


The Molecular Basis for Selectivity of the Cytotoxic Response of Lung Adenocarcinoma Cells to Cold Atmospheric Plasma

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
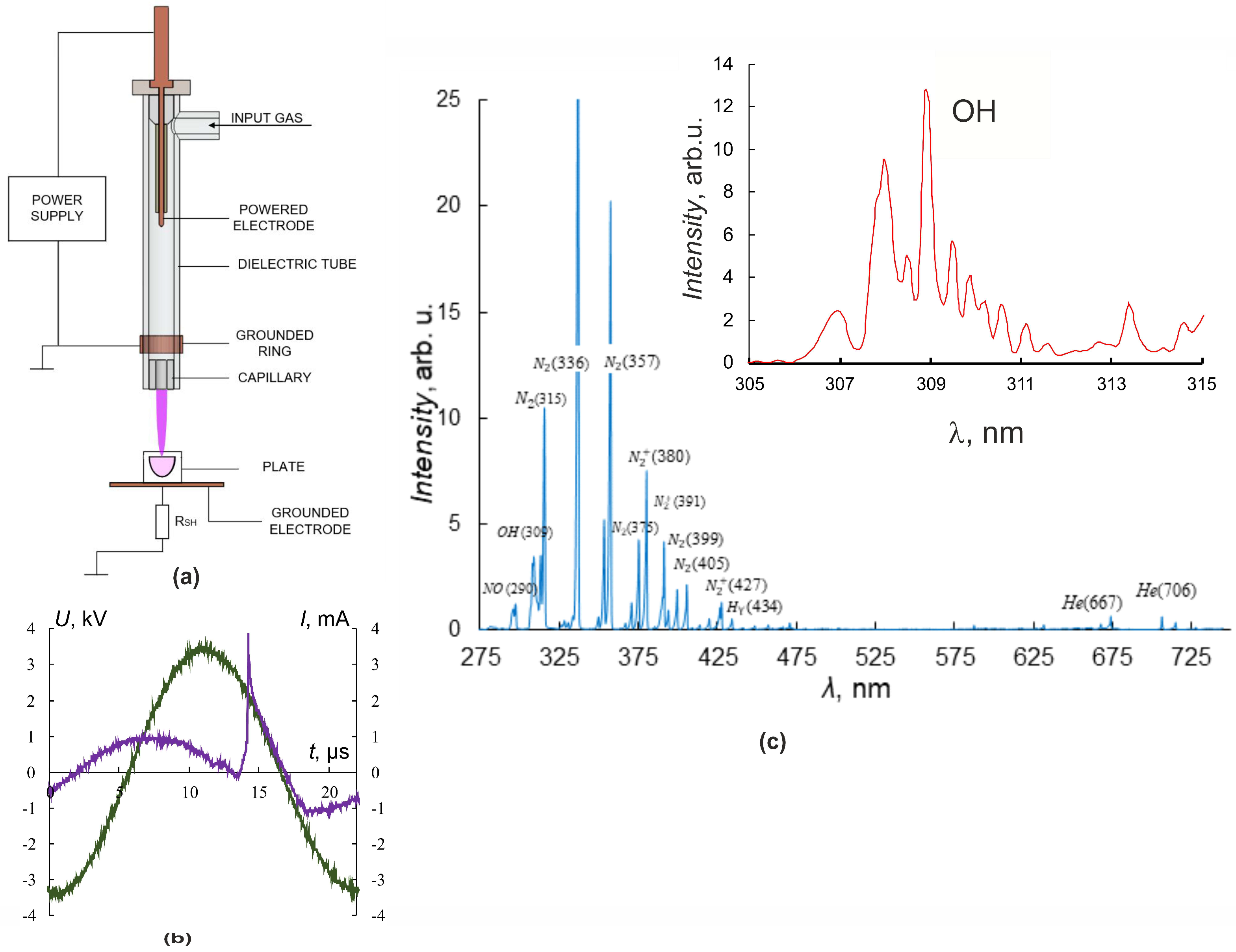
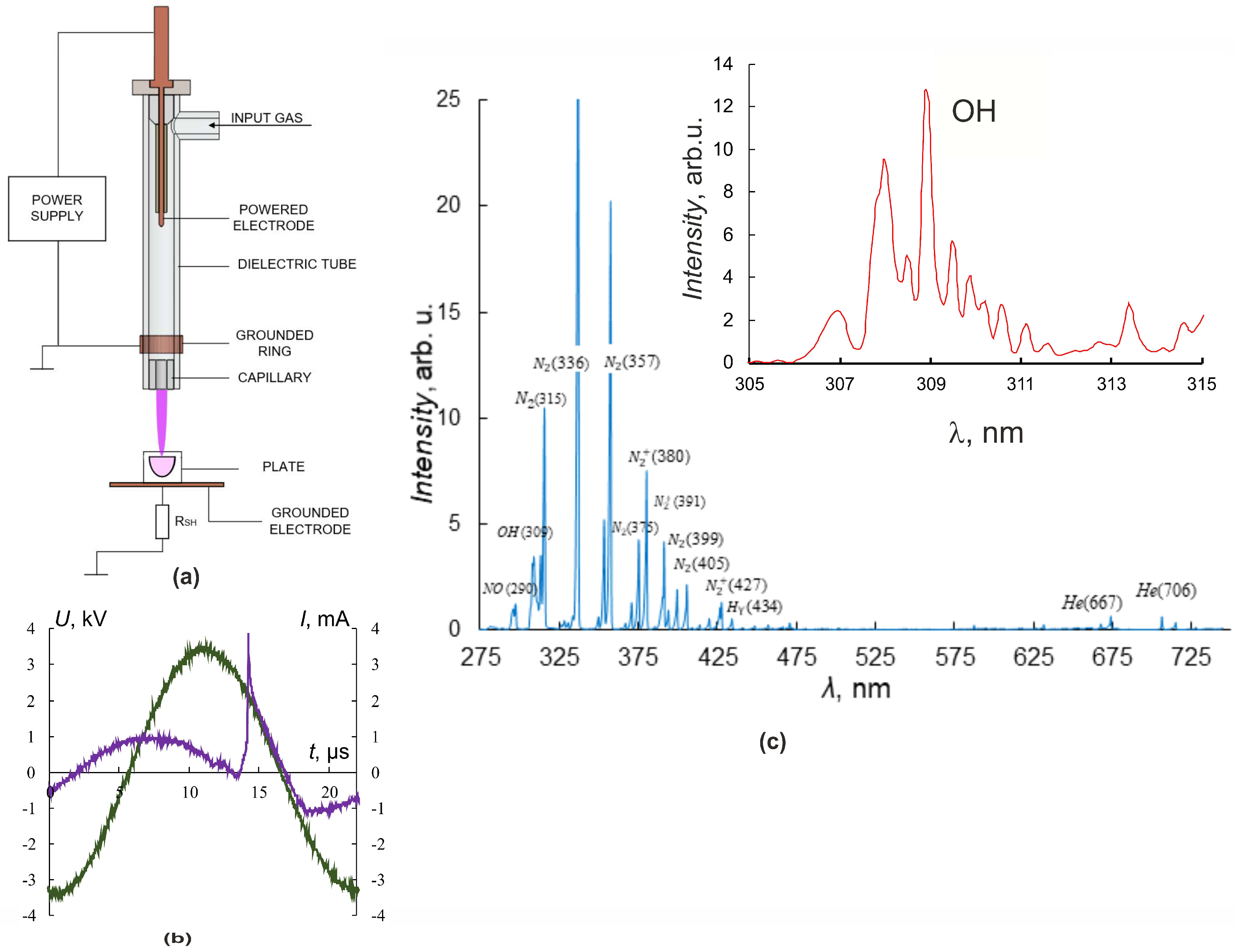
2.2. Experimental Plasma Jet Setup
2.3. Cell-Viability Assay
2.4. Western Blot
2.5. Cell-Cycle Analysis
2.6. Measurement of Extracellular Nitrite Ions
2.7. Transmission-Electron Microscopy
2.8. RNA Extraction and Real-Time Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
2.9. RNA-Seq and Transcriptome Analyses and Gene-Set-Enrichment Analysis
2.10. Statistical Analysis
3. Results
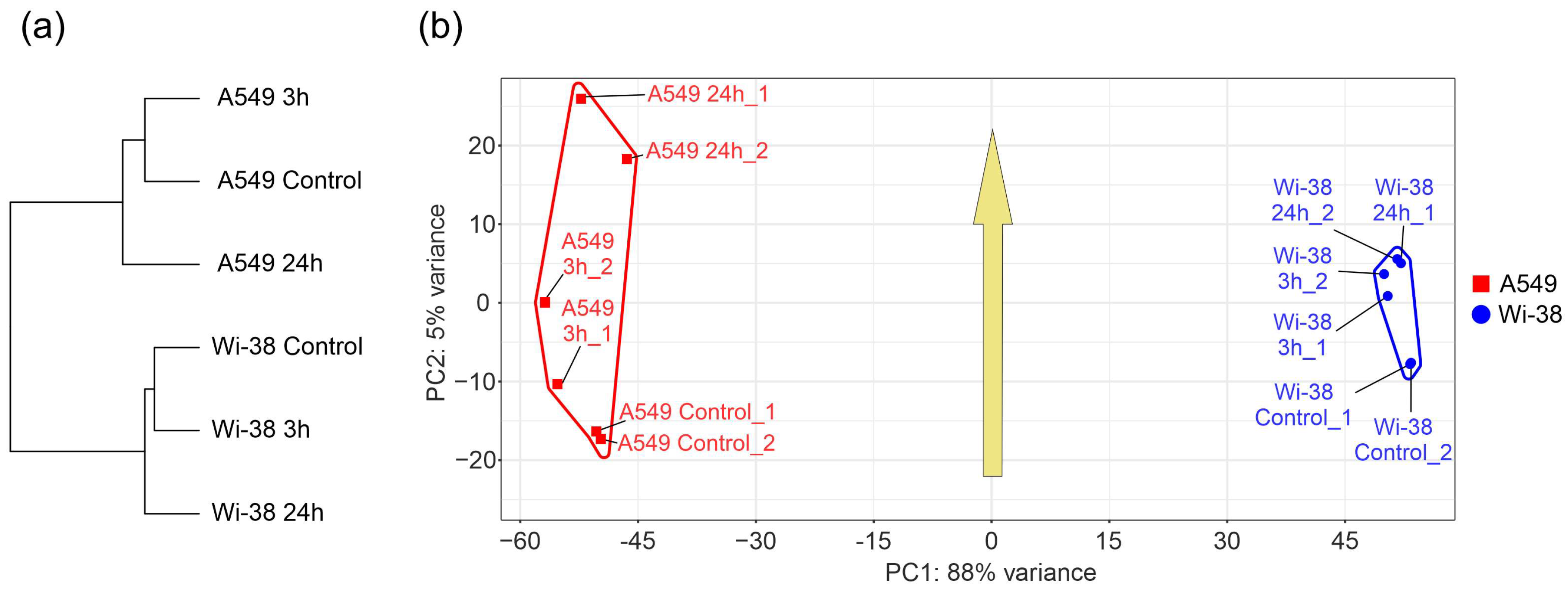
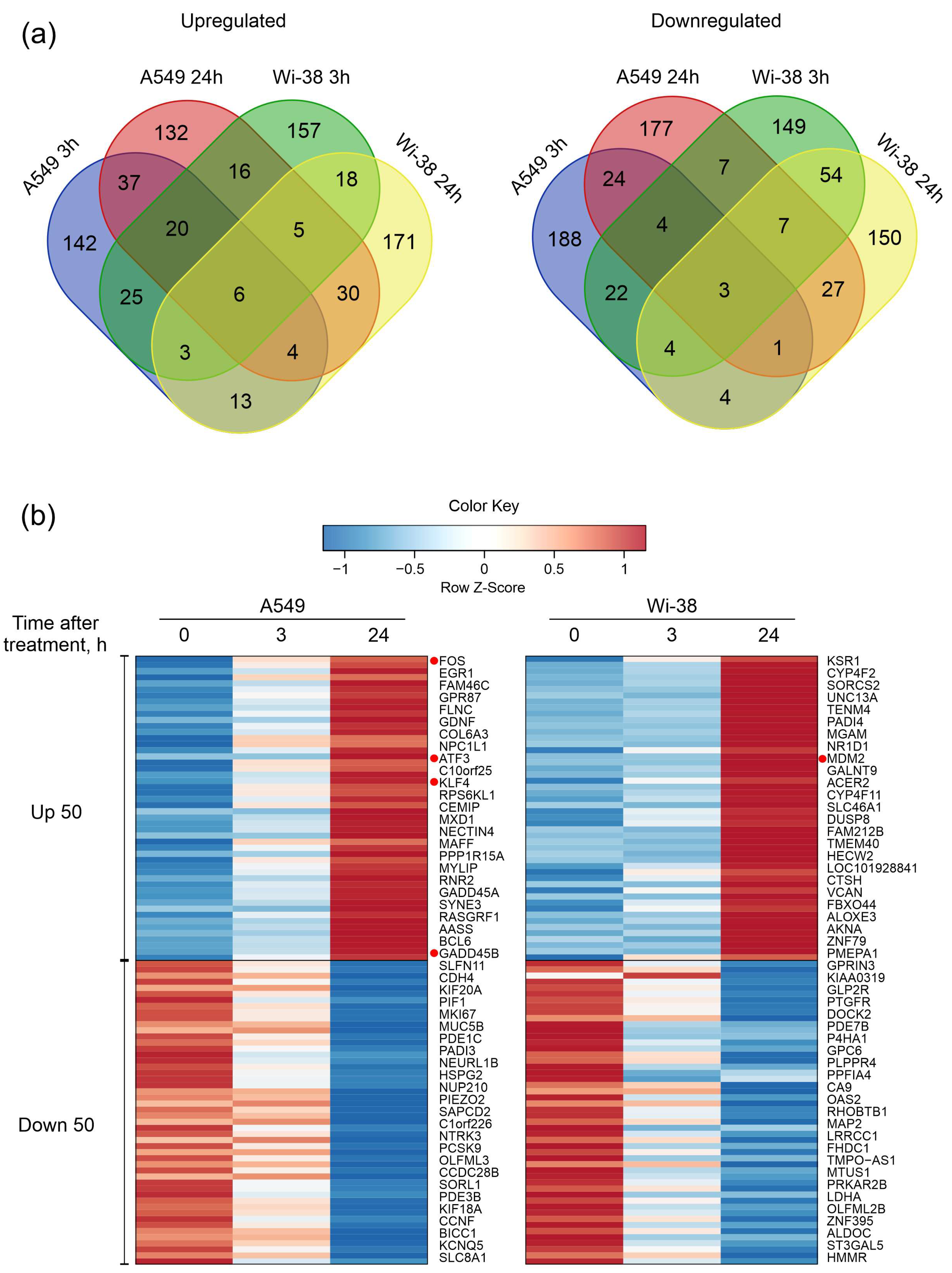
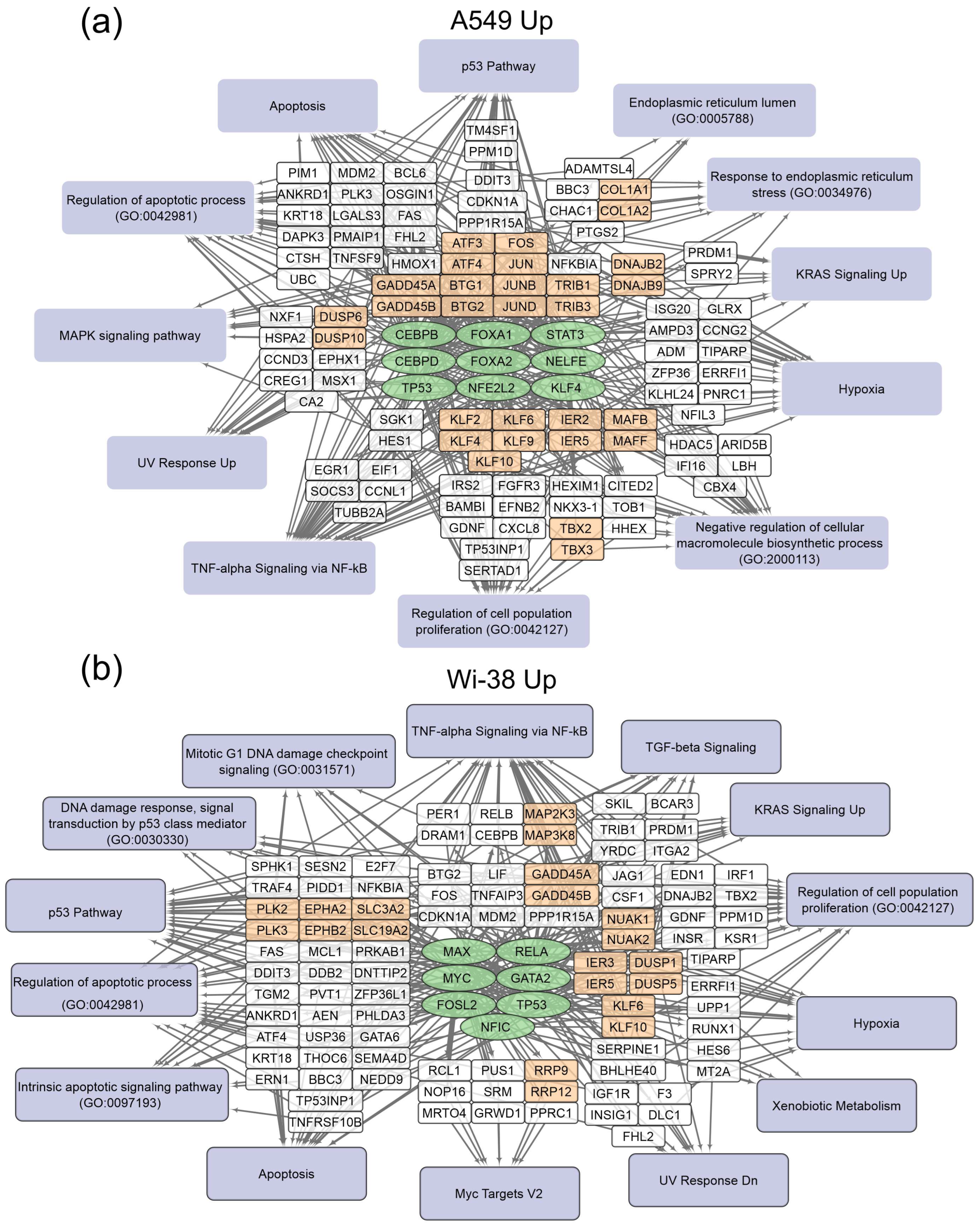
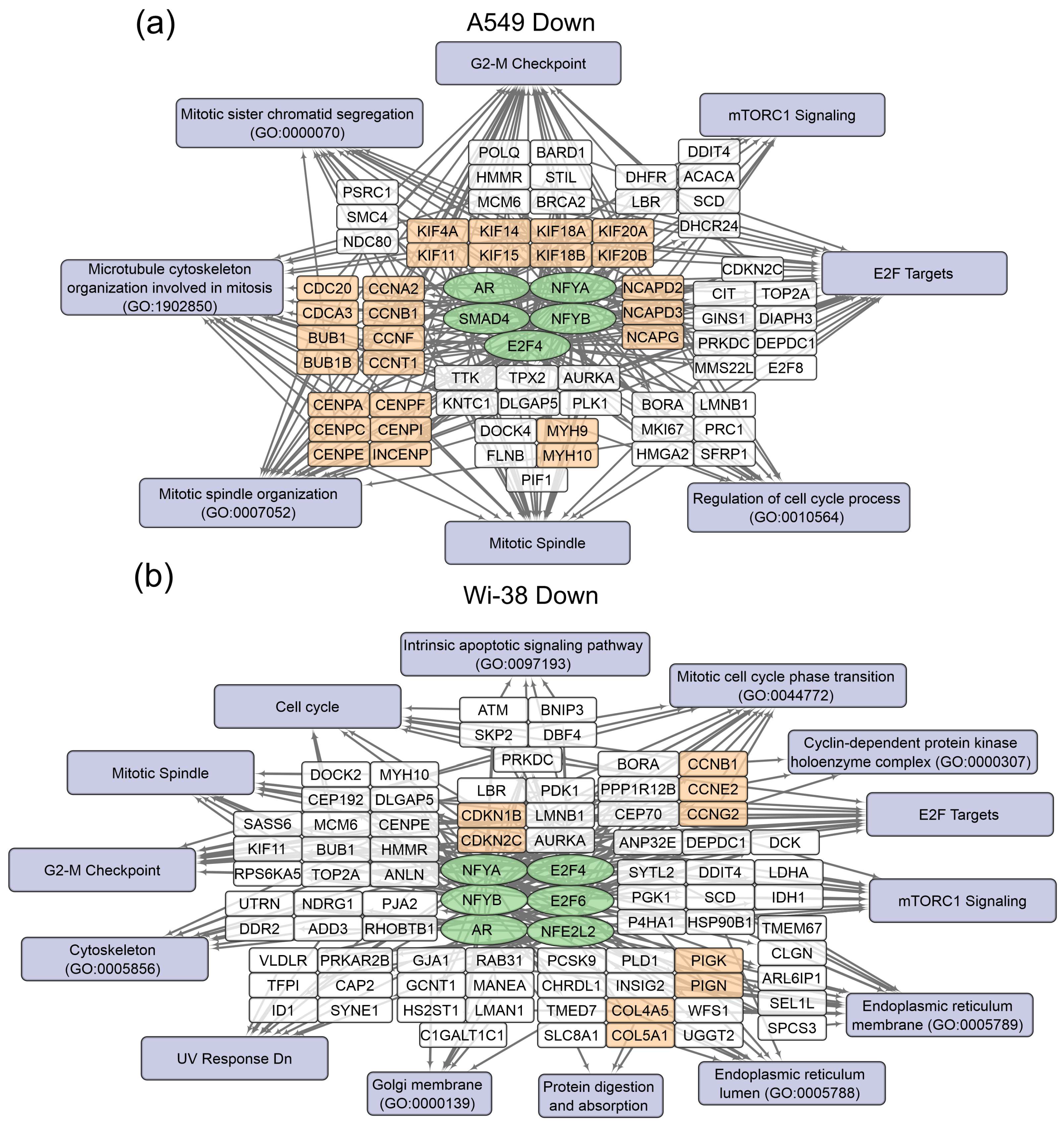
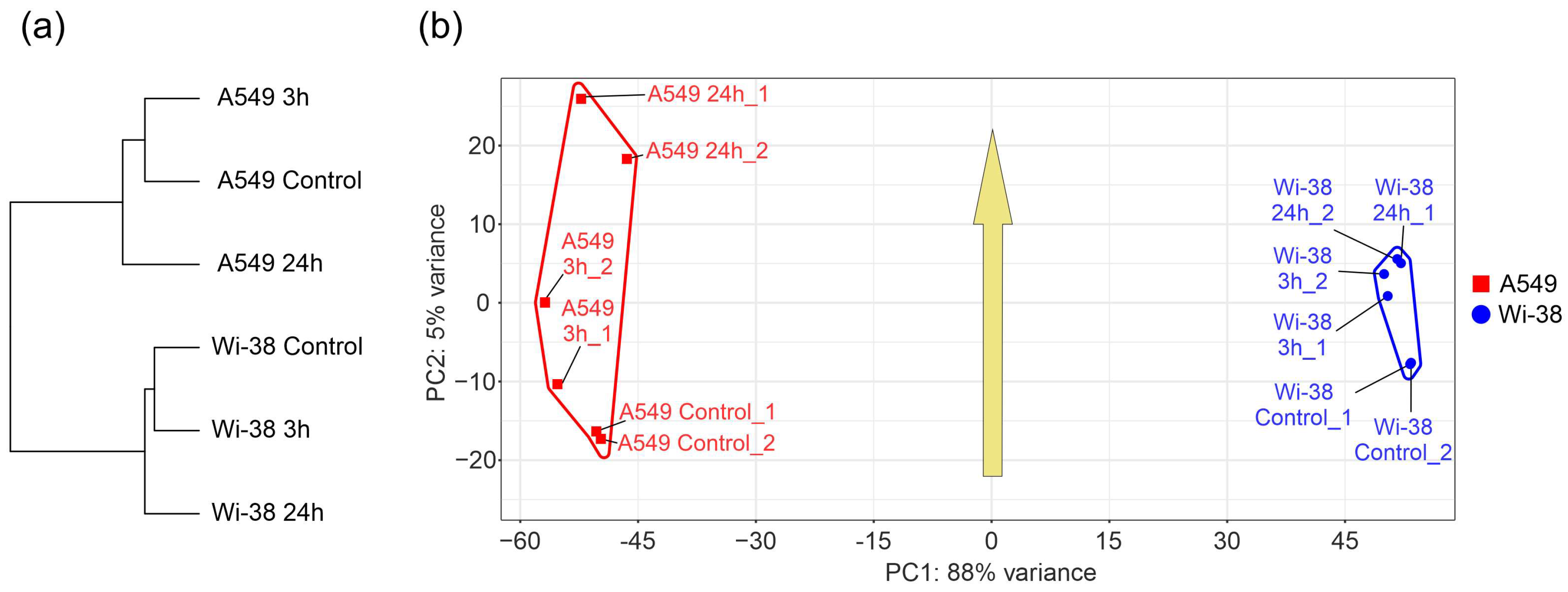
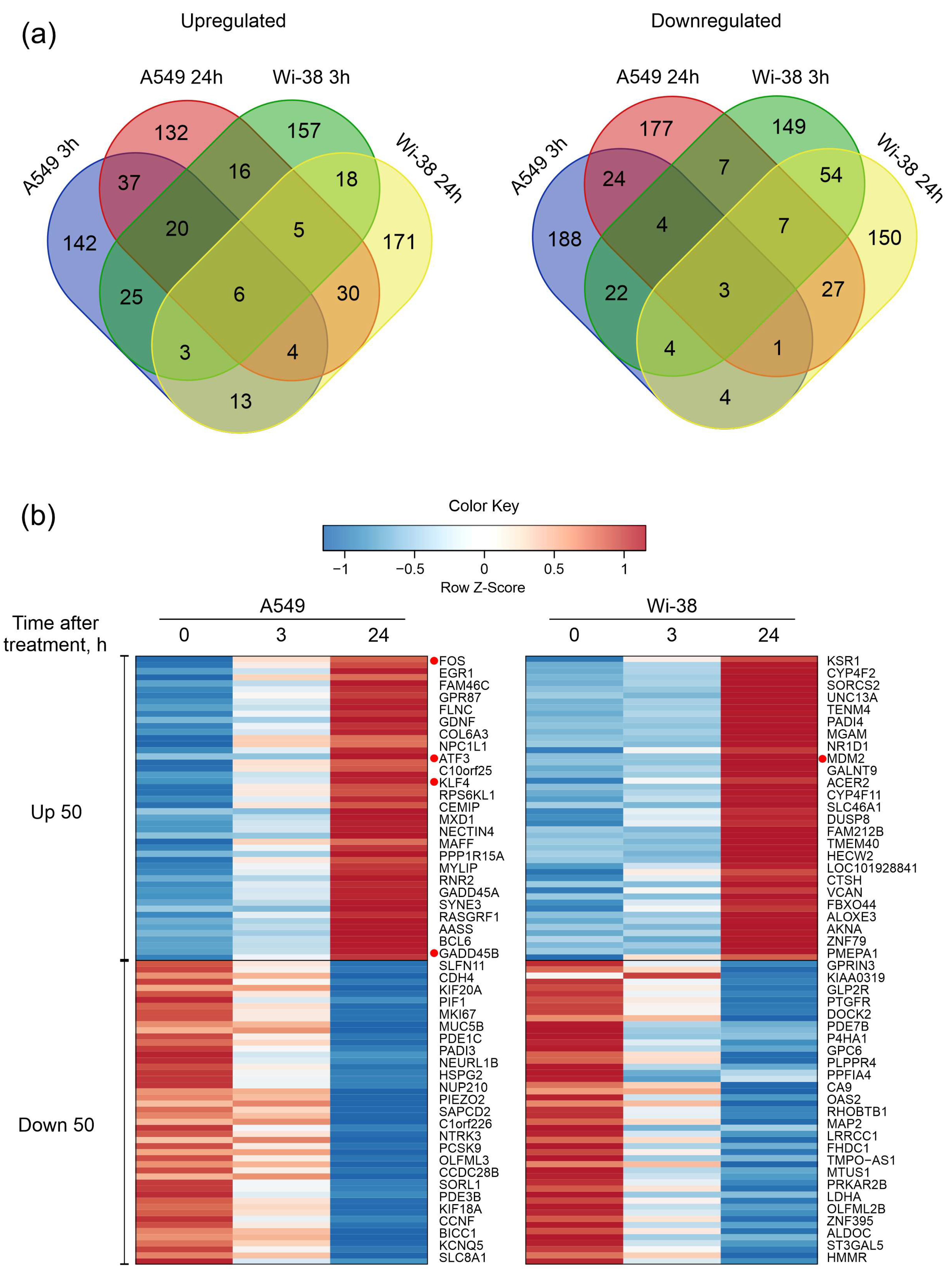
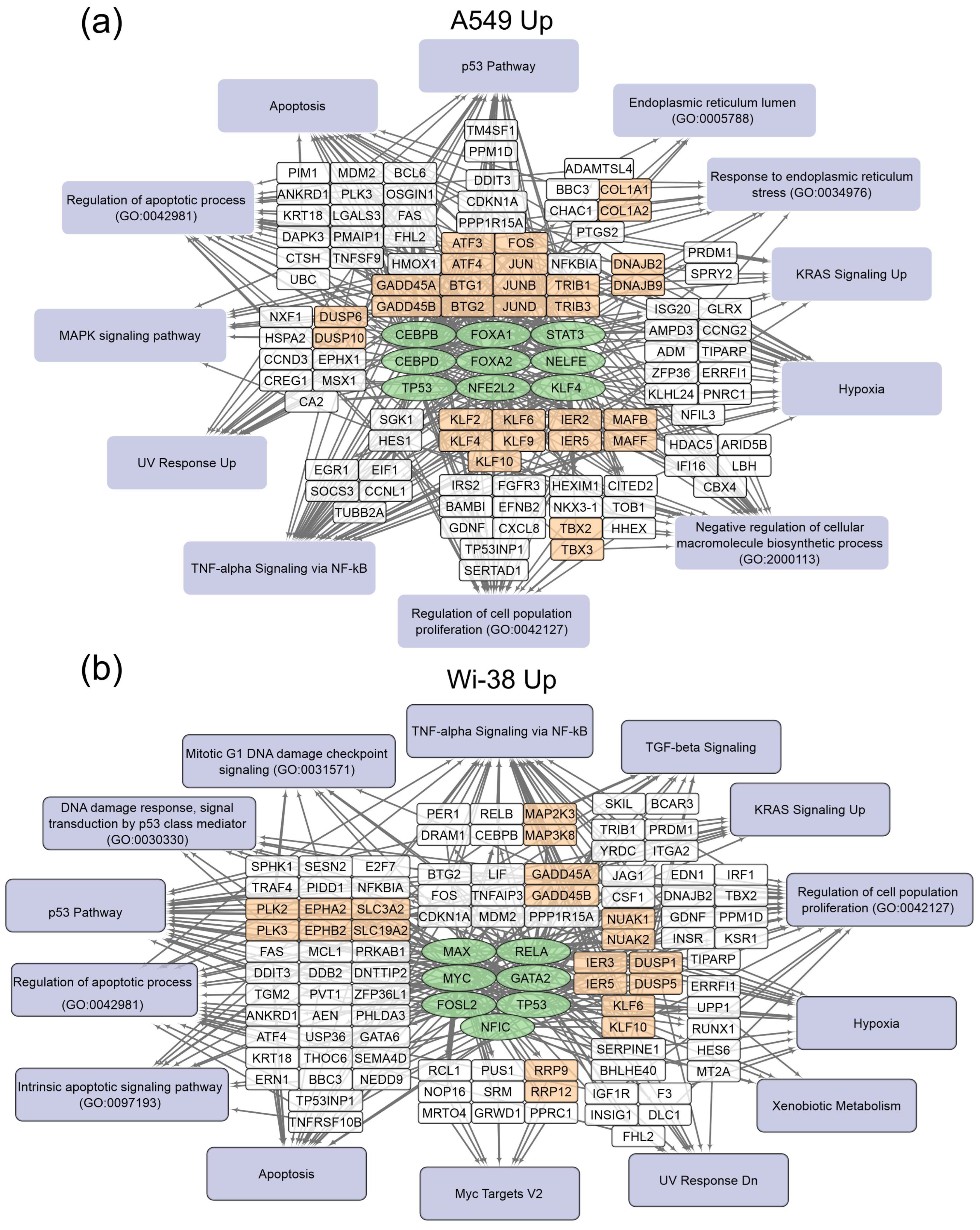
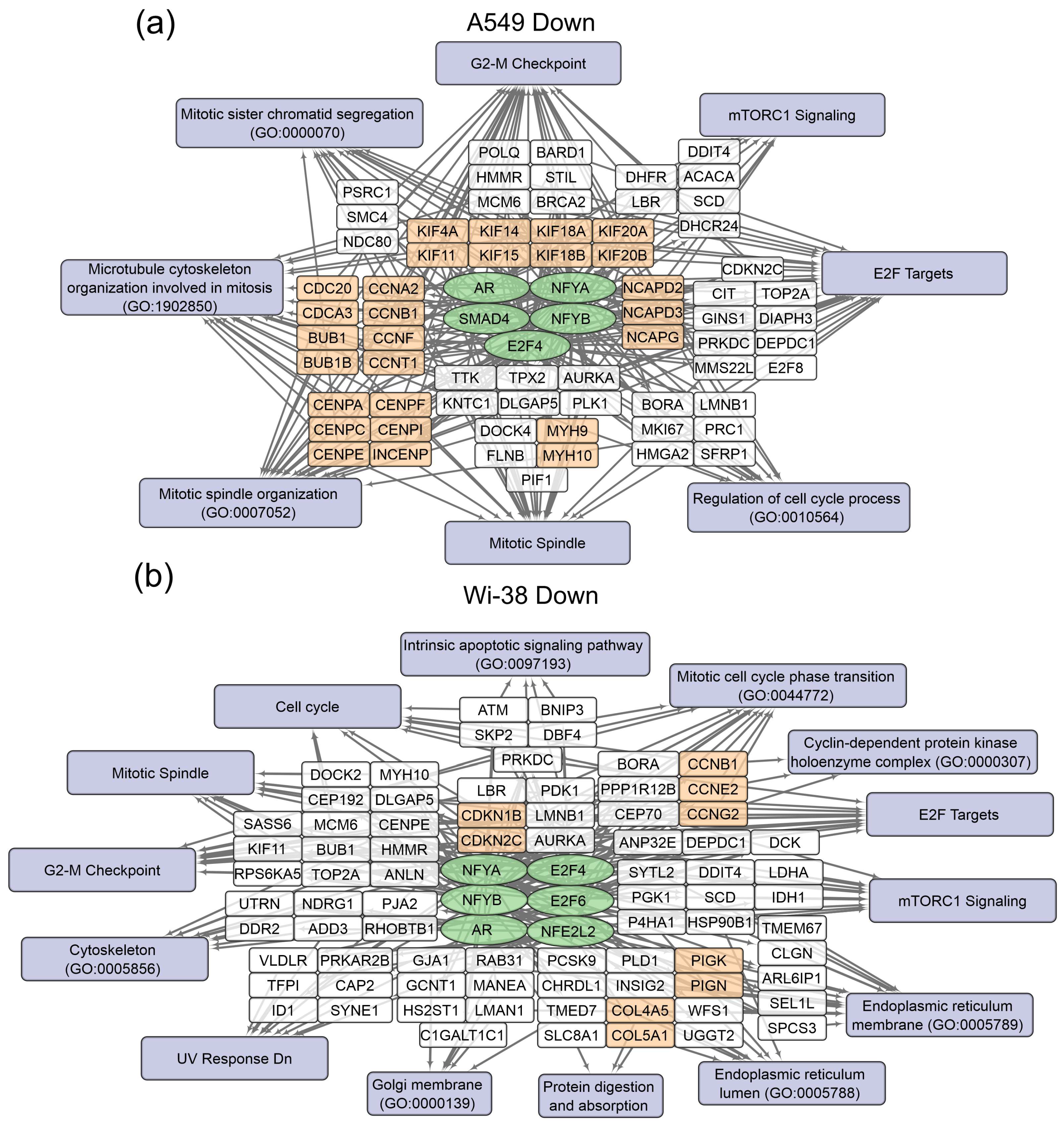
3.1. Transcriptome Analysis of A549 and Wi-38 Cells Treated with CAP
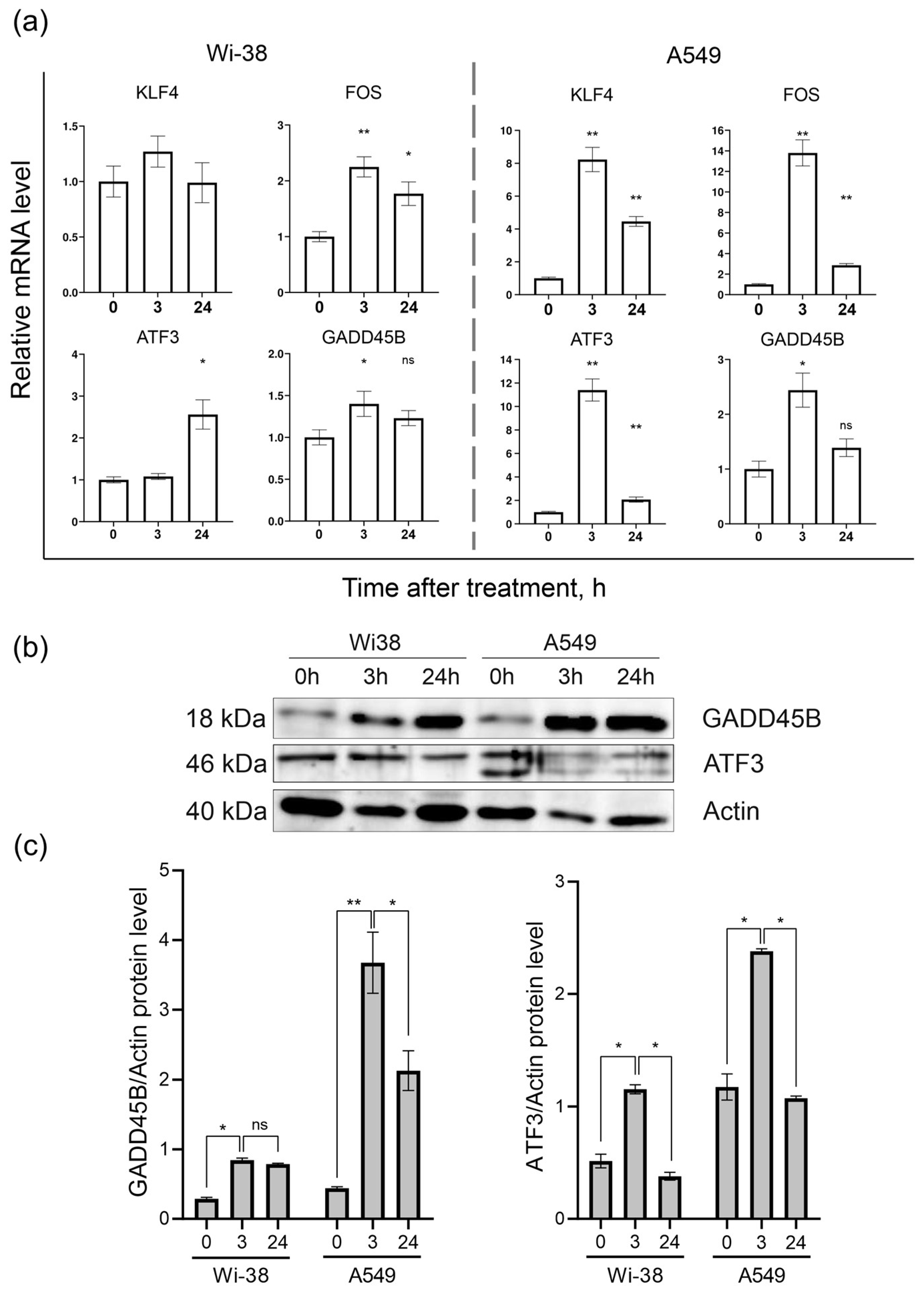
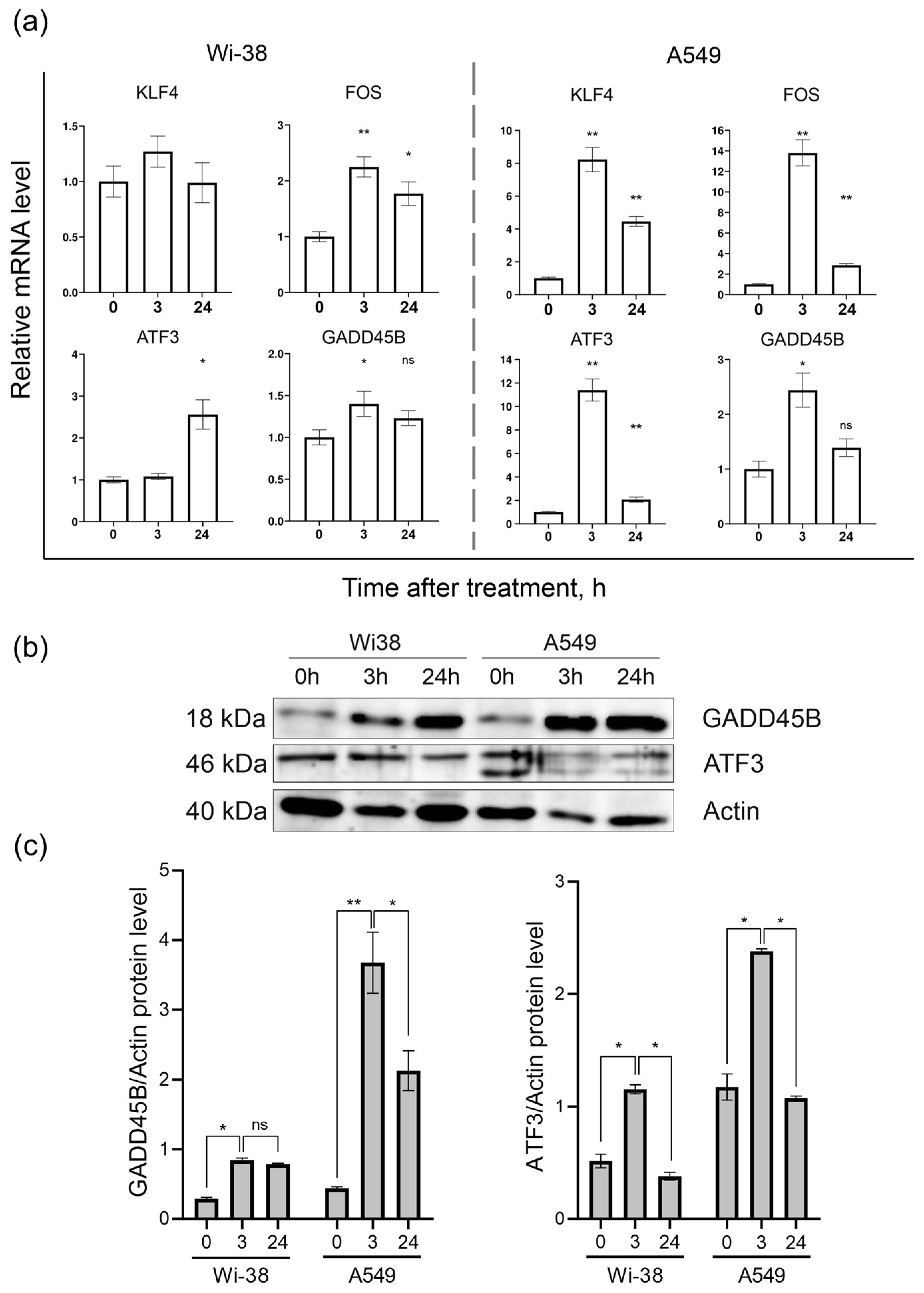
3.2. Real-Time PCR Analysis of Gene Expression and Protein Level
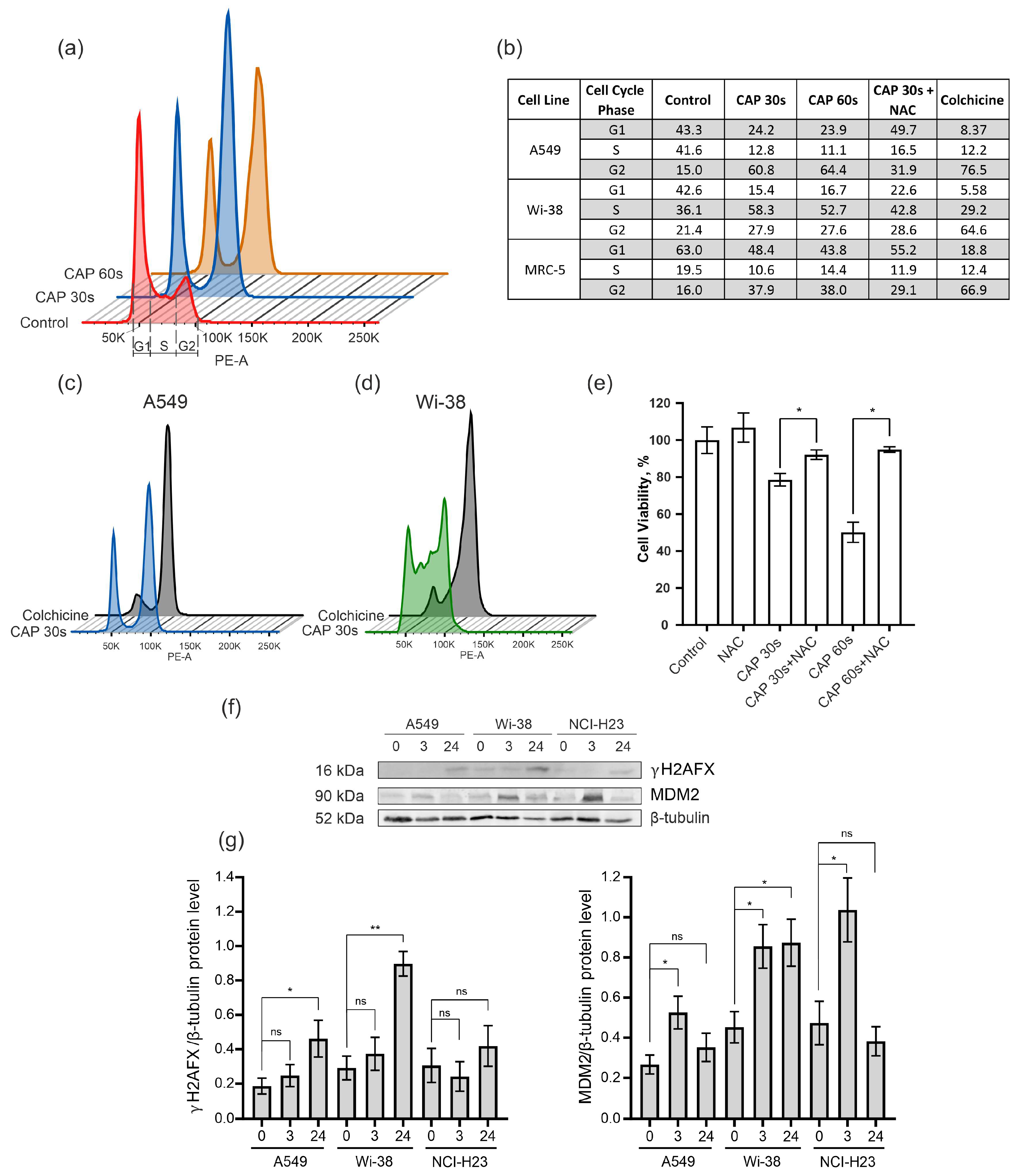
3.3. CAP Treatment Arrests the Cells in G2 Phase of Cell Cycle
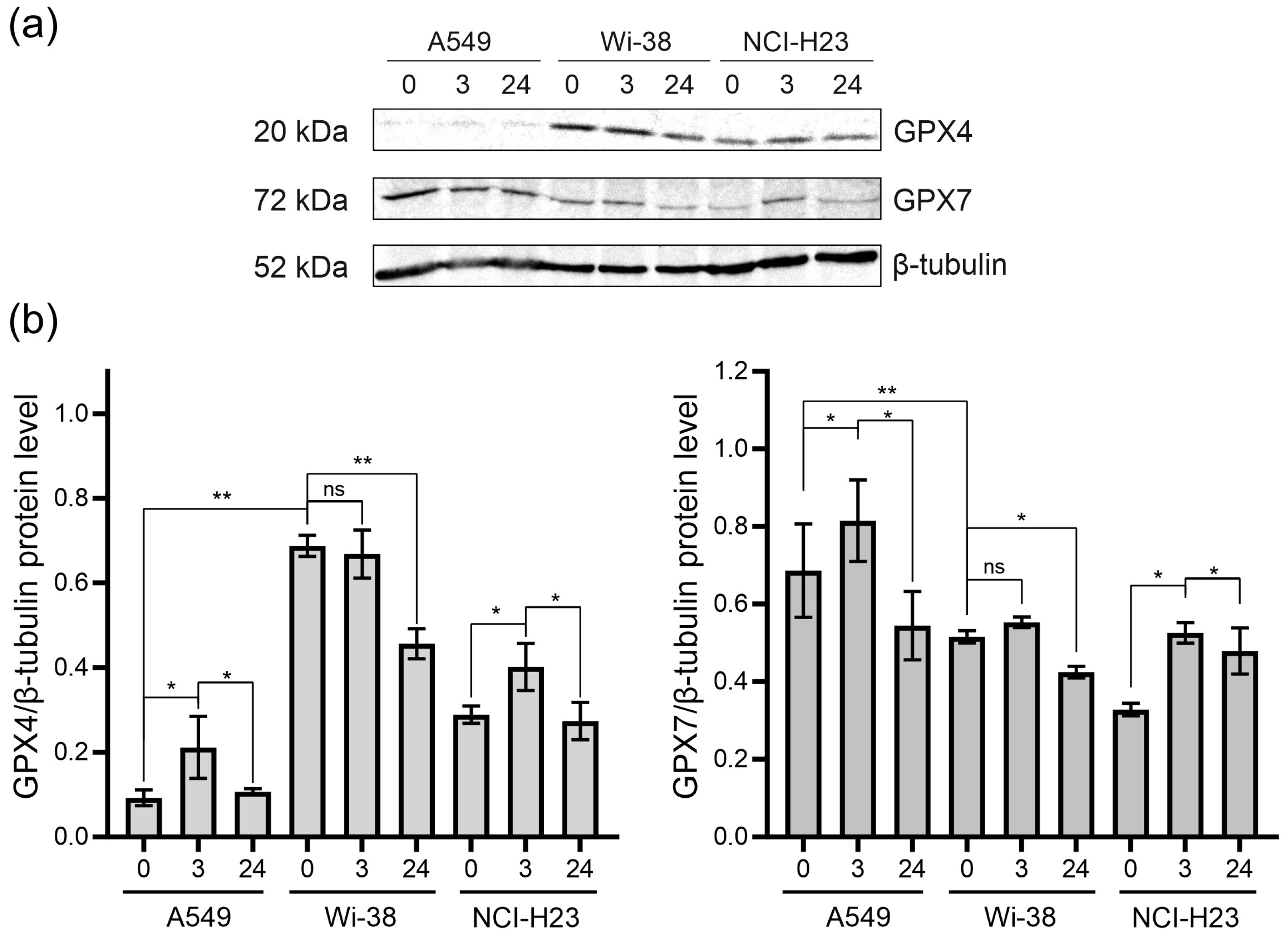
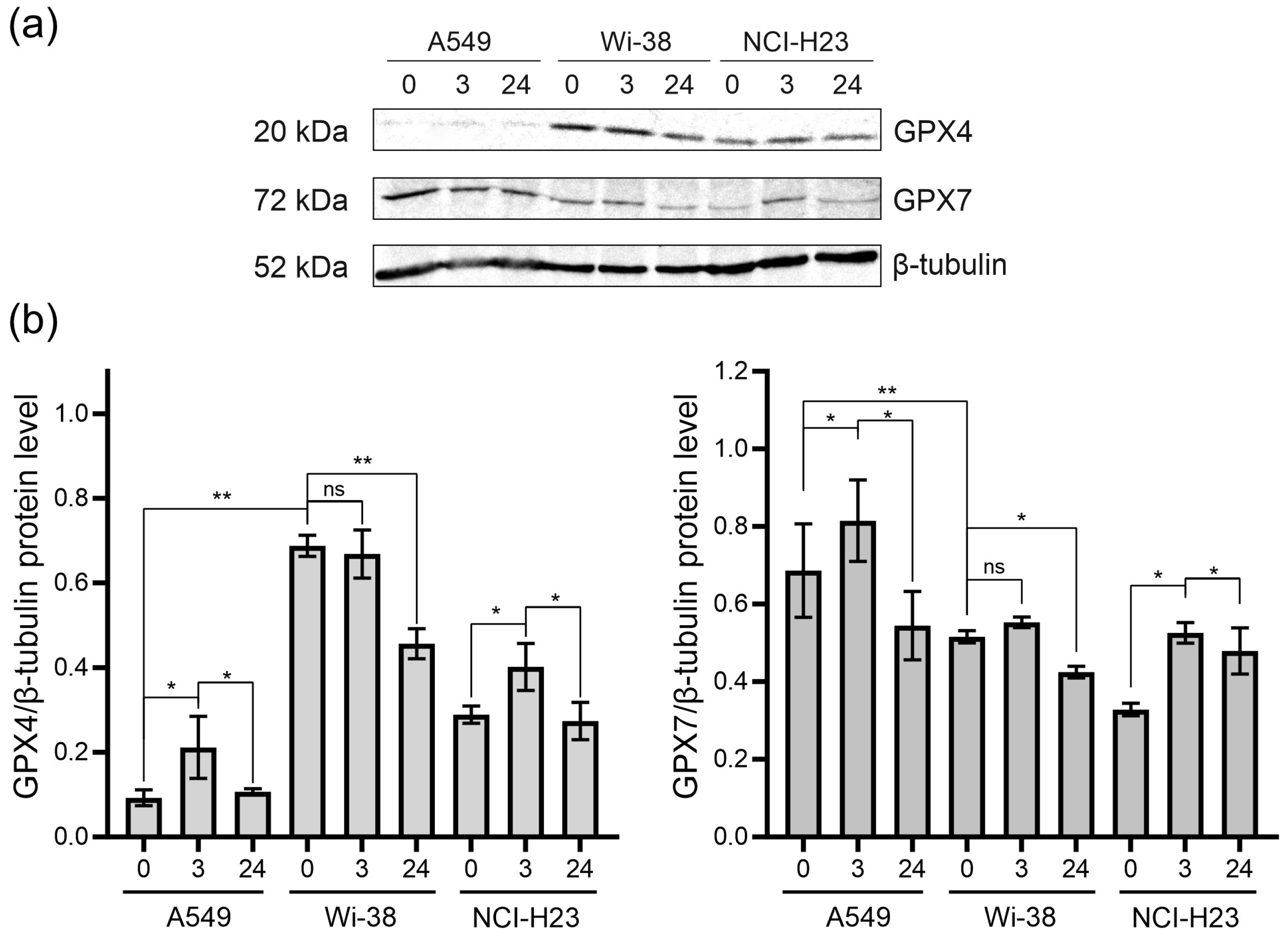
3.4. Analysis of GPX4 and GPX7 Responses to the CAP Treatment
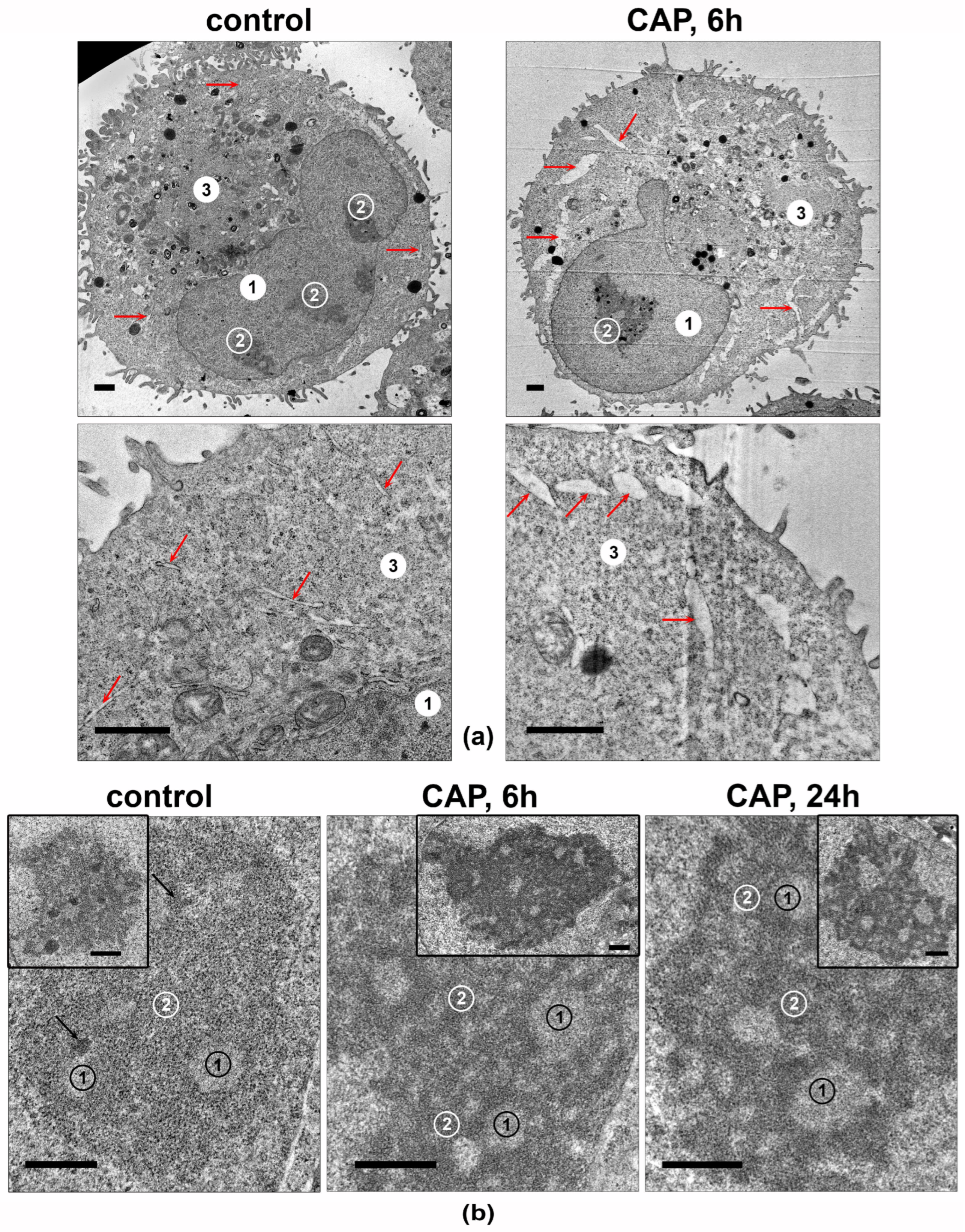
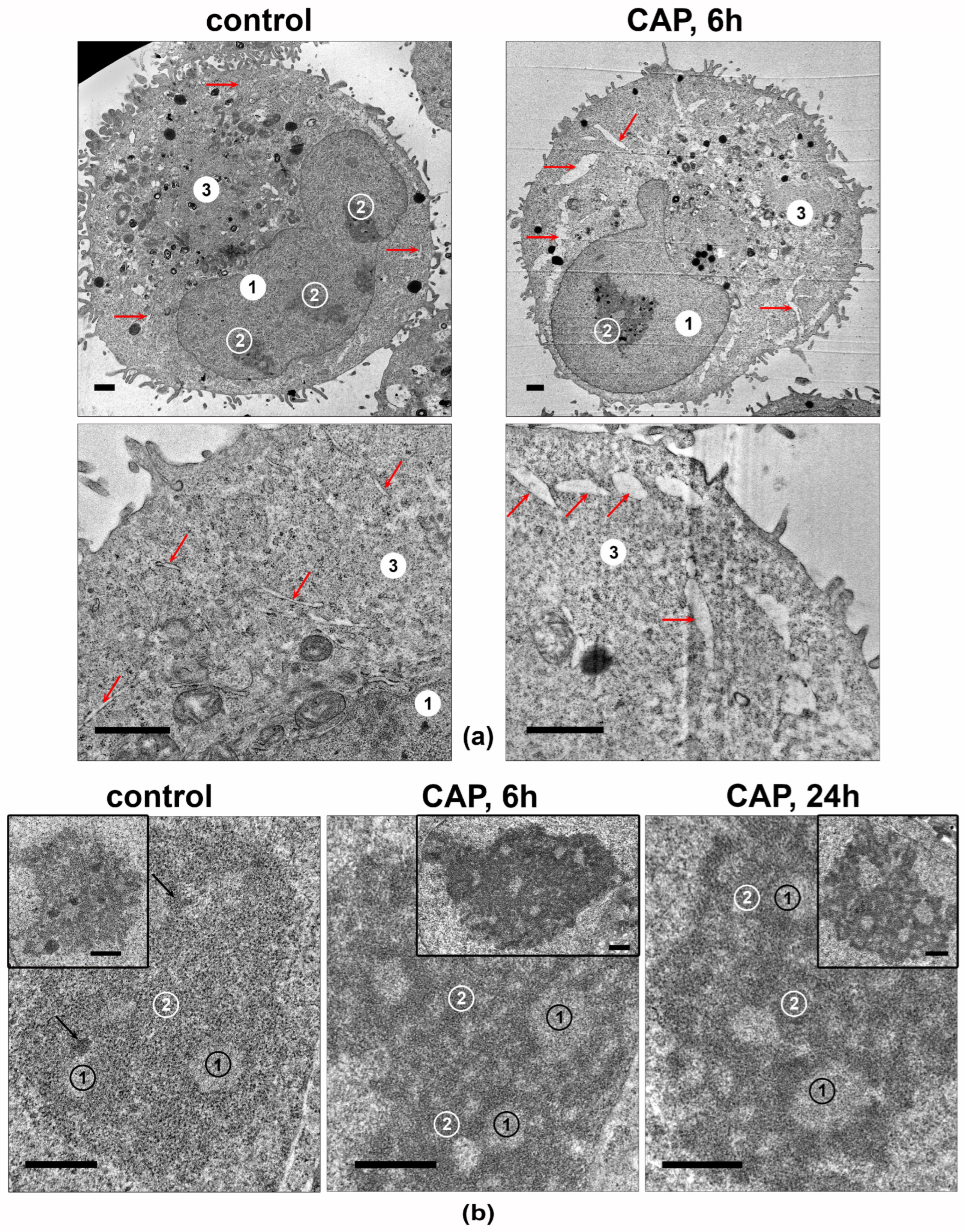
3.5. CAP Treatment Changes the Ultrastructure of Nucleolus in the A549 Cells
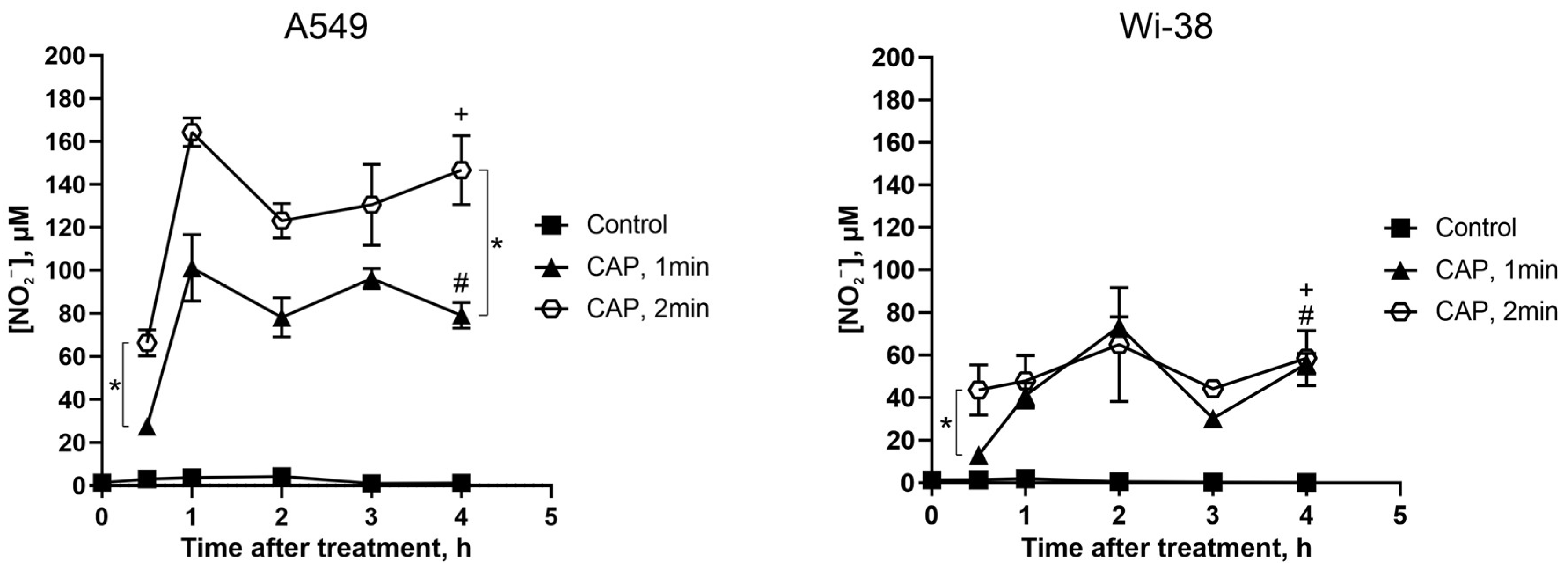
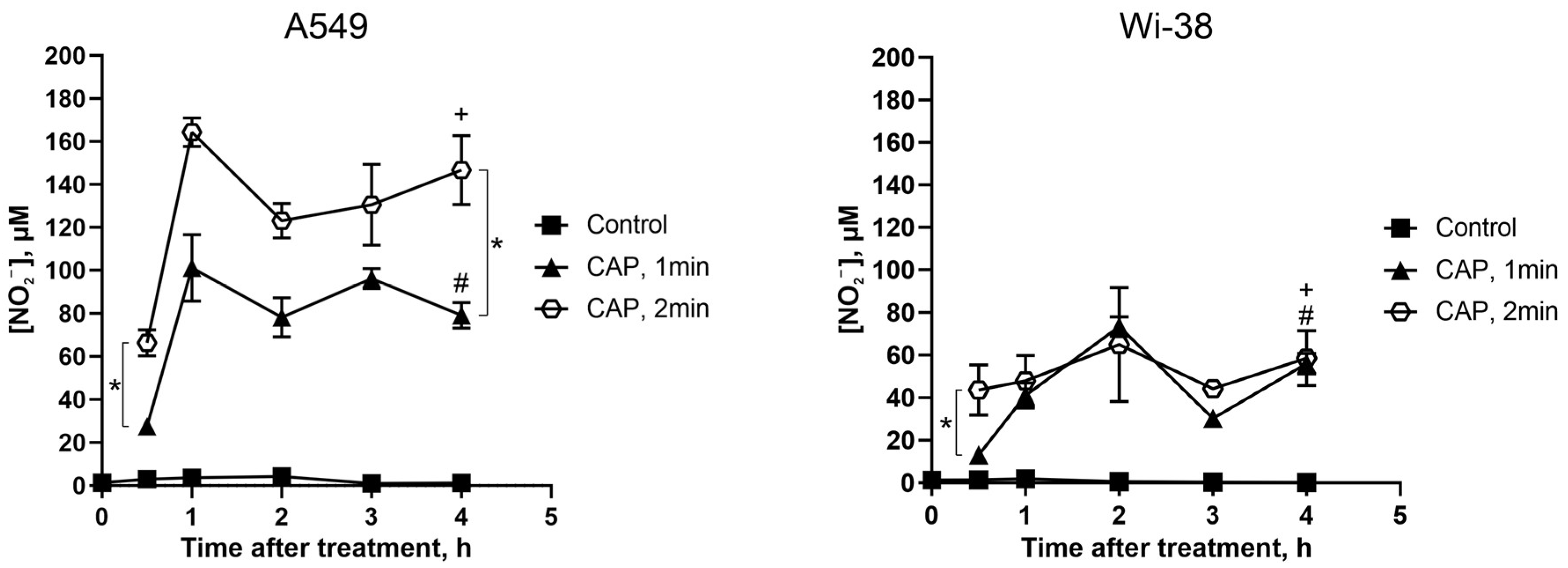
3.6. Direct CAP Exposure of the Cultivated Cells Led to Changes in the Extracellular Contents of Nitrite Ions
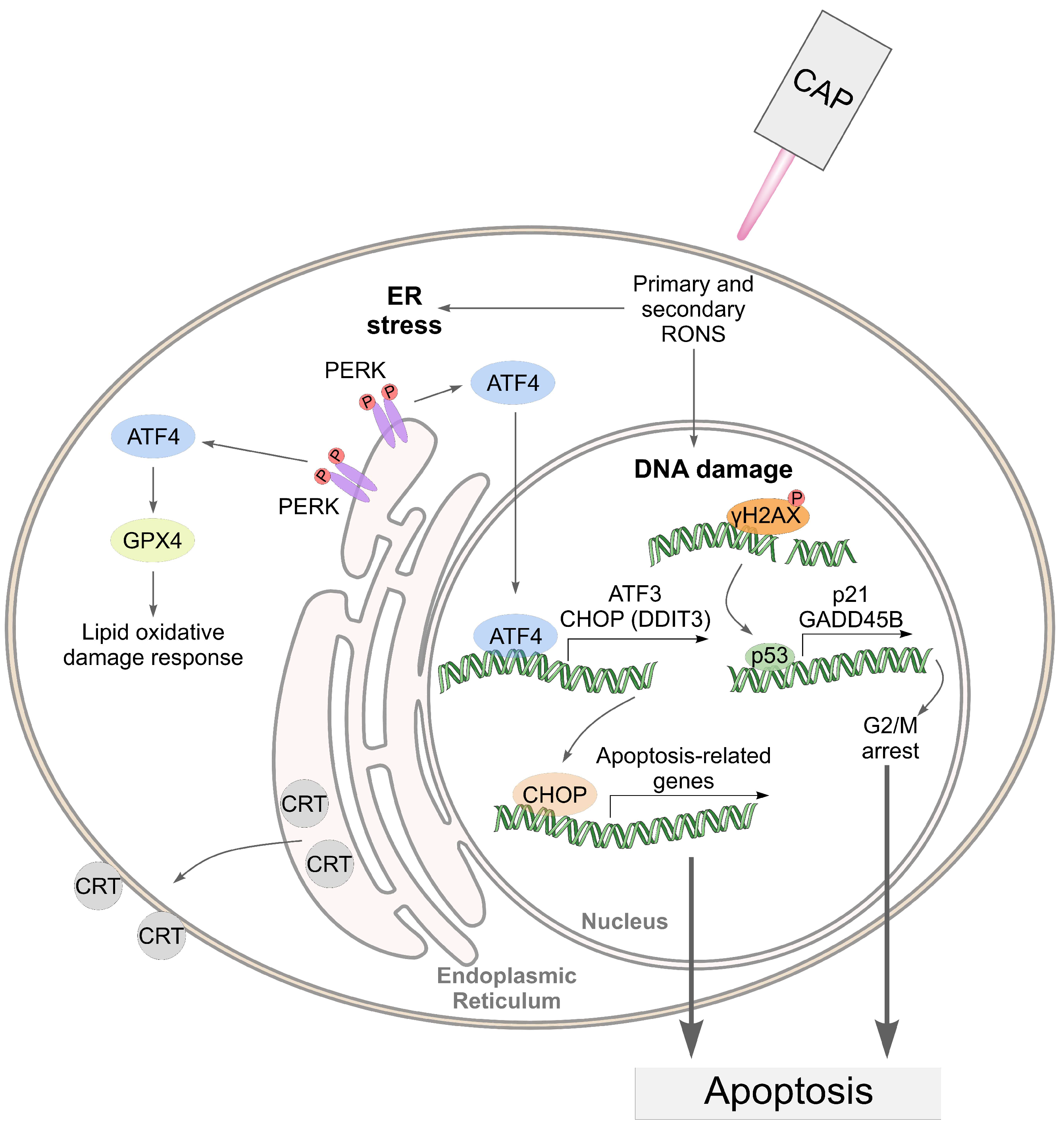
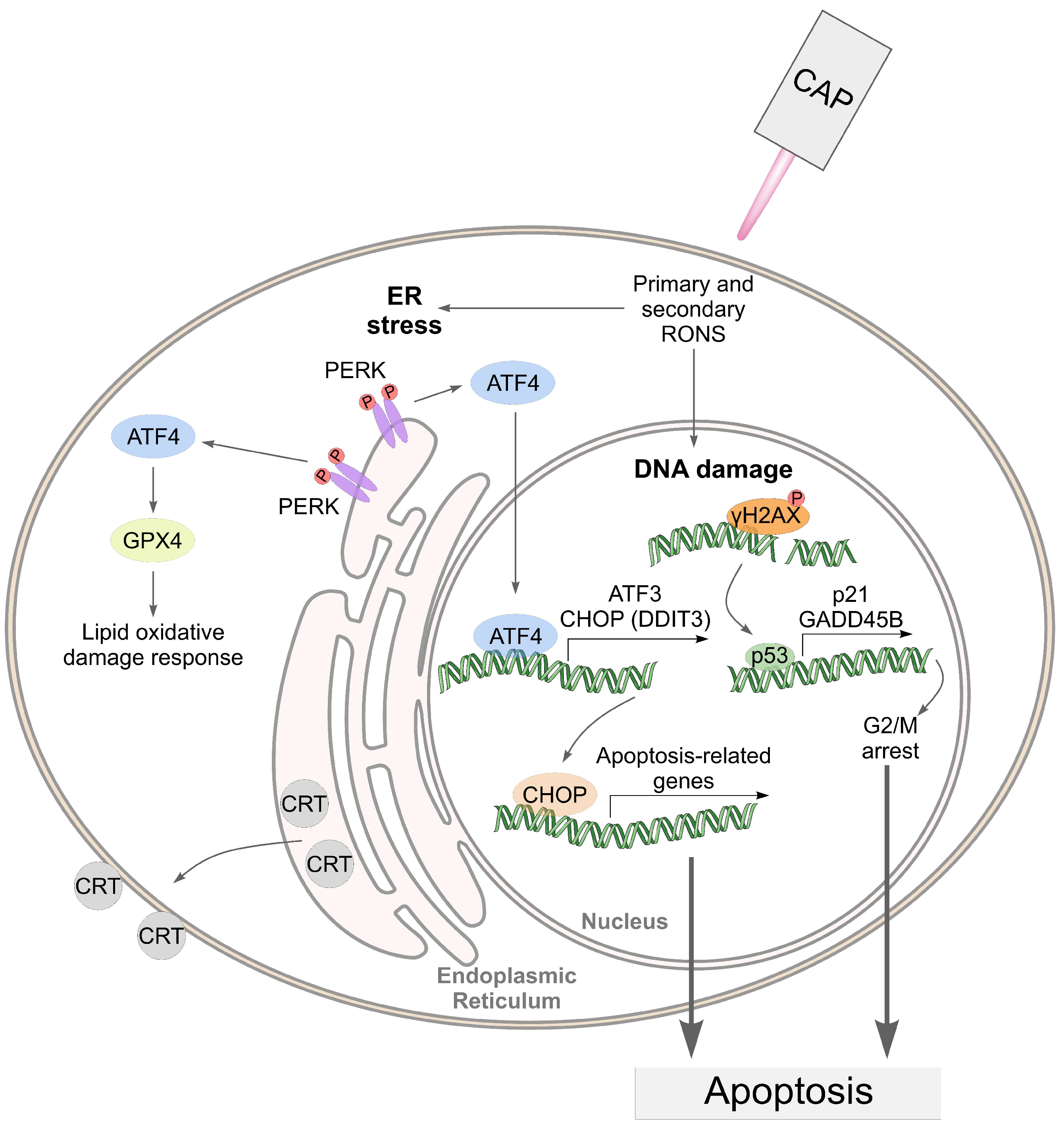
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welz, C.; Emmert, S.; Canis, M.; Becker, S.; Baumeister, P.; Shimizu, T.; Morfill, G.E.; Harréus, U.; Zimmermann, J.L. Cold Atmospheric Plasma: A Promising Complementary Therapy for Squamous Head and Neck Cancer. PLoS ONE 2015, 10, e0141827. [Google Scholar] [CrossRef] [PubMed]
- Golubitskaya, E.A.; Troitskaya, O.S.; Yelak, E.V.; Gugin, P.P.; Richter, V.A.; Schweigert, I.V.; Zakrevsky, D.E.; Koval, O.A. Cold Physical Plasma Decreases the Viability of Lung Adenocarcinoma Cells. Acta Nat. 2019, 11, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Patrakova, E.; Biryukov, M.; Troitskaya, O.; Gugin, P.; Milakhina, E.; Semenov, D.; Poletaeva, J.; Ryabchikova, E.; Novak, D.; Kryachkova, N.; et al. Chloroquine Enhances Death in Lung Adenocarcinoma A549 Cells Exposed to Cold Atmospheric Plasma Jet. Cells 2023, 12, 290. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, I.; Zakrevsky, D.; Gugin, P.; Yelak, E.; Golubitskaya, E.; Troitskaya, O.; Koval, O. Interaction of Cold Atmospheric Argon and Helium Plasma Jets with Bio-Target with Grounded Substrate Beneath. Appl. Sci. 2019, 9, 4528. [Google Scholar] [CrossRef]
