide reduction [10], dehydrochlorination [11], and isomerization [12,13]. Moreover, they may trap substrates in the absence of enzymatic activity, which enables the sequestration of toxic compounds; this function is called ligandin [14]. Canonical GSTs are subdivided into classes, designated by the names of the Greek letters—Alpha, Delta, Epsilon, etc.—abbreviated as Roman capitals A, D, E, etc. Class members are distinguished by Arabic numerals [15]. GST members exhibit their own distinct tissue-specific expression patterns, suggesting that they have different functions. This differential pattern of expression is observed both in mammals [16] and insects [17]. It is not surprising that GSTs are preferentially expressed in insect detoxification organs, such as the fat body, midgut, or epidermis [18], and in the mammalian liver [19]. Chemosensory organs are exposed to the external environment and, thus, to xenobiotics. Consequently, GST expression is also observed in these organs, which represent specific places in the animal body. The detoxifying system can advantageously protect the chemosensory system, and additionally or as a result of its activity, it modulates chemosensory detection. The molecular organization of the chemosensory organs has been well described in both mammals and insects (the main laboratory models are rodents and Drosophila). In this review, we detail the knowledge supporting the involvement of GSTs in chemoperception, describing their localization in these systems as well as their enzymatic capacity toward odorants, sapid molecules, and pheromones in insects and mammals.
2. The Involvement of GSTs in Mammalian Chemoperception
2.1. Chemoperception in Mammals
Chemoperception in mammals involves three components: olfaction, taste, and trigeminal sensations [20,21]. Olfaction is a fundamental sense that enables animals to locate their food and their sexual partners and to warn of danger, which is also of great importance for their well-being through the hedonic tone of food. In mammals, olfaction is assured by the olfactory system located in the nasal cavity. Odorant compounds are small, volatile chemicals, generally of a hydrophobic nature, that enter the body through the nostrils and solubilize within the nasal mucus. Odorants bind to olfactory receptors (ORs) located on the olfactory receptor neurons (ORNs) present in the main olfactory epithelium. Indeed, in addition to the olfactory epithelium, mammals have several accessory olfactory organs, such as the vomeronasal organ, the septal organ of Masera, and the Grueneberg ganglion, which all contain ORNs. These organs exhibit overlapping functions with the main olfactory system, but their specificities are not fully characterized. Each olfactory receptor neuron expresses only one type of OR [22]. ORNs are connected to the olfactory bulb, where the olfactory signal is processed and further transmitted to higher brain regions. This organization transforms a chemical signal into an electrical signal thanks to an efficient combinatorial code of odors permitted by the high diversity of olfactory receptors (380 for humans) [23].
In contrast to olfaction, which is important for various functions such as food search and enjoyment, reproduction, and survival, the sense of taste in mammals is exclusively devoted to the evaluation of food quality and is closely related to feeding behavior [24]. Taste compounds are divided into five qualities, namely, sweet (sugars), bitter (various compounds of organic or inorganic nature), sour (acidic compounds), salty (ionic inorganic compounds such as Na+), and umami (amino acids). In mammals, taste sensations result from the activation of taste receptors by taste compounds. Taste receptor cells assemble at the surface of the tongue or palate into small structures called “taste buds”, composed of approximately 100 cells. These taste buds are found in epithelial structures called “papillae”, classified into different types and present different structures and locations at the tongue surface. Papillae include fungiform, circumvallate, and foliate papillae. Taste receptor cells are linked by afferent nerves to the geniculate and petrosal ganglions, which mediate taste signals to the brain stem. Taste receptors from the T1R and T2R families are part of the class C and class A GPCR families, respectively [25]. They assemble through different functional homo- or heterodimers for T1Rs that are able to detect sweet (T1R2 + T1R3) and umami (T1R1 + T1R3) compounds. Monomeric T2Rs enable the detection of bitter molecules. Two other families of receptors have been proposed for the detection of sourness (PKD2L1) and saltiness (ENaC) [24].
In addition to the five qualities of taste, a few studies have evidenced a sixth sensation, which is more of a modulation of some of the basic tastes, namely the kokumi taste [26]. Kokumi means “rich taste” in Japanese and is associated with the reinforcement of umami, sweetness, and saltiness due to the presence of kokumi compounds. Kokumi compounds include both nonpeptide compounds and peptide compounds such as gamma-glutamyl peptides such as glutathione [27]. Kokumi compounds have been proposed to activate the calcium-sensing receptor (CaSR) to elicit the kokumi sensation [28]. Kokumi is an increasingly studied topic of research with great potential for flavor enhancement and food product development [26].
In addition to olfaction and taste, a third chemosensory sensation is related to the trigeminal system and enables the detection of trigeminal compounds such as astringent plant polyphenols, compounds present in foods eliciting a sensation of cooling (menthol) or burning (capsaicin), or carbonated drinks containing CO2 that trigger a prickly sensation [21]. Trigeminal compounds mainly stimulate transient receptor potential (TRP) channels present in sensory neurons of the oral mucosa, which convert chemical signals into electric activity. The identification of astringent compounds may either involve mechanoreceptors detecting changes in the friction forces at the surface of the oral mucosae or the transmembrane mucin MUC1, as recently proposed by our group [29].
Olfaction, taste, and trigeminal sensations can be modulated by the metabolic activities present in the vicinity of the chemosensory receptors [30]. This metabolism involves enzymes such as glutathione transferases (GSTs). These have shown to be linked to both olfaction and taste perception, as explained in the following section.
2.2. Roles of GSTs in Mammalian Chemoperception
Many drug-metabolizing enzymes are present in the nasal cavity, within the nasal epithelium and mucus [31,32], as well as in the oral cavity, within the saliva and the oral mucosa [33,34]. These enzymes assure the protection of the epithelia and, notably, the olfactory receptor neurons from damage provoked by xenobiotics entering the nasal (or oral) cavity. In addition to their role in detoxification, some of these enzymes handle flavor molecules, thus producing flavor metabolites in the oronasal sphere [35,36,37,38]. All these enzymes, in addition to proteins able to bind flavor compounds such as odorant-binding proteins (OBPs), have an impact on both the quality and quantity of flavor compounds that activate chemoreceptors [30]. These molecular mechanisms have been named “perireceptor events” because they occur in the environment surrounding the chemoreceptors. In the context of mammalian olfaction, a body of evidence indicates that glutathione transferase is an important perireceptor enzyme acting on odorant detection modulation. Additionally, pioneering studies have started to investigate the potential role of glutathione transferases in taste perception. Both of these aspects are treated in this part of the review. Among the seven canonical mammalian GSTs identified thus far (alpha, mu, theta, pi, omega, sigma, and zeta [15]), alpha, mu, and pi classes of GSTs are common [39] among mammals [40].
In 1992, Ben-Arie and coworkers showed that the olfactory epithelium is the extrahepatic tissue in a rat model that exhibits the highest glutathione transfer activity with the chemical substrate chlorodinitrobenzene (CDNB), thus showing the strong expression of GSTs and suggesting a potential role in olfaction [41]. All GST classes were shown to be expressed in chemosensory organs in different mammalian species (Table 1). Glutathione transferase of the mu class was demonstrated in the rat nasal mucus and olfactory epithelium [4], supporting a previous study already showing GST expression in rat chemosensory mucosae [42]. Additionally, in the same study, the ability of recombinant rat GSTM2 to catalyze the transfer of glutathione to various odorant compounds, including aldehydes, ketones, and epoxides, was shown. Using a competition assay, GSTM2 was also found to be able to bind a large variety of odorants, most likely due to its ligandin properties [4]. Similar results were obtained for human GSTA1 and GSTP1, which are two GSTs that play roles in glutathione and ligand transfer for many odorous molecules and are expressed in the human respiratory epithelium [43]. Immunohistochemistry experiments showed the localization of human GSTs in ciliated cells (at the surface of the epithelium from the human olfactory vicinity), thus facilitating the entry of odorants into the epithelium. Furthermore, the first X-ray structure of a GST bound to a metabolized odorant enabled a fine analysis of its active site and its capacity to specifically recognize odorant molecules [43]. The crystal structure of human GSTA1 bound to the metabolite glutathionyl-dihydrocinnamaldehyde was analyzed and revealed the ability of the hydrophobic site of GSTA1 to strongly adapt to small hydrophobic volatile compounds. This property facilitates the binding of cinnamaldehyde and promotes the formation of the glutathione conjugate through a nucleophilic substitution, suppressing the carbon double bond of cinnamaldehyde [43].
GST expression levels were also shown to be associated with olfactory dysfunction. Zinc deficiency is linked to olfactory and gustatory dysfunction in mammals [44,45]. Interestingly, in rats, zinc deficiency is associated with a reduction in GST mRNA in some cell types of the olfactory epithelium (supporting cells [46]).
