
Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells


Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Culture
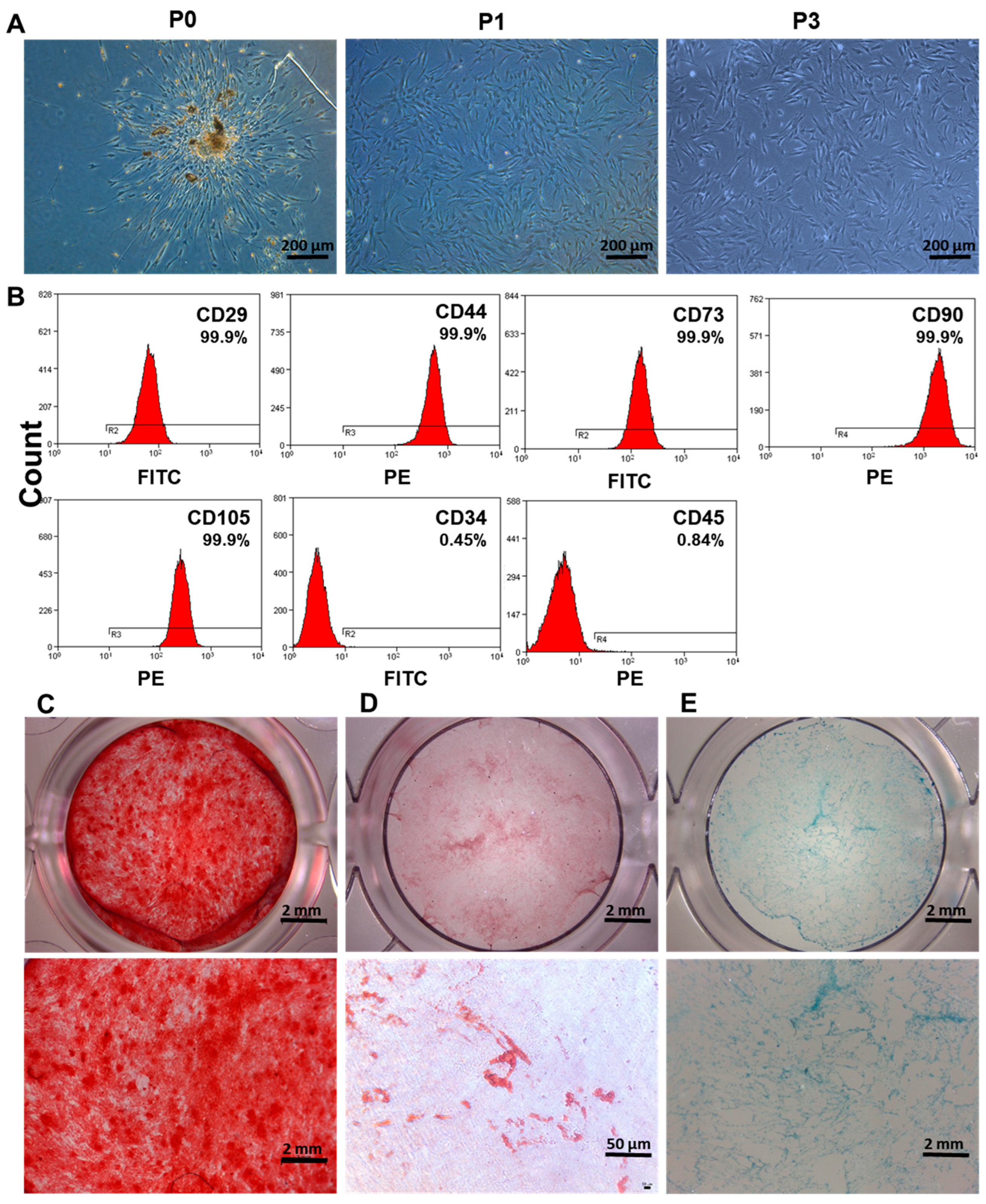
2.2. Identification of Surface Markers by Flow Cytometry
2.3. Multilineage Differentiation of DPSCs
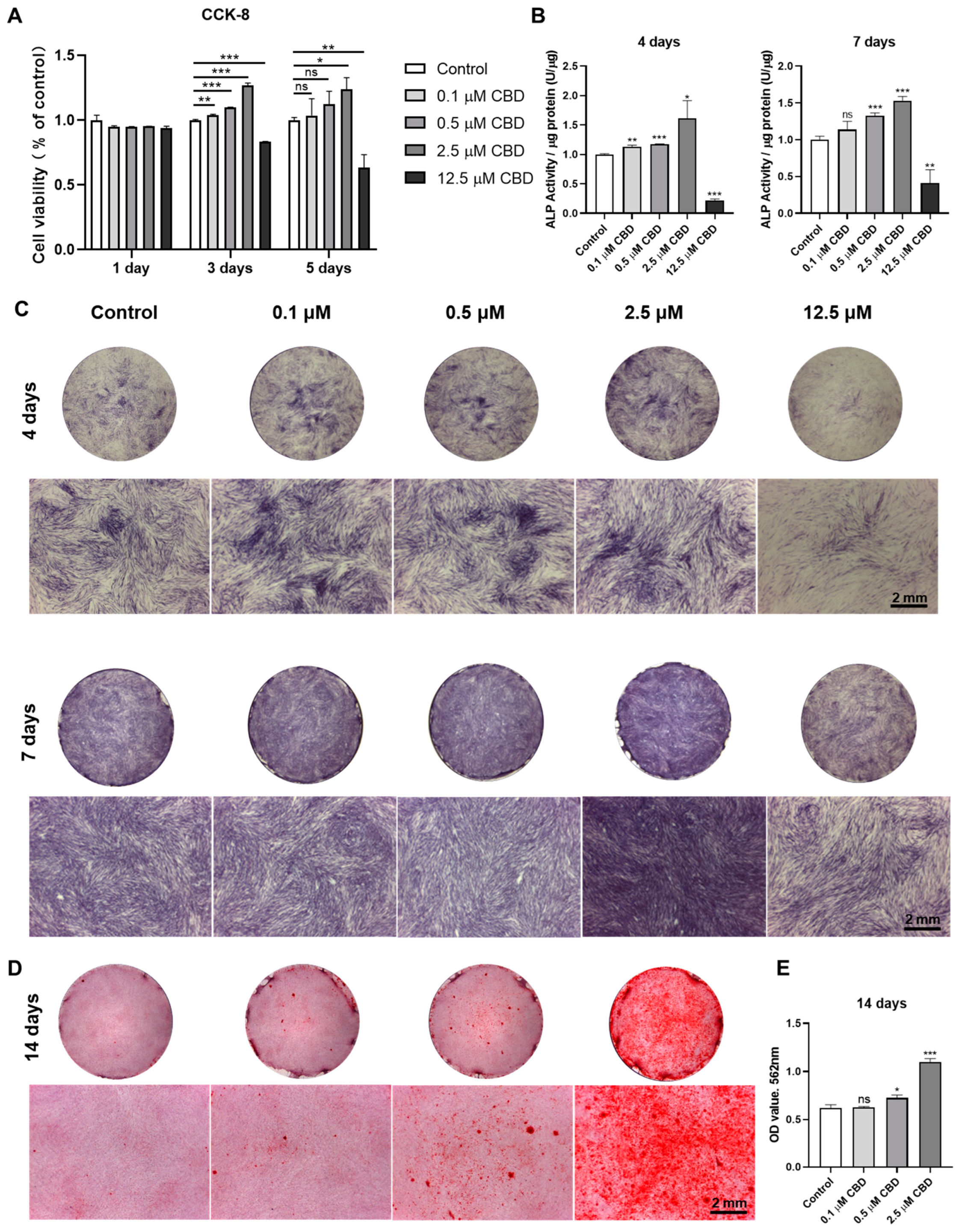
2.4. Cell Counting Kit-8 (CCK-8) Assay
2.5. Alkaline Phosphatase (ALP) Staining and Activity
2.6. Alizarin Red S (ARS) Staining
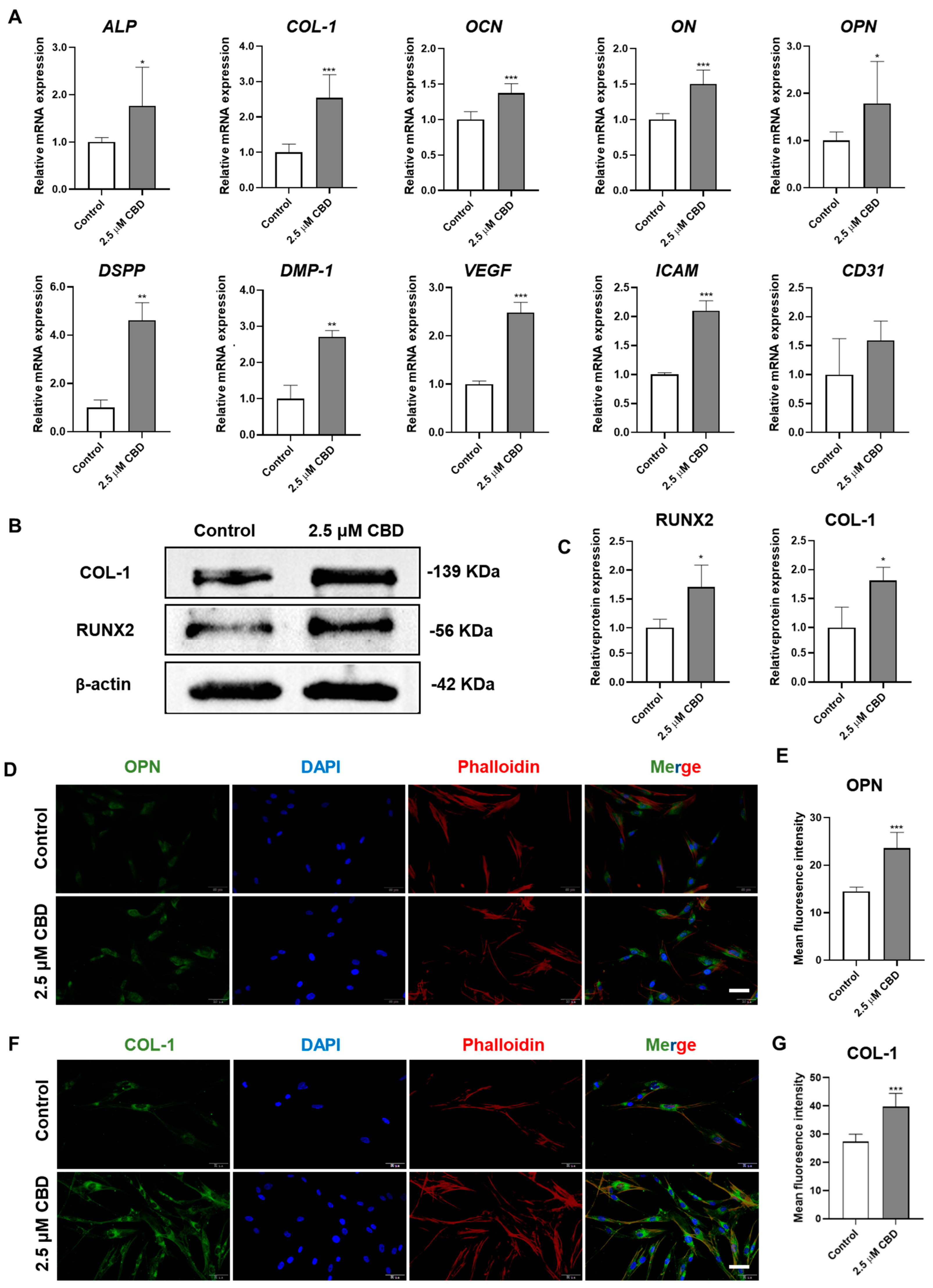
2.7. RNA Isolation and Real-Time Quantitative PCR (RT-qPCR) Analysis
2.8. Western Blot Assay
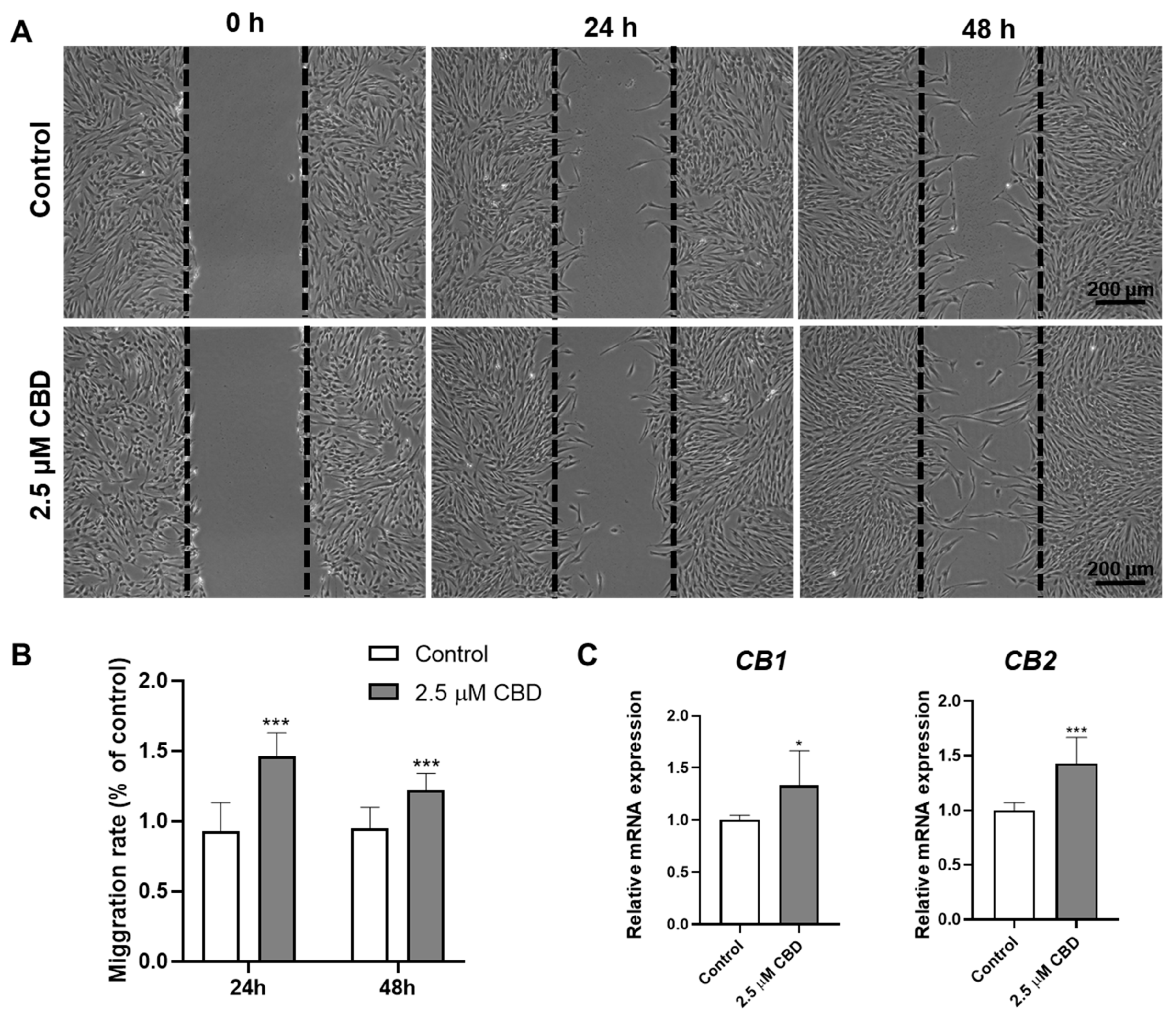
2.9. Scratch Wound Healing Assay
2.10. Immunofluorescence Staining
2.11. Statistical Analysis
3. Results
3.1. Isolation and Characterization of DPSCs
3.2. CBD-Induced Proliferation and Osteogenic/Odontogenic Differentiation of DPSCs
3.3. CBD Upregulated the Expression of Osteogenic/Odontogenic and Angiogenic Markers in DPSCs
3.4. CBD Promoted DPSCs Migration and Upregulated the Expression of CBD Receptors CB1 and CB2 in DPSCs
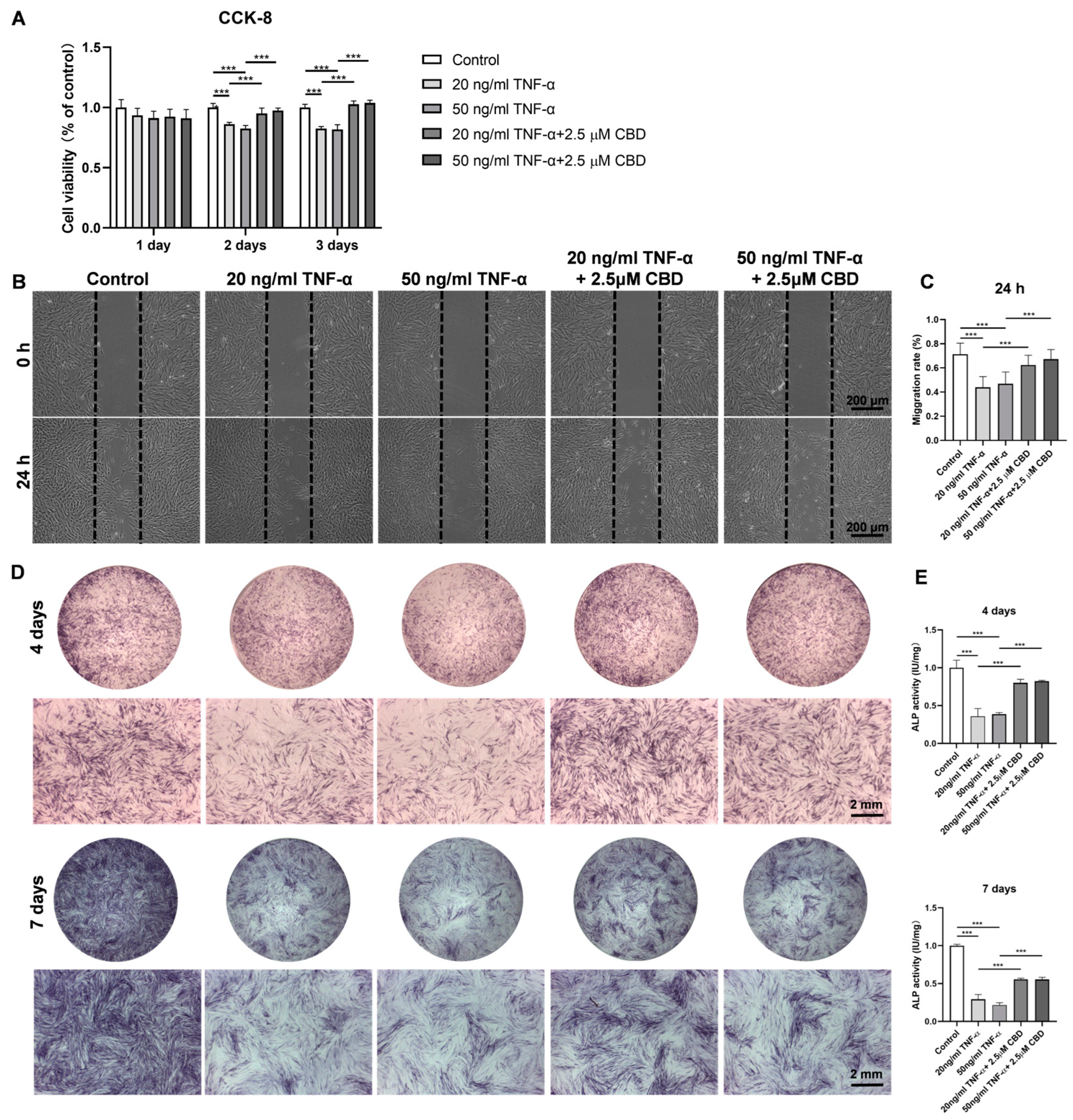
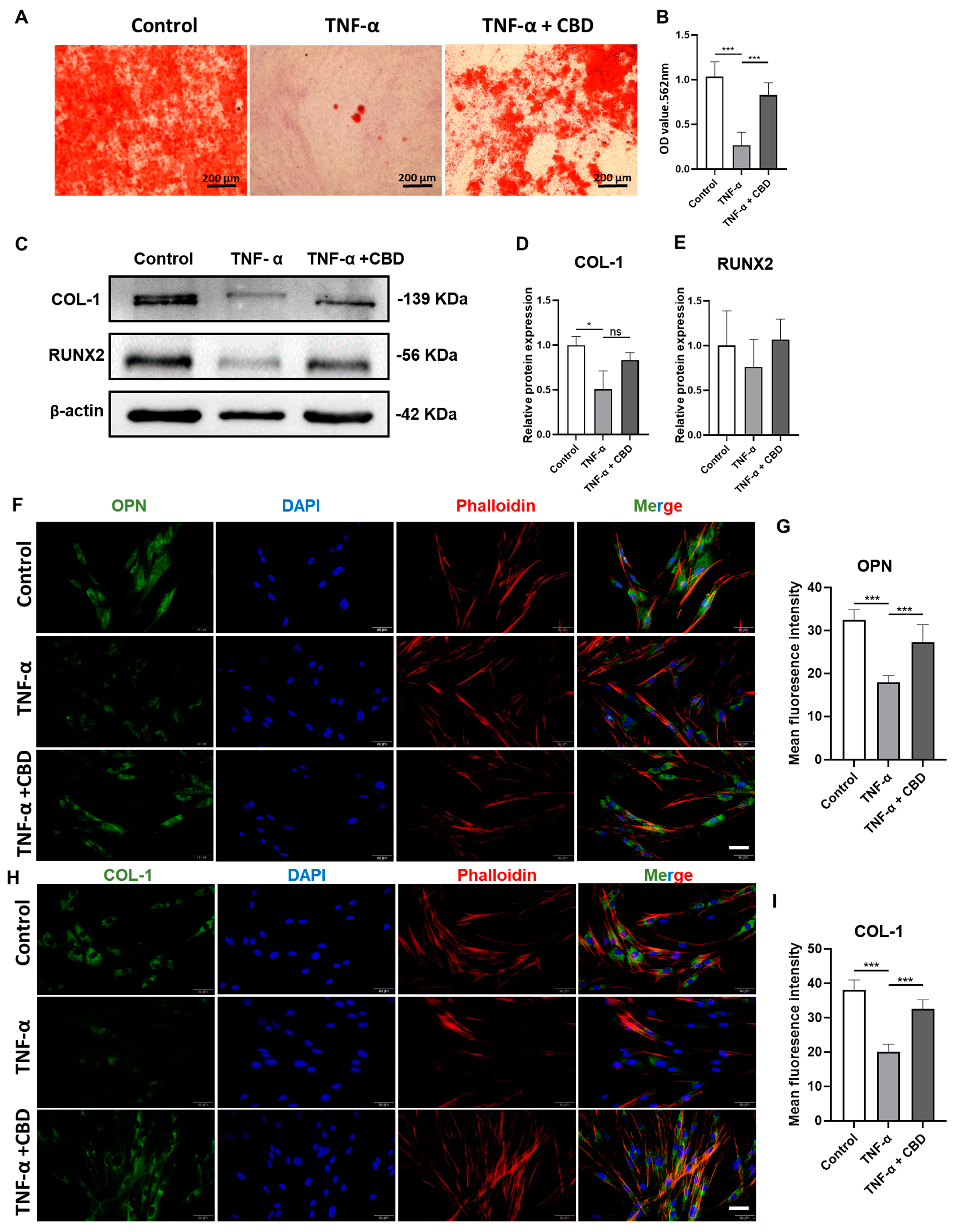
3.5. CBD Alleviated the TNF-α-Mediated Inhibitory Effect in DPSCs Viability, Migration, and Osteogenic/Odontogenic Differentiation
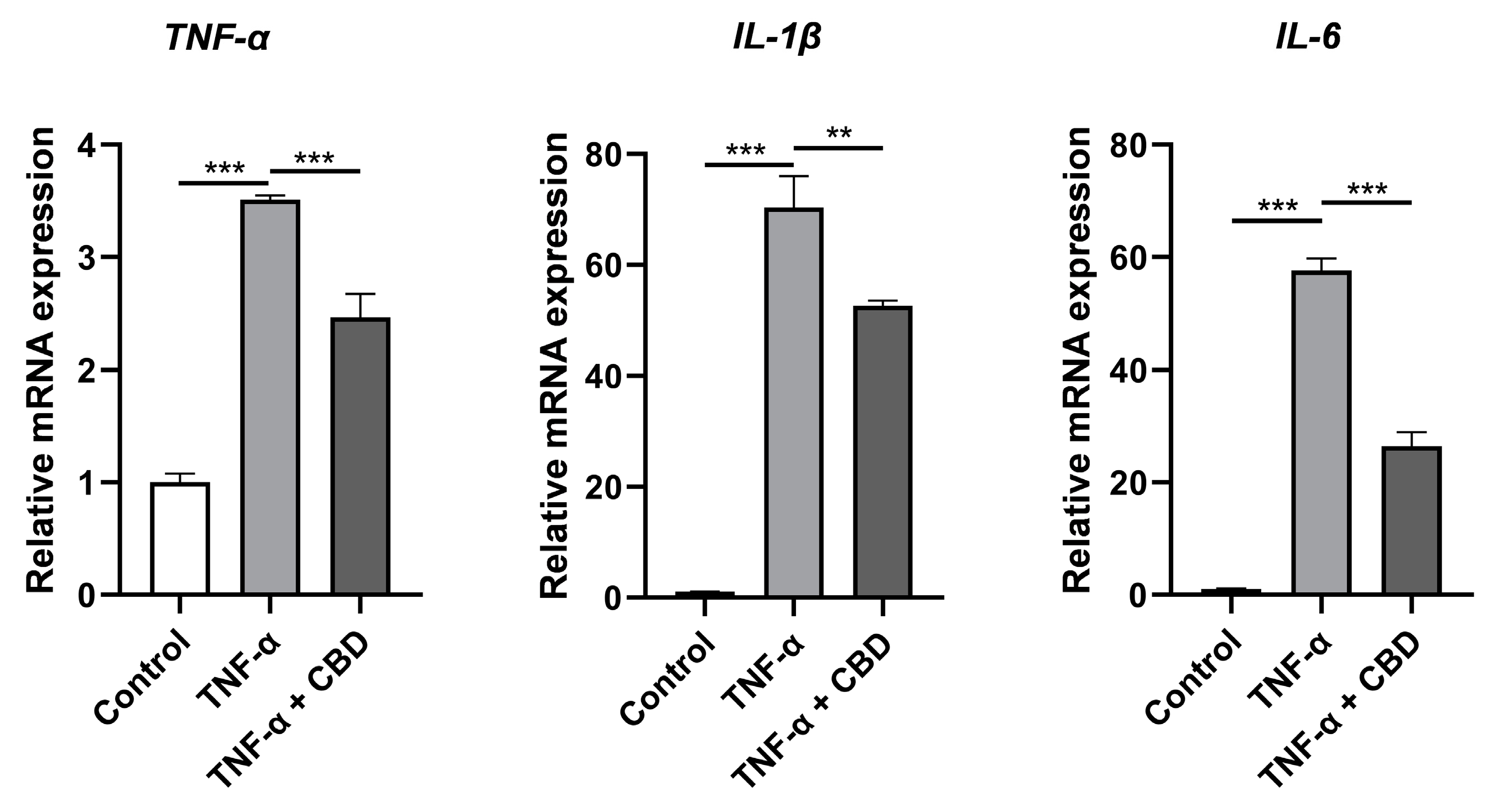
3.6. CBD Attenuated TNF-α-Induced Expression of Inflammatory Markers in DPSCs
4. Discussion
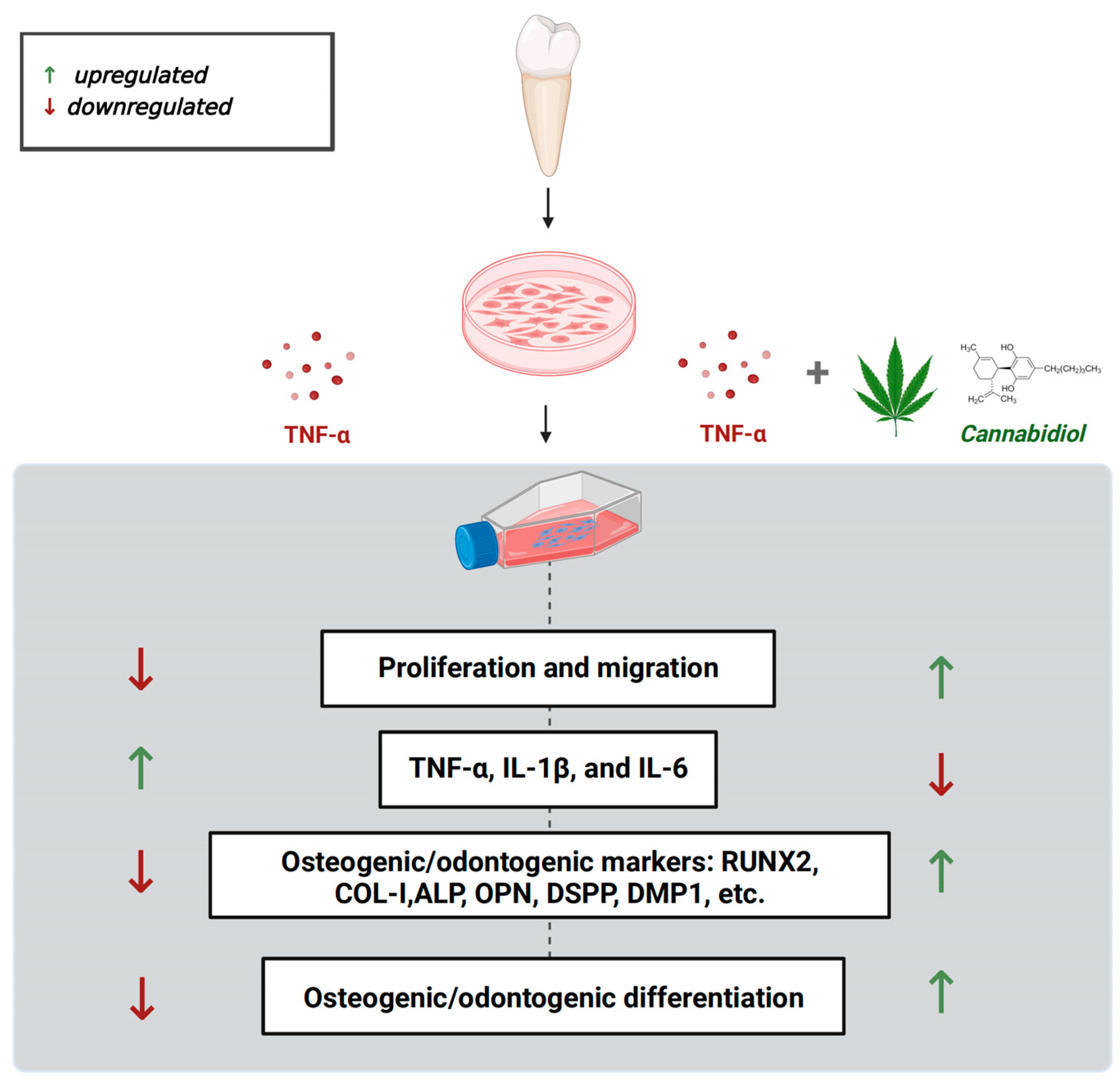
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haapasalo, M. Black-pigmented gram-negative anaerobes in endodontic infections. FEMS Immunol. Med. Microbiol. 1993, 6, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.L.; Liewehr, F.R. Innate immune responses of the dental pulp to caries. J. Endod. 2007, 33, 643–651. [Google Scholar] [CrossRef]
- Xuan, K.; Li, B.; Guo, H.; Sun, W.; Kou, X.; He, X.; Zhang, Y.; Sun, J.; Liu, A.; Liao, L.; et al. Deciduous autologous tooth stem cells regenerate dental pulp after implantation into injured teeth. Sci. Transl. Med. 2018, 10, eaaf3227. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.; Chen, C.; Kou, X.; Li, B.; Xuan, K.; Shi, S.; Jin, Y. Pulp Stem Cell-Mediated Functional Pulp Regeneration. J. Dent. Res. 2018, 98, 27–35. [Google Scholar] [CrossRef]
