
Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies




Abstract
1. Introduction
2. Polyphenols
2.1. Polyphenol Flavonol
2.1.1. Quercetin
2.1.2. Apigenin
2.1.3. Baicalin
2.1.4. Cyanidin-3-Glucoside
2.1.5. Daidzein
2.1.6. Delphinidin
2.1.7. Epigallocatechin-3-Gallate
2.1.8. Fisetin
2.1.9. Formononetin
2.1.10. Genistein
2.1.11. Glycyrrhiza Compounds
2.1.12. Licochalcone
2.1.13. Luteolin
2.2. Polyphenol Lignans
2.2.1. Arctigenin
2.2.2. Honokiol
2.2.3. Magnolol
2.2.4. Obovatol
2.2.5. Silibinin
2.3. Polyphenol Stilbenes
Resveratrol
2.4. Other Polyphenols
2.4.1. Curcumin
2.4.2. Ellagitannin
2.4.3. Gallic Acid
2.4.4. Gossypol
3. Terpenoids
3.1. Artemisinin
3.2. Betulinic Acid
3.3. Germacrone
3.4. Ginsenosides
3.5. Glycyrrhizin
3.6. Lycopene
3.7. Oridonin
3.8. Thymoquinone
3.9. Ursolic Acid
3.10. β-Elemonic Acid
4. Taxanes
4.1. Cabazitaxel
4.2. Docetaxel (DTX)
4.3. Paclitaxel (PTX)
5. Alkaloids
5.1. Anibamine
5.2. Berberine
5.3. Capsaicin
5.4. Neferine
5.5. Piperine
5.6. Sanguinarine
6. Other
6.1. Gambogic Acid
6.2. Glucoraphanin
6.2.1. Sulforaphane
6.2.2. Phenethyl-Isothiocyanate
6.2.3. Indole-3-Carbinol
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front. Public Health 2022, 10, 811044. [Google Scholar] [CrossRef] [PubMed]
- Crawford, E.D.; Heidenreich, A.; Lawrentschuk, N.; Tombal, B.; Pompeo, A.C.L.; Mendoza-Valdes, A.; Miller, K.; Debruyne, F.M.J.; Klotz, L. Androgen-targeted therapy in men with prostate cancer: Evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019, 22, 24–38. [Google Scholar] [CrossRef]
- Dong, L.; Zieren, R.C.; Xue, W.; de Reijke, T.M.; Pienta, K.J. Metastatic prostate cancer remains incurable, why? Asian J. Urol. 2019, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Patel, J.; He, M.; Foulk, B.; Kraft, K.; Smirnov, D.A.; Twardowski, P.; Kortylewski, M.; Bhargava, V.; Jones, J.O. Identification of mechanisms of resistance to treatment with abiraterone acetate or enzalutamide in patients with castration-resistant prostate cancer (CRPC). Cancer 2018, 124, 1216–1224. [Google Scholar] [CrossRef]
- Gjyrezi, A.; Xie, F.; Voznesensky, O.; Khanna, P.; Calagua, C.; Bai, Y.; Kung, J.; Wu, J.; Corey, E.; Montgomery, B.; et al. Taxane resistance in prostate cancer is mediated by decreased drug-target engagement. J. Clin. Investig. 2020, 130, 3287–3298. [Google Scholar] [CrossRef]
- Erdogan, S.; Turkekul, K.; Dibirdik, I.; Doganlar, O.; Doganlar, Z.B.; Bilir, A.; Oktem, G. Midkine downregulation increases the efficacy of quercetin on prostate cancer stem cell survival and migration through PI3K/AKT and MAPK/ERK pathway. Biomed. Pharm. 2018, 107, 793–805. [Google Scholar] [CrossRef]
- Aalinkeel, R.; Bindukumar, B.; Reynolds, J.L.; Sykes, D.E.; Mahajan, S.D.; Chadha, K.C.; Schwartz, S.A. The dietary bioflavonoid, quercetin, selectively induces apoptosis of prostate cancer cells by down-regulating the expression of heat shock protein 90. Prostate 2008, 68, 1773–1789. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Apigenin suppresses insulin-like growth factor I receptor signaling in human prostate cancer: An in vitro and in vivo study. Mol. Carcinog. 2009, 48, 243–252. [Google Scholar] [CrossRef]
- Pandey, M.; Kaur, P.; Shukla, S.; Abbas, A.; Fu, P.; Gupta, S. Plant flavone apigenin inhibits HDAC and remodels chromatin to induce growth arrest and apoptosis in human prostate cancer cells: In vitro and in vivo study. Mol. Carcinog. 2012, 51, 952–962. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Apigenin-induced prostate cancer cell death is initiated by reactive oxygen species and p53 activation. Free Radic. Biol. Med. 2008, 44, 1833–1845. [Google Scholar] [CrossRef]
