
LncRNAs and CircRNAs as Strategies against Pathological Conditions Caused by a Hypoxic/Anoxic State


Abstract
:1. Introduction
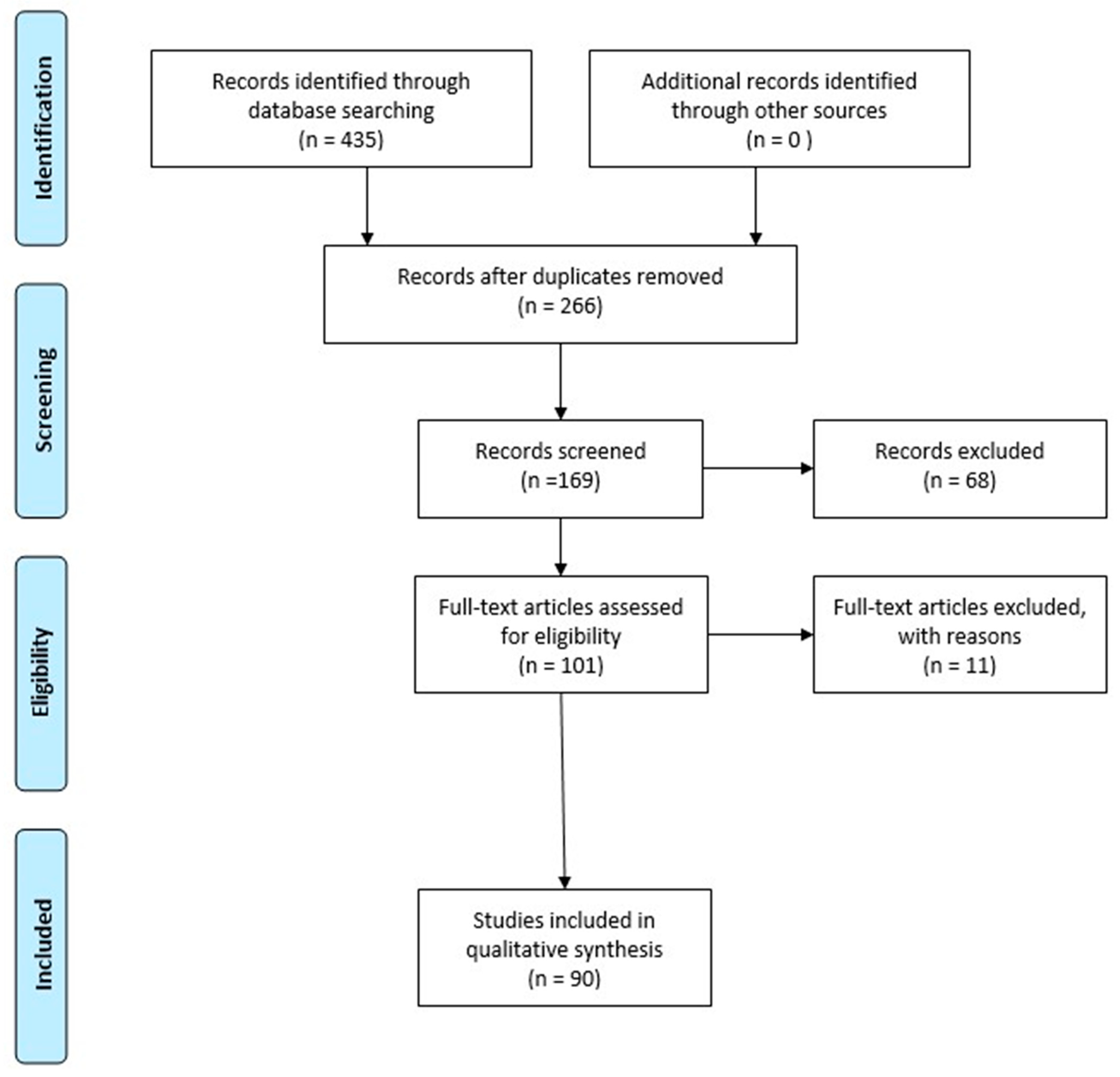
2. Methodology
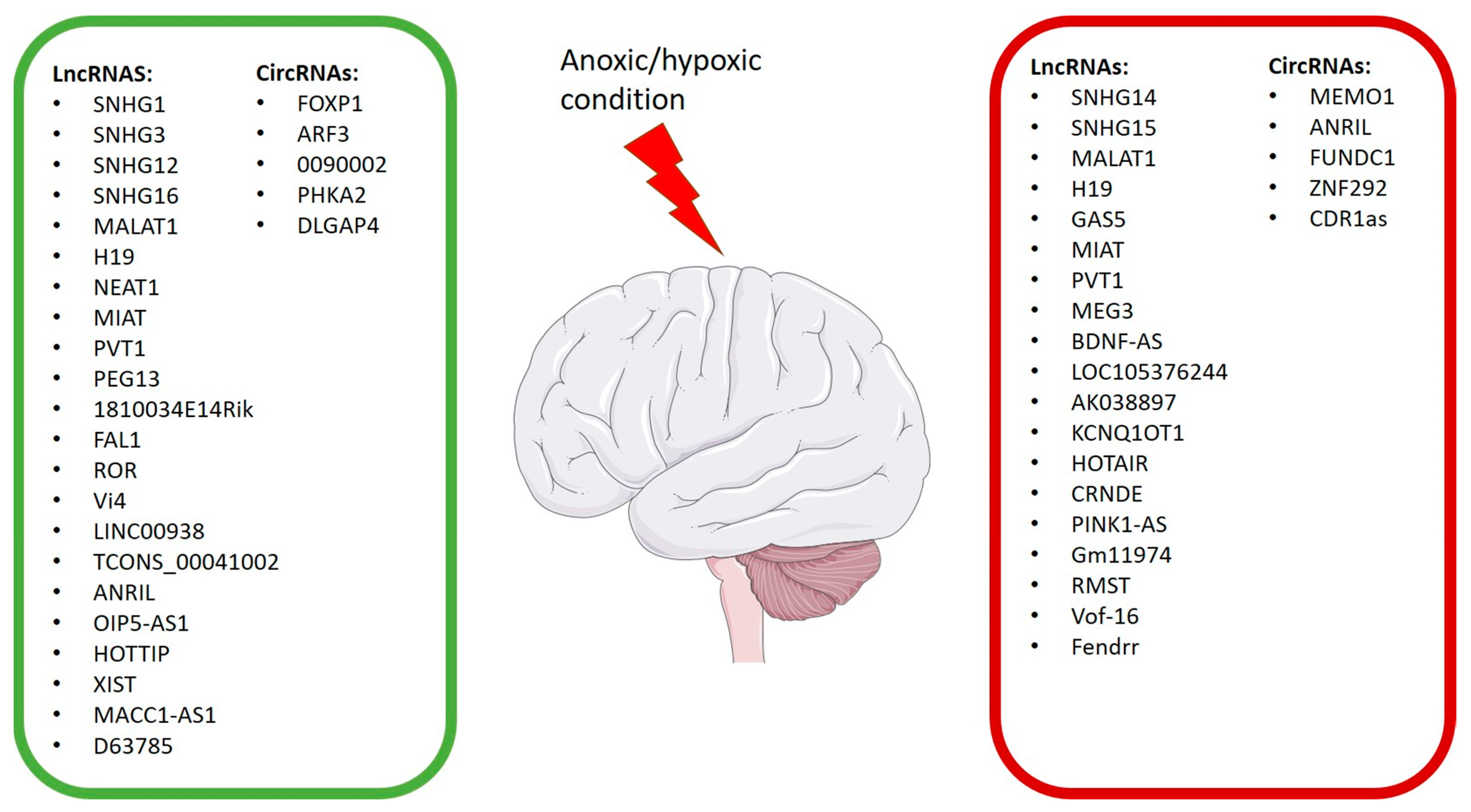
3. LncRNAs and circRNAs in Hypoxic Brain Damage
4. Studies about lncRNAs in Hypoxic/Anoxic Conditions
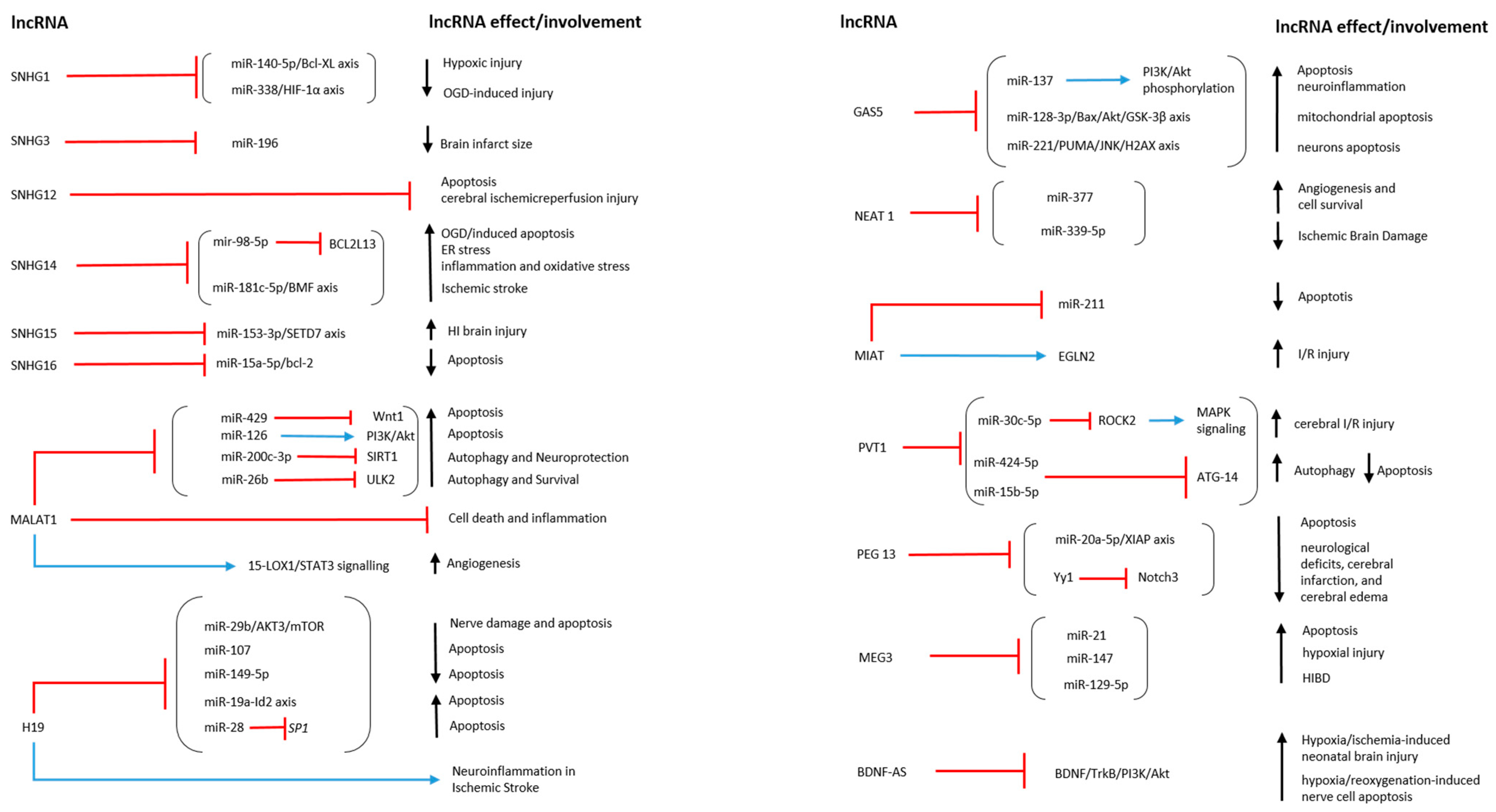
4.1. SNHG Family in Hypoxic/Anoxic Conditions
4.2. LncRNA MALAT1 in Hypoxic/Anoxic Conditions
4.3. LncRNA H19 in Hypoxic/Anoxic Conditions
4.4. LncRNA GAS5 in Hypoxic/Anoxic Conditions
4.5. LncRNA NEAT1 in Hypoxic/Anoxic Conditions
4.6. LncRNA MIAT in Hypoxic/Anoxic Conditions
4.7. LncRNA PVT1 in Hypoxic/Anoxic Conditions
4.8. LncRNA PEG13 in Hypoxic/Anoxic Conditions
4.9. LncRNA MEG3 in Hypoxic/Anoxic Conditions
4.10. LncRNA BDNF-AS in Hypoxic/Anoxic Conditions
4.11. Other lncRNAs That Play a Role in Exacerbating Hypoxic/Anoxic Conditions
4.12. Other lncRNAs Possessing Neuroprotective Properties
4.13. Experimental Studies That Reported the Effect on lncRNAs Modulation in Hypoxic/Anoxic Conditions Induced by Natural or Chemical Compounds
4.14. Experimental Studies That Have Focused on the Change in the Expression of Multiple lncRNAs in Hypoxic/Anoxic Conditions
5. Studies about CircRNAs in Hypoxic/Anoxic Conditions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rainaldi, M.A.; Perlman, J.M. Pathophysiology of Birth Asphyxia. Clin. Perinatol. 2016, 43, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Sandroni, C.; Cronberg, T.; Sekhon, M. Brain injury after cardiac arrest: Pathophysiology, treatment, and prognosis. Intensive Care Med. 2021, 47, 1393–1414. [Google Scholar] [CrossRef] [PubMed]
- Moppett, I.K. Traumatic brain injury: Assessment, resuscitation and early management. Br. J. Anaesth. 2007, 99, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Keel, M.; Trentz, O. Pathophysiology of polytrauma. Injury 2005, 36, 691–709. [Google Scholar] [CrossRef] [PubMed]
- Winstanley, E.L.; Mahoney, J.J., 3rd; Castillo, F.; Comer, S.D. Neurocognitive impairments and brain abnormalities resulting from opioid-related overdoses: A systematic review. Drug Alcohol Dep. 2021, 226, 108838. [Google Scholar] [CrossRef] [PubMed]
- Busl, K.M.; Greer, D.M. Hypoxic-ischemic brain injury: Pathophysiology, neuropathology and mechanisms. NeuroRehabilitation 2010, 26, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.S.; Holmes, P.A.; Koutroumanidis, M.A. Hypoxic-ischaemic brain injury. Pract. Neurol. 2011, 11, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Kurinczuk, J.J.; White-Koning, M.; Badawi, N. Epidemiology of neonatal encephalopathy and hypoxic-ischaemic encephalopathy. Early Hum. Dev. 2010, 86, 329–338. [Google Scholar] [CrossRef]
- Yang, X.; Niu, X.; Xiao, Y.; Lin, K.; Chen, X. MiRNA expression profiles in healthy OSAHS and OSAHS with arterial hypertension: Potential diagnostic and early warning markers. Respir. Res. 2018, 19, 194. [Google Scholar] [CrossRef]
- Shao, H.; Shen, P.; Chen, J. Expression Profile Analysis and Image Observation of miRNA in Serum of Patients with Obstructive Sleep Apnea-Hypopnea Syndrome. Contrast Med. Mol. Imag. 2021, 2021, 9731502. [Google Scholar] [CrossRef]
- Peters, M.M.C.; Sampaio-Pinto, V.; da Costa Martins, P.A. Non-coding RNAs in endothelial cell signalling and hypoxia during cardiac regeneration. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118515. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, J.; Lu, Q.; Huang, K.; Yang, B.; Reilly, J.; Jiang, N.; Shu, X.; Shang, L. An update on the functional roles of long non-coding RNAs in ischemic injury (Review). Int. J. Mol. Med. 2022, 50, 91. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Gedansky, A.; Hassett, C.E.; Price, C.; Fan, T.H.; Stephens, R.S.; Nyquist, P.; Uchino, K.; Cho, S.M. Pathophysiology of Brain Injury and Neurological Outcome in Acute Respiratory Distress Syndrome: A Scoping Review of Preclinical to Clinical Studies. Neur. Care 2021, 35, 518–527. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, D.; Verheijen, B.M.; Pasterkamp, R.J. Circular RNAs: Novel Regulators of Neuronal Development. Front. Mol. Neurosci. 2016, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Motti, D.; Bixby, J.L.; Lemmon, V.P. MicroRNAs and neuronal development. Semin. Fetal Neonatal Med. 2012, 17, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zuo, X.; Deng, H.; Liu, X.; Liu, L.; Ji, A. Roles of long noncoding RNAs in brain development, functional diversification and neurodegenerative diseases. Brain Res. Bull. 2013, 97, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Liu, D.Z.; Jickling, G.C.; Sharp, F.R.; Yin, K.J. MicroRNA-based therapeutics in central nervous system injuries. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2018, 38, 1125–1148. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wei, H.; Wu, J.Q. Coding and long non-coding gene expression changes in the CNS traumatic injuries. Cell. Mol. Life Sci. CMLS 2022, 79, 123. [Google Scholar] [CrossRef]
- Hartford, C.C.R.; Lal, A. When Long Noncoding Becomes Protein Coding. Mol. Cell. Biol. 2020, 40, e00528-19. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nature reviews. Mol. Cell. Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, B.; Biscarini, S.; Morlando, M. Non-coding RNAs in Nervous System Development and Disease. Front. Cell Dev. Biol. 2020, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Hamblin, M.H.; Yin, K.J. Non-coding RNAs in the regulation of blood-brain barrier functions in central nervous system disorders. Fluids Barriers CNS 2022, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (lncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non-Coding RNA 2021, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Min, L.; Qiu, X.; Wu, X.; Liu, C.; Ma, J.; Zhang, D.; Zhu, L. Biological Function of Long Non-coding RNA (LncRNA) Xist. Front. Cell Dev. Biol. 2021, 9, 645647. [Google Scholar] [CrossRef]
- Marques, A.C.; Ponting, C.P. Intergenic lncRNAs and the evolution of gene expression. Curr. Opin. Genet. Dev. 2014, 27, 48–53. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Pandey, V.; Bhattacharyya, M.; Dey, A. Regulatory role of long non coding RNAs (lncRNAs) in neurological disorders: From novel biomarkers to promising therapeutic strategies. Asian J. Pharm. Sci. 2021, 16, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Du, Y.; Liu, J.; Gareev, I.; Yang, G.; Kang, X.; Wang, X.; Beylerli, O.; Chen, X. Hypoxia related long non-coding RNAs in ischemic stroke. Non-Coding RNA Res. 2021, 6, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Schaukowitch, K.; Kim, T.K. Emerging epigenetic mechanisms of long non-coding RNAs. Neuroscience 2014, 264, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yang, F.; Cao, H.; Liang, Z. Malat1 regulates serum response factor through miR-133 as a competing endogenous RNA in myogenesis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 3054–3064. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.; Pontarelli, F.; Prendergast, E.; Dharap, A. Discovery of novel stroke-responsive lncRNAs in the mouse cortex using genome-wide RNA-seq. Neurobiol. Dis. 2017, 108, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Dharap, A.; Nakka, V.P.; Vemuganti, R. Effect of focal ischemia on long noncoding RNAs. Stroke 2012, 43, 2800–2802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hamblin, M.H.; Yin, K.J. The long noncoding RNA Malat1: Its physiological and pathophysiological functions. RNA Biol. 2017, 14, 1705–1714. [Google Scholar] [CrossRef]
