Abstract
Thickness of lipid bilayer membranes is a key physical parameter determining membrane permeability and stability with respect to formation of through pores. Most membrane inclusions or impurities like amphipathic peptides, transmembrane peptides, lipid inclusions of a different molecular shape, lipid domains, and protein-lipid domains, locally deform the membrane. The detailed structure of the locally deformed region of the membrane is a kind of “fingerprint” for the inclusion type. However, most experimental methods allow determining only averaged parameters of membranes with incorporated inclusions, thus preventing the direct obtaining of the characteristics of the inclusion. Here we developed a model that allows the obtaining of characteristic parameters of three types of membrane inclusions (amphipathic peptides, transmembrane peptides, monolayer lipid patches) from experimentally observable dependencies of the average thickness of lipid bilayer on the surface concentration of the inclusions. In the case of amphipathic peptides, the model provided the peptide parameters that were in qualitative agreement with the available experimental data.
1. Introduction
The lipid bilayer is a main structural element of biological membranes. Membranes are used by cells as weakly permeable, thin envelopes that perform a barrier function. This function allows cells to maintain the composition of cytoplasm, different from the composition of extracellular milieu, and the composition of cell organells different from that of cytoplasm. Membrane stability and, consequently, the efficiency of performing its barrier function strongly depends on membrane thickness: generally, lipid bilayers with smaller thickness are softer and less stable [1,2].
Substances that violate the barrier function of membranes are considered promising antimicrobial agents. These antibiotics include, but are not limited to, amphipathic peptides [3,4,5], membrane-disrupting and penetrating lipocations [6,7], channel-forming peptides [8,9,10,11], polyene antibiotics [12,13]. Amphipathic and channel-forming peptides (like gramicidin A) are definitely known to locally decrease the thickness of lipid bilayers [14,15,16,17], that is observed by means of X-Ray diffraction, molecular dynamics, or inferred from comparison of known bilayer thickness and the length of the membrane-permeating structure formed by the above-mentioned compounds.
Generally, most membrane inclusions or impurities (amphipathic peptides, transmembrane peptides, lipid inclusions of different molecular shape, lipid and protein-lipid domains [18,19], etc.) deform the membrane [20,21]. The lateral length of deformation decay in “conventional” membranes (e.g., formed from dioleoylphosphatidylcholine, DOPC) is of the order of several nanometers [21,22,23]. When the membrane-deforming objects are far-separated, their induced deformations are independent, and their energy is additive. Upon approaching, the deformed zones near the inclusions overlap; the overlapping leads to an effective membrane-mediated lateral interaction [22,23] on the distances of several nanometers, that exceeds typical distances of interactions like screened Debye, van der Waals and dipole–dipole interactions under physiological conditions [24,25,26,27]. The membrane-mediated interaction is significant at large surface concentrations of inclusions. But even in a low concentration, membrane inclusions alter membrane mechanical properties like local bilayer thickness and elastic rigidity [14,15,28,29]. A local disturbance of the lipid bilayer structure contributes to measurable average elastic properties of the membrane that can be observed experimentally. However, the relationship of the local disturbance and the resulting measured deviation of the average characteristics is not straightforward. At the same time, the structure of membrane deformations locally induced by the membrane inclusion depends on the inclusion shape and elastic properties. The establishment of the relationship between nanoscopic, local, and observable average deformations of the membrane may allow extraction of the information about the structure or configuration of the membrane-embedded inclusions. In this present study, we determined how the average membrane thickness changes with the addition of α-helical amphipathic peptides, transmembrane peptides, or a small amount of an impurity lipid differing from the bulk lipid of the membrane.
Spatial distribution of membrane deformations induced by inclusions can be determined in a framework of an adequate theory of elasticity. Such theories formulate possible deformational modes of the considered elastic medium (lipid membrane), and relate deformations and surface density of the elastic energy [20,21,22,23]. The total elastic energy thus becomes a functional on the surface density of the elastic energy. In order to determine the spatial distribution and energy of membrane deformations, one should minimize this functional. A correct minimization problem requires appropriate boundary conditions for the energy functional. As a rule, only lipid deformations are allowed, while membrane-embedded peptides and proteins are deemed absolutely rigid and undeformable. Such rigid membrane inclusions are supposed to impose boundary conditions onto the deformations. In this elastic approach, details of the structure and configuration of the membrane-embedded peptide or protein are completely reflected by the boundary conditions they impose onto the membrane deformations [20,21,22,23], for example, an orientation of lipid molecules and local membrane thickness. Utilizing experimental data on the dependence of membrane thickness on the surface concentration of inclusions, our developed model principally allows the obtaining of boundary conditions imposed by amphipathic peptides and transmembrane peptides. From comparison of calculation results with available experimental data [14,15], we quantitatively determined the boundary director characterizing an average orientation of adjacent lipid molecules for two types (melittin and magainin) of amphipathic peptides incorporated into a lipid monolayer of a membrane.
2. Methods
Let us consider a one-component lipid bilayer containing some membrane-deforming inclusions. Such inclusions may be lipid inclusions differing from the main lipid of the membrane; α-helical amphipathic peptide like magainin or melitin; a rigid transmembrane peptide like WALP [30,31]. The ground state of the pure membrane without any inclusions is planar. As the inclusions induce deformations, the ground state of the membrane with even single inclusion would not be a planar configuration [20,21]. Here we determine ground states of lipid membranes with inclusions of different types.