The involvement of GSTs in odorant metabolism has become increasingly documented; however, less evidence is available concerning the significance of these molecular mechanisms in odorant perception. In this context, experiments with a rabbit model enabled us to reveal some clues on the role of GSTs in odorant signal termination. It was shown that the mammary pheromone, corresponding to the compound 2-methylbut-2-enal (2MB2), triggers the grasping of the mother rabbit mammae. Newborn rabbits are blind, and suckling-related behavior allows them to survive despite the short time that allows the mother to feed them [47]. This chemically triggered behavior is critical for pups, which are constrained to finding nipples within the five minutes of daily nursing. It has been shown that the mammary pheromone is metabolized to a glutathione conjugate in the olfactory epithelium of newborn rabbits, in accordance with a high early expression of glutathione transferases in this tissue [48]. Furthermore, it has been shown that this metabolism is also present in the nasal mucus of newborn rabbits due to the presence of glutathione transferases in this biological fluid based on proteomic analysis [49]. Additionally, the deregulation of this metabolism by in vivo washing of the nasal mucus, thus diminishing the glutathione conjugation of 2MB2, led to increased sensitivity of the behavior response in newborns exposed to the mammary pheromone. Similar results were obtained when the 2MB2 metabolism was reduced due to competition with another odorant substrate catalyzed by the same enzyme [50]. Decreasing the glutathione conjugation of the mammary pheromone allowed us to record behavioral responses with concentrations of mammary pheromone that were usually inactive. Glutathione conjugation to the mammary pheromone modifies its structure and thus terminates the odorant signal. This metabolization thus enables the newborn rabbit to remain responsive to the mammary pheromone by reinitializing the chemical signal. Rabbit GSTs catalyze glutathione conjugation with 2MB2 but can ensure the role of odorant signal termination for a wide range of other odorant compounds in mammals, including humans.
In addition to the nasal cavity, GSTs are expressed in the oral cavity, particularly in taste bud cells [51]. In rats, GSTM and GSTP were found to be expressed in both circumvallate and foliate papillae. The expression seems to be GST member-dependent; GSTA is not found in these papillae. Moreover, the results obtained in human and rat olfactory epithelia show differences with regard to the type of GST expressed in this tissue depending on the species, highlighting differences in their expression in chemosensory tissue between mammalian species [4,43]. GST was also found to be expressed in mammalian saliva, including human saliva [52]. Glutathione is found at a concentration of approximately 1 g/L in human saliva [53], enabling the GSH saturation of GSTP, the main human salivary GST, and then allowing it to catalyze glutathione conjugation at the maximal rate. Additionally, this salivary expression was shown to be modulated in humans in association with food behavior. Indeed, increased GST expression in response to specific diets, such as those rich in broccoli or coffee, was observed [52,54]. Recently, a study explored a possible link between GST expression in saliva and bitter taste perception [55]. GSTA1 and GSTP1 were identified in a cohort of 104 people, all of whom expressed the 2 GSTs in saliva. Additionally, people exhibiting ageusia or dysgeusia, included in this study, showed significantly lower salivary GSTA1 levels than those in the saliva of the control group, suggesting possible relationships between salivary GST levels and taste function. In the same study, GSTA1 and GSTP1 interacted with various bitter compounds, including flavonoids and isothiocyanates. This last family of compounds is metabolized within the saliva [55]. The X-ray structures obtained between GSTs and isothiocyanates showed that different binding sites exist, whether the interactions imply glutathione conjugation (binding in the active site) or covalent adduction to an exposed cysteine in the GSTA1 ligand site.
All these elements suggest the involvement of GSTs in mammalian taste perception, in addition to olfaction. In addition, the level of expression of GST is correlated with disorders related to both taste and smell.

Table 1.
Location and classes of GSTs within the mammal’s chemosensory organs.
Table 1.
Location and classes of GSTs within the mammal’s chemosensory organs.
| Mammal Species | Location | GST Classes | Ref. |
|---|---|---|---|
| Canis lupus familiaris | Saliva | Alpha, Mu, and Omega | [56] |
| Equus caballus | Saliva | Pi | [57] |
| Homo sapiens | Olfactory mucus | Alpha and Pi | [58,59] |
| Olfactory epithelium | Alpha, Mu, and Pi | [43] | |
| Saliva | Alpha, Kappa, Mu Omega, Theta, and Pi | [60,61] | |
| Mus musculus | Sensory cilia | Alpha, Kappa, Mu, Omega, Tau, and Zeta | [62] |
| Saliva | Omega | [63] | |
| Oryctolagus cuniculus | Olfactory mucus | Alpha, Mu, and Pi | [49] |
| Ovis aries | Saliva | Alpha | [57] |
| Rattus norvegicus | Sensory cilia | Alpha and Mu | [64] |
| Olfactory epithelium | Alpha, Mu, and Pi | [4,41] | |
| Olfactory mucus | Alpha, Mu, and Pi | [4] |
3. The Involvement of GSTs in Insect Chemoperception
3.1. Chemoperception in Insects
Insects constitute the largest class of living animal species. Due to their small size, they have developed multiple mechanisms to limit toxic xenobiotic effects, including the enhancement of metabolic detoxification [65,66], which reduces penetration through the cuticle, or behavioral avoidance [67]. Insects can taste through many parts of their body. The proboscis organs used for feeding and sucking allow insects to taste food during ingestion before it reaches the digestive system. In addition to the proboscis, insects are able to detect tastants with their legs, wings, and ovipositor organs. Consequently, due to their small size compared with food, they are generally already in contact with it before ingesting it, making it advantageous to taste it with their legs before eating or with the ovipositor organ before laying eggs. Although the taste systems of mammals and insects evolved independently, they enable the detection of similar qualities, including sweet, salty, and bitter stimuli. Insects are able to detect carbonation as a taste modality through gustatory neurons [68], similar to mammals, through carbonic anhydrase IV, which produces protons that activate a proton-gated channel [69]. Interestingly, carbonation detection is also possible in both mammals and insects through the olfactory system [69]. Insects have gustatory sensory neurons that mediate the recognition of water [70]; to date, it has not been established whether other animals can taste the water. In mammals, taste receptors are not hosted by neurons. Neurons are in contact with the taste cells that carry gustatory receptors and are located in taste buds. In insects, gustatory receptors are directly carried by gustatory neurons, in contrast to vertebrate gustatory neurons, which are housed in cells that are indirectly in contact with neurons. Gustatory neurons are housed within the hundreds of gustatory sensilla distributed on the surface of the different sensory organs except the proboscis, which also includes internal sensilla [71].
In insects, the equivalent of the mammalian nose is the antenna and the maxillary palps. Although they do not exhibit any evolutionary relationship with mammals, olfaction is also supported by olfactory neurons in insects. Indeed, the olfactory sensilla cover the distal segment of the antenna, and the maxillary palps host the olfactory neurons. Dendrites of olfactory neurons that express olfactory neurons are located in the sensilla lymph within the sensilla. Odor molecules pass through pores or slits in the sensillum cuticle and enter the sensillum lymph [72]. Insect olfactory receptors are not homologs of vertebrate olfactory receptors [73], suggesting different evolutionary origins compared with those found in vertebrates. Consequently, although the organizational features of the olfactory systems of vertebrates and insects appear very similar, these structures may not share a common evolutionary heritage [74].
Insect GRs and ORs, which are membrane proteins, do not show any homology to those of vertebrates [75,76] and consequently do not belong to the GPCR family. However, both GRs and ORs evolved from an ancestral protein, and in addition to sharing the same sequence identity, they share the same inverted transmembrane topology as vertebrate olfactory GPCRs. The expansion of genes coding for the insect GR and OR has occurred only in insects [77]. In contrast to the monomeric ORs of vertebrates, insect ORs form heteromers with a conserved OR receptor also called Orco (i.e., the OR coreceptor). One specific OR is expressed in each insect olfactory neuron in addition to Orco, as in vertebrate olfactory neurons, where one specific OR is expressed in each olfactory neuron. It is unclear whether insect GRs can function alone as multimers or with other insect GRs due to the observation that multiple GR genes are expressed in a single GR neuron [78].
Even if the chemosensory systems in insects and mammals present similar biological organizations, they do not share any evolutionary links. However, in both chemosensory systems, glutathione transferases from the same common ancestor are expressed, most likely sharing similar physiological roles.