- Zhu, H.; Xing, Z.; Zhao, Y.; Hao, Z.; Li, M. The Role of Circular RNAs in Brain Injury. Neuroscience 2020, 428, 50–59. [Google Scholar] [CrossRef]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; Christoff, A.P.; Margis, R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J. Biotechnol. 2016, 238, 42–51. [Google Scholar] [CrossRef]
- Leung, A.K.; Sharp, P.A. MicroRNA functions in stress responses. Mol. Cell 2010, 40, 205–215. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glazar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Zhao, F.; Qu, Y.; Liu, J.; Liu, H.; Zhang, L.; Feng, Y.; Wang, H.; Gan, J.; Lu, R.; Mu, D. Microarray Profiling and Co-Expression Network Analysis of LncRNAs and mRNAs in Neonatal Rats Following Hypoxic-ischemic Brain Damage. Sci. Rep. 2015, 5, 13850. [Google Scholar] [CrossRef]
- Minhas, G.; Mathur, D.; Ragavendrasamy, B.; Sharma, N.K.; Paanu, V.; Anand, A. Hypoxia in CNS Pathologies: Emerging Role of miRNA-Based Neurotherapeutics and Yoga Based Alternative Therapies. Front. Neurosci. 2017, 11, 386. [Google Scholar] [CrossRef]
- Gutierrez, G. Cellular energy metabolism during hypoxia. Crit. Care Med. 1991, 19, 619–626. [Google Scholar] [CrossRef]
- Zauner, A.; Daugherty, W.P.; Bullock, M.R.; Warner, D.S. Brain oxygenation and energy metabolism: Part I-biological function and pathophysiology. Neurosurgery 2002, 51, 289–301, discussion 302. [Google Scholar]
- Boutilier, R.G. Mechanisms of cell survival in hypoxia and hypothermia. J. Exp. Biol. 2001, 204, 3171–3181. [Google Scholar] [CrossRef] [PubMed]
- Loor, G.; Schumacker, P.T. Role of hypoxia-inducible factor in cell survival during myocardial ischemia-reperfusion. Cell Death Differ. 2008, 15, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Buga, A.M.; Di Napoli, M.; Popa-Wagner, A. Preclinical models of stroke in aged animals with or without comorbidities: Role of neuroinflammation. Biogerontology 2013, 14, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Sandu, R.E.; Buga, A.M.; Uzoni, A.; Petcu, E.B.; Popa-Wagner, A. Neuroinflammation and comorbidities are frequently ignored factors in CNS pathology. Neural Regen. Res. 2015, 10, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Katschinski, D.M. Is there a molecular connection between hypoxia and aging? Exp. Gerontol. 2006, 41, 482–484. [Google Scholar] [CrossRef] [PubMed]
- Venø, M.T.; Hansen, T.B.; Venø, S.T.; Clausen, B.H.; Grebing, M.; Finsen, B.; Holm, I.E.; Kjems, J. Spatio-temporal regulation of circular RNA expression during porcine embryonic brain development. Genome Biol. 2015, 16, 245. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.J.; Cao, S.Q.; Gao, L.B.; Wang, Y.Y.; Zhou, B.; Hu, X.; Pu, Y.; Li, Z.L.; Wang, Q.; Xiao, X.; et al. Circular Ribonucleic Acid Expression Profile in Mouse Cortex after Traumatic Brain Injury. J. Neurotrauma 2019, 36, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Gokool, A.; Anwar, F.; Voineagu, I. The Landscape of Circular RNA Expression in the Human Brain. Biol. Psychiatry 2020, 87, 294–304. [Google Scholar] [CrossRef]
- Mehta, S.L.; Pandi, G.; Vemuganti, R. Circular RNA Expression Profiles Alter Significantly in Mouse Brain After Transient Focal Ischemia. Stroke 2017, 48, 2541–2548. [Google Scholar] [CrossRef]
- Saeinasab, M.; Atlasi, Y.; Matin, M.M. Functional role of lncRNAs in gastrointestinal malignancies: The peculiar case of small nucleolar RNA host gene family. FEBS J. 2022. [Google Scholar] [CrossRef]
- Tan, A.Q.; Zheng, Y.F. The Roles of SNHG Family in Osteoblast Differentiation. Genes 2022, 13, 2268. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Gu, X.; Zheng, Q.; Guo, Z.; Shan, D.; Wang, J.; Zhu, H. Long noncoding RNA SNHG4: A novel target in human diseases. Cancer Cell Int. 2021, 21, 583. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.W.; Lou, X.Q.; Liu, Z.L.; Zhang, N.; Pang, L. LncRNA SNHG1 protects SH-SY5Y cells from hypoxic injury through miR-140-5p/Bcl-XL axis. Int. J. Neurosci. 2021, 131, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zi, X.H. LncRNA SNHG1 alleviates OGD induced injury in BMEC via miR-338/HIF-1α axis. Brain Res. 2019, 1714, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, M.F.; Zhu, L.H.; Qiao, L.X.; Zhao, R.B.; Xia, Z.K. Long non-coding RNA Snhg3 protects against hypoxia/ischemia-induced neonatal brain injury. Exp. Mol. Pathol. 2020, 112, 104343. [Google Scholar] [CrossRef]
- Cheng, Y.; Jiang, Y.; Sun, Y.; Jiang, H. The role of long non-coding RNA SNHG12 in neuroprotection following cerebral ischemic injury. Neuroreport 2019, 30, 945–952. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, Y.; Cai, J.; Zhang, D.; Liu, S.; Pang, B. LncRNA SNHG12 Improves Cerebral Ischemic-reperfusion Injury by Activating SIRT1/FOXO3a Pathway through I nhibition of Autophagy and Oxidative Stress. Curr. Neurovasc. Res. 2020, 17, 394–401. [Google Scholar] [CrossRef]
- Zhang, G.; Guo, J.; Zeng, J.; Zhang, X.; Chen, R.; Wang, G.; Liang, W. LncRNA SNHG14 is beneficial to oxygen glucose deprivation/reoxygenation-induced neuro-2a cell injury via mir-98-5p sequestration-caused BCL2L13 upregulation. Metab. Brain Dis. 2022, 37, 2005–2016. [Google Scholar] [CrossRef]
- Bu, X.; Zhao, Y.; Chang, M.; Ge, X. Downregulation of lncRNA SNHG14 alleviates neurons injury by modulating the miR-181c-5p/BMF axis in ischemic stroke. Brain Res. Bull. 2021, 174, 379–388. [Google Scholar] [CrossRef]
- Fu, J.; Huang, Y.; Xian, L. LncRNA SNHG15 regulates hypoxic-ischemic brain injury via miR-153-3p/SETD7 axis. Histol. Histopathol. 2022, 37, 1113–1125. [Google Scholar] [CrossRef]
- Teng, H.; Li, M.; Qian, L.; Yang, H.; Pang, M. Long non-coding RNA SNHG16 inhibits the oxygen-glucose deprivation and reoxygenation-induced apoptosis in human brain microvascular endothelial cells by regulating miR-15a-5p/bcl-2. Mol. Med. Rep. 2020, 22, 2685–2694. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Li, H.F.; He, M.H.; Yan, J.Y.; Yang, M.; Zhang, F.X.; Wang, R.R.; Wang, Q.Y.; Zhang, J.P. Long non-coding RNA MALAT1 sponges microRNA-429 to regulate apoptosis of hippocampal neurons in hypoxic-ischemic brain damage by regulating WNT1. Brain Res. Bull. 2019, 152, 1–10. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, H.; Li, W.J.; Liu, Y.H. LncRNA MALAT1 Promotes OGD-Induced Apoptosis of Brain Microvascular Endothelial Cells by Sponging miR-126 to Repress PI3K/Akt Signaling Pathway. Neurochem. Res. 2020, 45, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Han, X.; Mao, Z.; Xin, Y.; Maharjan, S.; Zhang, B. MALAT1 lncRNA Induces Autophagy and Protects Brain Microvascular Endothelial Cells Against Oxygen-Glucose Deprivation by Binding to miR-200c-3p and Upregulating SIRT1 Expression. Neuroscience 2019, 397, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, J.; Tang, N. Long noncoding RNA Malat1 is a potent autophagy inducer protecting brain microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury by sponging miR-26b and upregulating ULK2 expression. Neuroscience 2017, 354, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, X.; Liu, K.; Hamblin, M.H.; Yin, K.J. Long Noncoding RNA Malat1 Regulates Cerebrovascular Pathologies in Ischemic Stroke. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Qu, Y.; Suo, R.; Zhu, Y. Long non-coding RNA MALAT1 regulates angiogenesis following oxygen-glucose deprivation/reoxygenation. J. Cell. Mol. Med. 2019, 23, 2970–2983. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, L.; Chen, J.; Shen, H.; Chen, Z. Mechanisms Underlying Abnormal Expression of lncRNA H19 in Neonatal Hypoxic-Ischemic Encephalopathy. Am. J. Perinatol. 2022, 39, 844–852. [Google Scholar] [CrossRef]