We introduce a Cartesian coordinate system Oxyz, the origin of which coincides with the center of the membrane inclusion; the Oxy plane is parallel to the plane of the pure membrane in its ground state. The state of the lipid bilayer can be completely characterized by five functions: Hu(x, y) and Hl(x, y), which are z-coordinates of the upper and lower monolayer neutral surfaces, respectively; M(x, y), which is a z-coordinate of the monolayer interface; nu(x, y) and nl(x, y), which are the unit vector fields set on the neutral surfaces of the upper and lower monolayers, respectively. Vectors nu(x, y) and nl(x, y) are called directors, and they characterize an average orientation of lipid hydrophobic chains [32,33]. In the ground state of the pure membrane without inclusions: Hu(x, y) = h, Hl(x, y) = −h, M(x, y) = 0, nu(x, y) = (0, 0, −1) and nl(x, y) = (0, 0, +1), where h is the thickness of the hydrophobic part of the lipid monolayer, i.e., the distance from the monolayer interface to the monolayer neutral surface [34].
When the membrane is deformed by a peptide or lipid inclusion, functions Hu(x, y), Hl(x, y), M(x, y), nu(x, y), and nl(x, y) generally deviate from constants. When a single inclusion is incorporated into the membrane, the average thickness of the membrane can be determined as follows:
where S is the total area of the membrane; ; the integration is performed over the membrane area S. The integral Δ is accumulated only in the vicinity of membrane inclusions, because the value of (Hu − Hl − 2h) generally decays exponentially with increasing distance from the inclusion [20,21,22,23].
In the case of N identical inclusions incorporated into the membrane in a low surface concentration C, we can neglect lateral interactions between the inclusions and write down the following linear expression for the average thickness of the membrane :
So, in a low concentration limit the average membrane thickness is completely characterized by the concentration of inclusions and the thickness disturbance Δ induced by single inclusion.
In order to calculate Δ for several types of membrane inclusions, we utilized the theory of elasticity of lipid membranes originally developed by Hamm and Kozlov [33] and further generalized in the works [21,22,23] to include additional deformational modes. The corresponding functional of the monolayer elastic energy:
accounts for the deformations of splay (the first and second terms), tilt (the third term), lateral tension (the fourth term), lateral compression (the fifth term), Gaussian splay (the sixth term), and twist (the last term), respectively; the integration is performed over the neutral surface of the lipid monolayer Sm. The energy functional (3) is quadratic in deformations, and it is valid for small deformations only; in particular, within the required accuracy, nl,u = (nx(x, y), ny(x, y), ±1). Here, B, Kt, Ka, KG, and Krot are elastic moduli (per monolayer) of splay, tilt, lateral compression, Gaussian splay, and twist, respectively; J0 is the spontaneous curvature of the monolayer; σ is the monolayer lateral tension; α is the relative change of the area per lipid molecule at the monolayer neutral surface; is the Gaussian splay; is the divergence of the director along the neutral surface; is the vector of rotor of the director (T is transposition); is the vector of gradient of H(x, y). Taking into account that the bulk modulus of membranes is very high [35], we assume that the hydrophobic part of a lipid monolayer is locally volumetrically incompressible, i.e., the volume of its any small element does not change upon deformations. This condition imposes a constraint on the deformation fields. Within this condition, the elastic energy functional (3) for the bilayer membrane can be rewritten as:
where the indices “u” and “l” correspond to the upper and lower monolayers, respectively; the integration is performed over the neutral surfaces of the respective monolayers.
All deformations should be limited everywhere, and should vanish at large distances from the inclusion, i.e.,
Depending on the type of the membrane inclusions, they impose different boundary conditions on the elastic energy functional (4). Below, we explicitly consider boundary conditions imposed onto the deformation fields by amphipathic peptides, transmembrane peptides and lipid inclusions.
2.1. Amphipathic Peptide
For definiteness, we considered α-helical amphipathic peptides. Such peptides consist of hydrophobic and polar (or charged) amino acids combined in such a way that one side-surface of α-helix is hydrophobic, while the opposite side-surface is polar. In order to hide their hydrophobic surfaces from contact with water, amphipathic peptides partially incorporate into the membrane monolayer. The incorporation requires a lateral shifting of polar lipid heads away, resulting in a jump of lipid orientation on the left and right boundaries of the amphipathic peptide. This effect can be formally expressed as the following boundary conditions imposed on the director field [23,36] at the peptide outline Γ at the neutral surface of the upper monolayer (Figure 1a):
where nn,u and nτ,u are, respectively, the normal and tangential to the contour Γ components of the director field nu of the upper monolayer. The exact value of n0 is unknown; it depends on the details of peptide structure and lipid-peptide intermolecular interactions. However, based on geometric meaning of the director, n0 can be estimated as (Figure 1b) [23,36]:
where rp is the radius of α-helix. The boundary director is directed beneath the incorporated amphipathic peptide (Figure 1b). Of note, as the elastic energy functional (4) is quadratic in deformations, the value of Δ should be linear on n0: Δ = ηn0, where η is some constant independent on n0. The same should hold for amplitudes of all deformation fields in (4).
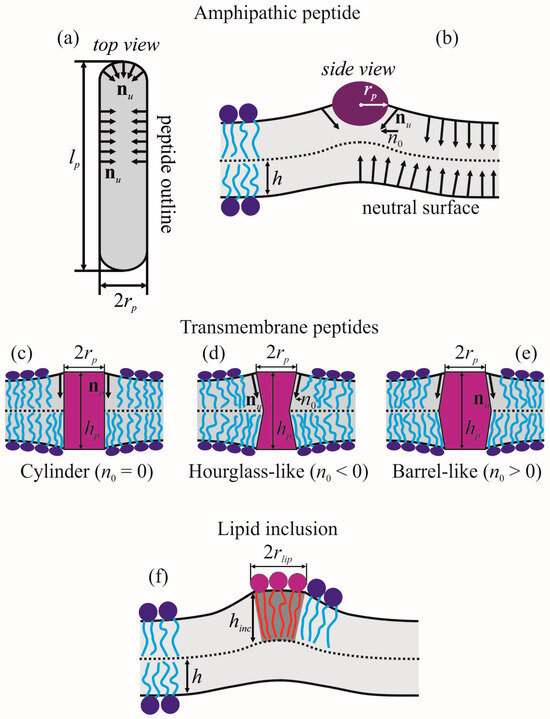
Figure 1.