3.2. Roles of GSTs in Insect Chemoperception
In addition to the Omega, Sigma, Theta, and Zeta GST classes found in insects and shared with mammals, two other classes are observed: Delta and Epsilon GSTs. Delta GSTs are found in insects and are observed in a more general manner in arthropods, such as crustaceans [9]. Epsilon GSTs appear more specific to insects and were hypothesized to be insect-specific [79]. The numbers of Delta and Epsilon GSTs are variable from one insect species to another, mostly due to duplication events that occur in each insect species (Table 2). This gene-coding GST duplication might be associated with functional differentiation during insect evolution and is related to environmental adaptation. Gene duplication followed by sequence divergence is a key process during evolution, allowing the creation of novel gene functions [80]. Interactions of insects with plants, and especially plant chemicals and their adaptations to them, appear to be the most likely major driving force in herbivorous insect evolution [81]. Plant molecules can be toxic to insects, and consequently, GSTs and detoxifying enzymes are essential for insect survival. GSTs detoxify a broad range of plant molecules, generally with an overlap of GSTs for the same substrate [82,83,84]. Signatures of a positive selection of Delta GSTs suggest that they may have evolved under positive selection in the herbivorous [85] lineage after the transition of insects to herbivory > 350 Ma [86]. This adaptation phenomenon can be rapid; indeed, anthropological pressure toward insects for insecticide resistance has been suggested to promote Musca domestica gst gene amplification [87,88]. The main classes of GST diversification appear to be the Epsilon and Delta classes in various insects, such as Anopheles gambiae, Drosophilia melanogaster, or Tribolium castaneum [79,89]. In this context, it is not surprising to observe numerous insect adaptations toward insecticides [90,91] due to the Delta and Epsilon GSTs in the role of A. gambiae GSTE1 and GSTE2 in the DDT resistance [92]. The chemical resistance promoted by GSTs involves various chemicals, such as pyrethroids or neonicotinoids and 2,2-dichlorovinyl dimethylphosphate for Diaphorina citri [93] and Rynochophorus phoenicis, respectively [94]. In contrast, Apis mellifera, known to be highly sensitive to insecticides, presents only one Delta GST (including two isoforms) and no Epsilon GST. It is not excluded that some GSTs resulting from functional differentiation appear with different functions not related at all to xenobiotic metabolism, such as GSTE14 in D. melanogaster, which is involved in ecdysone biosynthesis [13,95,96]. Additionally, GSTs formed during the diversification process can also be specific in metabolizing some molecules without a functional overlap from other GSTs within the same insect species. For example, the deletion of epsilon and omega GSTs in the Asian gypsy moth, Lymantria dispar, affected its adaptability to salicin and rutin produced by its host, the poplar tree [97].
Table 2.
Number of identified canonical GSTs in different insect species.
As shown in vertebrate chemosensory organs, GSTs are also expressed in insect chemosensory organs (Table 3). This expression is advantageous to protect these sensitive organs where neurons are directly exposed to xenobiotics. GSTs were shown to be expressed in the antennae of various orders of insect species, such as the dipteran D. melanogaster [17,134], various lepidopteran species [107,130,135,136,137,138,139] such as Manduca sexta [140] or Spodoptera littoralis [141], and in the Coleoptera antennae of Agrilus planipennis [142] or Dendroctonus valens [143]. Table 3 shows the diversity of insect species expressing GSTs within their sensory organs. Although a limited number of studies have analyzed GST expression, GSTs appear to be ubiquitously expressed in antennae. To support this hypothesis, GST expression in two particular insect species can be highlighted. The only Delta GST found in A. melifera is expressed in its antennae [144]. Ticks have a unique chemosensory organ presumed to function similarly to insect antennae, the fore-tarsal Haller’s organ. GSTs were found to be expressed in this organ of the dog tick, Dermacentor variabilis [145]. As in mammals, GSTs were proposed to protect the chemosensory organs so they could participate in odorant clearance and consequently signal termination. Antennal GSTs were shown to be active toward the model substrate CDNB (1-chloro-2,4-dinitrobenzene); for example, most antennal Drosophila GSTs [146,147]. The ability to conjugate CDNB was also observed for the antennal-specific GST identified in Bombyx mori [137]. The selective pressure to conserve efficient odorant clearance is crucial for flying insects, which need to reinitiate odorant perception as quickly as possible to follow the odorant volute. The detection of odorant food sources can be diversified, probably involving different GSTs. However, pheromone detection can also involve more specialized GSTs, as shown for an antenna-specific Delta GST found in Manducta sexta. This GST was shown to metabolize trans-2-hexenal, a plant-derived green leaf aldehyde known to stimulate the olfactory system of M. sexta. This GST was proposed to be involved in the signal termination of a complex mixture of aldehyde molecules forming the sex pheromone bouquet [140]. A delta GST found in the antennae of Grapholita molesta shows high activity toward a sex pheromone component, (Z)-8-dodecenyl alcohol [148]. The role of GST in sex pheromone detection is also supported by the differential expression of GST depending on the insect sex. For example, antennal-specific genes of a GST belonging to the Delta class were significantly more highly expressed in male Helicoverpa armigera antennae compared with females [139]. Despite all the different studies, to the best of the authors’ knowledge, the cellular localization of GSTs within the olfactory sensilla is not known to date. Moreover, expression within the sensory lymph, where the olfactory neurons are located, has not been validated experimentally. The same question about localization exists for insect gustatory sensilla. GSTs have been identified in diverse taste organs, such as the labellum, in insects belonging to the dipteran and ledidopteran orders (Table 3); however, the cell types and localization within the lymph of gustatory sensilla are not shown. Food containing bitter molecules such as glucosinolates and isothiocyanates led to GST overexpression in aphids in a general manner [149]. Additionally, other results have shown the same regulation in chemosensory organs. Isothiocyanates were shown to increase the expression of GST Delta in Drosophila labellum [5]. This modulation can be hypothesized to affect food habits. The results showed that the loss of bitter taste receptors observed in D. suzukii in comparison to D. melanogaster was proposed to contribute to the evolutionary shift in oviposition preference between the two species [150], knowing that ovipositor organs are taste-sensitive. In the same study, the taste difference between these two Drosophila species showed an associated differential expression of xenobiotic metabolism enzymes, including delta and epsilon GSTs. Again, this observation supports a direct link between these enzymes and the taste biochemistry in insects.
Table 3.
Identification of GSTs in the chemosensory organs of various insect species.
4. Discussion
GST appears to be a main actor in the mammalian and insect detoxifying systems. In insects, Delta and Epsilon GST diversification were shown to be associated with chemical resistance toward numerous molecules found naturally in plants, such as isothiocyanates, or resulting from human activity, such as pesticides. Interestingly, GSTs were found to be involved in the metabolism of similar molecules both in mammals and insects after the separation of these lineages [156]. Isothiocyanate molecules found in terrestrial plants were shown to modulate GST expression in mammals and insects [5,54,157]. These observations support the idea that GST gene diversification offers advantageous opportunities during evolution to build functional chemosensory systems to face molecular diversification in plant molecules. It may also have offered some species better adaptability to their environment, although this advantage is less important for species with a more specific ecological niche, such as Apis melifera. Interestingly, both olfactory and gustatory organs appeared independently in mammals and insects during their evolution and present an analogous organization as the chemosensory neurons wearing membrane olfactory receptors. In both cases, the receptors evolved from a different membrane protein ancestor, whereas the GSTs found in these two systems evolved from the same ancestral GST (homologous). In the insect lineage, GSTs of the Delta class have been found in almost all studies analyzing the contents of the antennae, indicating the importance of this class in the olfactory organs.
Crustaceans and insects share Delta and Epsilon GSTs, indicating their appearance before the separation of the lineages and consequently before the evolutionary elaboration of their olfactory systems, which did not share the same protein actors [158]. This observation supports that these specific GST classes did not appear to support the physiology of the chemosensory organs, and it is more likely that different members of existing classes of GSTs were randomly used in the chemosensory organs, highlighting the versatility of GSTs. One of the main biological functions of chemosensory GSTs is the protection of the chemosensory organs where neurons are exposed. The lifespan of olfactory neurons in rodents is approximately 40 days [159], although xenobiotic metabolism enzymes already contribute to extending this life; thus, neurons must continuously be replenished. In this context, it is not surprising to find numerous GST members as key players in xenobiotic metabolism in both mammalian and insect chemosensory systems (Table 3). In addition to chemoprotection, GSTs modulate their perception when these xenobiotic molecules are perceived by decreasing the xenobiotic concentration. This metabolic activity can also be involved in the termination of the signal if the metabolites are no longer perceived, or in the perception of a new signal if the metabolite is perceived by other chemoreceptors (Figure 1). To date, GSTs have only been shown to be involved in signal termination in mammals (rabbits) and insects. Due to the increased steric hindrance after glutathione conjugation, it is more likely that the chance of molecules binding receptors will decrease. However, it is not excluded that, after GST catalysis, those molecules can be perceived differently. Other GST activities, such as glutathione conjugation, are more susceptible to generating molecules that can be perceived, including isomerization activity. Indeed, chemosensory receptors are known to be stereospecific. Additionally, the ligandin capacity of GSTs has been proposed to help the diffusion of hydrophobic molecules as odorants in hydrophilic olfactory mucus surrounding the olfactory receptors [4]. A similar role in the sensillary lymph surrounding the insect olfactory receptors is not excluded if GSTs are expressed within the lymph. GST localization in sensilla lymphs is only supported by the observation of a signal peptide in some insect GSTs that are expressed in sensilla [141]. Interestingly, an increase in diffusion can also be proposed for all chemosensory molecules as a consequence of glutathione conjugation, as suggested for bitter molecules [55]. After glutathione conjugation, molecules are more hydrophilic. Thus, if it does not impact the ability of the molecules to bind to their receptors or their affinity ranges, they are most likely rapidly perceived.
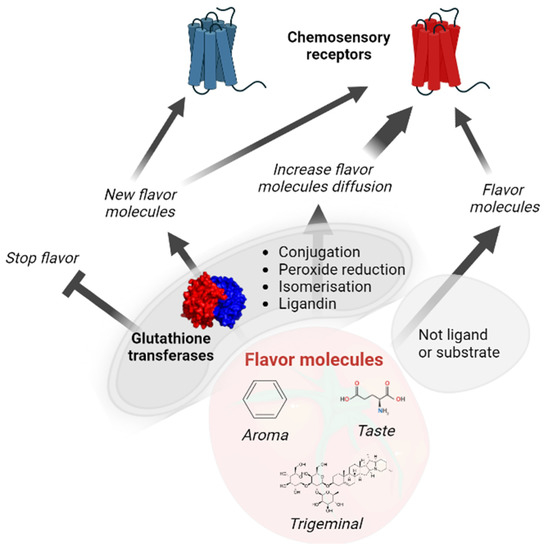
Figure 1.