- Troitskaya, O.; Golubitskaya, E.; Biryukov, M.; Varlamov, M.; Gugin, P.; Milakhina, E.; Richter, V.; Schweigert, I.; Zakrevsky, D.; Koval, O. Non-Thermal Plasma Application in Tumor-Bearing Mice Induces Increase of Serum HMGB1. Int. J. Mol. Sci. 2020, 21, 5128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yan, C.; Kong, S.; Jia, T.; Chu, Z.; Yang, L.; Wu, J.; Geng, S.; Guo, K. Biosafety and Differentially Expressed Genes Analysis of Melanoma Cells Treated with Cold Atmospheric Plasma. J. Biophotonics 2022, 15, e202100403. [Google Scholar] [CrossRef]
- Yan, D.; Xu, W.; Yao, X.; Lin, L.; Sherman, J.H.; Keidar, M. The Cell Activation Phenomena in the Cold Atmospheric Plasma Cancer Treatment. Sci. Rep. 2018, 8, 15418. [Google Scholar] [CrossRef]
- Li, W.; Yu, H.; Ding, D.; Chen, Z.; Wang, Y.; Wang, S.; Li, X.; Keidar, M.; Zhang, W. Cold Atmospheric Plasma and Iron Oxide-Based Magnetic Nanoparticles for Synergetic Lung Cancer Therapy. Free Radic. Biol. Med. 2019, 130, 71–81. [Google Scholar] [CrossRef]
- Chen, Z.; Simonyan, H.; Cheng, X.; Gjika, E.; Lin, L.; Canady, J.; Sherman, J.; Young, C.; Keidar, M. A Novel Micro Cold Atmospheric Plasma Device for Glioblastoma Both In Vitro and In Vivo. Cancers 2017, 9, 61. [Google Scholar] [CrossRef]
- Patrakova, E.A.; Biryukov, M.M.; Troitskaya, O.S.; Novak, D.D.; Milakhina, E.V.; Gugin, P.P.; Zakrevsky, D.E.; Schweigert, I.V.; Koval, O.A. Cytotoxic Activity of a Cold Atmospheric Plasma Jet in Relation to a 3D Cell Model of Human Breast Cancer. Cell Tissue Biol. 2023, 17, 233–246. [Google Scholar] [CrossRef]
- Privat-Maldonado, A.; Gorbanev, Y.; Dewilde, S.; Smits, E.; Bogaerts, A. Reduction of Human Glioblastoma Spheroids Using Cold Atmospheric Plasma: The Combined Effect of Short- and Long-Lived Reactive Species. Cancers 2018, 10, 394. [Google Scholar] [CrossRef]
- Laroussi, M.; Lu, X.; Keidar, M. Perspective: The Physics, Diagnostics, and Applications of Atmospheric Pressure Low Temperature Plasma Sources Used in Plasma Medicine. J. Appl. Phys. 2017, 122, 020901. [Google Scholar] [CrossRef]