- Fang, H.; Li, H.F.; Pan, Q.; Yang, M.; Zhang, F.X.; Wang, R.R.; Wang, Q.Y.; Zhang, J.P. Long Noncoding RNA H19 Overexpression Protects against Hypoxic-Ischemic Brain Damage by Inhibiting miR-107 and Up-Regulating Vascular Endothelial Growth Factor. Am. J. Pathol. 2021, 191, 503–514. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, T.; Wang, P.; Shen, Y.; Huang, T. Overexpression of Long Noncoding RNA H19 Inhibits Cardiomyocyte Apoptosis in Neonatal Rats with Hypoxic-Ischemic Brain Damage Through the miR-149-5p/LIF/PI3K/Akt Axis. Biopreserv. Biobank. 2021, 19, 376–385. [Google Scholar] [CrossRef]
- Xiao, Z.; Qiu, Y.; Lin, Y.; Medina, R.; Zhuang, S.; Rosenblum, J.S.; Cui, J.; Li, Z.; Zhang, X.; Guo, L. Blocking lncRNA H19-miR-19a-Id2 axis attenuates hypoxia/ischemia induced neuronal injury. Aging 2019, 11, 3585–3600. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, X.; Guo, R.; Meng, J. Protective effects of lncRNA H19 silence against hypoxia-induced injury in PC-12 cells by regulating miR-28. Int. J. Biol. Macromol. 2019, 121, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, H.; Fan, Z.; Li, G.; Ma, Q.; Tao, Z.; Wang, R.; Feng, J.; Luo, Y. Long Noncoding RNA H19 Promotes Neuroinflammation in Ischemic Stroke by Driving Histone Deacetylase 1-Dependent M1 Microglial Polarization. Stroke 2017, 48, 2211–2221. [Google Scholar] [CrossRef]
- Jing, W.; Tuxiu, X.; Xiaobing, L.; Guijun, J.; Lulu, K.; Jie, J.; Lu, Y.; Liying, Z.; Xiaoxing, X.; Jingjun, L. LncRNA GAS5/miR-137 Is a Hypoxia-Responsive Axis Involved in Cardiac Arrest and Cardiopulmonary Cerebral Resuscitation. Front. Immunol. 2021, 12, 790750. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Z.; Wang, H. Downregulation of lncRNA GAS5 prevents mitochondrial apoptosis and hypoxic-ischemic brain damage in neonatal rats through the microRNA-128-3p/Bax/Akt/GSK-3β axis. Neuroreport 2021, 32, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.B.; Lai, L.F.; Xie, G.B.; Ding, C.; Xu, X.; Wang, Y. LncRNAGAS5 sponges miRNA-221 to promote neurons apoptosis by up-regulated PUMA under hypoxia condition. Neurol. Res. 2020, 42, 8–16. [Google Scholar] [CrossRef]
- Zhou, Z.W.; Zheng, L.J.; Ren, X.; Li, A.P.; Zhou, W.S. LncRNA NEAT1 facilitates survival and angiogenesis in oxygen-glucose deprivation (OGD)-induced brain microvascular endothelial cells (BMECs) via targeting miR-377 and upregulating SIRT1, VEGFA, and BCL-XL. Brain Res. 2019, 1707, 90–98. [Google Scholar] [CrossRef]
- Zhao, J.; He, L.; Yin, L. lncRNA NEAT1 Binds to MiR-339-5p to Increase HOXA1 and Alleviate Ischemic Brain Damage in Neonatal Mice. Mol. Ther. Nucleic Acids 2020, 20, 117–127. [Google Scholar] [CrossRef]
- Li, E.Y.; Zhao, P.J.; Jian, J.; Yin, B.Q.; Sun, Z.Y.; Xu, C.X.; Tang, Y.C.; Wu, H. LncRNA MIAT overexpression reduced neuron apoptosis in a neonatal rat model of hypoxic-ischemic injury through miR-211/GDNF. Cell Cycle 2019, 18, 156–166. [Google Scholar] [CrossRef]
- Li, S.; Fu, J.; Wang, Y.; Hu, C.; Xu, F. LncRNA MIAT enhances cerebral ischaemia/reperfusion injury in rat model via interacting with EGLN2 and reduces its ubiquitin-mediated degradation. J. Cell. Mol. Med. 2021, 25, 10140–10151. [Google Scholar] [CrossRef]
- Zhang, H.; Li, M.; Liang, J.; Li, M.; Sun, X. Long Non-coding RNA PVT1 Inhibits miR-30c-5p to Upregulate Rock2 to Modulate Cerebral Ischemia/Reperfusion Injury Through MAPK Signaling Pathway Activation. Mol. Neurobiol. 2021, 58, 6032–6048. [Google Scholar] [CrossRef]
- Zhang, P.; Gong, S.; Li, S.; Yuan, Z. PVT1 alleviates hypoxia-induced endothelial apoptosis by enhancing autophagy via the miR-15b-5p/ATG14 and miR-424-5p/ATG14 axis. Biochem. Biophys. Res. Commun. 2023, 671, 1–9. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Y.; Xue, H.; Zhang, Q.; Zhang, Y.; Shen, Y.; Bing, X. Long Non-coding RNA Peg13 Alleviates Hypoxic-Ischemic Brain Damage in Neonatal Mice via miR-20a-5p/XIAP Axis. Neurochem. Res. 2022, 47, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, C.; Fan, H.; Du, Y.; Zhang, R.; Zhan, S.; Zhang, G.; Bu, N. Gli2-induced lncRNA Peg13 alleviates cerebral ischemia-reperfusion injury by suppressing Yy1 transcription in a PRC2 complex-dependent manner. Metab. Brain Dis. 2023, 38, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Liang, H. Silencing MEG3 protects PC12 cells from hypoxic injury by targeting miR-21. Artif. Cells Nanomed. Biotechnol. 2020, 48, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Rao, J.; Yuan, J.; Gao, L.; Huang, W.; Zhao, L.; Ren, J. Long non-coding RNA MEG3 functions as a competing endogenous RNA to regulate ischemic neuronal death by targeting miR-21/PDCD4 signaling pathway. Cell Death Dis. 2017, 8, 3211. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Dong, Z.; Liu, N.; Xie, F.; Wang, N. Maternally Expressed Gene 3 (MEG3) Enhances PC12 Cell Hypoxia Injury by Targeting MiR-147. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 2457–2469. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.M.; Liu, J.; Wang, Y.; Zhang, M.H. Silencing of long noncoding RNA MEG3 enhances cerebral protection of dexmedetomidine against hypoxic-ischemic brain damage in neonatal mice by binding to miR-129-5p. J. Cell. Biochem. 2019, 120, 7978–7988. [Google Scholar] [CrossRef]
- Qiao, L.X.; Zhao, R.B.; Wu, M.F.; Zhu, L.H.; Xia, Z.K. Silencing of long non-coding antisense RNA brain-derived neurotrophic factor attenuates hypoxia/ischemia-induced neonatal brain injury. Int. J. Mol. Med. 2020, 46, 653–662. [Google Scholar] [CrossRef]
- Zhong, J.B.; Li, X.; Zhong, S.M.; Liu, J.D.; Chen, C.B.; Wu, X.Y. Knockdown of long noncoding antisense RNA brain-derived neurotrophic factor attenuates hypoxia/reoxygenation-induced nerve cell apoptosis through the BDNF-TrkB-PI3K/Akt signaling pathway. Neuroreport 2017, 28, 910–916. [Google Scholar] [CrossRef]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, Y.; Zhao, M.; Gao, Z. Neuro-protective roles of long non-coding RNA MALAT1 in Alzheimer’s disease with the involvement of the microRNA-30b/CNR1 network and the following PI3K/AKT activation. Exp. Mol. Pathol. 2020, 117, 104545. [Google Scholar] [CrossRef] [PubMed]
- Abrishamdar, M.; Jalali, M.S.; Rashno, M. MALAT1 lncRNA and Parkinson’s Disease: The role in the Pathophysiology and Significance for Diagnostic and Therapeutic Approaches. Mol. Neurobiol. 2022, 59, 5253–5262. [Google Scholar] [CrossRef] [PubMed]