Schematic representation of boundary conditions imposed by amphipathic peptide, transmembrane peptides and lipid inclusion. (a) amphipathic peptide, top view; (b) amphipathic peptide, side view along the longitudinal axis of the peptide α-helix; (c) cylindrical transmembrane peptide (n0 = 0); (d) hourglass-like transmembrane peptide (n0 < 0); (e) barrel-like transmembrane peptide (n0 > 0); (f) lipid inclusion.
2.2. Transmembrane Peptide
Transmembrane peptide incorporated into a lipid bilayer imposes boundary conditions both on the director field and membrane thickness at the peptide boundary outlines at the neutral surfaces of the upper and lower monolayers [22,37]:
where hp is the length of transmembrane part of the peptide; n0u, n0l are projection of boundary directors onto Oxy plane (Figure 1c–e). We assumed that outlines of the peptide boundary on the neutral surfaces of the upper (Γu) and lower (Γl) monolayers are identical circles of the radius rp and their center coordinates are (x, y) = (0, 0); thus, Γu = Γl = Γ. For definiteness, we considered the case of symmetric transmembrane peptides, n0u = n0l = n0. Negative n0 corresponds to the hourglass-like shape of the peptide (Figure 1d), positive n0—to barrel-like shape (Figure 1e), and n0 = 0—to cylindrical peptides (Figure 1c) [37]. For a quadratic energy functional (4), Δ should linearly depend on n0 and (hp − 2h), i.e., it can be expressed in the form Δ = βn0 + γ(hp − 2h), where β, γ are constants that do not depend on n0 and hp.
2.3. Lipid Inclusion
Lipid inclusions do not impose any specific boundary conditions, except continuity of the neutral surface and director (Figure 1f). Unlike undeformable peptide inclusions, lipid inclusions can be deformed and the energy of their deformation should be calculated in the framework of the same elastic theory as that of the surrounding membrane, e.g., utilizing the elastic energy functional (4). However, generally, the elastic parameters of lipid inclusions differ from those of the membrane, and this difference should be taken into account. As a lipid inclusion, we considered a circular patch of lipid monolayer of the radius rlip, the center of which has coordinates (x, y) = (0, 0). The patch is located in the upper monolayer of the membrane (Figure 1f).
We can determine how Δ depends on spontaneous curvature of lipid inclusion J0inc. If we fix the value of the radial component of the director at the inclusion boundary, nr(rlip) = n, and the thickness of the bilayer at the inclusion boundary [Hu(rlip) − Hl(rlip)] = d, we can write down the following expression for the elastic energy W:
where hinc is the thickness of the lipid inclusion monolayer; α1,2, β1,2, γ1,2 are constants depending on elastic parameters of the bulk lipid and the lipid of the inclusion except spontaneous curvature J0inc and thickness hinc. The first and second square brackets correspond to energies of membrane deformations in the circle of the radius rlip and in the outer region, respectively. Minimizing (9) with respect to n and d, we obtain what n ∝ (kJ0inc + b), where k and b are constants that do not depend on J0inc. As the elastic energy functional (4) is quadratic on deformations, we can conclude that Δ = (kJ0inc + b).
In order to obtain the values of the parameters η, β, γ, k, b, we numerically minimized the elastic energy functional (4) using the finite elements method, essentially as described in details in the works [22,23,36,37]. The minimization routine was written in Mathematica 11.3 (Wolfram Research, Champaign, IL, USA) using its standard functions; the corresponding program can be obtained from the authors by request.
2.4. Parameters
To obtain quantitative results, the following parameters were used. For DOPC, which was considered as the main lipid component of the membrane: B = 10 kBT [2], Kt = 40 mN/m [33], Ka = 133 mN/m [2], σ = 0.1 mN/m, Krot = B/2 = 5 kBT, KG = −B/2 = −5 kBT [23,36,37,38], J0 = −0.091 nm−1 [39], h = 1.45 nm [2,34]. In the work [14], the average thickness of the membrane made from palmitoyloleoylphosphatidylcholine (POPC):palmitoyloleoylphosphatidylserine (POPS) 3:1 was measured as a function of the mole fraction of adsorbed amphipathic peptide melittin. For this lipid mixture, we utilized the following parameters: B = 11 kBT [2], Kt = 40 mN/m [33], Ka = 117 mN/m [2], σ = 0.1 mN/m, Krot = B/2 = 5.5 kBT, KG = −B/2 = −5.5 kBT [23,36,37,38], J0 = 0 [39], h = 1.46 nm [2,34]. For the amphipathic peptide: lp = 5 nm, rp = 0.65 nm. For the transmembrane peptide: rp = 0.65 nm. For the lipid inclusion: rlip = 0.5 nm or rlip = 0.662 nm in the case of thin lipid inclusion, like photoswitchable lipid OptoDArG, Kt = 40 mN/m [33], Ka = 133 mN/m, σ = 0.1 mN/m, Krot = B/2 = 5 kBT, KG = −B/2 = −5 kBT.
3. Results
3.1. Amphipathic Peptide
Numerically calculated shapes Hu(x, y), Hl(x, y) of neutral surfaces of the upper and lower monolayers of DOPC membrane with incorporated amphipathic peptide are illustrated in Figure 2a,b for particular case of n0 = −0.4, as derived from Equation (7) for rp = 0.65 nm, h = 1.45 nm. The membrane thickness [Hu(x, y) − Hl(x, y)] is presented in Figure 2c. The obtained dependence of Δ on n0 for DOPC membrane is: Δ = 9.16 × n0 (in the units of nm3); for POPC:POPS 3:1 membrane: Δ = 9.36 × n0 (in the units of nm3). The similarity of the proportionality coefficient between Δ and n0 for DOPC and POPC:POPS membrane is not surprising, as the elastic parameters of these lipids differ, but slightly.