- Canady, J.; Murthy, S.R.K.; Zhuang, T.; Gitelis, S.; Nissan, A.; Ly, L.; Jones, O.Z.; Cheng, X.; Adileh, M.; Blank, A.T.; et al. The First Cold Atmospheric Plasma Phase I Clinical Trial for the Treatment of Advanced Solid Tumors: A Novel Treatment Arm for Cancer. Cancers 2023, 15, 3688. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, I.V.; Zakrevsky, D.E.; Gugin, P.P.; Milakhina, E.V.; Biryukov, M.M.; Keidar, M.; Koval, O.A. Effect of Voltage Pulse Duration on Electrophysical and Thermal Characteristics of Cold Atmospheric Plasma Jet. Plasma Sources Sci. Technol. 2022, 31, 114004. [Google Scholar] [CrossRef]
- Graves, D.B. The Emerging Role of Reactive Oxygen and Nitrogen Species in Redox Biology and Some Implications for Plasma Applications to Medicine and Biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar] [CrossRef]
- Mitra, S.; Nguyen, L.N.; Akter, M.; Park, G.; Choi, E.H.; Kaushik, N.K. Impact of ROS Generated by Chemical, Physical, and Plasma Techniques on Cancer Attenuation. Cancers 2019, 11, 1030. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive Oxygen Species: Drivers of Physiological and Pathological Processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef]
- Von Woedtke, T.; Schmidt, A.; Bekeschus, S.; Wende, K.; Weltmann, K.-D. Plasma Medicine: A Field of Applied Redox Biology. In Vivo 2019, 33, 1011–1026. [Google Scholar] [CrossRef]
- Semmler, M.L.; Bekeschus, S.; Schäfer, M.; Bernhardt, T.; Fischer, T.; Witzke, K.; Seebauer, C.; Rebl, H.; Grambow, E.; Vollmar, B.; et al. Molecular Mechanisms of the Efficacy of Cold Atmospheric Pressure Plasma (CAP) in Cancer Treatment. Cancers 2020, 12, 269. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Starkova, T.Y.; Kuzmin, A.A.; Tomilin, A.N. Physiological Signaling Functions of Reactive Oxygen Species in Stem Cells: From Flies to Man. Front. Cell Dev. Biol. 2021, 9, 714370. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, Oxidative Stress and the Biology of Ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative Stress in Cancer Cell Metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Flohé, L. Regulatory Phenomena in the Glutathione Peroxidase Superfamily. Antioxid. Redox Signal. 2020, 33, 498–516. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Sahun, M.; Biscop, E.; Verswyvel, H.; De Waele, J.; De Backer, J.; Theys, C.; Cuypers, B.; Laukens, K.; Berghe, W.V.; et al. Acquired Non-Thermal Plasma Resistance Mediates a Shift towards Aerobic Glycolysis and Ferroptotic Cell Death in Melanoma. Drug Resist. Updates 2023, 67, 100914. [Google Scholar] [CrossRef]
- Lertpatipanpong, P.; Sillapachaiyaporn, C.; Oh, G.; Kang, Y.-H.; Hwang, C.-Y.; Baek, S.J. Effect of Cold Atmospheric Microwave Plasma (CAMP) on Wound Healing in Canine Keratinocytes. Front. Cell Dev. Biol. 2023, 11, 1105692. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Yu, L.; Zou, F.; Hu, H.; Liu, K.; Lin, Z. Gene Expression Profiling and Functional Analysis Reveals That P53 Pathway-Related Gene Expression Is Highly Activated in Cancer Cells Treated by Cold Atmospheric Plasma-Activated Medium. PeerJ 2017, 5, e3751. [Google Scholar] [CrossRef]