- Ming, Y.; Deng, Z.; Tian, X.; Jia, Y.; Ning, M.; Cheng, S. m6A Methyltransferase METTL3 Reduces Hippocampal Neuron Apoptosis in a Mouse Model of Autism through the MALAT1/SFRP2/Wnt/β-catenin Axis. Psychiatry Investig. 2022, 19, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Chen, B.; Zhu, Z.; Du, C.; Gao, S.; Zhao, G.; Zhao, P.; Wang, Y.; Wang, A.; Schwartz, Z.; et al. Long noncoding RNA (lncRNA) H19: An essential developmental regulator with expanding roles in cancer, stem cell differentiation, and metabolic diseases. Genes Dis. 2023, 10, 1351–1366. [Google Scholar] [CrossRef] [PubMed]
- Bond, C.S.; Fox, A.H. Paraspeckles: Nuclear bodies built on long noncoding RNA. J. Cell Biol. 2009, 186, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Ishii, N.; Ozaki, K.; Sato, H.; Mizuno, H.; Susumu, S.; Takahashi, A.; Miyamoto, Y.; Ikegawa, S.; Kamatani, N.; Hori, M.; et al. Identification of a novel non-coding RNA, MIAT, that confers risk of myocardial infarction. J. Human Genet. 2006, 51, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Yao, W.; Wang, J.; Ma, X.; Xiao, W.; Li, H.; Xia, D.; Yang, Y.; Deng, K.; Xiao, H.; et al. LncRNAs expression signatures of renal clear cell carcinoma revealed by microarray. PLoS ONE 2012, 7, e42377. [Google Scholar] [CrossRef]
- Zou, B.; Wang, D.; Xu, K.; Liu, J.L.; Yuan, D.Y.; Meng, Z.; Zhang, B. Prognostic value of long non-coding RNA plasmacytoma variant translocation1 in human solid tumors: A meta-analysis. Medicine 2019, 98, e16087. [Google Scholar] [CrossRef]
- Barel, O.; Shalev, S.A.; Ofir, R.; Cohen, A.; Zlotogora, J.; Shorer, Z.; Mazor, G.; Finer, G.; Khateeb, S.; Zilberberg, N.; et al. Maternally inherited Birk Barel mental retardation dysmorphism syndrome caused by a mutation in the genomically imprinted potassium channel KCNK9. American J. Human Genet. 2008, 83, 193–199. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Mehta, K.R.; Danila, D.C.; Scolavino, S.; Johnson, S.R.; Klibanski, A. A pituitary-derived MEG3 isoform functions as a growth suppressor in tumor cells. J. Clin. Endocrinol. Metab. 2003, 88, 5119–5126. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhong, Y.; Wang, Y.; Zhang, X.; Batista, D.L.; Gejman, R.; Ansell, P.J.; Zhao, J.; Weng, C.; Klibanski, A. Activation of p53 by MEG3 non-coding RNA. J. Biol. Chem. 2007, 282, 24731–24742. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Wei, D.; Chen, S.; Chen, X.; Li, S.; Chen, W.; He, W. MiR-145 protected the cell viability of human cerebral cortical neurons after oxygen-glucose deprivation by downregulating EPHA4. Life Sci. 2019, 231, 116517. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Zhang, L.; Hu, W.; Wu, J.; Zhang, W. Long non-coding RNA AK038897 aggravates cerebral ischemia/reperfusion injury via acting as a ceRNA for miR-26a-5p to target DAPK1. Exp. Neurol. 2019, 314, 100–110. [Google Scholar] [CrossRef]
- Yi, M.; Li, Y.; Wang, D.; Zhang, Q.; Yang, L.; Yang, C. KCNQ1OT1 Exacerbates Ischemia-Reperfusion Injury Through Targeted Inhibition of miR-140-3P. Inflammation 2020, 43, 1832–1845. [Google Scholar] [CrossRef]
- Wang, Y.; Mao, J.; Li, X.; Wang, B.; Zhou, X. lncRNA HOTAIR mediates OGD/R-induced cell injury and angiogenesis in a EZH2-dependent manner. Exp. Ther. Med. 2022, 23, 99. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.H.; Lai, F.F.; Chen, S.; Yan, C.X.; Zhang, B.H.; Fang, C.Z.; Wang, G.H. Silencing of long non-coding RNA CRNDE promotes autophagy and alleviates neonatal hypoxic-ischemic brain damage in rats. Mol. Cell. Biochem. 2020, 472, 1–8. [Google Scholar] [CrossRef]
- Yang, Z.B.; Xiang, Y.; Zuo, M.L.; Mao, L.; Hu, G.H.; Song, G.L.; Sheikh, M.S.A.; Tan, L.M. lncRNA PINK1-AS Aggravates Cerebral Ischemia/Reperfusion Oxidative Stress Injury through Regulating ATF2 by Sponging miR-203. Oxidative Med. Cell. Longev. 2022, 2022, 1296816. [Google Scholar] [CrossRef]
- Cai, J.; Shangguan, S.; Li, G.; Cai, Y.; Chen, Y.; Ma, G.; Miao, Z.; Liu, L.; Deng, Y. Knockdown of lncRNA Gm11974 protect against cerebral ischemic reperfusion through miR-766-3p/NR3C2 axis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3847–3853. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, M.; Yan, F.; Cong, Y. Knockdown of RMST Impedes Neuronal Apoptosis and Oxidative Stress in OGD/R-Induced Ischemic Stroke Via Depending on the miR-377/SEMA3A Signal Network. Neurochem. Res. 2021, 46, 584–594. [Google Scholar] [CrossRef]
- Tan, Y.; Du, R.L.; Su, Z.Y.; Xue, L.L.; Liu, J.; Zhang, B.L.; Huang, S.J.; Li, L.J.; Xi-Yang, Y.B.; Xiong, L.L. Vof-16 knockout improves the recovery from hypoxic-ischemic brain damage of neonatal rats. Brain Res. 2020, 1748, 147070. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Zheng, Y.Y.; Zhou, H.J.; Zhang, X.X.; Wu, P.; Zhu, S.M. LncRNA-Fendrr protects against the ubiquitination and degradation of NLRC4 protein through HERC2 to regulate the pyroptosis of microglia. Mol. Med. 2021, 27, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, X.L.; Ji, B.Y.; Cao, X.; Yu, L.J.; Zhang, Y.; Bao, X.Y.; Xu, Y.; Jin, J.L. LncRNA-1810034E14Rik reduces microglia activation in experimental ischemic stroke. J. Neuroinflamm. 2019, 16, 75. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Fu, J.; Wang, Y. The lncRNA FAL1 protects against hypoxia-reoxygenation- induced brain endothelial damages through regulating PAK1. J. Bioenerg. Biomembr. 2020, 52, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; An, Y.; Tang, Y. Long noncoding RNA-regulator of reprogramming alleviates hypoxia-induced cerebral injury in mice model and human via modulating apoptosis complexes. J. Integr. Neurosci. 2019, 18, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.L.; Xue, L.L.; Du, R.L.; Zhou, H.L.; Tan, Y.X.; Ma, Z.; Jin, Y.; Zhang, Z.B.; Xu, Y.; Hu, Q.; et al. Vi4-miR-185-5p-Igfbp3 Network Protects the Brain From Neonatal Hypoxic Ischemic Injury via Promoting Neuron Survival and Suppressing the Cell Apoptosis. Front. Cell Dev. Biol. 2020, 8, 529544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Le, M.; Li, J.; Huang, Q.; Chen, H.; Zhang, W.; Mao, H.; Sun, Q.; Li, A.; Zhao, Y.; et al. LINC00938 alleviates hypoxia ischemia encephalopathy induced neonatal brain injury by regulating oxidative stress and inhibiting JNK/p38 MAPK signaling pathway. Exp. Neurol. 2023, 367, 114449. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.L.; Xue, L.L.; Du, R.L.; Xu, Y.; Niu, Y.J.; Hu, Q.; Zhou, H.L.; Liu, F.; Zhu, Z.Q.; Yu, C.Y.; et al. LncRNA TCONS_00041002 improves neurological outcomes in neonatal rats with hypoxic-ischemic encephalopathy by inhibiting apoptosis and promoting neuron survival. Exp. Neurol. 2021, 346, 113835. [Google Scholar] [CrossRef]
- Li, H.; Qi, J.; Wei, J.; Xu, B.; Min, S.; Wang, L.; Si, Y.; Qiu, H. Long non-coding RNA ANRIL mitigates neonatal hypoxic-ischemic brain damage via targeting the miR-378b/ATG3 axis. Am. J. Transl. Res. 2021, 13, 11585–11596. [Google Scholar]
- Chen, Y.; Liu, W.; Chen, M.; Sun, Q.; Chen, H.; Li, Y. Up-regulating lncRNA OIP5-AS1 protects neuron injury against cerebral hypoxia-ischemia induced inflammation and oxidative stress in microglia/macrophage through activating CTRP3 via sponging miR-186-5p. Int. Immunopharmacol. 2021, 92, 107339. [Google Scholar] [CrossRef]