- Rechenberg, D.K.; Galicia, J.C.; Peters, O.A. Biological Markers for Pulpal Inflammation: A Systematic Review. PLoS ONE 2016, 11, e0167289. [Google Scholar] [CrossRef]
- Bender, I.B. Pulpal pain diagnosis—A review. J. Endod. 2000, 26, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.L.; Mann, V.; Rahbaran, S.; Lewsey, J.; Gulabivala, K. Outcome of primary root canal treatment: Systematic review of the literature—Part 2. Influence of clinical factors. Int. Endod. J. 2008, 41, 6–31. [Google Scholar] [CrossRef]
- Su, Y.; Wang, C.; Ye, L. Healing rate and post-obturation pain of single- versus multiple-visit endodontic treatment for infected root canals: A systematic review. J. Endod. 2011, 37, 125–132. [Google Scholar] [CrossRef]
- Duggal, M.; Tong, H.J.; Alansary, M.; Twati, W.; Day, P.F.; Nazzal, H. Interventions for the endodontic management of non-vital traumatised immature permanent anterior teeth in children and adolescents: A systematic review of the evidence and guidelines of the European Academy of Paediatric Dentistry. Eur. Arch. Paediatr. Dent. 2017, 18, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Heng, B.C.; Lo, E.C.M.; Zhang, C. Current Advance and Future Prospects of Tissue Engineering Approach to Dentin/Pulp Regenerative Therapy. Stem Cells Int. 2016, 2016, 9204574. [Google Scholar] [CrossRef]
- Lim, W.L.; Liau, L.L.; Ng, M.H.; Chowdhury, S.R.; Law, J.X. Current Progress in Tendon and Ligament Tissue Engineering. Tissue Eng. Regen. Med. 2019, 16, 549–571. [Google Scholar] [CrossRef]
- Xie, Q.; Liu, R.; Jiang, J.; Peng, J.; Yang, C.; Zhang, W.; Wang, S.; Song, J. What is the impact of human umbilical cord mesenchymal stem cell transplantation on clinical treatment? Stem Cell Res. Ther. 2020, 11, 519. [Google Scholar] [CrossRef]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef]