- Schweigert, I.V.; Alexandrov, A.L.; Zakrevsky, D.E. Self-Organization of Touching-Target Current with Ac Voltage in Atmospheric Pressure Plasma Jet for Medical Application Parameters. Plasma Sources Sci. Technol. 2020, 29, 12LT02. [Google Scholar] [CrossRef]
- Schweigert, I.; Zakrevsky, D.; Milakhina, E.; Gugin, P.; Biryukov, M.; Patrakova, E.; Koval, O. A Grounded Electrode beneath Dielectric Targets, Including Cancer Cells, Enhances the Impact of Cold Atmospheric Plasma Jet. Plasma Phys. Control. Fusion. 2022, 64, 044015. [Google Scholar] [CrossRef]
- Zarubin, I.A.; Labusov, V.A.; Babin, S.A. Characteristics of Compact Spectrometers with Diffraction Gratings of Different Typ. Zavod. Lab. Diagn. Mater. 2019, 85, 117–121. [Google Scholar] [CrossRef]
- Koval, O.; Kochneva, G.; Tkachenko, A.; Troitskaya, O.; Sivolobova, G.; Grazhdantseva, A.; Nushtaeva, A.; Kuligina, E.; Richter, V. Recombinant Vaccinia Viruses Coding Transgenes of Apoptosis-Inducing Proteins Enhance Apoptosis But Not Immunogenicity of Infected Tumor Cells. BioMed Res. Int. 2017, 2017, 3620510. [Google Scholar] [CrossRef] [PubMed]
- Bagamanshina, A.V.; Troitskaya, O.S.; Nushtaeva, A.A.; Yunusova, A.Y.; Starykovych, M.O.; Kuligina, E.V.; Kit, Y.Y.; Richter, M.; Wohlfromm, F.; Kähne, T.; et al. Cytotoxic and Antitumor Activity of Lactaptin in Combination with Autophagy Inducers and Inhibitors. BioMed Res. Int. 2019, 2019, 4087160. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Ruijter, J.M.; Benes, V.; Van Den Hoff, M.J.B. Web-Based LinRegPCR: Application for the Visualization and Analysis of (RT)-qPCR Amplification and Melting Data. BMC Bioinform. 2021, 22, 398. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase Update, a Database of Eukaryotic Repetitive Elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Hartley, S.W.; Mullikin, J.C. QoRTs: A Comprehensive Toolset for Quality Control and Data Processing of RNA-Seq Experiments. BMC Bioinform. 2015, 16, 224. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Raghu, G.; Berk, M.; Campochiaro, P.A.; Jaeschke, H.; Marenzi, G.; Richeldi, L.; Wen, F.-Q.; Nicoletti, F.; Calverley, P.M.A. The Multifaceted Therapeutic Role of N-Acetylcysteine (NAC) in Disorders Characterized by Oxidative Stress. Curr. Neuropharmacol. 2021, 19, 1202–1224. [Google Scholar] [CrossRef]
- Mokhtari, V.; Afsharian, P.; Shahhoseini, M.; Kalantar, M.; Moini, A. A Review on Various Uses of N-Acetyl Cysteine. Cell J. 2017, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Firsanov, D.V.; Solovjeva, L.V.; Mikhailov, V.M.; Svetlova, M.P. Methods for the Detection of DNA Damage. In Genome Stability; Elsevier: Amsterdam, The Netherlands, 2016; pp. 635–649. ISBN 978-0-12-803309-8. [Google Scholar]