- Wang, Y.; Li, G.; Zhao, L.; Lv, J. Long noncoding RNA HOTTIP alleviates oxygen-glucose deprivation-induced neuronal injury via modulating miR-143/hexokinase 2 pathway. J. Cell. Biochem. 2018, 119, 10107–10117. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Bai, X.; Liu, C.; Hu, Z. Long noncoding RNA XIST participates hypoxia-induced angiogenesis in human brain microvascular endothelial cells through regulating miR-485/SOX7 axis. Am. J. Transl. Res. 2019, 11, 6487–6497. [Google Scholar] [PubMed]
- Yan, G.; Zhao, H.; Hong, X. LncRNA MACC1-AS1 attenuates microvascular endothelial cell injury and promotes angiogenesis under hypoxic conditions via modulating miR-6867-5p/TWIST1 in human brain microvascular endothelial cells. Ann. Transl. Med. 2020, 8, 876. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, Y.; Chen, J.P.; Li, D.Z.; Jiang, Q.; Wu, T.; Zhou, X.Z. Oxygen glucose deprivation/re-oxygenation-induced neuronal cell death is associated with Lnc-D63785 m6A methylation and miR-422a accumulation. Cell Death Dis. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Xiaoqing, S.; Yinghua, C.; Xingxing, Y. The autophagy in ischemic stroke: A regulatory role of non-coding-RNAs. Cell. Signal. 2023, 104, 110586. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, W.; Zhang, Y.; Hu, Z.; Guo, H.; Lv, J.; Du, H. Dexmedetomidine had neuroprotective effects on hippocampal neuronal cells via targeting lncRNA SHNG16 mediated microRNA-10b-5p/BDNF axis. Mol. Cell. Biochem. 2020, 469, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Sun, Y.; Liu, H.; Li, Y.; An, N.; Wang, L.; Zhang, H.; Yang, F.; Xing, Y.; Gao, Y. Network Pharmacology and Bioinformatics Methods Reveal the Mechanism of Berberine in the Treatment of Ischaemic Stroke. Evid.-Based Complement. Altern. Med. eCAM 2022, 2022, 5160329. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, F.; Xu, H.; Yang, F.; Zhou, G.; Tong, M.; Li, Y.; Yang, S. Melatonin affects hypoxia-inducible factor 1alpha and ameliorates delayed brain injury following subarachnoid hemorrhage via H19/miR-675/HIF1A/TLR4. Bioengineered 2022, 13, 4235–4247. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Walayat, A.; Fu, Y.; Liu, B.; Zhang, L.; Xiao, D. The Regulatory Role of H19/miR-181a/ATG5 Signaling in Perinatal Nicotine Exposure-Induced Development of Neonatal Brain Hypoxic-Ischemic Sensitive Phenotype. Int. J. Mol. Sci. 2022, 23, 6885. [Google Scholar] [CrossRef]
- Qiao, P.; Yan, H.; Wang, J. EGb761 Protects Brain Microvascular Endothelial Cells Against Oxygen-Glucose Deprivation-Induced Injury Through lncRNA Rmst/miR-150 Axis. Neurochem. Res. 2020, 45, 2398–2408. [Google Scholar] [CrossRef]
- Yang, F.; Duan, H.; Ye, N.; Zeng, Y.; Yang, P.; Shao, B.; Wang, C.; Lin, G. beta-Asarone Protects PC12 Cells Against Hypoxia-Induced Injury Via Negatively Regulating RPPH1/MiR-542-3p/DEDD2 Axis. Cell Transplant. 2022, 31, 9636897221079336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Ouyang, B.; Ji, X.Y.; Liu, M.F. Gastrodin Alleviates Cerebral Ischaemia/Reperfusion Injury by Inhibiting Pyroptosis by Regulating the lncRNA NEAT1/miR-22-3p Axis. Neurochem. Res. 2021, 46, 1747–1758. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ye, C.; Cheng, S.; Chen, J. Propofol Downregulates lncRNA MALAT1 to Alleviate Cerebral Ischemia-Reperfusion Injury. Inflammation 2021, 44, 2580–2591. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ye, X.; Xie, H.; Liu, Y.; Xu, Y.; Wang, Y.; Zhou, Y.; Zhou, N.; Wang, J. Profiling analysis reveals the potential contribution of long non-coding RNAs to preterm white matter injury. Life Sci. 2020, 255, 117815. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Qu, Y.; Wang, D.; Zhu, Y. The Proangiogenic Roles of Long NonCoding RNAs Revealed by RNA-Sequencing Following Oxygen-Glucose Deprivation/Re-Oxygenation. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2019, 52, 708–727. [Google Scholar] [CrossRef]
- Zhao, R.B.; Zhu, L.H.; Li, H.J.; Fan, Z.M.; Xia, Z.K. High-throughput sequencing analysis of lncRNAs in hippocampus tissues with hypoxic-ischemic brain damage. Int. J. Clin. Exp. Pathol. 2018, 11, 5265–5277. [Google Scholar] [PubMed]
- He, J.; Chen, K.; Sui, Y.; Yang, Q. Effects of N-methyl-D-aspartate receptor knockdown and hypoxia/reoxygenation injury on the neuronal proteome and transcriptome. Front. Mol. Neurosci. 2022, 15, 1004375. [Google Scholar] [CrossRef]
- Ewida, H.A.; Zayed, R.K.; Darwish, H.A.; Shaheen, A.A. Circulating lncRNAs HIF1A-AS2 and LINLK-A: Role and Relation to Hypoxia-Inducible Factor-1α in Cerebral Stroke Patients. Mol. Neurobiol. 2021, 58, 4564–4574. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, X.; Cheng, R.; Hou, X.; Chen, Y.; Feng, Y.; Qiu, J. Analysis of long non-coding RNA expression profiles in neonatal rats with hypoxic-ischemic brain damage. J. Neurochem. 2019, 149, 346–361. [Google Scholar] [CrossRef]
- Wei, L.; Li, X.; Wang, L.; Song, Y.; Dong, H. Comprehensive Analysis of RNA Expression Profile Identifies Hub miRNA-circRNA Interaction Networks in the Hypoxic Ischemic Encephalopathy. Comput. Math. Methods Med. 2021, 2021, 6015473. [Google Scholar] [CrossRef]
- Jiang, L.; Li, H.; Fan, Z.; Zhao, R.; Xia, Z. Circular RNA expression profiles in neonatal rats following hypoxic-ischemic brain damage. Int. J. Mol. Med. 2019, 43, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhuang, S.; Huang, Y.; Yang, X.; Fu, Y.; Yu, L.; Zhao, Y. Expression profile of circular RNAs in the peripheral blood of neonates with hypoxic-ischemic encephalopathy. Mol. Med. Rep. 2020, 22, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Li, L.; Yu, B.; Jiao, L.; Han, Z.; Zhao, H.; Li, G.; Ma, Y.; Luo, Y. Circular RNA profiling of neutrophil transcriptome provides insights into asymptomatic Moyamoya disease. Brain Res. 2019, 1719, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.B.; Wu, Y.T.; Guo, X.X.; Xiang, C.; Chen, P.S.; Qin, W.; Shi, Z.S. Circular RNA hsa_circ_0007990 as a blood biomarker for unruptured intracranial aneurysm with aneurysm wall enhancement. Front. Immunol. 2022, 13, 1061592. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, W.; Gu, L.; Zhu, L.; Liang, T.; Liang, X.; Zhong, Q.; Zhang, R.; Nan, A.; Su, L. CircFOXP1 alleviates brain injury after acute ischemic stroke by regulating STAT3/apoptotic signaling. Transl. Res. J. Lab. Clin. Med. 2023, 257, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Ge, B.; Xu, S.; Chen, X.; Yang, H. Up-regulation of circARF3 reduces blood-brain barrier damage in rat subarachnoid hemorrhage model via miR-31-5p/MyD88/NF-kappaB axis. Aging 2021, 13, 21345–21363. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Dai, M.; Liu, X.; Wang, H.; Wang, C.; Fan, X.; Fang, W. Hsa_circ_0090002 regulates miR-186-5p/HECTD1 axis to mediate brain microvascular endothelial cell dysfunction. Brain Res. Bull. 2022, 178, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Jing, Y.X.; Zhou, Z.W.; Yang, J.W. Knockdown of circRNA-Memo1 Reduces Hypoxia/Reoxygenation Injury in Human Brain Endothelial Cells Through miRNA-17-5p/SOS1 Axis. Mol. Neurobiol. 2022, 59, 2085–2097. [Google Scholar] [CrossRef]