- Lee, M.; Jeong, S.Y.; Ha, J.; Kim, M.; Jin, H.J.; Kwon, S.-J.; Chang, J.W.; Choi, S.J.; Oh, W.; Yang, Y.S.; et al. Low immunogenicity of allogeneic human umbilical cord blood-derived mesenchymal stem cells in vitro and in vivo. Biochem. Biophys. Res. Commun. 2014, 446, 983–989. [Google Scholar] [CrossRef]
- Achilleos, A.; Trainor, P.A. Neural crest stem cells: Discovery, properties and potential for therapy. Cell Res. 2012, 22, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Alge, D.L.; Zhou, D.; Adams, L.L.; Wyss, B.K.; Shadday, M.D.; Woods, E.J.; Chu, T.M.G.; Goebel, W.S. Donor-matched comparison of dental pulp stem cells and bone marrow-derived mesenchymal stem cells in a rat model. J. Tissue Eng. Regen. Med. 2009, 4, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, C.; Kimura, K.; Nakayama, T.; Terashita, M. Temporal and spatial expression of c-jun and jun-B proto-oncogenes in pulp cells involved with reparative dentinogenesis after cavity preparation of rat molars. J. Dent. Res. 1999, 78, 673–680. [Google Scholar] [CrossRef]
- Ratajczak, J.; Bronckaers, A.; Dillen, Y.; Gervois, P.; Vangansewinkel, T.; Driesen, R.B.; Wolfs, E.; Lambrichts, I.; Hilkens, P. The Neurovascular Properties of Dental Stem Cells and Their Importance in Dental Tissue Engineering. Stem Cells Int. 2016, 2016, 9762871. [Google Scholar] [CrossRef]
- Suchanek, J.; Soukup, T.; Visek, B.; Ivancakova, R.K.; Kucerova, L.; Mokry, J. Dental pulp stem cells and their characterization. Biomed. Pap. 2009, 153, 31–35. [Google Scholar] [CrossRef]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef]
- Martellucci, S.; Manganelli, V.; Santacroce, C.; Santilli, F.; Piccoli, L.; Sorice, M.; Mattei, V. Role of Prion protein-EGFR multimolecular complex during neuronal differentiation of human dental pulp-derived stem cells. Prion 2018, 12, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Mata, M.; Milian, L.; Oliver, M.; Zurriaga, J.; Sancho-Tello, M.; de Llano, J.J.M.; Carda, C. In Vivo Articular Cartilage Regeneration Using Human Dental Pulp Stem Cells Cultured in an Alginate Scaffold: A Preliminary Study. Stem Cells Int. 2017, 2017, 8309256. [Google Scholar] [CrossRef]