- Brooks, C.L.; Gu, W. P53 Ubiquitination: Mdm2 and Beyond. Mol. Cell 2006, 21, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research Progress of Glutathione Peroxidase Family (GPX) in Redoxidation. Front. Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef]
- Qin, H.; Qiu, H.; He, S.-T.; Hong, B.; Liu, K.; Lou, F.; Li, M.; Hu, P.; Kong, X.; Song, Y.; et al. Efficient Disinfection of SARS-CoV-2-like Coronavirus, Pseudotyped SARS-CoV-2 and Other Coronaviruses Using Cold Plasma Induces Spike Protein Damage. J. Hazard. Mater. 2022, 430, 128414. [Google Scholar] [CrossRef]
- Ding, H.; Wang, T.; Sun, Y.; Zhang, Y.; Wei, J.; Cai, R.; Guo, C.; Yuan, Y.; Yue, T. Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus Acidoterrestris in Apple Juice. Foods 2023, 12, 1531. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xu, R.; Gou, L.; Liu, Z.; Zhao, Y.; Liu, D.; Zhang, L.; Chen, H.; Kong, M.G. Mechanism of Virus Inactivation by Cold Atmospheric-Pressure Plasma and Plasma-Activated Water. Appl. Environ. Microbiol. 2018, 84, e00726-18. [Google Scholar] [CrossRef]
- Kong, X.; Xue, S.; Li, H.; Yang, W.; Martynovich, E.F.; Ning, W.; Wang, R. Simulation Study on an Atmospheric Pressure Plasma Jet Interacting with a Single Fiber: Effects of the Fiber’s Permittivity. Plasma Sources Sci. Technol. 2022, 31, 095010. [Google Scholar] [CrossRef]
- Kong, X.; Li, H.; Yang, W.; Li, S.; Yang, D.; Ning, W.; Wang, R. Atmospheric Pressure Plasma Jet Impinging on Fiber Arrays: Penetration Pattern Determined by Fiber Spacing. Appl. Phys. Lett. 2023, 122, 084101. [Google Scholar] [CrossRef]
- Yan, D.; Malyavko, A.; Wang, Q.; Ostrikov, K.; Sherman, J.H.; Keidar, M. Multi-Modal Biological Destruction by Cold Atmospheric Plasma: Capability and Mechanism. Biomedicines 2021, 9, 1259. [Google Scholar] [CrossRef]
- Volotskova, O.; Hawley, T.S.; Stepp, M.A.; Keidar, M. Targeting the Cancer Cell Cycle by Cold Atmospheric Plasma. Sci. Rep. 2012, 2, 636. [Google Scholar] [CrossRef]
- Moniruzzaman, R.; Rehman, M.U.; Zhao, Q.-L.; Jawaid, P.; Mitsuhashi, Y.; Imaue, S.; Fujiwara, K.; Ogawa, R.; Tomihara, K.; Saitoh, J.; et al. Roles of Intracellular and Extracellular ROS Formation in Apoptosis Induced by Cold Atmospheric Helium Plasma and X-Irradiation in the Presence of Sulfasalazine. Free Radic. Biol. Med. 2018, 129, 537–547. [Google Scholar] [CrossRef]