- Yang, X.; Li, X.; Zhong, C.; Peng, J.; Pang, J.; Peng, T.; Wan, W.; Li, X. Circular RNA circPHKA2 Relieves OGD-Induced Human Brain Microvascular Endothelial Cell Injuries through Competitively Binding miR-574-5p to Modulate SOD2. Oxidative Med. Cell. Longev. 2021, 2021, 3823122. [Google Scholar] [CrossRef]
- Jiang, S.; Zhao, G.; Lu, J.; Jiang, M.; Wu, Z.; Huang, Y.; Huang, J.; Shi, J.; Jin, J.; Xu, X.; et al. Silencing of circular RNA ANRIL attenuates oxygen-glucose deprivation and reoxygenation-induced injury in human brain microvascular endothelial cells by sponging miR-622. Biol. Res. 2020, 53, 27. [Google Scholar] [CrossRef]
- Bai, X.; Liu, X.; Wu, H.; Feng, J.; Chen, H.; Zhou, D. CircFUNDC1 knockdown alleviates oxygen-glucose deprivation-induced human brain microvascular endothelial cell injuries by inhibiting PTEN via miR-375. Neurosci. Lett. 2022, 770, 136381. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, H.; Zhang, J.; Dong, Y. Circular RNA cZNF292 silence alleviates OGD/R-induced injury through up-regulation of miR-22 in rat neural stem cells (NSCs). Artif. Cells Nanomed. Biotechnol. 2020, 48, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; He, W.; Xia, J.; Huang, Q.; Yang, J.; Gu, W.P.; Zhang, N.; Liu, Y.H. Human umbilical cord mesenchymal stem cells-derived exosomal circDLGAP4 promotes angiogenesis after cerebral ischemia-reperfusion injury by regulating miR-320/KLF5 axis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2023, 37, e22733. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.H.; Wang, M.; Lin, P.C. Gymnadenia conopsea (L.) R. Br.: Comprehensive review of propagation and breeding, traditional uses, chemical composition, pharmacology, quality control, and processing. J. Ethnopharmacol. 2023, 306, 116205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, L.; Liang, C.; Zhou, L.; Tan, L.; Zong, Y.; Wu, L.; Liu, T. Expression Profiles of Long Noncoding RNAs in Mice with High-Altitude Hypoxia-Induced Brain Injury Treated with Gymnadenia conopsea (L.) R. Br. Neuropsychiatr. Dis. Treat. 2020, 16, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Tasbihgou, S.R.; Barends, C.R.M.; Absalom, A.R. The role of dexmedetomidine in neurosurgery. Best Pract. Res. Clin. Anaesthesiol. 2021, 35, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y. Circular RNA cerebellar degeneration-related protein 1 antisense RNA (Circ-CDR1as) downregulation induced by dexmedetomidine treatment protects hippocampal neurons against hypoxia/reoxygenation injury through the microRNA-28-3p (miR-28-3p)/tumor necrosis factor receptor-associated factor-3 (TRAF3) axis. Bioengineered 2021, 12, 10512–10524. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Studies that Summarized the Involvement of LncRNAs SNHG Family in Hypoxic/Anoxic Conditions | |||
| SH-SY5Y OGD/R-induced | SNHG1 | The overexpression of SNHG1 or the KO of miR-140-5p alleviated hypoxia-induced damage. Overall, the results proved that lncRNA SNHG1 could be novel therapeutic target in hypoxic brain injury | [63] |
| Primary BMEC mice OGD/R-induced | SNHG1 | Snhg1 silencing led to worsening the BMEC apoptosis that was OGD/R-induced, so confirmed the protective effect of lncRNA Snhg1in in IS model | [64] |
| C57Bl/6 mice with ligation of the right internal carotid artery. Primary hippocampal cell culture with shRNA or miRNA mimic transfection | SNHG3 | Snhg3 silencing led to reduction in cell viability and promoted the apoptosis, in vivo Snhg3 overexpression improved brain neurological function in the animals and reduced HIBD | [65] |
| OGD/R-induced primary neurons | SNHG12 | SNHG12 knockdown exacerbated apoptosis induced by OGD/R | [66] |
| HT22 cells of I/R model | SNHG12 | SNHG12 increased cell activity and inhibited oxidative stress through inhibition of SIRT1/FOXO3a signaling-mediated autophagy | [67] |
| N2A cells ODG/R-induced MCAO/R mice | SNHG14 | SNHG14 silencing led to an increase in miR-98-5p and BCL2L13 depletion. So, SNHG14 is beneficial to OGD/R | [68] |
| MCAO male SD rats OGD-induced primary cortical neurons of SD embryos rat | SNHG14 | SNHG14 silencing led to a reduction in the infarct volume and improved neurological deficits in MCAO rats | [69] |
| H/I mice model PC12 cells OGD-stimulated | SNHG15 | SNHG15 suppression alleviated H/I brain injury by modulating the miR-153-3p/SETD7 axis | [70] |
| HBMECs OGD/R-induced | SNHG16 | The overexpression of SNHG16 led to a decrease in miR-15a-5p and increased the Bcl-2 expression levels, as well as reduced apoptosis and enhanced cell survival | [71] |
| Studies that Summarized the Involvement of LncRNA MALAT1 In Hypoxic/Anoxic Conditions | |||
| C57BL/6 mice with right common carotid artery occlusion C57BL/6 mice hippocampal neurons with H/I | MALAT1 | MALAT1 acting as a miR-429 sponge led to an increment in WNT1 expression promoting apoptosis, as shown by expression levels of cleaved caspase-3 and the ratio of Bax to Bcl-2 both in vitro and vivo | [72] |
| HBMECs OGD-induced | MALAT1 | Through sponging miR-126, MALAT1 inhibited proliferation and induced apoptosis of OGD-induced HBMECs | [73] |
| bEnd.3 OGD-induced | MALAT1 | The MALAT1-miR-200c-3p-SIRT1 pathway could be interesting to modulate for influencing autophagy and cell survival for neuroprotection | [74] |
| BMECs of C57BL/6J mice OGD/R-induced | MALAT1 | MALAT1 increased autophagy and cell survival in BMECs | [75] |
| MCAO mice BMECs OGD-induced | MALAT1 | MALAT1 interactions reduced cell death and inflammation | [76] |
| MCAO C57BL/6J mice BMECs OGD/R-induced | MALAT1 | Following OGD/R, MALAT1 regulated angiogenesis | [77] |
| Studies that Summarized the Involvement of LncRNA H19 In Hypoxic/Anoxic Conditions | |||
| Neonatal HIE SD rats | H19 | H19 overexpression reduced both the nervous damage and autophagy of brain tissue, as well as improved neurological functions | [78] |
| Primary neuron cells rat OGD-induced HIBD SD rat induced by partial occlusion of carotid artery | H19 | HIBD can be prevented by overexpression of the lncRNA H19 | [79] |
| HIBD neonatal rats | H19 | H19 overexpression suppressed apoptosis in rat cardiomyocytes | [80] |
| Blood samples of IS patients MCAO/R rats SH-SY5Y cells OGD-induced | H19 | Hypoxia/ischemia-induced neuronal injury was attenuated by blocking the lncRNA H19-miR-19a-Id2 axis | [81] |
| PC-12 cells induced by hypoxia | H19 | Overexpression of lncRNA H19 led to an increase in cell damage induced by hypoxic conditions | [82] |
| IS patients MCAO mouse BV2 cell OGD-induced | H19 | H19 could be a useful marker as well as a potential therapeutic target for IS patients | [83] |
| Studies that Summarized the Involvement of LncRNA GAS5 in Hypoxic/Anoxic Conditions | |||
| Blood of CA/CCR patients OGD/R-inducted astrocytes | GAS5 | Inhibiting PI3K/Akt signaling through INPP4B, lncRNAGAS5/miR-137 is a hypoxia-responsive axis involved in astrocyte–microglia crosstalk. | [84] |
| HIBD neonatal rat model established and treated with shRNA-GAS5 or antagomir miR-128-3p | GAS5 | Through the microRNA-128-3p/Bax/Akt/GSK-3β axis, downregulation of GAS5 prevents mitochondrial apoptosis and HIBD in neonatal rats | [85] |