- Zou, T.; Jiang, S.; Dissanayaka, W.L.; Heng, B.C.; Xu, J.; Gong, T.; Huang, X.; Zhang, C. Sema4D/PlexinB1 promotes endothelial differentiation of dental pulp stem cells via activation of AKT and ERK1/2 signaling. J. Cell Biochem. 2019, 120, 13614–13624. [Google Scholar] [CrossRef]
- Monache, S.D.; Martellucci, S.; Clementi, L.; Pulcini, F.; Santilli, F.; Mei, C.; Piccoli, L.; Angelucci, A.; Mattei, V. In Vitro Conditioning Determines the Capacity of Dental Pulp Stem Cells to Function as Pericyte-Like Cells. Stem Cells Dev. 2019, 28, 695–706. [Google Scholar] [CrossRef]
- Sasaki, J.; Zhang, Z.; Oh, M.; Pobocik, A.; Imazato, S.; Shi, S.; Nör, J. VE-Cadherin and Anastomosis of Blood Vessels Formed by Dental Stem Cells. J. Dent. Res. 2020, 99, 437–445. [Google Scholar] [CrossRef]
- Smith, A.J.; Duncan, H.F.; Diogenes, A.; Simon, S.; Cooper, P.R. Exploiting the Bioactive Properties of the Dentin-Pulp Complex in Regenerative Endodontics. J. Endod. 2016, 42, 47–56. [Google Scholar] [CrossRef]
- Leng, S.; Liu, L.; Xu, W.; Yang, F.; Du, J.; Ye, L.; Huang, D.; Zhang, L. Inflammation down regulates stromal cell-derived factor 1alpha in the early phase of pulpitis. Cytokine 2022, 158, 155983. [Google Scholar] [CrossRef]
- Yang, X.; van der Kraan, P.M.; Bian, Z.; Fan, M.; Walboomers, X.F.; Jansen, J.A. Mineralized tissue formation by BMP2-transfected pulp stem cells. J. Dent. Res. 2009, 88, 1020–1025. [Google Scholar] [CrossRef]
- Aksel, H.; Ozturk, S.; Serper, A.; Ulubayram, K. VEGF/BMP-2 loaded three-dimensional model for enhanced angiogenic and odontogenic potential of dental pulp stem cells. Int. Endod. J. 2018, 51, 420–430. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; Di Nisio, M.; Duffy, S.; Hernandez, A.V.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for Medical Use: A Systematic Review and Meta-analysis. JAMA 2015, 313, 2456–2473. [Google Scholar] [CrossRef]