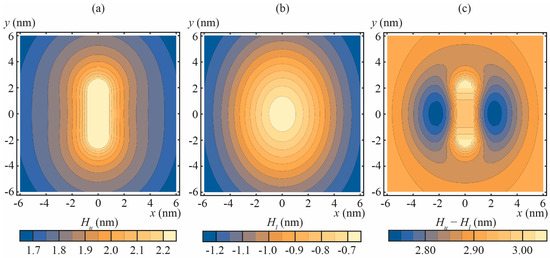
Figure 2.
The shapes of the neutral surfaces of the upper and lower monolayers and the thickness of the DOPC membrane with incorporated amphipathic peptide imposing the normal projection of the boundary director |n0| = 0.4. (a) the shape of the neutral surface of the upper monolayer, Hu(x, y); (b) the shape of the neutral surface of the lower monolayer, Hl(x, y); (c) the thickness of the membrane, [Hu(x, y) − Hl(x, y)].
3.2. Transmembrane Peptide
Radial distribution of the thickness of the DOPC membrane with incorporated transmembrane peptide is shown in Figure 3 for different values of the transmembrane peptide length and boundary director. Due to the symmetry with respect to the monolayer interface, [Hu(x, y) − Hl(x, y)] = 2Hu(x, y) = −2Hl(x, y). The determined dependence of Δ on n0 and hp is: Δ = 10.72 × n0 + 3.96 × (hp − 2h) (in the units of nm3).
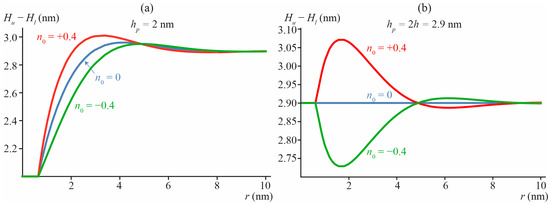
Figure 3.
The thickness of the DOPC membrane [Hu(x, y) − Hl(x, y)] with incorporated transmembrane peptide of the length: (a) hp = 2 nm; (b) hp = 2h = 2.9 nm (no mismatch with the surrounding membrane), imposing the boundary director: n0 = −0.4 (hourglass-like peptide, green curves), 0 (cylindrical peptide, blue curves), +0.4 (barrel-like peptide, red curves). The radial coordinate r is the distance to the center of the peptide: r = (x2 + y2)1/2.
Figure 3a illustrates a deviation of the membrane thickness in the vicinity of short transmembrane peptide, the length hp = 2 nm of which is smaller than the bilayer equilibrium thickness 2h = 2.9 nm. In Figure 3b, the case of no mismatch between the transmembrane peptide length and bilayer thickness (hp = 2h = 2.9 nm) is illustrated. From Figure 3b, it follows that even when hp = 2h, but n0 ≠ 0 (red and green curves in Figure 3b), deformations of the membrane arise leading to an alteration of the local thickness of the bilayer.
3.3. Lipid Inclusion
We considered two types of lipid inclusions. The first type is “common” lipid inclusion with splay modulus ranging from 5 to 20 kBT, the monolayer thickness hinc of which varies from 1.2 to 1.7 nm and spontaneous curvature ranges from −0.6 to +0.2 nm−1. This type corresponds to “common” lipids like DOPC, POPC, etc. In addition, we especially considered the second type of so-called “thin” lipid inclusions with hinc = 0.7 nm. This type models photoswitchable lipids having cis-trans isomerizable diazobenzene group in one or two hydrophobic chains, like PhoDAG and OptoDArG [40,41,42], respectively. In cis-conformation such lipids are characterized by a small monolayer thickness and strongly negative spontaneous curvature [43], while in trans-conformation these lipids can be considered as “common” ones. Photoswitchable lipids are a convenient biophysical tool, although their physical parameters are not characterized well to date.
Calculated shapes of membrane neutral surfaces, Hu(x, y) and Hl(x, y), and membrane thickness [Hu(x, y) − Hl(x, y)] are presented in Figure 4 for particular cases of “common” lipid inclusion with Binc = 10 kBT, J0inc = 0, and hinc = h, h + 0.1 nm, h − 0.1 nm. From Figure 4c, it follows that even in the case of no thickness mismatch, hinc = h, the membrane thickness [Hu(x, y) − Hl(x, y)] can still locally deviate from 2h if J0inc ≠ J0 (see blue curve in Figure 4c). In addition, a difference in the spontaneous curvatures of the bulk lipid and lipid inclusion can lead to a local increase in the membrane thickness even in the case of hinc < h (see red curve in Figure 4c). Of note, the typical length of bilayer thickness relaxation is of the order of 3–4 nm (Figure 4c).
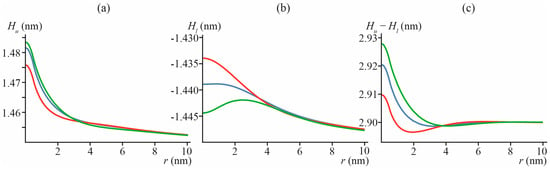
Figure 4.
Shapes of neutral surfaces of membrane monolayers, (a) Hu(x, y), and (b) Hl(x, y), and membrane thickness (c) [Hu(x, y) − Hl(x, y)], in particular case of Bimp = 10 kBT, J0inc = 0 ≠ J0 = −0.091 nm−1, hinc = h = 1.45 nm (blue), hinc = 1.35 nm (red) and hinc = 1.55 nm (green), as a function of distance to lipid inclusion center r = (x2 + y2)1/2.
In Figure 5, we presented the calculated values of Δ for “common” lipid inclusion with splay modulus Binc = 5, 10, and 20 kBT, spontaneous curvature J0inc varying from −0.6 to 0.2 nm−1 and thickness of the inclusion hinc varying from 1.2 to 1.7 nm. Of note, in the case of zero mismatch, hinc = h, the average thickness of the membrane is strictly 2h (i.e., Δ = 0) independently on Binc and J0inc values, even though the deformations do arise and membrane local thickness deviates from 2h (c.f. blue curve in Figure 4c).