| Primary rat cortical cell hypoxia-treated B35 cell with hypoxic treated MCAO SD rats | GAS5 | GAS5 can lead to worsening of cell apoptotic processes in hypoxic conditions targeting miR-221/PUMA axis. Consequently, GAS5 could be useful in ischemic stroke treatment | [86] |
| Studies that Summarized the Involvement of LncRNA NEAT1 in Hypoxic/Anoxic Conditions | |||
| BMECs OGD-induced | NEAT1 | NEAT1 facilitates survival and angiogenesis in OGD-induced BMECs via targeting miR-377 | [87] |
| Neonatal HIBD mouse model was established via right common carotid artery occlusion | NEAT1 | The findings demonstrated that NEAT1 alleviated HIBD in mice by binding to miR-339-5p | [88] |
| Studies that Summarized the Involvement of LncRNA MIAT in Hypoxic/Anoxic Conditions | |||
| Neuro2A cells OGD-induced Neonatal rat with permanent unilateral carotid ligation | MIAT | The overexpression of MIAT reduced neuron apoptosis and relieved HI injury of neonatal rats through miR-211/GDNF | [89] |
| MCAO SD rats Primary rat neurons OGD/R- stimulated | MIAT | MIAT exacerbated I/R injury by disrupting redox homeostasis in neurons | [90] |
| Studies that Summarized the Involvement of LncRNA PVT1 in Hypoxic/Anoxic Conditions | |||
| MCAO mice Primary neurons OGD/R-induced | PVT1 | PVT1 silencing led to a reduction in infarct volume and improved neurological behavior in MCAO mice | [91] |
| HUVECs OGD-induced | PVT1 | PVT1 alleviated hypoxia-induced endothelial apoptosis by enhancing autophagy through the miR-15b-5p/ATG14 and miR-424-5p/ATG14 pathways | [92] |
| Studies that Summarized the Involvement of LncRNA Peg13 in Hypoxic/Anoxic Conditions | |||
| Neonatal HIBD mice Mouse hippocampal neurons OGD-induced | PEG13 | Peg13 exerted an anti-apoptotic role, acting as a sponge for miR-20a-5p targeting XIAP | [93] |
| bEnd.3 cells OGD/R-induced MCAO mice | PEG13 | Peg13 alleviated I/R-induced neurological deficit, cerebral infarct, and cerebral edema | [94] |
| Studies that Summarized the Involvement of LncRNA MEG3 in Hypoxic/Anoxic Conditions | |||
| PC12 cells induced by hypoxia | MEG3 | MEG3 silencing performed by siRNA played a neuroprotective function, preventing hypoxia-induced injury in PC12 via the modulation of protein involved in apoptosis and cell proliferation | [95] |
| MCAO C57BL/6J mice | MEG3 | Silencing MEG3 resulted in being protective against I/R-induced ischemic damage in vivo and improved overall neurological functions | [96] |
| PC12 cells cultivated in hypoxic conditions | MEG3 | MEG3 led to an increase in hypoxic damage in PC12 cells by targeting miR147 and the downstream target Sox2 | [97] |
| 260 neonatal female or male C57/BL6 mice with right cervical vessels ligated with a double bandage | MEG3 | MEG3 silencing improved the therapeutic effect of DEX on HIBD in neonatal mice through the expression of miR-129-5p | [98] |
| Studies that Summarized the Involvement of LncRNA BDNF-AS in Hypoxic/Anoxic Conditions | |||
| Mice received ligation of the unilateral common carotid artery Primary hippocampal neuron with hypoxic and oxidative stress induced | BDNF-AS | Silenced BDNF-AS lncRNA attenuated HI events | [99] |
| HCN2 and human astrocytes after H/R | BDNF-AS | lncRNA BDNF-AS knockdown suppressed apoptosis | [100] |
| Models | LncRNAs Involved | Conclusions | Ref. |
|---|---|---|---|
| Neuronal cells OGD-induced Blood of 44 IS patients and 37 healthy controls | LOC105376244 | LOC105376244 results in being dysregulated in IS patients and it is in turn regulated by miR-145–EPHA4 interaction | [113] |
| Neuronal cells OGD/R-stimulated Mouse brains subjected to transient focal ischemia | AK038897 | AK038897 plays a role in cerebral I/R injury | [114] |
| PC12 cells OGD/R-stimulated Plasma of patients with cerebral ischemia reperfusion | KCNQ1OT1 | Upregulated KCNQ1OT1 aggravates cerebral I/R injury | [115] |
| Plasma of neonatal HIE patients hBMVECs OGD/R-induced | HOTAIR | LncRNA HOTAIR modulating EZH2, led to apoptosis of hBMVECs | [116] |
| HIBD rat model Primary cortical neurons of rats OGD-stimulated | CRNDE | The silencing of CRNDE alleviated brain injury after HI and inhibited neuronal apoptosis, both by promoting autophagy | [117] |
| H/R SH-SY5Y cells MCAO rat model | PINK1-AS | PINK1-AS/miR-203/ATF2 axis is involved in NOX2 expression regulation | [118] |
| N2a cells OGD-stimulated | Gm11974 | Gm11974 suppression provides protection against cerebral I/R injury, suggesting that this protective effect is mediated by the miR-766-3p/NR3C2 axis | [119] |
| N2a cells OGD/R-exposed | RMST | RMST decrement causes the abrogation of the OGD/R-triggered apoptosis and oxidative stress | [120] |
| HIE neonatal SD male rats | Vof-16 | Vof-16 KO improves neurological damage as well as spatial learning and memory functions in HIE rats | [121] |
| MCAO mice BV-2 cells were exposed to high glucose followed by H/R | Fendrr | Fendrr is able to protect against the ubiquitination and degradation of NLRC4 protein through E3 ubiquitin ligase HERC2, thereby accelerating the pyroptosis of microglia | [122] |
| Models | LncRNAs Involved | Conclusions | Ref. |
|---|---|---|---|
| MCAO mice Microglial cell OGD-induced | 1810034E14Rik | The overexpression of lncRNA1810034E14Rik leads to a reduction in inflammatory cytokine levels and brain damage | [123] |
| OGD/R-induced HBMVECs | FAL1 | Overexpressed FAL1 reduces inflammation | [124] |
| MCAO mice OGD/R-induced HBMECs | ROR | Upregulation in the expression of ROR inhibits the activation of TNF-α/P-ASK-1-mediated apoptosis, which ultimately alleviates hypoxia in brain tissue | [125] |
| HIE SD neonatal rats | Vi4 | LncRNA Vi4, by miR-185-5p modulation, leads to an improvement in motor functions, learning. and memory deficits | [126] |
| HIE whole blood (8 infants) OGD/R-induced SH-SY5Y | LINC00938 | LINC00938 contributes to the promotion of neuronal cell survival. It achieves this by inhibiting the activation of p-JNK and p-p38 MAPK signaling pathways | [127] |
| HIE neonate rats | TCONS_00041002 | TCONS_00041002 participates in the cell apoptosis and neuronal survival of HIE and represents a potential new target for the treatment of HIE | [128] |
| PC12 cells OGD-induced | ANRIL | ANRIL is able to reduce HIBD by targeting the miR-378b/ATG3 axis | [129] |
| Serum of stroke patients MCAO/R rats OGD/R-induced microglial cells | OIP5-AS1 | lncRNA OIP5-AS upregulation exerts neuroprotective effects | [130] |
| MCAO C57BL/6 mice Primary cortical neurons OGD-induced | HOTTIP | Overexpression of HOTTIP leads to an increase in cell viability and reduces apoptosis, as well as increasing and improving glycolytic metabolism via the upregulation of HK-2 expression levels, leading to the reduction in OGD-induced neuronal injury | [131] |
| HBMECs induced by hypoxia | XIST | The study results suggest that XIST involving miR-485-3p/SOX7 axis exerts an important role in maintaining angiogenesis processes after hypoxia | [132] |
| HBMECs induced by hypoxia | MACC1-AS1 | The study found that lncRNA MACC1-AS1, by modulating the miR-6867-5p/TWIST1 axis, could help to discover emerging therapies useful for IS | [133] |