- Liu, C.; Qi, X.; Yang, D.; Neely, A.; Zhou, Z. The effects of cannabis use on oral health. Oral Dis. 2019, 26, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Bab, I.; Zimmer, A.; Melamed, E. Cannabinoids and the skeleton: From marijuana to reversal of bone loss. Ann. Med. 2009, 41, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Mandolino, G.; Galuppo, M.; Bramanti, P.; Mazzon, E. Cannabinoids: New promising agents in the treatment of neurological diseases. Molecules 2014, 19, 18781–18816. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Muramatsu, T.; Sato, M.; Ichikawa, H.; Sahara, Y.; Tazaki, M.; Shibukawa, Y. TRPV1-mediated calcium signal couples with cannabinoid receptors and sodium-calcium exchangers in rat odontoblasts. Cell Calcium 2012, 52, 124–136. [Google Scholar] [CrossRef]
- Qi, X.; Liu, C.; Li, G.; Luan, H.; Li, S.; Yang, D.; Zhou, Z. Investigation of in vitro odonto/osteogenic capacity of cannabidiol on human dental pulp cell. J. Dent. 2021, 109, 103673. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar]
- Vasudevan, K.; Stahl, V. Cannabinoids infused mouthwash products are as effective as chlorhexidine on inhibition of total-culturable bacterial content in dental plaque samples. J. Cannabis Res. 2020, 2, 20. [Google Scholar] [CrossRef]
- Huang, G.T.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef]
- Petrescu, N.; Jurj, A.; Sorițău, O.; Lucaciu, O.; Dirzu, N.; Raduly, L.; Berindan-Neagoe, I.; Cenariu, M.; Boșca, B.; Campian, R.; et al. Cannabidiol and Vitamin D3 Impact on Osteogenic Differentiation of Human Dental Mesenchymal Stem Cells. Medicina 2020, 56, 607. [Google Scholar]
- Yu, C.; Abbott, P.V. An overview of the dental pulp: Its functions and responses to injury. Aust. Dent. J. 2007, 52, S4–S6. [Google Scholar]
- Zhu, X.; Liu, J.; Yu, Z.; Chen, C.A.; Aksel, H.; Azim, A.A.; Huang, G.T. A Miniature Swine Model for Stem Cell-Based De Novo Regeneration of Dental Pulp and Dentin-Like Tissue. Tissue Eng. Part C Methods 2018, 24, 108–120. [Google Scholar] [CrossRef]
- Rattanawarawipa, P.; Pavasant, P.; Osathanon, T.; Sukarawan, W. Effect of lithium chloride on cell proliferation and osteogenic differentiation in stem cells from human exfoliated deciduous teeth. Tissue Cell 2016, 48, 425–431. [Google Scholar] [CrossRef]