- Abduvokhidov, D.; Yusupov, M.; Shahzad, A.; Attri, P.; Shiratani, M.; Oliveira, M.C.; Razzokov, J. Unraveling the Transport Properties of RONS across Nitro-Oxidized Membranes. Biomolecules 2023, 13, 1043. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Ma, J.; Yu, K.N.; Li, W.; Cheng, C.; Bao, L.; Han, W. Non-Thermal Plasma Treatment Altered Gene Expression Profiling in Non-Small-Cell Lung Cancer A549 Cells. BMC Genom. 2015, 16, 435. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Tong, T.; Fan, W.; Fan, F.; Antinore, M.J.; Zhu, X.; Mazzacurati, L.; Li, X.; Petrik, K.L.; Rajasekaran, B.; et al. GADD45-Induced Cell Cycle G2-M Arrest Associates with Altered Subcellular Distribution of Cyclin B1 and Is Independent of P38 Kinase Activity. Oncogene 2002, 21, 8696–8704. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-N.; Oh, C.; Chang, J.W.; Liu, L.; Lim, M.A.; Jin, Y.L.; Piao, Y.; Kim, H.J.; Won, H.-R.; Lee, S.E.; et al. EGR1/GADD45α Activation by ROS of Non-Thermal Plasma Mediates Cell Death in Thyroid Carcinoma. Cancers 2021, 13, 351. [Google Scholar] [CrossRef]
- Oh, C.; Won, H.-R.; Kang, W.S.; Kim, D.-W.; Jung, S.-N.; Im, M.A.; Liu, L.; Jin, Y.L.; Piao, Y.; Kim, H.J.; et al. Head and Neck Cancer Cell Death Due to Mitochondrial Damage Induced by Reactive Oxygen Species from Nonthermal Plasma-Activated Media: Based on Transcriptomic Analysis. Oxidative Med. Cell. Longev. 2021, 2021, 9951712. [Google Scholar] [CrossRef]
- Jiang, H.-Y.; Wek, S.A.; McGrath, B.C.; Lu, D.; Hai, T.; Harding, H.P.; Wang, X.; Ron, D.; Cavener, D.R.; Wek, R.C. Activating Transcription Factor 3 Is Integral to the Eukaryotic Initiation Factor 2 Kinase Stress Response. Mol. Cell. Biol. 2004, 24, 1365–1377. [Google Scholar] [CrossRef]
- Wek, R.C.; Jiang, H.-Y.; Anthony, T.G. Coping with Stress: eIF2 Kinases and Translational Control. Biochem. Soc. Trans. 2006, 34, 7–11. [Google Scholar] [CrossRef]
- Troitskaya, O.; Novak, D.; Varlamov, M.; Biryukov, M.; Nushtaeva, A.; Kochneva, G.; Zakrevsky, D.; Schweigert, I.; Richter, V.; Koval, O. Immunological Effects of Cold Atmospheric Plasma-Treated Cells in Comparison with Those of Cells Treated with Lactaptin-Based Anticancer Drugs. Biophysica 2022, 2, 266–280. [Google Scholar] [CrossRef]
- Troitskaya, O.S.; Novak, D.D.; Richter, V.A.; Koval, O.A. Immunogenic Cell Death in Cancer Therapy. Acta Nat. 2022, 14, 40–53. [Google Scholar] [CrossRef]
- Lin, A.; Gorbanev, Y.; De Backer, J.; Van Loenhout, J.; Van Boxem, W.; Lemière, F.; Cos, P.; Dewilde, S.; Smits, E.; Bogaerts, A. Non-Thermal Plasma as a Unique Delivery System of Short-Lived Reactive Oxygen and Nitrogen Species for Immunogenic Cell Death in Melanoma Cells. Adv. Sci. 2019, 6, 1802062. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Ai, H.; Li, K.; Yao, X.; Zhu, W.; Liu, L.; Yu, C.; Song, Z.; Bao, Y.; Huang, Y.; et al. Prohibitin 2 Localizes in Nucleolus to Regulate Ribosomal RNA Transcription and Facilitate Cell Proliferation in RD Cells. Sci. Rep. 2018, 8, 1479. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Grummt, I. Ribosome Biogenesis and Cell Growth: mTOR Coordinates Transcription by All Three Classes of Nuclear RNA Polymerases. Oncogene 2006, 25, 6384–6391. [Google Scholar] [CrossRef] [PubMed]