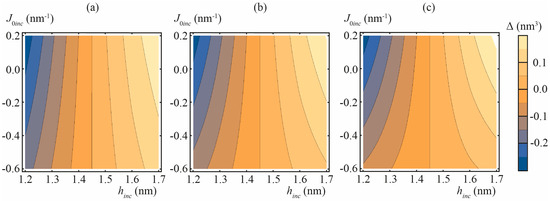
Figure 5.
Calculated dependencies of Δ on thickness hinc and spontaneous curvature J0inc of “common” lipid inclusion. (a) Binc = 5 kBT; (b) Binc = 10 kBT; (c) Binc = 20 kBT.
The calculated values of Δ for “thin” lipid inclusion (hinc = 0.7 nm) are presented in Figure 6 for different spontaneous curvatures and splay moduli of the inclusion.
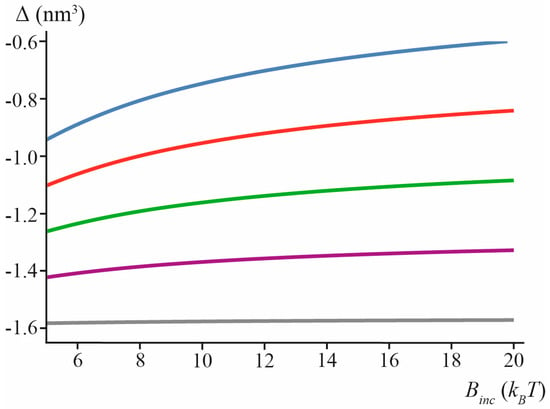
Figure 6.
Calculated values of Δ in the case of “thin” lipid inclusion (hinc = 0.7 nm) for J0inc = −0.6 nm−1 (blue), J0inc = −0.4 nm−1 (red), J0inc = −0.2 nm−1 (green), J0inc = 0 (purple), J0inc = +0.2 nm−1 (gray) as a function of inclusion splay modulus, Binc.
4. Discussion
Most membrane inclusions cause elastic deformations of lipid bilayers. A detailed structure of the locally deformed region of the membrane is a kind of “fingerprint” for the inclusion type. However, most experimental methods allow determining only averaged parameters of membranes with incorporated inclusions, thus preventing from direct obtaining of the characteristics of the inclusion. A derivation of the structural information from the measured average parameters is an inverse problem, the solution of which is always model-dependent. Molecular dynamics (MD) allows determining configurations of membrane-embedded inclusions within an atomistic resolution. However, structural information obtained from MD may be too detailed: it is preferable to characterize membrane inclusions by a few number of effective parameters, while MD provides coordinates of all atoms of the modeled system, i.e., enormous number of the parameters. The data of MD should thus be somehow averaged to converge to one or several effective characteristics of the system. However, the averaging may yield the parameter differing from those obtained from physical experiments [17] or from the theory of elasticity [44,45].
In the present work, we developed a model that allows solving the inverse problem: deriving one to several effective elastic parameters from the measured average thickness of the membrane for three types of membrane-deforming inclusions: amphipathic peptides, transmembrane peptides, lipid inclusions. In the framework of the model, amphipathic peptides of equal length are characterized by the only parameter, the normal projection of the boundary director, n0. Dependencies of the average thickness of the membrane on the mole fraction of incorporated amphipathic peptides were determined in works [14,15] for melittin in DOPC membrane [14] and magainin in POPC:POPS 3:1 membrane [15]. For mole fractions smaller than about 1/65 peptide/lipid, the dependencies are linear, and thus they can be readily fitted by the linear relation Δ = ηn0 where the only fitting parameter is n0 (Δ is measured in [14,15]; η was calculated here). This yields n0 = −0.42 for melittin and n0 = −0.48 for magainin. Of note, the estimation of the boundary director based on its oversimplified geometric interpretation, Equation (7), yields n0 = −0.4 that is surprisingly close to the determined value for melittin. In our recent work on membrane-mediated lateral interactions of amphipathic peptides, we used the same value of the boundary director [46]. This difference in the projections of the boundary directors is in a qualitative agreement with available data on magainin and melittin structures. When incorporated into a membrane, the longitudinal axis of magainin α-helix is about parallel to the membrane plane, and the molecule can be approximately considered as a cylinder [47,48]. Melittin has a relatively flexible loop in its structure, and can be approximately considered as two α-helical non-equal parts connected by this loop [48,49]. The smaller α-helical part of melittin is more hydrophobic than the larger one, and this part is incorporated deeper into the membrane [48,49], that should result in smaller (by the absolute value) projection of the boundary director n0; this agrees qualitatively with the predictions of our model. Magainin and melittin are known as classical antimicrobial peptides that effectively disrupt membranes by forming through pores in them [3,50,51,52,53,54]. The projection of the boundary director n0 is a key parameter determining the energy landscape of pore formation: the larger the |n0|, the higher the poration efficiency should be [55].
The model developed here can be utilized to retrieve characteristic parameters of the considered types of membrane inclusions. This requires measurements of dependencies of the average membrane thickness on the inclusion surface concentration in a low concentration (linear) regime. As transmembrane peptides and lipid inclusions are each described by two effective parameters (n0 and hp, J0inc and hinc, respectively) their determination requires at least two dependencies of the average membrane thickness on the inclusion surface concentration, measured in two different membranes. Unfortunately, to date, the measurements were not performed for transmembrane peptides, lipid inclusions, or photoswitchable lipids (as far as we know).