Role of glutathione transferases in chemosensory modulation.
To conclude, numerous questions about GSTs and their function in the chemosensory system remain open. For instance, their roles in trigeminal perception have never been studied to date in any organism. Numerous GST members are produced in bacteria, opening questions regarding their role in both mammalian and insect chemoperception in view of recent advances showing the potential roles of bacterial enzymes in human chemosensory perception [160,161] and insect chemoperception [162]. GST functions in chemosensory perception seem to be conserved in non-evolutionarily related chemosensory systems. Thus, the knowledge obtained from experimental data in one lineage should be tested in other lineages to enhance the understanding of their functions.
Author Contributions
M.S. and F.N. wrote the manuscript, V.B., S.F., M.M., P.S., A.N., F.L., M.B., F.C. and J.-M.H. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The authors received funding for this work: F.N., Agence Nationale de la Recherche grant number ANR-16-CE21-0004-01 (https://anr.fr). J.-M.H., ANR-18-CE92-0018-01, M.S., ANR-22-CE21-0001, F.C. ANR-20-CE21-0002.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Valen, L. A new evolutionary law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Booth, J.; Boyland, E.; Sims, P. An enzyme from rat liver catalysing conjugations with glutathione. Biochem. J. 1961, 79, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Mannervik, B. Five decades with glutathione and the GSTome. J. Biol. Chem. 2012, 287, 6072–6083. [Google Scholar] [CrossRef]
- Heydel, J.M.; Menetrier, F.; Belloir, C.; Canon, F.; Faure, P.; Lirussi, F.; Chavanne, E.; Saliou, J.M.; Artur, Y.; Canivenc-Lavier, M.C.; et al. Characterization of rat glutathione transferases in olfactory epithelium and mucus. PLoS ONE 2019, 14, e0220259. [Google Scholar] [CrossRef]
- Gonzalez, D.; Fraichard, S.; Grassein, P.; Delarue, P.; Senet, P.; Nicolai, A.; Chavanne, E.; Mucher, E.; Artur, Y.; Ferveur, J.F.; et al. Characterization of a Drosophila glutathione transferase involved in isothiocyanate detoxification. Insect Biochem. Mol. Biol. 2018, 95, 33–43. [Google Scholar] [CrossRef]
- Hayes, J.D.; Strange, R.C. Glutathione S-transferase polymorphisms and their biological consequences. Pharmacology 2000, 61, 154–166. [Google Scholar] [CrossRef]
- Morel, F.; Aninat, C. The glutathione transferase kappa family. Drug Metab. Rev. 2011, 43, 281–291. [Google Scholar] [CrossRef]
- Ladner, J.E.; Parsons, J.F.; Rife, C.L.; Gilliland, G.L.; Armstrong, R.N. Parallel evolutionary pathways for glutathione transferases: Structure and mechanism of the mitochondrial class kappa enzyme rGSTK1-1. Biochemistry 2004, 43, 352–361. [Google Scholar] [CrossRef]
- Roncalli, V.; Cieslak, M.C.; Passamaneck, Y.; Christie, A.E.; Lenz, P.H. Glutathione S-Transferase (GST) Gene Diversity in the Crustacean Calanus finmarchicus--Contributors to Cellular Detoxification. PLoS ONE 2015, 10, e0123322. [Google Scholar] [CrossRef]
- Hurst, R.; Bao, Y.; Jemth, P.; Mannervik, B.; Williamson, G. Phospholipid hydroperoxide glutathione peroxidase activity of human glutathione transferases. Biochem. J. 1998, 332 Pt 1, 97–100. [Google Scholar] [CrossRef]
- Clark, A.G.; Shamaan, N.A. Evidence that DDT-dehydrochlorinase from the house fly is a glutathione S-transferase. Pestic. Biochem. Physiol. 1984, 22, 249–261. [Google Scholar] [CrossRef]
- Johansson, A.S.; Mannervik, B. Human glutathione transferase A3-3, a highly efficient catalyst of double-bond isomerization in the biosynthetic pathway of steroid hormones. J. Biol. Chem. 2001, 276, 33061–33065. [Google Scholar] [CrossRef]
- Škerlová, J.; Lindström, H.; Gonis, E.; Sjödin, B.; Neiers, F.; Stenmark, P.; Mannervik, B. Structure and steroid isomerase activity of Drosophila glutathione transferase E14 essential for ecdysteroid biosynthesis. FEBS Lett. 2019, 594, 1187–1195. [Google Scholar] [CrossRef]
- Kamisaka, K.; Gatmaitan, Z.; Moore, C.L.; Arias, I.M. Ligandin Reverses Bilirubin Inhibition of Liver Mitochondrial Respiration In Vitro. Pediatr. Res. 1975, 9, 903–905. [Google Scholar]
- Mannervik, B.; Board, P.G.; Hayes, J.D.; Listowsky, I.; Pearson, W.R. Nomenclature for mammalian soluble glutathione transferases. Methods Enzymol. 2005, 401, 1–8. [Google Scholar] [CrossRef]
- Sherratt, P.J.; Hayes, J.D. 9 Glutathione S-Transferases; Costas, I., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Gonis, E.; Fraichard, S.; Chertemps, T.; Hecker, A.; Schwartz, M.; Canon, F.; Neiers, F. Expression Patterns of Drosophila Melanogaster Glutathione Transferases. Insects 2022, 13, 612. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, Z.; Lin, X.; Feng, Q.; Zheng, S. Structure and expression of glutathione S-transferase genes from the midgut of the Common cutworm, Spodoptera litura (Noctuidae) and their response to xenobiotic compounds and bacteria. J. Insect Physiol. 2011, 57, 1033–1044. [Google Scholar] [CrossRef]
- Knight, T.R.; Choudhuri, S.; Klaassen, C.D. Constitutive mRNA expression of various glutathione S-transferase isoforms in different tissues of mice. Toxicol. Sci. 2007, 100, 513–524. [Google Scholar] [CrossRef]
- Sullivan, S.L. Mammalian chemosensory receptors. Neuroreport 2002, 13, A9–A17. [Google Scholar] [CrossRef]
- Filiou, R.P.; Lepore, F.; Bryant, B.; Lundstrom, J.N.; Frasnelli, J. Perception of trigeminal mixtures. Chem. Senses 2015, 40, 61–69. [Google Scholar] [CrossRef]
- Su, C.Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, D.A.; Zuker, C.S.; Ryba, N.J. Common sense about taste: From mammals to insects. Cell 2009, 139, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Front. Pharmacol. 2020, 11, 587664. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, L.; Lametsch, R. Current progress in kokumi-active peptides, evaluation and preparation methods: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 1230–1241. [Google Scholar] [CrossRef]
- Toelstede, S.; Hofmann, T. Kokumi-active glutamyl peptides in cheeses and their biogeneration by Penicillium roquefortii. J. Agric. Food Chem. 2009, 57, 3738–3748. [Google Scholar] [CrossRef]
- Maruyama, Y.; Yasuda, R.; Kuroda, M.; Eto, Y. Kokumi substances, enhancers of basic tastes, induce responses in calcium-sensing receptor expressing taste cells. PLoS ONE 2012, 7, e34489. [Google Scholar] [CrossRef]
- Canon, F.; Belloir, C.; Bourillot, E.; Brignot, H.; Briand, L.; Feron, G.; Lesniewska, E.; Nivet, C.; Septier, C.; Schwartz, M.; et al. Perspectives on Astringency Sensation: An Alternative Hypothesis on the Molecular Origin of Astringency. J. Agric. Food Chem. 2021, 69, 3822–3826. [Google Scholar] [CrossRef]
- Boichot, V.; Muradova, M.; Nivet, C.; Proskura, A.; Heydel, J.M.; Canivenc-Lavier, M.C.; Canon, F.; Neiers, F.; Schwartz, M. The role of perirecetor events in flavor perception. Front. Food Sci. Technol. 2022, 2, 989291. [Google Scholar] [CrossRef]
- Sarkar, M.A. Drug metabolism in the nasal mucosa. Pharm. Res. 1992, 9, 1–9. [Google Scholar] [CrossRef]
- Heydel, J.M.; Faure, P.; Neiers, F. Nasal odorant metabolism: Enzymes, activity and function in olfaction. Drug Metab. Rev. 2019, 51, 224–245. [Google Scholar] [CrossRef]
- Mallery, S.R.; Budendorf, D.E.; Larsen, M.P.; Pei, P.; Tong, M.; Holpuch, A.S.; Larsen, P.E.; Stoner, G.D.; Fields, H.W.; Chan, K.K.; et al. Effects of human oral mucosal tissue, saliva, and oral microflora on intraoral metabolism and bioactivation of black raspberry anthocyanins. Cancer Prev. Res. 2011, 4, 1209–1221. [Google Scholar] [CrossRef]