- Schönhofen, P.; De Medeiros, L.M.; Bristot, I.J.; Lopes, F.M.; De Bastiani, M.A.; Kapczinski, F.; Crippa, J.A.S.; Castro, M.A.A.; Parsons, R.B.; Klamt, F. Cannabidiol Exposure During Neuronal Differentiation Sensitizes Cells Against Redox-Active Neurotoxins. Mol. Neurobiol. 2014, 52, 26–37. [Google Scholar] [CrossRef]
- Liang, C.; Liao, L.; Tian, W. Stem Cell-based Dental Pulp Regeneration: Insights From Signaling Pathways. Stem Cell Rev. Rep. 2021, 17, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Iohara, K.; Murakami, M.; Takeuchi, N.; Osako, Y.; Ito, M.; Ishizaka, R.; Utunomiya, S.; Nakamura, H.; Matsushita, K.; Nakashima, M. A novel combinatorial therapy with pulp stem cells and granulocyte colony-stimulating factor for total pulp regeneration. Stem Cells Transl. Med. 2013, 2, 521–533. [Google Scholar] [CrossRef]
- Hilkens, P.; Bronckaers, A.; Ratajczak, J.; Gervois, P.; Wolfs, E.; Lambrichts, I. The Angiogenic Potential of DPSCs and SCAPs in an In Vivo Model of Dental Pulp Regeneration. Stem Cells Int. 2017, 2017, 2582080. [Google Scholar] [CrossRef] [PubMed]
- Katata, C.; Sasaki, J.; Li, A.; Abe, G.; Nör, J.; Hayashi, M.; Imazato, S. Fabrication of Vascularized DPSC Constructs for Efficient Pulp Regeneration. J. Dent. Res. 2021, 100, 1351–1358. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)-key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar] [PubMed]
- Schmuhl, E.; Ramer, R.; Salamon, A.; Peters, K.; Hinz, B. Increase of mesenchymal stem cell migration by cannabidiol via activation of p42/44 MAPK. Biochem. Pharmacol. 2014, 87, 489–501. [Google Scholar] [CrossRef]
- Galler, K.; Weber, M.; Korkmaz, Y.; Widbiller, M.; Feuerer, M. Inflammatory Response Mechanisms of the Dentine-Pulp Complex and the Periapical Tissues. Int. J. Mol. Sci. 2021, 22, 1480. [Google Scholar] [CrossRef]