- Mirzoeva, S.; Franzen, C.A.; Pelling, J.C. Apigenin inhibits TGF-beta-induced VEGF expression in human prostate carcinoma cells via a Smad2/3- and Src-dependent mechanism. Mol. Carcinog. 2014, 53, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Hu, X.; Xing, Z.; Xing, R.; Lv, R.; Cheng, X.; Su, J.; Zhou, Z.; Xu, Z.; Nilsson, S.; et al. Baicalein inhibits prostate cancer cell growth and metastasis via the caveolin-1/AKT/mTOR pathway. Mol. Cell Biochem. 2015, 406, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Chen, Q.; Liu, Y.; Wen, X. Baicalein suppresses the androgen receptor (AR)-mediated prostate cancer progression via inhibiting the AR N-C dimerization and AR-coactivators interaction. Oncotarget 2017, 8, 105561–105573. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.C.; Chen, R.; Yang, T.N.; Zhang, F.; Zhao, D. Baicalein inhibits the proliferative activity of human prostate cancer cell line PC3 by downregulating Ezrin. J. Biol. Regul. Homeost. Agents 2020, 34, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Jongsomchai, K.; Leardkamolkarn, V.; Mahatheeranont, S. A rice bran phytochemical, cyanidin 3-glucoside, inhibits the progression of PC3 prostate cancer cell. Anat. Cell Biol. 2020, 53, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Rabiau, N.; Kossai, M.; Braud, M.; Chalabi, N.; Satih, S.; Bignon, Y.J.; Bernard-Gallon, D.J. Genistein and daidzein act on a panel of genes implicated in cell cycle and angiogenesis by polymerase chain reaction arrays in human prostate cancer cell lines. Cancer Epidemiol. 2010, 34, 200–206. [Google Scholar] [CrossRef]
- Vardi, A.; Bosviel, R.; Rabiau, N.; Adjakly, M.; Satih, S.; Dechelotte, P.; Boiteux, J.P.; Fontana, L.; Bignon, Y.J.; Guy, L.; et al. Soy phytoestrogens modify DNA methylation of GSTP1, RASSF1A, EPH2 and BRCA1 promoter in prostate cancer cells. In Vivo 2010, 24, 393–400. [Google Scholar]
- Ranjithkumar, R.; Saravanan, K.; Balaji, B.; Hima, S.; Sreeja, S.; Timane, S.R.; Ram Pravin Kumar, M.; Kabilan, S.; Ramanathan, M. Novel daidzein molecules exhibited anti-prostate cancer activity through nuclear receptor ERbeta modulation, in vitro and in vivo studies. J. Chemother. 2021, 33, 582–594. [Google Scholar] [CrossRef]
- Bin Hafeez, B.; Asim, M.; Siddiqui, I.A.; Adhami, V.M.; Murtaza, I.; Mukhtar, H. Delphinidin, a dietary anthocyanidin in pigmented fruits and vegetables: A new weapon to blunt prostate cancer growth. Cell Cycle 2008, 7, 3320–3326. [Google Scholar] [CrossRef]
- Lee, W.; Yun, J.M. Suppression of beta-catenin Signaling Pathway in Human Prostate Cancer PC3 Cells by Delphinidin. J. Cancer Prev. 2016, 21, 110–114. [Google Scholar] [CrossRef]
- Ko, H.; Jeong, M.H.; Jeon, H.; Sung, G.J.; So, Y.; Kim, I.; Son, J.; Lee, S.W.; Yoon, H.G.; Choi, K.C. Delphinidin sensitizes prostate cancer cells to TRAIL-induced apoptosis, by inducing DR5 and causing caspase-mediated HDAC3 cleavage. Oncotarget 2015, 6, 9970–9984. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.H.; Ko, H.; Jeon, H.; Sung, G.J.; Park, S.Y.; Jun, W.J.; Lee, Y.H.; Lee, J.; Lee, S.W.; Yoon, H.G.; et al. Delphinidin induces apoptosis via cleaved HDAC3-mediated p53 acetylation and oligomerization in prostate cancer cells. Oncotarget 2016, 7, 56767–56780. [Google Scholar] [CrossRef]