| SH-SY5Y cells and primary murine neurons both OGD/R stimulated | D63785 | The silencing of lnc-D63785 leads to an increase in Mir-422a, thus reducing apoptosis | [134] |
| Model | Compound | LncRNAs Involved | Results | Ref. |
|---|---|---|---|---|
| MCAO rats OGD-treated HT22 hippocampal neurons | Rats DEX (25 μmol/kg, 50 μmol/kg, 100 μmol/kg body weight) injected via external jugular vein and HT22 neuronal cells treated with DEX (25 μM, 50 μM, and 100 μM) for 24 h | SNHG16 | DEX attenuates neurological damage, increasing neurons’ viability and promoting lncRNA SNHG16 and BDNF levels, as well as reducing miR-10b-5p expression | [136] |
| SH-SY5Y cells in hypoxia condition | Berberine at a doses of 10, 20, and 50 μM | H19, HOTAIR, CASC2 and LINC00943 | Berberine treatment reduces IS in in vitro model, through inhibition of lncRNA H19/EGFR/JNK1/c-Jun axis | [137] |
| C57BL/6 J mice with endovascular perforation | MT at a dose of 150 mg/kg via intraperitoneally 12 h after surgery | H19 | MT administration improves neurobehavioral deficits and reduces both apoptosis and inflammation levels, leading to an increase in H19 and miR-675, as well as a reduction in HIF-1α and TLR4 | [138] |
| Pregnant SD rats and male pups incised the right common carotid artery and exposed to hypoxia | Nicotine at a dose of 102 mg/mL infused subcutaneously by osmotic mini-pumps | H19 | Nicotine exposure leads to an increase in H19 and miR-181a, as well as a decrease in ATG5 and thus the possible onset of the brain phenotype sensitive to neonatal H/I, while miR-181a block induces a reduction in H/I induced by perinatal nicotine exposure | [139] |
| OGD-induced bEnd.3 cells | EGb761 at a dose of 100 μg/mL 2 h before OGD exposure | RMST | EGb761 pretreatment leads to apoptosis reduction as well as an increase in lncRNA RMST and miR-150 | [140] |
| PC12 cells treated with CoCl2 | β-Asarone (20, 30, and 45 μg/mL) | RPPH1 | β-Asarone exerts neuroprotective effects, reducing hypoxia-induced damage, by suppressing RPPH1/miR-542-3p/DEDD2 axis | [141] |
| MCAO rats OGD/R rat primary cortical neurons | GAS 50 mg/kg | NEAT1 | GAS attenuates I/R-induced inflammation in neuronal cells via lncRNA NEAT1/miR-22-3p axis | [142] |
| MCAO/R Kunming mice PC12 cells H/R-induced | PC12 cells pre-treated with PPF 50 μM 1 h before H/R induction and Rats infused intravenously with PPF (60 mg/kg, 10 min before MCAO | MALAT1 | PPF exerts neuroprotective effects thanks to increments in MALAT1 and miR-182-5p, as well as a reduction in TLR4 expression | [143] |
| Models | LncRNAs Involved | Conclusions | Ref. |
|---|---|---|---|
| Neonatal rats with the right common carotid artery permanently ligated Primary OPC OGD-induced | ↑106 ↓104 | The study suggests the contribution of lncRNAs to the development of preterm WMI | [144] |
| BEnd.3 of mouse | ↑18 ↓15 | Multiple lncRNAs exhibit altered expression, with a focus on lncRNA Gpr137b-ps being a potential candidate to promote angiogenesis after I/R | [145] |
| Neonatal rats with the left common carotid artery ligated and transferred in a low-oxygen chamber | 617 aberrantly expressed | HI injury alters the expression profiles of lncRNAs in neonatal rat brains | [146] |
| Mouse hippocampal neuron HT22 cell line after NMDAR knockdown and H/R | ↑XLOC_161072, ↑XLOC_065271, ↓XLOC_159404, ↓XLOC_031922 | lncRNAs respond to hypoxic stimulation | [147] |
| Serum of stroke patients | ↑HIF1A-AS2; ↓LINK-A | HIF1A-AS2 and LINK-A as diagnostic and prognostic markers | [148] |
| HIBD neonatal rats | ↑151 ↓177 | Several lncRNAs can be useful for therapeutic research in neonatal HIBD | [149] |
| Models | circRNA Involved | Compound | Results | Ref. |
|---|---|---|---|---|
| Two RNA-seq datasets (GSE164727 and GSE121178) | 2432 | - | circRNAs may play a role in HIE development through circRNA/miRNA interactions | [150] |
| Rats with the right common carotid artery permanently double ligated and sliced from the middle | ↑20 ↓46 | - | Potential biomarkers or novel therapeutic targets in the treatment of HIBD | [151] |
| 10 peripheral blood samples (6 mL) from neonates with HIE | ↑250 ↓206 | - | Potential relationship between circRNAs and HIE in neonates | [152] |
| 5 MMD patients | ↑54 ↓69 | - | Moreover, this study suggest that angiogenesis was involved in asymptomatic MMD | [153] |
| 8 patients harboring saccular UIAs with HR-VWI and 5 controls | 412 differentially expressed; RNA hsa_circ_0007990 | - | Biomarkers for detecting blood inflammation in people with UIAs accompanied by an AWE | [154] |
| OGD/R-induced A172 cells tMCAO model | CircFOXP1 | - | circFOXP1 reduced apoptosis by affecting Bax and caspase-3 expression. | [155] |
| BMECs OGD/R-induced Rats with proximal end of the external carotid artery ligated | CircARF3 | - | Brain hypoxia-induced damage reduction via miR-31-5p/MyD88/NF-κB axis | [156] |
| OGD/R-induced HBMECs | Circ0090002 | - | Brain hypoxia-induced damage reduction via circ_0090002/miR-186-5p/HECTD1 | [157] |
| Blood samples collected from 25 IS patients and 25 healthy controls+ HBMVECs | Memo1 | - | Circ-Memo1 silencing promotes cell viability, inhibiting the activation of ERK/NF-κB signaling pathway; reducing oxidative stress and inflammatory response; and inhibiting cell apoptosis | [158] |
| OGD/R-induced HBMECs | CircPHKA2 | - | circPHKA2 protects HBMEC from OGD-induced neurovascular injuries via circPHKA2/miR-574-5p/SOD2 axis | [159] |
| OGD/R-induced HBMECs | CircANRIL | - | Knockdown of circRNA_ANRIL improves OGD/R-induced cell damage, apoptosis, and inflammatory responses via circRNA /miR-622/NF-κB | [160] |
| OGD-induced HBMECs peripheral blood of IS patients | CircFUNDC1 | - | CircFUNDC1 knockdown alleviates OGD-induced HBEMCs injuries by targeting the miR-375/PTEN axis. | [161] |
| NSCs of rat treated with OGD/R | cZNF292 | - | cZNF292 silencing alleviated OGD/R-induced injury through the upregulation of miR-22 | [162] |
| H/R-treated HBMECs; MCAO rats | CircDLGAP4 | - | Potential key factor in CI/RI treatment | [163] |
| Mice with high-altitude hypoxia-induced brain injury | 23 differentially expressed | Gymnadenia conopsea (L.) R. Br. | Potential biomarkers or targets | [165] |
| HT-22 cells H/R-induced | CircCDR1as | DEX | Circ-CDR1as is downregulated after DEX treatment | [167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anchesi, I.; Schepici, G.; Mazzon, E. LncRNAs and CircRNAs as Strategies against Pathological Conditions Caused by a Hypoxic/Anoxic State. Biomolecules 2023, 13, 1622. https://doi.org/10.3390/biom13111622
Anchesi I, Schepici G, Mazzon E. LncRNAs and CircRNAs as Strategies against Pathological Conditions Caused by a Hypoxic/Anoxic State. Biomolecules. 2023; 13(11):1622. https://doi.org/10.3390/biom13111622
Chicago/Turabian StyleAnchesi, Ivan, Giovanni Schepici, and Emanuela Mazzon. 2023. "LncRNAs and CircRNAs as Strategies against Pathological Conditions Caused by a Hypoxic/Anoxic State" Biomolecules 13, no. 11: 1622. https://doi.org/10.3390/biom13111622
APA StyleAnchesi, I., Schepici, G., & Mazzon, E. (2023). LncRNAs and CircRNAs as Strategies against Pathological Conditions Caused by a Hypoxic/Anoxic State. Biomolecules, 13(11), 1622. https://doi.org/10.3390/biom13111622