- Xiao, J.; Zheng, Y.; Zhang, W.; Zhang, Y.; Cao, P.; Liang, Y.; Bao, L.; Shi, S.; Feng, X. General Control Nonrepressed Protein 5 Modulates Odontogenic Differentiation Through NF-kappaB Pathway in Tumor Necrosis Factor-alpha-Mediated Impaired Human Dental Pulp Stem Cells. Cell Reprogram. 2022, 24, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, S.; Li, R.; Chen, X.; Guo, L.; Zheng, Y. The detection of pro-inflammatory cytokines in exudates from dental pulp tissues. Cytokine 2022, 153, 155846. [Google Scholar] [CrossRef]
- Jia, M.; Li, Q.; Guo, J.; Shi, W.; Zhu, L.; Huang, Y.; Li, Y.; Wang, L.; Ma, S.; Zhuang, T.; et al. Deletion of BACH1 Attenuates Atherosclerosis by Reducing Endothelial Inflammation. Circ. Res. 2022, 130, 1038–1055. [Google Scholar] [CrossRef] [PubMed]
- Lisboa, S.F.; Hartmann, A.; Vila-Verde, C.; Guimarães, F.S.; Joca, S.R. The NLRP3 inflammasome in stress response: Another target for the promiscuous cannabidiol? Curr. Neuropharmacol. 2023, 21, 284–308. [Google Scholar] [CrossRef]
- Corpetti, C.; Del Re, A.; Seguella, L.; Palenca, I.; Rurgo, S.; De Conno, B.; Pesce, M.; Sarnelli, G.; Esposito, G. Cannabidiol inhibits SARS-Cov-2 spike (S) protein-induced cytotoxicity and inflammation through a PPARgamma-dependent TLR4/NLRP3/Caspase-1 signaling suppression in Caco-2 cell line. Phytother. Res. 2021, 35, 6893–6903. [Google Scholar] [CrossRef] [PubMed]
- Muthumalage, T.; Rahman, I. Cannabidiol differentially regulates basal and LPS-induced inflammatory responses in macrophages, lung epithelial cells, and fibroblasts. Toxicol. Appl. Pharmacol. 2019, 382, 114713. [Google Scholar] [CrossRef]
- Arora, S.; Cooper, P.R.; Friedlander, L.T.; Rizwan, S.; Seo, B.; Rich, A.M.; Hussaini, H.M. Potential application of immunotherapy for modulation of pulp inflammation: Opportunities for vital pulp treatment. Int. Endod. J. 2021, 54, 1263–1274. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Acc. No | Primer Sequence (5′–3′) | Product Length (bp) |
|---|---|---|---|
| ALP | NM_001127501.4 | F: AACATCAGGGACATTGACGTG R: GTATCTCGGTTTGAAGCTCTTCC | 159 |
| OPN | NM_001040060.2 | F: GAAGTTTCGCAGACCTGACAT R: GTATGCACCATTCAACTCCTCG | 91 |