- Albrecht, D.S.; Clubbs, E.A.; Ferruzzi, M.; Bomser, J.A. Epigallocatechin-3-gallate (EGCG) inhibits PC-3 prostate cancer cell proliferation via MEK-independent ERK1/2 activation. Chem. Biol. Interact. 2008, 171, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Asim, M.; Hafeez, B.B.; Adhami, V.M.; Tarapore, R.S.; Mukhtar, H. Green tea polyphenol EGCG blunts androgen receptor function in prostate cancer. FASEB J. 2011, 25, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Chuu, C.P.; Chen, R.Y.; Kokontis, J.M.; Hiipakka, R.A.; Liao, S. Suppression of androgen receptor signaling and prostate specific antigen expression by (-)-epigallocatechin-3-gallate in different progression stages of LNCaP prostate cancer cells. Cancer Lett. 2009, 275, 86–92. [Google Scholar] [CrossRef]
- Hagen, R.M.; Chedea, V.S.; Mintoff, C.P.; Bowler, E.; Morse, H.R.; Ladomery, M.R. Epigallocatechin-3-gallate promotes apoptosis and expression of the caspase 9a splice variant in PC3 prostate cancer cells. Int. J. Oncol. 2013, 43, 194–200. [Google Scholar] [CrossRef]
- Yeo, C.; Han, D.S.; Lee, H.J.; Lee, E.O. Epigallocatechin-3-Gallate Suppresses Vasculogenic Mimicry through Inhibiting the Twist/VE-Cadherin/AKT Pathway in Human Prostate Cancer PC-3 Cells. Int. J. Mol. Sci. 2020, 21, 439. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kwak, J.; Choi, H.K.; Choi, K.C.; Kim, S.; Lee, J.; Jun, W.; Park, H.J.; Yoon, H.G. EGCG suppresses prostate cancer cell growth modulating acetylation of androgen receptor by anti-histone acetyltransferase activity. Int. J. Mol. Med. 2012, 30, 69–74. [Google Scholar] [CrossRef]
- Modernelli, A.; Naponelli, V.; Giovanna Troglio, M.; Bonacini, M.; Ramazzina, I.; Bettuzzi, S.; Rizzi, F. EGCG antagonizes Bortezomib cytotoxicity in prostate cancer cells by an autophagic mechanism. Sci. Rep. 2015, 5, 15270. [Google Scholar] [CrossRef]
- Chien, C.S.; Shen, K.H.; Huang, J.S.; Ko, S.C.; Shih, Y.W. Antimetastatic potential of fisetin involves inactivation of the PI3K/Akt and JNK signaling pathways with downregulation of MMP-2/9 expressions in prostate cancer PC-3 cells. Mol. Cell Biochem. 2010, 333, 169–180. [Google Scholar] [CrossRef]
- Adhami, V.M.; Syed, D.N.; Khan, N.; Mukhtar, H. Dietary flavonoid fisetin: A novel dual inhibitor of PI3K/Akt and mTOR for prostate cancer management. Biochem. Pharm. 2012, 84, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Asim, M.; Afaq, F.; Abu Zaid, M.; Mukhtar, H. A novel dietary flavonoid fisetin inhibits androgen receptor signaling and tumor growth in athymic nude mice. Cancer Res. 2008, 68, 8555–8563. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Helewski, K.J.; Mizgala, E.; Krol, W. The dietary flavonol fisetin enhances the apoptosis-inducing potential of TRAIL in prostate cancer cells. Int. J. Oncol. 2011, 39, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, E.; Adhami, V.M.; Sechi, M.; Mukhtar, H. Dietary flavonoid fisetin binds to beta-tubulin and disrupts microtubule dynamics in prostate cancer cells. Cancer Lett. 2015, 367, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Hou, R.; Chen, J.; Mo, L.; Zhang, J.; Huang, Y.; Mo, Z. Formononetin-induced apoptosis of human prostate cancer cells through ERK1/2 mitogen-activated protein kinase inactivation. Horm. Metab. Res. 2012, 44, 263–267. [Google Scholar] [CrossRef]