Gramicidin A is a peptide, the transmembrane dimers of which are ion-conducting channels [9]. The length of the dimer may differ from the membrane thickness that leads to membrane deformations in the vicinity of both gramicidin monomers and dimers [10,16,17,21,23]. The deformations contribute to the energy of monomeric and dimeric states, and the channel characteristics depend on the elastic properties of the membrane [10,23]. For this reason, gramicidin is considered a molecular force sensor [10]. To properly describe gramicidin based on the theory of elasticity of membranes, one should know boundary conditions imposed by gramicidin monomer and dimers. The boundary conditions can principally be determined from the model developed here. The change of lipid bilayer thickness upon incorporation of gramicidin was measured in the work [56]. It was shown that gramicidin induces thinning of dimiristoylphosphatidylcholine (DMPS) and thickening of dilauroylphosphatidylcholine (DLPC) membranes. However, the peptide/lipid ratio in these experiments was very high, 1/10 peptide/lipid. This corresponds to only about 2.5 nm gramicidin–gramicidin separations, meaning that deformations induced by gramicidin molecules should definitely overlap. Besides, the deformations induce by gramicidin dimers are generally different from those induced by monomers. Thus, there are two types of membrane-deforming inclusions in such the system: gramicidin monomers and dimers. These inclusions are in equilibrium, and the equilibrium surface concentration of dimers depends on elastic parameters of the membrane. In principle, our elastic model can be generalized to describe this case of non-ideal mixture of gramicidin monomers and dimers, where all molecular species laterally interact with each other; the necessary theoretical background was partially developed in our recent work [23]. However, the experiment of the work [56] provides only one measured value—the membrane thickness—at single concentration of gramicidin. From this value, it is impossible to determine the constant of equilibrium and two sets of boundary conditions imposed by monomers and dimers onto the membrane; additional experimental data are required.
Recently, the energy of transition of single gramicidin dimer from DLPC to DMPC membrane was determined as 3.7 kBT from molecular dynamics simulations [17]. In this work, the dimer is characterized by the only parameter, its length hp. However, the length of gramicidin dimer was determined experimentally as hp = 2.2 nm, independently on the membrane composition [56]. In the oversimplified model, where gramicidin dimer is characterized by hp only, it is impossible to obtain the measured value of the transition energy using the value hp = 2.2 nm. In the work [17], it was supposed that the effective “deformational” dimer length should be larger than its actual length by 0.4 nm. However, if transmembrane dimer of gramicidin is characterized by two parameters, n0 and hp, one can estimate the value n0 = −0.45 [37] from the measured transition energy [17], using the actual length of the dimer hp = 2.2 nm. Thus, physical experiments and molecular dynamics modeling can be combined to derive boundary conditions imposed by various membrane-deforming inclusions.
Author Contributions
Conceptualization, O.V.K. and S.A.A.; methodology, O.V.K.; software, O.V.K.; validation, O.V.K. and S.A.A.; formal analysis, O.V.K. and S.A.A.; investigation, O.V.K. and S.A.A.; resources, O.V.K. and S.A.A.; data curation, O.V.K. and S.A.A.; writing—original draft preparation, O.V.K. and S.A.A.; visualization, O.V.K.; supervision, O.V.K. and S.A.A.; project administration, O.V.K.; funding acquisition, O.V.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Russian Science Foundation (grant #22-24-00661, https://rscf.ru/project/22-24-00661/, accessed on 10 November 2023).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Evans, E.; Heinrich, V.; Ludwig, F.; Rawicz, W. Dynamic tension spectroscopy and strength of biomembranes. Biophys. J. 2003, 85, 2342–2350. [Google Scholar] [CrossRef] [PubMed]
- Rawicz, W.; Olbrich, K.C.; McIntosh, T.; Needham, D.; Evans, E. Effect of chain length and unsaturation on elasticity of lipid bilayers. Biophys. J. 2000, 79, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Peinado, C.; Dias, S.A.; Domingues, M.M.; Benfield, A.H.; Freire, J.M.; Rádis-Baptista, G.; Gaspar, D.; Castanho, M.A.R.B.; Craik, D.J.; Henriques, S.T.; et al. Mechanisms of bacterial membrane permeabilization by crotalicidin (Ctn) and its fragment Ctn (15–34), antimicrobial peptides from rattlesnake venom. J. Biol. Chem. 2018, 293, 1536–1549. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Balandin, S.V.; Ovchinnikova, T. Structure and biological functions of β-hairpin antimicrobial peptides. Acta Naturae 2015, 7, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Parvez, F.; Alam, J.M.; Dohra, H.; Yamazaki, M. Elementary processes of antimicrobial peptide PGLa-induced pore formation in lipid bilayers. Biochim. Biophys. Acta 2018, 1860, 2262–2271. [Google Scholar] [CrossRef]