| OCN | NM_199173.6 | F: CACTCCTCGCCCTATTGGC R: CCCTCCTGCTTGGACACAAAG | 112 |
| COL-I | NM_000088.4 | F: GAGGGCCAAGACGAAGACATC R: CAGATCACGTCATCGCACAAC | 140 |
| TNF-α | NM_000594.4 | F: GAGGCCAAGCCCTGGTATG R: CGGGCCGATTGATCTCAGC | 91 |
| IL-1β | NM_000576.3 | F: ATGATGGCTTATTACAGTGGCAA R: GTCGGAGATTCGTAGCTGGA | 132 |
| IL-6 | NM_001371096.1 | F: ACTCACCTCTTCAGAACGAATTG R: CCATCTTTGGAAGGTTCAGGTTG | 149 |
| DSPP | NM_014208.3 | F: TGGCGATGCAGGTCACAAT R: CCATTCCCACTAGGACTCCCA | 249 |
| DMP1 | NM_001079911.3 | F: CACTCAAGATTCAGGTGGCAG R: TCTGAGATGCGAGACTTCCTAAA | 75 |
| ICAM | NM_000201.3 | F: ATGCCCAGACATCTGTGTCC R: GGGGTCTCTATGCCCAACAA | 112 |
| VEGF | NM_001025366.3 | F: GGAGGCAGAGAAAAGAGAAAGTGT R: TAAGAGAGCAAGAGAGAGCAAAAGA | 175 |
| CD31 | NM_000442.5 | F: CTCCAGACTCCACCACCTTAC R: GAACTTTGCCTATTTCTTACCA | 243 |
| GAPDH | NM_001357943.2 | F: GAAGGTGAAGGTCGGAGTCA R: GAAGATGGTGATGGGATTTC | 172 |
| ON CB1 CB2 | NM_001309444.2 NM_001365874.3 XM_047444833.1 | F:TCTTCCCTGTACACTGGCAGTTC R:AGCTCGGTGTGGGAGAGGTA F:GTGTTCCACCGCAAAGATAGC R:GGGGCCTGTGAATGGATATGT F:AGCCCTCATACCTGTTCATTGG R:GTGAAGGTCATAGTCACGCTG | 73 130 154 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Zeng, L.; Zhang, Z.; Zhu, G.; Xu, Z.; Xia, J.; Weng, J.; Li, J.; Pathak, J.L. Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells. Biomolecules 2023, 13, 118. https://doi.org/10.3390/biom13010118
Yu L, Zeng L, Zhang Z, Zhu G, Xu Z, Xia J, Weng J, Li J, Pathak JL. Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells. Biomolecules. 2023; 13(1):118. https://doi.org/10.3390/biom13010118
Chicago/Turabian StyleYu, Lina, Liting Zeng, Zeyu Zhang, Guanxiong Zhu, Zidan Xu, Junyi Xia, Jinlong Weng, Jiang Li, and Janak Lal Pathak. 2023. "Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells" Biomolecules 13, no. 1: 118. https://doi.org/10.3390/biom13010118
APA StyleYu, L., Zeng, L., Zhang, Z., Zhu, G., Xu, Z., Xia, J., Weng, J., Li, J., & Pathak, J. L. (2023). Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells. Biomolecules, 13(1), 118. https://doi.org/10.3390/biom13010118