- Huang, W.J.; Bi, L.Y.; Li, Z.Z.; Zhang, X.; Ye, Y. Formononetin induces the mitochondrial apoptosis pathway in prostate cancer cells via downregulation of the IGF-1/IGF-1R signaling pathway. Pharm. Biol. 2013, 52, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bi, L.; Ye, Y.; Chen, J. Formononetin induces apoptosis in PC-3 prostate cancer cells through enhancing the Bax/Bcl-2 ratios and regulating the p38/Akt pathway. Nutr. Cancer 2014, 66, 656–661. [Google Scholar] [CrossRef]
- Liu, X.J.; Li, Y.Q.; Chen, Q.Y.; Xiao, S.J.; Zeng, S.E. Up-regulating of RASD1 and apoptosis of DU-145 human prostate cancer cells induced by formononetin in vitro. Asian Pac. J. Cancer Prev. 2014, 15, 2835–2839. [Google Scholar] [CrossRef]
- Lee, J.; Ju, J.; Park, S.; Hong, S.J.; Yoon, S. Inhibition of IGF-1 signaling by genistein: Modulation of E-cadherin expression and downregulation of beta-catenin signaling in hormone refractory PC-3 prostate cancer cells. Nutr. Cancer 2012, 64, 153–162. [Google Scholar] [CrossRef]
- Hirata, H.; Hinoda, Y.; Shahryari, V.; Deng, G.; Tanaka, Y.; Tabatabai, Z.L.; Dahiya, R. Genistein downregulates onco-miR-1260b and upregulates sFRP1 and Smad4 via demethylation and histone modification in prostate cancer cells. Br. J. Cancer 2014, 110, 1645–1654. [Google Scholar] [CrossRef]
- Chiyomaru, T.; Yamamura, S.; Fukuhara, S.; Yoshino, H.; Kinoshita, T.; Majid, S.; Saini, S.; Chang, I.; Tanaka, Y.; Enokida, H.; et al. Genistein inhibits prostate cancer cell growth by targeting miR-34a and oncogenic HOTAIR. PLoS ONE 2013, 8, e70372. [Google Scholar] [CrossRef] [PubMed]
- Seon, M.R.; Lim, S.S.; Choi, H.J.; Park, S.Y.; Cho, H.J.; Kim, J.K.; Kim, J.; Kwon, D.Y.; Park, J.H. Isoangustone A present in hexane/ethanol extract of Glycyrrhiza uralensis induces apoptosis in DU145 human prostate cancer cells via the activation of DR4 and intrinsic apoptosis pathway. Mol. Nutr. Food Res. 2010, 54, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Seon, M.R.; Park, S.Y.; Kwon, S.J.; Lim, S.S.; Choi, H.J.; Park, H.; Lim, D.Y.; Kim, J.S.; Lee, C.H.; Kim, J.; et al. Hexane/ethanol extract of Glycyrrhiza uralensis and its active compound isoangustone A induce G1 cycle arrest in DU145 human prostate and 4T1 murine mammary cancer cells. J. Nutr. Biochem. 2012, 23, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lim, S.S.; Kim, J.K.; Kang, I.J.; Kim, J.S.; Lee, C.; Kim, J.; Park, J.H. Hexane-ethanol extract of Glycyrrhiza uralensis containing licoricidin inhibits the metastatic capacity of DU145 human prostate cancer cells. Br. J. Nutr. 2010, 104, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Hsieh, T.C.; Guo, J.; Kunicki, J.; Lee, M.Y.; Darzynkiewicz, Z.; Wu, J.M. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2004, 322, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhou, Q.; Shi, X.L.; Jiang, B.H. Luteolin inhibits insulin-like growth factor 1 receptor signaling in prostate cancer cells. Carcinogenesis 2007, 28, 713–723. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Son, Y.O.; Budhraja, A.; Wang, X.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.C.; Kim, D.; Divya, S.P.; et al. Luteolin inhibits human prostate tumor growth by suppressing vascular endothelial growth factor receptor 2-mediated angiogenesis. PLoS ONE 2012, 7, e52279. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Phan, T.; Gordon, D.; Chung, S.; Henning, S.M.; Vadgama, J.V. Arctigenin in combination with quercetin synergistically enhances the antiproliferative effect in prostate cancer cells. Mol. Nutr. Food Res. 2015, 59, 250–261. [Google Scholar] [CrossRef]