- Schwartz, M.; Neiers, F.; Charles, J.P.; Heydel, J.M.; Munoz-Gonzalez, C.; Feron, G.; Canon, F. Oral enzymatic detoxification system: Insights obtained from proteome analysis to understand its potential impact on aroma metabolization. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5516–5547. [Google Scholar] [CrossRef]
- Canon, F.; Neiers, F.; Guichard, E. Saliva and Flavor Perception: Perspectives. J. Agric. Food Chem. 2018, 66, 7873–7879. [Google Scholar] [CrossRef]
- Munoz-Gonzalez, C.; Brule, M.; Martin, C.; Feron, G.; Canon, F. Molecular mechanisms of aroma persistence: From noncovalent interactions between aroma compounds and the oral mucosa to metabolization of aroma compounds by saliva and oral cells. Food Chem. 2022, 373, 131467. [Google Scholar] [CrossRef]
- Robert-Hazotte, A.; Faure, P.; Menetrier, F.; Folia, M.; Schwartz, M.; Le Quere, J.L.; Neiers, F.; Thomas-Danguin, T.; Heydel, J.M. Nasal Odorant Competitive Metabolism Is Involved in the Human Olfactory Process. J. Agric. Food Chem. 2022, 70, 8385–8394. [Google Scholar] [CrossRef]
- Ployon, S.; Brule, M.; Andriot, I.; Morzel, M.; Canon, F. Understanding retention and metabolization of aroma compounds using an in vitro model of oral mucosa. Food Chem. 2020, 318, 126468. [Google Scholar] [CrossRef]
- Mannervik, B.; Alin, P.; Guthenberg, C.; Jensson, H.; Tahir, M.K.; Warholm, M.; Jornvall, H. Identification of three classes of cytosolic glutathione transferase common to several mammalian species: Correlation between structural data and enzymatic properties. Proc. Natl. Acad. Sci. USA 1985, 82, 7202–7206. [Google Scholar] [CrossRef]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef]
- Ben-Arie, N.; Khen, M.; Lancet, D. Glutathione S-transferases in rat olfactory epithelium: Purification, molecular properties and odorant biotransformation. Biochem. J. 1993, 292 Pt 2, 379–384. [Google Scholar] [CrossRef]
- Krishna, N.S.; Getchell, T.V.; Getchell, M.L. Differential expression of alpha, mu, and pi classes of glutathione S-transferases in chemosensory mucosae of rats during development. Cell Tissue Res. 1994, 275, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Menetrier, F.; Heydel, J.M.; Chavanne, E.; Faure, P.; Labrousse, M.; Lirussi, F.; Canon, F.; Mannervik, B.; Briand, L.; et al. Interactions Between Odorants and Glutathione Transferases in the Human Olfactory Cleft. Chem. Senses 2020, 45, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, K.; Tandler, B. Dividing type II cell in rabbit taste bud. Anat Rec. 1986, 214, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.M.; Cox, M.E.; Solomons, N. Zinc and the special senses. Ann. Intern. Med. 1983, 99, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Kudo, H.; Doi, Y.; Nishino, T.; Nara, S.; Hamasaki, K.; Fujimoto, S. Dietary zinc deficiency decreases glutathione S-transferase expression in the rat olfactory epithelium. J. Nutr. 2000, 130, 38–44. [Google Scholar] [CrossRef][Green Version]
- Schaal, B.; Coureaud, G.; Langlois, D.; Ginies, C.; Semon, E.; Perrier, G. Chemical and behavioural characterization of the rabbit mammary pheromone. Nature 2003, 424, 68–72. [Google Scholar] [CrossRef]
- Legendre, A.; Faure, P.; Tiesset, H.; Potin, C.; Jakob, I.; Sicard, G.; Schaal, B.; Artur, Y.; Coureaud, G.; Heydel, J.M. When the nose must remain responsive: Glutathione conjugation of the mammary pheromone in the newborn rabbit. Chem. Senses 2014, 39, 425–437. [Google Scholar] [CrossRef]
- Robert-Hazotte, A.; Faure, P.; Neiers, F.; Potin, C.; Artur, Y.; Coureaud, G.; Heydel, J.M. Nasal mucus glutathione transferase activity and impact on olfactory perception and neonatal behavior. Sci. Rep. 2019, 9, 3104. [Google Scholar] [CrossRef]
- Hanser, H.I.; Faure, P.; Robert-Hazotte, A.; Artur, Y.; Duchamp-Viret, P.; Coureaud, G.; Heydel, J.M. Odorant-odorant metabolic interaction, a novel actor in olfactory perception and behavioral responsiveness. Sci. Rep. 2017, 7, 10219. [Google Scholar] [CrossRef]
- Nishino, T.; Kudo, H.; Doi, Y.; Maeda, M.; Hamasaki, K.; Morita, M.; Fujimoto, S. Immunocytochemistry of glutathione S-transferase in taste bud cells of rat circumvallate and foliate papillae. Chem. Senses 2001, 26, 179–188. [Google Scholar] [CrossRef][Green Version]
- Fabrini, R.; Bocedi, A.; Camerini, S.; Fusetti, M.; Ottaviani, F.; Passali, F.M.; Topazio, D.; Iavarone, F.; Francia, I.; Castagnola, M.; et al. Inactivation of human salivary glutathione transferase P1-1 by hypothiocyanite: A post-translational control system in search of a role. PLoS ONE 2014, 9, e112797. [Google Scholar] [CrossRef]
- Derindag, G.; Akgul, H.M.; Kiziltunc, A.; Ozkan, H.I.; Kiziltunc Ozmen, H.; Akgul, N. Evaluation of saliva glutathione, glutathione peroxidase, and malondialdehyde levels in head-neck radiotherapy patients. Turk J. Med. Sci. 2021, 51, 644–649. [Google Scholar] [CrossRef]
- Sreerama, L.; Hedge, M.W.; Sladek, N.E. Identification of a class 3 aldehyde dehydrogenase in human saliva and increased levels of this enzyme, glutathione S-transferases, and DT-diaphorase in the saliva of subjects who continually ingest large quantities of coffee or broccoli. Clin. Cancer Res. 1995, 1, 1153–1163. [Google Scholar]
- Schwartz, M.; Brignot, H.; Feron, G.; Hummel, T.; Zhu, Y.; von Koskull, D.; Heydel, J.M.; Lirussi, F.; Canon, F.; Neiers, F. Role of human salivary enzymes in bitter taste perception. Food Chem. 2022, 386, 132798. [Google Scholar] [CrossRef]
- Franco-Martinez, L.; Gelemanovic, A.; Horvatic, A.; Contreras-Aguilar, M.D.; Dabrowski, R.; Mrljak, V.; Ceron, J.J.; Martinez-Subiela, S.; Tvarijonaviciute, A. Changes in Serum and Salivary Proteins in Canine Mammary Tumors. Animals 2020, 10, 741. [Google Scholar] [CrossRef]
- de Sousa-Pereira, P.; Cova, M.; Abrantes, J.; Ferreira, R.; Trindade, F.; Barros, A.; Gomes, P.; Colaco, B.; Amado, F.; Esteves, P.J.; et al. Cross-species comparison of mammalian saliva using an LC-MALDI based proteomic approach. Proteomics 2015, 15, 1598–1607. [Google Scholar] [CrossRef]
- Debat, H.; Eloit, C.; Blon, F.; Sarazin, B.; Henry, C.; Huet, J.C.; Trotier, D.; Pernollet, J.C. Identification of human olfactory cleft mucus proteins using proteomic analysis. J. Proteome Res. 2007, 6, 1985–1996. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Wang, H.; Jaen, C.; Haneoka, M.; Saito, N.; Nakamura, J.; Adappa, N.D.; Cohen, N.A.; Dalton, P. The human olfactory cleft mucus proteome and its age-related changes. Sci. Rep. 2018, 8, 17170. [Google Scholar] [CrossRef]
- Pappa, E.; Vastardis, H.; Mermelekas, G.; Gerasimidi-Vazeou, A.; Zoidakis, J.; Vougas, K. Saliva Proteomics Analysis Offers Insights on Type 1 Diabetes Pathology in a Pediatric Population. Front. Physiol. 2018, 9, 444. [Google Scholar] [CrossRef]
- Grassl, N.; Kulak, N.A.; Pichler, G.; Geyer, P.E.; Jung, J.; Schubert, S.; Sinitcyn, P.; Cox, J.; Mann, M. Ultra-deep and quantitative saliva proteome reveals dynamics of the oral microbiome. Genome Med. 2016, 8, 44. [Google Scholar] [CrossRef]
- Kuhlmann, K.; Tschapek, A.; Wiese, H.; Eisenacher, M.; Meyer, H.E.; Hatt, H.H.; Oeljeklaus, S.; Warscheid, B. The membrane proteome of sensory cilia to the depth of olfactory receptors. Mol. Cell Proteom. 2014, 13, 1828–1843. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Zweyer, M.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Dataset on the comparative proteomic profiling of mouse saliva and serum from wild type versus the dystrophic mdx-4cv mouse model of dystrophinopathy. Data Brief 2018, 21, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Mayer, U.; Ungerer, N.; Klimmeck, D.; Warnken, U.; Schnolzer, M.; Frings, S.; Mohrlen, F. Proteomic analysis of a membrane preparation from rat olfactory sensory cilia. Chem. Senses 2008, 33, 145–162. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Bass, C.; Feyereisen, R.; Vontas, J. The Role of Cytochrome P450s in Insect Toxicology and Resistance. Annu. Rev. Entomol. 2022, 67, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Furlong, M.J. Behavior as a mechanism of insecticide resistance: Evaluation of the evidence. Curr. Opin. Insect Sci. 2017, 21, 19–25. [Google Scholar] [CrossRef]