- Sokolov, S.S.; Smirnova, E.A.; Markova, O.V.; Kireeva, N.A.; Kirsanov, R.S.; Khailova, L.S.; Knorre, D.A.; Severin, F.F. Lipophilic cations rescue the growth of yeast under the conditions of glycolysis overflow. Biomolecules 2020, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Rokitskaya, T.I.; Khailova, L.S.; Korshunova, G.A.; Antonenko, Y.N. Efficiency of mitochondrial uncoupling by modified butyltriphenylphosphonium cations and fatty acids correlates with lipophilicity of cations: Protonophoric vs leakage mechanisms. Biochim. Biophys. Acta 2023, 1865, 184183. [Google Scholar] [CrossRef]
- Qian, S.; Wang, W.; Yang, L.; Huang, H.W. Structure of the alamethicin pore reconstructed by x-ray diffraction analysis. Biophys. J. 2008, 94, 3512–3522. [Google Scholar] [CrossRef]
- Bamberg, E.; Läuger, P. Channel formation kinetics of gramicidin A in lipid bilayer membranes. J. Membr. Biol. 1973, 11, 177–194. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Collingwood, S.A.; Ingólfsson, H.I.; Kapoor, R.; Andersen, O.S. Lipid bilayer regulation of membrane protein function: Gramicidin channels as molecular force probes. J. R. Soc. Interface 2010, 7, 373–395. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Antonenko, Y.N.; Kotova, E.A. Photodynamic inactivation of gramicidin channels: A flash-photolysis study. Biochim. Biophys. Acta 1996, 1275, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Tevyashova, A.N.; Bychkova, E.N.; Solovieva, S.E.; Zatonsky, G.V.; Grammatikova, N.E.; Isakova, E.B.; Mirchink, E.P.; Treshchalin, I.D.; Pereverzeva, E.R.; Bykov, E.E.; et al. Discovery of amphamide, a drug candidate for the second generation of polyene antibiotics. ACS Infect. Dis. 2020, 6, 2029–2044. [Google Scholar] [CrossRef] [PubMed]
- Chulkov, E.G.; Ostroumova, O.S. Phloretin modulates the rate of channel formation by polyenes. Biochim. Biophys. Acta 2016, 1858, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.Y.; Lee, M.T.; Huang, H.W. Evidence for membrane thinning effect as the mechanism for peptide-induced pore formation. Biophys. J. 2003, 84, 3751–3758. [Google Scholar] [CrossRef] [PubMed]
- Ludtke, S.; He, K.; Huang, H. Membrane thinning caused by magainin 2. Biochemistry 1995, 34, 16764–16769. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; He, S.; Bennett, W.D.; Bilodeau, C.L.; Andersen, O.S.; Lightstone, F.C.; Ingólfsson, H.I. Atomistic characterization of gramicidin channel formation. J. Chem. Theory Comput. 2020, 17, 7–12. [Google Scholar] [CrossRef]
- Park, S.; Yeom, M.S.; Andersen, O.S.; Pastor, R.W.; Im, W. Quantitative characterization of protein–lipid interactions by free energy simulation between binary bilayers. J. Chem. Theory Comput. 2019, 15, 6491–6503. [Google Scholar] [CrossRef]
- Lillemeier, B.F.; Pfeiffer, J.R.; Surviladze, Z.; Wilson, B.S.; Davis, M.M. Plasma membrane-associated proteins are clustered into islands attached to the cytoskeleton. Proc. Natl. Acad. Sci. USA 2006, 103, 18992–18997. [Google Scholar] [CrossRef]
- Petruzielo, R.S.; Heberle, F.A.; Drazba, P.; Katsaras, J.; Feigenson, G.W. Phase behavior and domain size in sphingomyelin-containing lipid bilayers. Biochim. Biophys. Acta 2013, 1828, 1302–1313. [Google Scholar] [CrossRef]
- Zemel, A.; Ben-Shaul, A.; May, S. Perturbation of a lipid membrane by amphipathic peptides and its role in pore formation. Eur. Biophys. J. 2005, 34, 230–242. [Google Scholar] [CrossRef]
- Kondrashov, O.V.; Galimzyanov, T.R.; Pavlov, K.V.; Kotova, E.A.; Antonenko, Y.N.; Akimov, S.A. Membrane elastic deformations modulate gramicidin A transbilayer dimerization and lateral clustering. Biophys. J. 2018, 115, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, O.V.; Kuzmin, P.I.; Akimov, S.A. Hydrophobic mismatch controls the mode of membrane-mediated interactions of transmembrane peptides. Membranes 2022, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, O.V.; Akimov, S.A. Regulation of antimicrobial peptide activity via tuning deformation fields by membrane-deforming inclusions. Int. J. Mol. Sci. 2022, 23, 326. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.J.; Fotiadis, D.; Scheuring, S.; Müller, S.A.; Engel, A. Electrostatically balanced subnanometer imaging of biological specimens by atomic force microscope. Biophys. J. 1999, 76, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- LeNeveu, D.M.; Rand, R.P.; Parsegian, V.A. Measurement of forces between lecithin bilayers. Nature 1976, 259, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J.N. Intermolecular and Surface Forces, 3rd ed.; Academic Press: San Diego, CA, USA, 2011; ISBN 978-0-12-391927-4. [Google Scholar]
- Lipowsky, R.; Sackmann, E. Handbook of Biological Physics. Chapter 11. Generic Interactions of Flexible Membranes; Elsevier Science B.V.: Amsterdam, The Netherlands, 1995; pp. 521–602. ISBN 0-444-81975-4. [Google Scholar]
- Bashkirov, P.V.; Kuzmin, P.I.; Chekashkina, K.; Arrasate, P.; Vera Lillo, J.; Shnyrova, A.V.; Frolov, V.A. Reconstitution and real-time quantification of membrane remodeling by single proteins and protein complexes. Nat. Protoc. 2020, 15, 2443–2469. [Google Scholar] [CrossRef] [PubMed]
- Vitkova, V.; Méléard, P.; Pott, T.; Bivas, I. Alamethicin influence on the membrane bending elasticity. Eur. Biophys. J. 2006, 35, 281–286. [Google Scholar] [CrossRef]
- Bereau, T.; Bennett, W.D.; Pfaendtner, J.; Deserno, M.; Karttunen, M. Folding and insertion thermodynamics of the transmembrane WALP peptide. J. Chem. Phys. 2015, 143, 243127. [Google Scholar] [CrossRef] [PubMed]