- Hao, Q.; Diaz, T.; Verduzco, A.D.R.; Magyar, C.E.; Zhong, J.; Elshimali, Y.; Rettig, M.B.; Henning, S.M.; Vadgama, J.V.; Wang, P. Arctigenin inhibits prostate tumor growth in high-fat diet fed mice through dual actions on adipose tissue and tumor. Sci. Rep. 2020, 10, 1403. [Google Scholar] [CrossRef]
- Sun, B.L.; Cai, E.B.; Zhao, Y.; Wang, Y.; Yang, L.M.; Wang, J.Y. Arctigenin Triggers Apoptosis and Autophagy via PI3K/Akt/mTOR Inhibition in PC-3M Cells. Chem. Pharm. Bull. 2021, 69, 472–480. [Google Scholar] [CrossRef]
- Hahm, E.R.; Singh, S.V. Honokiol causes G0-G1 phase cell cycle arrest in human prostate cancer cells in association with suppression of retinoblastoma protein level/phosphorylation and inhibition of E2F1 transcriptional activity. Mol. Cancer Ther. 2007, 6, 2686–2695. [Google Scholar] [CrossRef] [PubMed]
- Hahm, E.R.; Arlotti, J.A.; Marynowski, S.W.; Singh, S.V. Honokiol, a constituent of oriental medicinal herb magnolia officinalis, inhibits growth of PC-3 xenografts in vivo in association with apoptosis induction. Clin. Cancer Res. 2008, 14, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Szczepanski, M.J.; Lee, Y.J. Magnolol induces apoptosis via inhibiting the EGFR/PI3K/Akt signaling pathway in human prostate cancer cells. J. Cell Biochem. 2009, 106, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Yuk, D.Y.; Song, H.S.; Yoon, D.Y.; Jung, J.K.; Moon, D.C.; Lee, B.S.; Hong, J.T. Growth inhibitory effects of obovatol through induction of apoptotic cell death in prostate and colon cancer by blocking of NF-kappaB. Eur. J. Pharm. 2008, 582, 17–25. [Google Scholar] [CrossRef]
- Roy, S.; Kaur, M.; Agarwal, C.; Tecklenburg, M.; Sclafani, R.A.; Agarwal, R. p21 and p27 induction by silibinin is essential for its cell cycle arrest effect in prostate carcinoma cells. Mol. Cancer Ther. 2007, 6, 2696–2707. [Google Scholar] [CrossRef]
- Agarwal, C.; Tyagi, A.; Kaur, M.; Agarwal, R. Silibinin inhibits constitutive activation of Stat3, and causes caspase activation and apoptotic death of human prostate carcinoma DU145 cells. Carcinogenesis 2007, 28, 1463–1470. [Google Scholar] [CrossRef]
- Wu, K.; Zeng, J.; Li, L.; Fan, J.; Zhang, D.; Xue, Y.; Zhu, G.; Yang, L.; Wang, X.; He, D. Silibinin reverses epithelial-to-mesenchymal transition in metastatic prostate cancer cells by targeting transcription factors. Oncol. Rep. 2010, 23, 1545–1552. [Google Scholar]
- Singh, R.P.; Raina, K.; Sharma, G.; Agarwal, R. Silibinin inhibits established prostate tumor growth, progression, invasion, and metastasis and suppresses tumor angiogenesis and epithelial-mesenchymal transition in transgenic adenocarcinoma of the mouse prostate model mice. Clin. Cancer Res. 2008, 14, 7773–7780. [Google Scholar] [CrossRef]
- Kumar, A.; Dhar, S.; Rimando, A.M.; Lage, J.M.; Lewin, J.R.; Zhang, X.; Levenson, A.S. Epigenetic potential of resveratrol and analogs in preclinical models of prostate cancer. Ann. N. Y. Acad. Sci. 2015, 1348, 1–9. [Google Scholar] [CrossRef]
- Ganapathy, S.; Chen, Q.; Singh, K.P.; Shankar, S.; Srivastava, R.K. Resveratrol enhances antitumor activity of TRAIL in prostate cancer xenografts through activation of FOXO transcription factor. PLoS ONE 2010, 5, e15627. [Google Scholar] [CrossRef]