- Fischler, W.; Kong, P.; Marella, S.; Scott, K. The detection of carbonation by the Drosophila gustatory system. Nature 2007, 448, 1054–1057. [Google Scholar] [CrossRef]
- Scott, K. Out of thin air: Sensory detection of oxygen and carbon dioxide. Neuron 2011, 69, 194–202. [Google Scholar] [CrossRef]
- Cameron, P.; Hiroi, M.; Ngai, J.; Scott, K. The molecular basis for water taste in Drosophila. Nature 2010, 465, 91–95. [Google Scholar] [CrossRef]
- Falk, R.; Bleiser-Avivi, N.; Atidia, J. Labellar taste organs of Drosophila melanogaster. J. Morphol. 1976, 150, 327–341. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Structure and function of insect olfactory sensilla. Ciba Found Symp. 1996, 200, 158–174; discussion 157–174. [Google Scholar] [CrossRef]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Hildebrand, J.G. Olfactory systems: Common design, uncommon origins? Curr. Opin. Neurobiol. 1999, 9, 634–639. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Carlson, J.R. Candidate taste receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Isono, K.; Morita, H. Molecular and cellular designs of insect taste receptor system. Front. Cell Neurosci. 2010, 4, 20. [Google Scholar] [CrossRef]
- Sanchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Shi, H.; Pei, L.; Gu, S.; Zhu, S.; Wang, Y.; Zhang, Y.; Li, B. Glutathione S-transferase (GST) genes in the red flour beetle, Tribolium castaneum, and comparative analysis with five additional insects. Genomics 2012, 100, 327–335. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by Gene Duplication; Allen & Unwin; Springer: London, UK; New York, NY, USA, 1970; p. xv. 160p. [Google Scholar]
- Mitter, C.; Farrell, B.; Wiegmann, B. The phylogenetic study of adaptive zones: Has phytophagy promoted insect diversification? Am. Nat. 1988, 132, 107–128. [Google Scholar] [CrossRef]
- Saruta, F.; Yamada, N.; Yamamoto, K. Functional Analysis of an Epsilon-Class Glutathione S-Transferase From Nilaparvata lugens (Hemiptera: Delphacidae). J. Insect Sci. 2019, 19, 14. [Google Scholar] [CrossRef]
- Yang, J.; Kong, X.D.; Zhu-Salzman, K.; Qin, Q.M.; Cai, Q.N. The Key Glutathione S-Transferase Family Genes Involved in the Detoxification of Rice Gramine in Brown Planthopper Nilaparvata lugens. Insects 2021, 12, 1055. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Xu, Z.; Zou, H.; Liu, J.; Chen, S.; Feng, Q.; Zheng, S. Glutathione S-transferase SlGSTE1 in Spodoptera litura may be associated with feeding adaptation of host plants. Insect Biochem. Mol. Biol. 2016, 70, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Gloss, A.D.; Vassao, D.G.; Hailey, A.L.; Nelson Dittrich, A.C.; Schramm, K.; Reichelt, M.; Rast, T.J.; Weichsel, A.; Cravens, M.G.; Gershenzon, J.; et al. Evolution in an ancient detoxification pathway is coupled with a transition to herbivory in the drosophilidae. Mol. Biol. Evol. 2014, 31, 2441–2456. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.C. Early history of Arthropod and vascular plant associations. Annu. Rev. Earth Planet. Sci. 1998, 26, 329–377. [Google Scholar] [CrossRef]
- Syvanen, M.; Zhou, Z.; Wharton, J.; Goldsbury, C.; Clark, A. Heterogeneity of the glutathione transferase genes encoding enzymes responsible for insecticide degradation in the housefly. J. Mol. Evol. 1996, 43, 236–240. [Google Scholar] [CrossRef]
- Wang, J.Y.; McCommas, S.; Syvanen, M. Molecular cloning of a glutathione S-transferase overproduced in an insecticide-resistant strain of the housefly (Musca domestica). Mol. Gen. Genet. MGG 1991, 227, 260–266. [Google Scholar] [CrossRef]
- Paton, M.G.; Karunaratne, S.H.; Giakoumaki, E.; Roberts, N.; Hemingway, J. Quantitative analysis of gene amplification in insecticide-resistant Culex mosquitoes. Biochem. J. 2000, 346 Pt 1, 17–24. [Google Scholar] [CrossRef]
- Yamamoto, K.; Higashiura, A.; Hirowatari, A.; Yamada, N.; Tsubota, T.; Sezutsu, H.; Nakagawa, A. Characterisation of a diazinon-metabolising glutathione S-transferase in the silkworm Bombyx mori by X-ray crystallography and genome editing analysis. Sci. Rep. 2018, 8, 16835. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ichinose, H.; Aso, Y.; Banno, Y.; Kimura, M.; Nakashima, T. Molecular characterization of an insecticide-induced novel glutathione transferase in silkworm. Biochim. Biophys. Acta 2011, 1810, 420–426. [Google Scholar] [CrossRef]
- Ortelli, F.; Rossiter, L.C.; Vontas, J.; Ranson, H.; Hemingway, J. Heterologous expression of four glutathione transferase genes genetically linked to a major insecticide-resistance locus from the malaria vector Anopheles gambiae. Biochem. J. 2003, 373, 957–963. [Google Scholar] [CrossRef]
- Yu, X.; Killiny, N. RNA interference of two glutathione S-transferase genes, Diaphorina citri DcGSTe2 and DcGSTd1, increases the susceptibility of Asian citrus psyllid (Hemiptera: Liviidae) to the pesticides fenpropathrin and thiamethoxam. Pest Manag. Sci. 2018, 74, 638–647. [Google Scholar] [CrossRef]
- Bamidele, O.S.; Ajele, J.O.; Olajuyigbe, F.M.; Gaber, Y. An evaluation of glutathione transferase associated with Dichlorvos degradation in African palm weevil (Rynchophorus phoenicis) larva. Cogent Biol. 2017, 3, 1286764. [Google Scholar] [CrossRef]
- Chanut-Delalande, H.; Hashimoto, Y.; Pelissier-Monier, A.; Spokony, R.; Dib, A.; Kondo, T.; Bohere, J.; Niimi, K.; Latapie, Y.; Inagaki, S.; et al. Pri peptides are mediators of ecdysone for the temporal control of development. Nat. Cell Biol. 2014, 16, 1035–1044. [Google Scholar] [CrossRef]
- Enya, S.; Ameku, T.; Igarashi, F.; Iga, M.; Kataoka, H.; Shinoda, T.; Niwa, R. A Halloween gene noppera-bo encodes a glutathione S-transferase essential for ecdysteroid biosynthesis via regulating the behaviour of cholesterol in Drosophila. Sci. Rep. 2014, 4, 6586. [Google Scholar] [CrossRef]
- Ma, J.; Sun, L.; Zhao, H.; Wang, Z.; Zou, L.; Cao, C. Functional identification and characterization of GST genes in the Asian gypsy moth in response to poplar secondary metabolites. Pestic. Biochem. Physiol. 2021, 176, 104860. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Wang, Z.Q.; Zhu, J.Y.; Liu, N.Y. Identification and characterization of detoxification genes in two cerambycid beetles, Rhaphuma horsfieldi and Xylotrechus quadripes (Coleoptera: Cerambycidae: Clytini). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 243–244, 110431. [Google Scholar] [CrossRef]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle-plant interface. Genome Biol. 2016, 17, 227. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Wu, Z.; Bin, S.; He, H.; Wang, Z.; Li, M.; Lin, J. Differential Expression Analysis of Chemoreception Genes in the Striped Flea Beetle Phyllotreta striolata Using a Transcriptomic Approach. PLoS ONE 2016, 11, e0153067. [Google Scholar] [CrossRef]
- Dai, L.; Ma, J.; Ma, M.; Zhang, H.; Shi, Q.; Zhang, R.; Chen, H. Characterisation of GST genes from the Chinese white pine beetle Dendroctonus armandi (Curculionidae: Scolytinae) and their response to host chemical defence. Pest Manag. Sci. 2016, 72, 816–827. [Google Scholar] [CrossRef]