- Holt, A.; Rougier, L.; Réat, V.; Jolibois, F.; Saurel, O.; Czaplicki, J.; Killian, J.A.; Milon, A. Order parameters of a transmembrane helix in a fluid bilayer: Case study of a WALP peptide. Biophys. J. 2010, 98, 1864–1872. [Google Scholar] [CrossRef]
- Helfrich, W. Elastic properties of lipid bilayers: Theory and possible experiments. Z. Naturforsch. C 1973, 28, 693–703. [Google Scholar] [CrossRef]
- Hamm, M.; Kozlov, M.M. Elastic energy of tilt and bending of fluid membranes. Eur. Phys. J. E 2000, 3, 323–335. [Google Scholar] [CrossRef]
- Leikin, S.; Kozlov, M.M.; Fuller, N.L.; Rand, R.P. Measured effects of diacylglycerol on structural and elastic properties of phospholipid membranes. Biophys. J. 1996, 71, 2623–2632. [Google Scholar] [CrossRef]
- Nagle, J.F.; Wilkinson, D.A. Lecithin bilayers. Density measurement and molecular interactions. Biophys. J. 1978, 23, 159–175. [Google Scholar] [CrossRef]
- Kondrashov, O.V.; Akimov, S.A. Effect of solid support and membrane tension on adsorption and lateral interaction of amphipathic peptides. J. Chem. Phys. 2022, 157, 074902. [Google Scholar] [CrossRef]
- Kondrashov, O.V.; Pinigin, K.V.; Akimov, S.A. Characteristic lengths of transmembrane peptides controlling their tilt and lateral distribution between membrane domains. Phys. Rev. E 2021, 104, 044411. [Google Scholar] [CrossRef]
- Hu, M.; de Jong, D.H.; Marrink, S.J.; Deserno, M. Gaussian curvature elasticity determined from global shape transformations and local stress distributions: A comparative study using the MARTINI model. Faraday Discuss. 2013, 161, 365–382. [Google Scholar] [CrossRef]
- Kollmitzer, B.; Heftberger, P.; Rappolt, M.; Pabst, G. Monolayer spontaneous curvature of raft-forming membrane lipids. Soft Matter 2013, 9, 10877–10884. [Google Scholar] [CrossRef]
- Erkan-Candag, H.; Krivic, D.; Gsell, M.A.; Aleksanyan, M.; Stockner, T.; Dimova, R.; Tiapko, O.; Groschner, K. Characterization of DAG binding to TRPC channels by target-dependent cis–trans isomerization of OptoDArG. Biomolecules 2022, 12, 799. [Google Scholar] [CrossRef]
- Morstein, J.; Impastato, A.C.; Trauner, D. Photoswitchable lipids. ChemBioChem 2021, 22, 73–83. [Google Scholar] [CrossRef]
- Frank, J.A.; Yushchenko, D.A.; Hodson, D.J.; Lipstein, N.; Nagpal, J.; Rutter, G.A.; Rhee, J.-S.; Gottschalk, A.; Brose, N.; Schultz, C.; et al. Photoswitchable diacylglycerols enable optical control of protein kinase C. Nat. Chem. Biol. 2016, 12, 755–762. [Google Scholar] [CrossRef]
- Pfeffermann, J.; Eicher, B.; Boytsov, D.; Hannesschlaeger, C.; Galimzyanov, T.R.; Glasnov, T.N.; Pabst, G.; Akimov, S.A.; Pohl, P. Photoswitching of model ion channels in lipid bilayers. J. Photochem. Photobiol. B 2021, 224, 112320. [Google Scholar] [CrossRef]
- Campelo, F.; McMahon, H.T.; Kozlov, M.M. The hydrophobic insertion mechanism of membrane curvature generation by proteins. Biophys. J. 2008, 95, 2325–2339. [Google Scholar] [CrossRef]
- Sodt, A.J.; Pastor, R.W. Molecular modeling of lipid membrane curvature induction by a peptide: More than simply shape. Biophys. J. 2014, 106, 1958–1969. [Google Scholar] [CrossRef]
- Kondrashov, O.V.; Galimzyanov, T.R.; Jiménez-Munguía, I.; Batishchev, O.V.; Akimov, S.A. Membrane-mediated interaction of amphipathic peptides can be described by a one-dimensional approach. Phys. Rev. E 2019, 99, 022401. [Google Scholar] [CrossRef]
- Bechinger, B.; Zasloff, M.; Opella, S.J. Structure and orientation of the antibiotic peptide magainin in membranes by solid-state nuclear magnetic resonance spectroscopy. Protein Sci. 1993, 2, 2077–2084. [Google Scholar] [CrossRef]
- Santo, K.P.; Berkowitz, M.L. Difference between magainin-2 and melittin assemblies in phosphatidylcholine bilayers: Results from coarse-grained simulations. J. Phys. Chem. B 2012, 116, 3021–3030. [Google Scholar] [CrossRef]
- Chen, C.H.; Wiedman, G.; Khan, A.; Ulmschneider, M.B. Absorption and folding of melittin onto lipid bilayer membranes via unbiased atomic detail microsecond molecular dynamics simulation. Biochim. Biophys. Acta 2014, 1838, 2243–2249. [Google Scholar] [CrossRef]
- Huang, H.W. Molecular mechanism of antimicrobial peptides: The origin of cooperativity. Biochim. Biophys. Acta 2006, 1758, 1292–1302. [Google Scholar] [CrossRef]
- Hasan, M.; Karal, M.A.S.; Levadnyy, V.; Yamazaki, M. Mechanism of initial stage of pore formation induced by antimicrobial peptide magainin 2. Langmuir 2018, 34, 3349–3362. [Google Scholar] [CrossRef]
- Fuertes, G.; Giménez, D.; Esteban-Martín, S.; Sánchez-Munoz, O.L.; Salgado, J. A lipocentric view of peptide-induced pores. Eur. Biophys. J. 2011, 40, 399–415. [Google Scholar] [CrossRef]
- Yang, L.; Weiss, T.M.; Lehrer, R.I.; Huang, H.W. Crystallization of antimicrobial pores in membranes: Magainin and protegrin. Biophys. J. 2000, 79, 2002–2009. [Google Scholar] [CrossRef] [PubMed]
- Last, N.B.; Schlamadinger, D.E.; Miranker, A.D. A common landscape for membrane-active peptides. Protein Sci. 2013, 22, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, O.V.; Akimov, S.A. The possibility of pore formation in lipid membranes by several molecules of amphipathic peptides. Biol. Membr. 2022, 39, 384–397. [Google Scholar] [CrossRef]
- Harroun, T.A.; Heller, W.T.; Weiss, T.M.; Yang, L.; Huang, H.W. Experimental evidence for hydrophobic matching and membrane-mediated interactions in lipid bilayers containing gramicidin. Biophys. J. 1999, 76, 937–945. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).