- Jang, Y.G.; Go, R.E.; Hwang, K.A.; Choi, K.C. Resveratrol inhibits DHT-induced progression of prostate cancer cell line through interfering with the AR and CXCR4 pathway. J. Steroid Biochem. Mol. Biol. 2019, 192, 105406. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Zhou, C.; Shi, Y.; Lu, H.; Xu, R.; He, X. Nuclear transcription factor Nrf2 suppresses prostate cancer cells growth and migration through upregulating ferroportin. Oncotarget 2016, 7, 78804–78812. [Google Scholar] [CrossRef] [PubMed]
- Mitani, T.; Harada, N.; Tanimori, S.; Nakano, Y.; Inui, H.; Yamaji, R. Resveratrol inhibits hypoxia-inducible factor-1alpha-mediated androgen receptor signaling and represses tumor progression in castration-resistant prostate cancer. J. Nutr. Sci. Vitaminol. 2014, 60, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Dorai, T.; Gehani, N.; Katz, A. Therapeutic potential of curcumin in human prostate cancer. II. Curcumin inhibits tyrosine kinase activity of epidermal growth factor receptor and depletes the protein. Mol. Urol. 2000, 4, 1–6. [Google Scholar]
- Hilchie, A.L.; Furlong, S.J.; Sutton, K.; Richardson, A.; Robichaud, M.R.; Giacomantonio, C.A.; Ridgway, N.D.; Hoskin, D.W. Curcumin-induced apoptosis in PC3 prostate carcinoma cells is caspase-independent and involves cellular ceramide accumulation and damage to mitochondria. Nutr. Cancer 2010, 62, 379–389. [Google Scholar] [CrossRef]
- Teiten, M.H.; Gaascht, F.; Cronauer, M.; Henry, E.; Dicato, M.; Diederich, M. Anti-proliferative potential of curcumin in androgen-dependent prostate cancer cells occurs through modulation of the Wingless signaling pathway. Int. J. Oncol. 2011, 38, 603–611. [Google Scholar] [CrossRef]
- Khurana, N.; Sikka, S.C. Targeting Crosstalk between Nrf-2, NF-kappaB and Androgen Receptor Signaling in Prostate Cancer. Cancers 2018, 10, 352. [Google Scholar] [CrossRef]
- Hong, J.H.; Ahn, K.S.; Bae, E.; Jeon, S.S.; Choi, H.Y. The effects of curcumin on the invasiveness of prostate cancer in vitro and in vivo. Prostate Cancer Prostatic Dis. 2006, 9, 147–152. [Google Scholar] [CrossRef]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.P.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B.E. Curcumin inhibits prostate cancer metastasis in vivo by targeting the inflammatory cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef]
- Vanella, L.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Cardile, V.; Kim, D.H.; Abraham, N.G.; Sorrenti, V. Apoptotic markers in a prostate cancer cell line: Effect of ellagic acid. Oncol. Rep. 2013, 30, 2804–2810. [Google Scholar] [CrossRef]
- Vanella, L.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Li Volti, G.; Cardile, V.; Abraham, N.G.; Sorrenti, V. Effects of ellagic Acid on angiogenic factors in prostate cancer cells. Cancers 2013, 5, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Naiki-Ito, A.; Chewonarin, T.; Tang, M.; Pitchakarn, P.; Kuno, T.; Ogawa, K.; Asamoto, M.; Shirai, T.; Takahashi, S. Ellagic acid, a component of pomegranate fruit juice, suppresses androgen-dependent prostate carcinogenesis via induction of apoptosis. Prostate 2015, 75, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Pitchakarn, P.; Chewonarin, T.; Ogawa, K.; Suzuki, S.; Asamoto, M.; Takahashi, S.; Shirai, T.; Limtrakul, P. Ellagic acid inhibits migration and invasion by prostate cancer cell lines. Asian Pac. J. Cancer Prev. 2013, 14, 2859–2863. [Google Scholar] [CrossRef] [PubMed]