- Stępiński, D. The Nucleolus, an Ally, and an Enemy of Cancer Cells. Histochem. Cell Biol. 2018, 150, 607–629. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5′ → 3′ | Product Size, bp |
|---|---|---|
| HPRT1 | F: CTCGAGATGTGATGAAGGAG | 258 |
| R: TATCTTCCACAATCAAGACATT | ||
| GAPDH | F: GAAGATGGTGATGGGATTTC | 226 |
| R: 5′-GAAGGTGAAGGTCGGAGT | ||
| KLF4 | F: GGGAGAAGACACTGCGTCAA | 229 |
| R: TCCAGGTCCAGGAGATCGTT | ||
| FOS | F: GCGTTGTGAAGACCATGACAG | 165 |
| R: GTGTATCAGTCAGCTCCCTCC | ||
| ATF3 | F: TTTGCTAACCTGACGCCCTT | 218 |
| R: TTGTTTCGGCACTTTGCAGC | ||
| GADD45B | F: GTACGAGTCGGCCAAGTTGA | 263 |
| R: CCGTGTGAGGGTTCGTGAC | ||
| GPX4 | F: GTGAGGCAAGACCGAAGTA | 275 |
| R: TCCACTTGATGGCATTTCCC | ||
| GPX7 | F: ACTTCAAGGCGGTCAACATC | 235 |
| R: CGGGCAAAGCTCTCAATCTC |
| Type of Regulation | Selected Terms | Representative Genes | Number of Genes |
|---|---|---|---|
| Up | CEBPB ENCODE | CBX4; GADD45A; SPRY4; ARID5B; KLHL24; FBXO32; ISG20; BCL6; NFIL3; DDIT3 | 23 |
| CEBPD ENCODE | PPP1R15A; HDAC5; BTG2; BTG1; TOB1; FAM117A; HHEX; NFIL3; JUNB; IER5 | 58 | |
| FOXA1 ENCODE | HEXIM1; PPP1R15A; CHIC2; BTG1; CXCL8; CITED2; FOS; DUSP6; BCL6; KLF9 | 29 | |
| FOXA2 ENCODE | HEXIM1; PPP1R15A; ERRFI1; CITED2; PTPRH; TM4SF1; DUSP6; DUSP10; KLF5; TP53INP2 | 36 | |
| KLF4 CHEA | ZNF296; CDKN1A; CITED2; ZFAND2A; ADM;GATA2; HOXB13; EFNB2; SOCS3; ZFP36 | 61 | |
| NELFE ENCODE | HEXIM1; EGR1; MYLIP; BTG1; GADD45B; GADD45A; CITED2; FAM46C; GDF15; FOS | 38 | |
| NFE2L2 CHEA | CHIC2; CDKN1A; BTG1; CITED2; IRS2; PTGS2; PPM1D; RND3; OASL; EFNB2 | 70 | |
| STAT3 ENCODE | EGR1; JUN; BTG2; BTG1; GADD45A; FOS; PRDM1; TOB1; PNRC1; ISG20 | 43 | |
| TP53 CHEA | CDKN1A; BTG2; BTG1; PRDM1; ADRB2; PPM1D; BBC3; SERTAD1; PRX; SESN2 | 38 | |
| Down | AR CHEA | FBN2; USP13; BNC2; PRKDC; LAMA3; ADAMTS12; SPATA5; LPP; PTPRF; SLC8A1 | 67 |
| E2F4 ENCODE | TOP2A; ARHGAP11A; CCNF; KIF14; HJURP; BUB1B; KIF11; TTF2; MKI67; SMC4 | 79 | |
| NFYA ENCODE | TOP2A;CCNF;KIF14;HJURP;TTF2;CDC20;TNFAIP8L1;GTSE1;LBR;DLGAP5 | 97 | |
| NFYB ENCODE | TOP2A;GLDC;CCNF;KIF14;HJURP;KIF11;TTF2;MKI67;LMNB1;CDC20 | 145 | |
| SMAD4 CHEA | PRELID2; NRP2; PKDCC; BNC2; LYPD1; PRICKLE2; NRXN3; ARHGAP26; PLEKHA7; SLC8A1 | 41 |
| Type of Regulation | Selected Terms | Representative Genes | Number of Genes |
|---|---|---|---|
| Up | FOSL2 ENCODE | GADD45A; ITGA2; TNFAIP3; NR1D1; DUSP14; ITPKC; BFSP1; DDIT3; ZNF219; ZBTB7B | 30 |