- Keeling, C.I.; Yuen, M.M.; Liao, N.Y.; Docking, T.R.; Chan, S.K.; Taylor, G.A.; Palmquist, D.L.; Jackman, S.D.; Nguyen, A.; Li, M.; et al. Draft genome of the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major forest pest. Genome Biol. 2013, 14, R27. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Ye, K.; Tu, X.F.; Lu, Y.J.; Thakur, K.; Jiang, L.; Wei, Z.J. Identification and expression profiles of twenty-six glutathione S-transferase genes from rice weevil, Sitophilus oryzae (Coleoptera: Curculionidae). Int. J. Biol. Macromol. 2018, 120, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; McKenna, D.; Scully, E.; Cook, S.C.; Dainat, B.; Egekwu, N.; Grubbs, N.; Lopez, D.; Lorenzen, M.D.; Reyna, S.M.; et al. Genome of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae), a worldwide parasite of social bee colonies, provides insights into detoxification and herbivory. Gigascience 2018, 7, giy138. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Markov, G.V.; Baskaran, P.; Herrmann, M.; Sommer, R.J.; Rodelsperger, C. Draft Genome of the Scarab Beetle Oryctes borbonicus on La Reunion Island. Genome Biol. Evol. 2016, 8, 2093–2105. [Google Scholar] [CrossRef]
- Liu, S.; Rao, X.J.; Li, M.Y.; Feng, M.F.; He, M.Z.; Li, S.G. Glutathione s-transferase genes in the rice leaffolder, cnaphalocrocis medinalis (Lepidoptera: Pyralidae): Identification and expression profiles. Arch Insect Biochem. Physiol. 2015, 90, 1–13. [Google Scholar] [CrossRef]
- Nair, P.M.; Choi, J. Identification, characterization and expression profiles of Chironomus riparius glutathione S-transferase (GST) genes in response to cadmium and silver nanoparticles exposure. Aquat. Toxicol. 2011, 101, 550–560. [Google Scholar] [CrossRef]
- Strode, C.; Wondji, C.S.; David, J.P.; Hawkes, N.J.; Lumjuan, N.; Nelson, D.R.; Drane, D.R.; Karunaratne, S.H.; Hemingway, J.; Black, W.C.t.; et al. Genomic analysis of detoxification genes in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 113–123. [Google Scholar] [CrossRef]
- Yan, L.; Yang, P.; Jiang, F.; Cui, N.; Ma, E.; Qiao, C.; Cui, F. Transcriptomic and phylogenetic analysis of Culex pipiens quinquefasciatus for three detoxification gene families. BMC Genom. 2012, 13, 609. [Google Scholar] [CrossRef]
- Niranjan Reddy, B.P.; Prasad, G.B.; Raghavendra, K. In silico characterization and comparative genomic analysis of the Culex quinquefasciatus glutathione S-transferase (GST) supergene family. Parasitol. Res. 2011, 109, 1165–1177. [Google Scholar] [CrossRef]
- Hu, F.; Dou, W.; Wang, J.J.; Jia, F.X.; Wang, J.J. Multiple glutathione S-transferase genes: Identification and expression in oriental fruit fly, Bactrocera dorsalis. Pest Manag. Sci. 2014, 70, 295–303. [Google Scholar] [CrossRef]
- Papanicolaou, A.; Schetelig, M.F.; Arensburger, P.; Atkinson, P.W.; Benoit, J.B.; Bourtzis, K.; Castanera, P.; Cavanaugh, J.P.; Chao, H.; Childers, C.; et al. The whole genome sequence of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), reveals insights into the biology and adaptive evolution of a highly invasive pest species. Genome Biol. 2016, 17, 192. [Google Scholar] [CrossRef]
- Aidlin Harari, O.; Santos-Garcia, D.; Musseri, M.; Moshitzky, P.; Patel, M.; Visendi, P.; Seal, S.; Sertchook, R.; Malka, O.; Morin, S. Molecular Evolution of the Glutathione S-Transferase Family in the Bemisia tabaci Species Complex. Genome Biol. Evol. 2020, 12, 3857–3872. [Google Scholar] [CrossRef]
- Bailey, E.; Field, L.; Rawlings, C.; King, R.; Mohareb, F.; Pak, K.H.; Hughes, D.; Williamson, M.; Ganko, E.; Buer, B.; et al. A scaffold-level genome assembly of a minute pirate bug, Orius laevigatus (Hemiptera: Anthocoridae), and a comparative analysis of insecticide resistance-related gene families with hemipteran crop pests. BMC Genom. 2022, 23, 45. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Rider, D.S.; Walsh, T.K.; De Vos, M.; Gordon, K.H.; Ponnala, L.; Macmil, S.L.; Roe, B.A.; Jander, G. Comparative analysis of detoxification enzymes in Acyrthosiphon pisum and Myzus persicae. Insect Mol. Biol. 2010, 19 (Suppl. S2), 155–164. [Google Scholar] [CrossRef]
- Zhou, W.W.; Li, X.W.; Quan, Y.H.; Cheng, J.; Zhang, C.X.; Gurr, G.; Zhu, Z.R. Identification and expression profiles of nine glutathione S-transferase genes from the important rice phloem sap-sucker and virus vector Laodelphax striatellus (Fallen) (Hemiptera: Delphacidae). Pest Manag. Sci. 2012, 68, 1296–1305. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.X.; Yu, L.L.; Fan, H.W.; Wang, Z.; Xu, H.J.; Xi, Y.; Zhu, Z.R.; Zhou, W.W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genome Biol. 2014, 15, 521. [Google Scholar] [CrossRef]
- Zhou, W.W.; Liang, Q.M.; Xu, Y.; Gurr, G.M.; Bao, Y.Y.; Zhou, X.P.; Zhang, C.X.; Cheng, J.; Zhu, Z.R. Genomic insights into the glutathione S-transferase gene family of two rice planthoppers, Nilaparvata lugens (Stal) and Sogatella furcifera (Horvath) (Hemiptera: Delphacidae). PLoS ONE 2013, 8, e56604. [Google Scholar] [CrossRef]
- Schama, R.; Pedrini, N.; Juarez, M.P.; Nelson, D.R.; Torres, A.Q.; Valle, D.; Mesquita, R.D. Rhodnius prolixus supergene families of enzymes potentially associated with insecticide resistance. Insect Biochem. Mol. Biol. 2016, 69, 91–104. [Google Scholar] [CrossRef]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 76. [Google Scholar] [CrossRef]
- Zhang, X.R.; Zhang, J.Q.; Shao, Y.Y.; Xing, X.R.; Wang, J.; Liu, Z.X.; Li, Y.J.; Ofori, A.D.; Tu, Q.B.; Wang, J.; et al. Identification of glutathione-S-transferase genes by transcriptome analysis in Meteorus pulchricornis (Hymenoptera: Braconidae) and their expression patterns under stress of phoxim and cypermethrin. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100607. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Johnson, R.M.; Berenbaum, M.R.; Ranson, H.; Cristino, A.S.; Claudianos, C. Metabolic enzymes associated with xenobiotic and chemosensory responses in Nasonia vitripennis. Insect Mol. Biol. 2010, 19 (Suppl. S1), 147–163. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Teng, Z.W.; Gu, G.X.; Guo, L.; Wang, F.; Xiao, S.; Wang, J.L.; Wang, B.B.; Fang, Q.; Wang, F.; et al. Genomic and transcriptomic analyses of glutathione S-transferases in an endoparasitoid wasp, Pteromalus puparum. Arch. Insect Biochem. Physiol. 2020, 103, e21634. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Lu, C.; Li, B.; Fang, S.; Zuo, W.; Dai, F.; Zhang, Z.; Xiang, Z. Identification, genomic organization and expression pattern of glutathione S-transferase in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1158–1164. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, C.Y.; Lyu, Z.H.; Lin, T. Multiple Glutathione S-Transferase Genes in Heortia vitessoides (Lepidoptera: Crambidae): Identification and Expression Patterns. J. Insect Sci. 2018, 18, 23. [Google Scholar] [CrossRef]
- Cheng, T.; Wu, J.; Wu, Y.; Chilukuri, R.V.; Huang, L.; Yamamoto, K.; Feng, L.; Li, W.; Chen, Z.; Guo, H.; et al. Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nat. Ecol. Evol. 2017, 1, 1747–1756. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.X.; Wang, W.L.; Zhang, B.X.; Li, S.G. Identification and characterisation of seventeen glutathione S-transferase genes from the cabbage white butterfly Pieris rapae. Pestic. Biochem. Physiol. 2017, 143, 102–110. [Google Scholar] [CrossRef]
- You, Y.; Xie, M.; Ren, N.; Cheng, X.; Li, J.; Ma, X.; Zou, M.; Vasseur, L.; Gurr, G.M.; You, M. Characterization and expression profiling of glutathione S-transferases in the diamondback moth, Plutella xylostella (L.). BMC Genom. 2015, 16, 152. [Google Scholar] [CrossRef]
- Huang, X.L.; Fan, D.S.; Liu, L.; Feng, J.N. Identification and characterization of glutathione S-transferase genes in the antennae of codling moth (Lepidoptera: Tortricidae). Ann. Entomol. Soc. Am. 2017, 110, 409–416. [Google Scholar] [CrossRef]
- Wang, X.; Fang, X.; Yang, P.; Jiang, X.; Jiang, F.; Zhao, D.; Li, B.; Cui, F.; Wei, J.; Ma, C.; et al. The locust genome provides insight into swarm formation and long-distance flight. Nat. Commun. 2014, 5, 2957. [Google Scholar] [CrossRef]
- Kirkness, E.F.; Haas, B.J.; Sun, W.; Braig, H.R.; Perotti, M.A.; Clark, J.M.; Lee, S.H.; Robertson, H.M.; Kennedy, R.C.; Elhaik, E.; et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2010, 107, 12168–12173. [Google Scholar] [CrossRef]
- Wei, D.D.; Chen, E.H.; Ding, T.B.; Chen, S.C.; Dou, W.; Wang, J.J. De novo assembly, gene annotation, and marker discovery in stored-product pest Liposcelis entomophila (Enderlein) using transcriptome sequences. PLoS ONE 2013, 8, e80046. [Google Scholar] [CrossRef]
- Younus, F.; Chertemps, T.; Pearce, S.L.; Pandey, G.; Bozzolan, F.; Coppin, C.W.; Russell, R.J.; Maibeche-Coisne, M.; Oakeshott, J.G. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 53, 30–43. [Google Scholar] [CrossRef]
- Corcoran, J.A.; Jordan, M.D.; Thrimawithana, A.H.; Crowhurst, R.N.; Newcomb, R.D. The Peripheral Olfactory Repertoire of the Lightbrown Apple Moth, Epiphyas postvittana. PLoS ONE 2015, 10, e0128596. [Google Scholar] [CrossRef]
- Liu, S.; Gong, Z.J.; Rao, X.J.; Li, M.Y.; Li, S.G. Identification of Putative Carboxylesterase and Glutathione S-transferase Genes from the Antennae of the Chilo suppressalis (Lepidoptera: Pyralidae). J. Insect Sci. 2015, 15, 103. [Google Scholar] [CrossRef]
- Tan, X.; Hu, X.M.; Zhong, X.W.; Chen, Q.M.; Xia, Q.Y.; Zhao, P. Antenna-specific glutathione S-transferase in male silkmoth Bombyx mori. Int. J. Mol. Sci. 2014, 15, 7429–7443. [Google Scholar] [CrossRef]
- Leal, W.S.; Ishida, Y.; Pelletier, J.; Xu, W.; Rayo, J.; Xu, X.; Ames, J.B. Olfactory proteins mediating chemical communication in the navel orangeworm moth, Amyelois transitella. PLoS ONE 2009, 4, e7235. [Google Scholar] [CrossRef]
- Wang, G.R.; Guo, Y.Y.; Wu, K.M. Cloning of a cDNA fragment of an antenna-specific gene in Helicoverpa armigera. Chin. J. Agric. Biotechnol. 2002, 1, 37–43. [Google Scholar] [CrossRef]
- Rogers, M.E.; Jani, M.K.; Vogt, R.G. An olfactory-specific glutathione-S-transferase in the sphinx moth Manduca sexta. J. Exp. Biol. 1999, 202, 1625–1637. [Google Scholar] [CrossRef]
- Durand, N.; Pottier, M.A.; Siaussat, D.; Bozzolan, F.; Maibeche, M.; Chertemps, T. Glutathione-S-Transferases in the Olfactory Organ of the Noctuid Moth Spodoptera littoralis, Diversity and Conservation of Chemosensory Clades. Front. Physiol. 2018, 9, 1283. [Google Scholar] [CrossRef]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Poland, T.; Qazi, S.S.; Doucet, D.; Cusson, M.; Beliveau, C.; Mittapalli, O. Identification of odor-processing genes in the emerald ash borer, Agrilus planipennis. PLoS ONE 2013, 8, e56555. [Google Scholar] [CrossRef]
- Gu, X.C.; Zhang, Y.N.; Kang, K.; Dong, S.L.; Zhang, L.W. Antennal Transcriptome Analysis of Odorant Reception Genes in the Red Turpentine Beetle (RTB), Dendroctonus valens. PLoS ONE 2015, 10, e0125159. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Xu, S.; Xie, C.; Geng, H.; Zhao, Y.; Li, J.; Huang, W.F.; Lin, Y.; Li, Z.; Su, S. Comparative transcriptome analysis of Apis mellifera antennae of workers performing different tasks. Mol. Genet Genom. 2018, 293, 237–248. [Google Scholar] [CrossRef]
- Carr, A.L.; Mitchell, R.D., III; Dhammi, A.; Bissinger, B.W.; Sonenshine, D.E.; Roe, R.M. Tick Haller’s Organ, a New Paradigm for Arthropod Olfaction: How Ticks Differ from Insects. Int. J. Mol. Sci. 2017, 18, 1563. [Google Scholar] [CrossRef] [PubMed]
- Saisawang, C.; Ketterman, A.J. Micro-plasticity of genomes as illustrated by the evolution of glutathione transferases in 12 Drosophila species. PLoS ONE 2014, 9, e109518. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, R.; Singh, S.P.; Mondal, A.K.; Benes, H.; Zimniak, P. Cloning, expression and biochemical characterization of one Epsilon-class (GST-3) and ten Delta-class (GST-1) glutathione S-transferases from Drosophila melanogaster, and identification of additional nine members of the Epsilon class. Biochem. J. 2003, 370, 661–669. [Google Scholar] [CrossRef]
- Li, G.W.; Chen, X.L.; Xu, X.L.; Wu, J.X. Degradation of sex pheromone and plant volatile components by an antennal glutathione S-transferase in the oriental fruit moth, Grapholita molesta Busck (Lepidoptera: Tortricidae). Arch Insect Biochem.Physiol. 2018, 99, e21512. [Google Scholar] [CrossRef]
- Francis, F.; Vanhaelen, N.; Haubruge, E. Glutathione S-transferases in the adaptation to plant secondary metabolites in the Myzus persicae aphid. Arch Insect Biochem. Physiol. 2005, 58, 166–174. [Google Scholar] [CrossRef]
- Dweck, H.K.; Talross, G.J.; Wang, W.; Carlson, J.R. Evolutionary shifts in taste coding in the fruit pest Drosophila suzukii. Elife 2021, 10, e64317. [Google Scholar] [CrossRef]
- Lombardo, F.; Salvemini, M.; Fiorillo, C.; Nolan, T.; Zwiebel, L.J.; Ribeiro, J.M.; Arca, B. Deciphering the olfactory repertoire of the tiger mosquito Aedes albopictus. BMC Genom. 2017, 18, 770. [Google Scholar] [CrossRef]
- Jordan, M.D.; Stanley, D.; Marshall, S.D.; De Silva, D.; Crowhurst, R.N.; Gleave, A.P.; Greenwood, D.R.; Newcomb, R.D. Expressed sequence tags and proteomics of antennae from the tortricid moth, Epiphyas postvittana. Insect Mol. Biol. 2008, 17, 361–373. [Google Scholar] [CrossRef]
- Ono, H.; Ozaki, K.; Yoshikawa, H. Identification of cytochrome P450 and glutathione-S-transferase genes preferentially expressed in chemosensory organs of the swallowtail butterfly, Papilio xuthus L. Insect Biochem. Mol. Biol. 2005, 35, 837–846. [Google Scholar] [CrossRef]
- Liu, H.; Tang, Y.; Wang, Q.; Shi, H.; Yin, J.; Li, C. Identification and Characterization of an Antennae-Specific Glutathione S-Transferase From the Indian Meal Moth. Front. Physiol. 2021, 12, 727619. [Google Scholar] [CrossRef]
- Legeai, F.; Malpel, S.; Montagne, N.; Monsempes, C.; Cousserans, F.; Merlin, C.; Francois, M.C.; Maibeche-Coisne, M.; Gavory, F.; Poulain, J.; et al. An Expressed Sequence Tag collection from the male antennae of the Noctuid moth Spodoptera littoralis: A resource for olfactory and pheromone detection research. BMC Genom. 2011, 12, 86. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Kolm, R.H.; Danielson, U.H.; Zhang, Y.; Talalay, P.; Mannervik, B. Isothiocyanates as substrates for human glutathione transferases: Structure-activity studies. Biochem. J. 1995, 311 Pt 2, 453–459. [Google Scholar] [CrossRef]
- Harzsch, S.; Krieger, J. Crustacean olfactory systems: A comparative review and a crustacean perspective on olfaction in insects. Prog. Neurobiol. 2018, 161, 23–60. [Google Scholar] [CrossRef]
- Graziadei, P.P.; Graziadei, G.A. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef]
- Neiers, F.; Gourrat, K.; Canon, F.; Schwartz, M. Metabolism of Cysteine Conjugates and Production of Flavor Sulfur Compounds by a Carbon-Sulfur Lyase from the Oral Anaerobe Fusobacterium nucleatum. J. Agric. Food Chem. 2022, 70, 9969–9979. [Google Scholar] [CrossRef]
- Schwartz, M.; Canon, F.; Feron, G.; Neiers, F.; Gamero, A. Impact of Oral Microbiota on Flavor Perception: From Food Processing to In-Mouth Metabolization. Foods 2021, 10, 2006. [Google Scholar] [CrossRef]
- Ai, S.; Zhang, Y.; Chen, Y.; Zhang, T.; Zhong, G.; Yi, X. Insect-Microorganism Interaction Has Implicates on Insect Olfactory Systems. Insects 2022, 13, 1084. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).