| GATA2 CHEA | BTG2; CDKN1A; TENM4; FHL2; KLHDC7A; CLN8; PPCDC; ZFP36L2; SLC9A1; FAM110A | 55 | |
| MAX ENCODE | HES6; PLK3; DYRK3; PANK1; DUSP1; INSR; SPHK1; PVT1; THUMPD2; TNFAIP3 | 81 | |
| MYC ENCODE | PLK3; DUSP1; PVT1; THUMPD2; AEN; SLC3A2; NR1D1; ZBTB5; SNHG5; ZFP36L2 | 58 | |
| NFIC ENCODE | TRAF4; GADD45A; DUSP1; ZFAS1; BHLHE40; MDM2; ZBTB7B; SLC25A45; PPM1D; DDB2 | 27 | |
| RELA ENCODE | CSF1; KSR1; ZFAS1; TNFAIP3; SLC3A2; ZFP36L2; FAM110A; ERN1; ACTA2; DNAJB2 | 38 | |
| TP53 CHEA | BTG2; CDKN1A; DYRK3; CEP85L; FHL2; ADRB2; PPM1D; EDA2R; BBC3; PIDD1 | 47 | |
| Down | AR CHEA | USP13; INSIG2; LRRC17; CHRDL1; SLC8A1; RHOBTB1; DTWD2; ADAMTS15; SH3BGRL; LEPR | 56 |
| E2F4 ENCODE | TOP2A; CDKN2C; BORA; FANCL; IFT80; KIF11; CTDSPL2; DCK; LMNB1; AURKA | 37 | |
| E2F6 ENCODE | HIBADH; CCDC126; SPIN4; CETN3; VLDLR; MANEA; LMNB1; PDK3; SLC25A40; FAM3C | 106 | |
| NFE2L2 CHEA | INSIG2; GCNT1; PPM1H; ADD3; TFPI; SLC4A4; NALCN; TMTC4; GJA1; PRKAR2B | 59 | |
| NFYA ENCODE | TOP2A; KDM3A; ARL6IP1; HIBADH; PIGN; DCUN1D4; CETN3; SEL1L; HMMR; TMTC4 | 81 | |
| NFYB ENCODE | TOP2A; ARL6IP1; HIBADH; PIGN; CETN3; KIF11; CTDSPL2; NXT2; LMNB1; HS2ST1 | 124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biryukov, M.; Semenov, D.; Kryachkova, N.; Polyakova, A.; Patrakova, E.; Troitskaya, O.; Milakhina, E.; Poletaeva, J.; Gugin, P.; Ryabchikova, E.; et al. The Molecular Basis for Selectivity of the Cytotoxic Response of Lung Adenocarcinoma Cells to Cold Atmospheric Plasma. Biomolecules 2023, 13, 1672. https://doi.org/10.3390/biom13111672
Biryukov M, Semenov D, Kryachkova N, Polyakova A, Patrakova E, Troitskaya O, Milakhina E, Poletaeva J, Gugin P, Ryabchikova E, et al. The Molecular Basis for Selectivity of the Cytotoxic Response of Lung Adenocarcinoma Cells to Cold Atmospheric Plasma. Biomolecules. 2023; 13(11):1672. https://doi.org/10.3390/biom13111672
Chicago/Turabian StyleBiryukov, Mikhail, Dmitriy Semenov, Nadezhda Kryachkova, Alina Polyakova, Ekaterina Patrakova, Olga Troitskaya, Elena Milakhina, Julia Poletaeva, Pavel Gugin, Elena Ryabchikova, and et al. 2023. "The Molecular Basis for Selectivity of the Cytotoxic Response of Lung Adenocarcinoma Cells to Cold Atmospheric Plasma" Biomolecules 13, no. 11: 1672. https://doi.org/10.3390/biom13111672
APA StyleBiryukov, M., Semenov, D., Kryachkova, N., Polyakova, A., Patrakova, E., Troitskaya, O., Milakhina, E., Poletaeva, J., Gugin, P., Ryabchikova, E., Zakrevsky, D., Schweigert, I., & Koval, O. (2023). The Molecular Basis for Selectivity of the Cytotoxic Response of Lung Adenocarcinoma Cells to Cold Atmospheric Plasma. Biomolecules, 13(11), 1672. https://doi.org/10.3390/biom13111672