- Mohammed Saleem, Y.I.; Albassam, H.; Selim, M. Urolithin A induces prostate cancer cell death in p53-dependent and in p53-independent manner. Eur. J. Nutr. 2020, 59, 1607–1618. [Google Scholar] [CrossRef]
- Wang, D.; Ozen, C.; Abu-Reidah, I.M.; Chigurupati, S.; Patra, J.K.; Horbanczuk, J.O.; Jozwik, A.; Tzvetkov, N.T.; Uhrin, P.; Atanasov, A.G. Vasculoprotective Effects of Pomegranate (Punica granatum L.). Front. Pharm. 2018, 9, 544. [Google Scholar] [CrossRef]
- Agarwal, C.; Tyagi, A.; Agarwal, R. Gallic acid causes inactivating phosphorylation of cdc25A/cdc25C-cdc2 via ATM-Chk2 activation, leading to cell cycle arrest, and induces apoptosis in human prostate carcinoma DU145 cells. Mol. Cancer Ther. 2006, 5, 3294–3302. [Google Scholar] [CrossRef]
- Chen, H.M.; Wu, Y.C.; Chia, Y.C.; Chang, F.R.; Hsu, H.K.; Hsieh, Y.C.; Chen, C.C.; Yuan, S.S. Gallic acid, a major component of Toona sinensis leaf extracts, contains a ROS-mediated anti-cancer activity in human prostate cancer cells. Cancer Lett. 2009, 286, 161–171. [Google Scholar] [CrossRef]
- Liu, K.C.; Huang, A.C.; Wu, P.P.; Lin, H.Y.; Chueh, F.S.; Yang, J.S.; Lu, C.C.; Chiang, J.H.; Meng, M.; Chung, J.G. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and -9 signaling pathways. Oncol. Rep. 2011, 26, 177–184. [Google Scholar] [CrossRef]
- Raina, K.; Rajamanickam, S.; Deep, G.; Singh, M.; Agarwal, R.; Agarwal, C. Chemopreventive effects of oral gallic acid feeding on tumor growth and progression in TRAMP mice. Mol. Cancer Ther. 2008, 7, 1258–1267. [Google Scholar] [CrossRef]
- Kaur, M.; Velmurugan, B.; Rajamanickam, S.; Agarwal, R.; Agarwal, C. Gallic acid, an active constituent of grape seed extract, exhibits anti-proliferative, pro-apoptotic and anti-tumorigenic effects against prostate carcinoma xenograft growth in nude mice. Pharm. Res. 2009, 26, 2133–2140. [Google Scholar] [CrossRef]
- Jiang, J.; Ye, W.; Lin, Y.C. Gossypol inhibits the growth of MAT-LyLu prostate cancer cells by modulation of TGFbeta/Akt signaling. Int. J. Mol. Med. 2009, 24, 69–75. [Google Scholar] [PubMed]
- Zhang, M.; Liu, H.; Tian, Z.; Griffith, B.N.; Ji, M.; Li, Q.Q. Gossypol induces apoptosis in human PC-3 prostate cancer cells by modulating caspase-dependent and caspase-independent cell death pathways. Life Sci. 2007, 80, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Karnak, D.; Xu, L. The Bcl-2-Beclin 1 interaction in (-)-gossypol-induced autophagy versus apoptosis in prostate cancer cells. Autophagy 2010, 6, 1201–1203. [Google Scholar] [CrossRef]
- Jiang, J.; Slivova, V.; Jedinak, A.; Sliva, D. Gossypol inhibits growth, invasiveness, and angiogenesis in human prostate cancer cells by modulating NF-kappaB/AP-1 dependent- and independent-signaling. Clin. Exp. Metastasis 2012, 29, 165–178. [Google Scholar] [CrossRef]
- Pang, X.; Wu, Y.; Wu, Y.; Lu, B.; Chen, J.; Wang, J.; Yi, Z.; Qu, W.; Liu, M. (-)-Gossypol suppresses the growth of human prostate cancer xenografts via modulating VEGF signaling-mediated angiogenesis. Mol. Cancer Ther. 2011, 10, 795–805. [Google Scholar] [CrossRef]
- Willoughby, J.A., Sr.; Sundar, S.N.; Cheung, M.; Tin, A.S.; Modiano, J.; Firestone, G.L. Artemisinin blocks prostate cancer growth and cell cycle progression by disrupting Sp1 interactions with the cyclin-dependent kinase-4 (CDK4) promoter and inhibiting CDK4 gene expression. J. Biol. Chem. 2009, 284, 2203–2213. [Google Scholar] [CrossRef]
- He, Q.; Shi, J.; Shen, X.L.; An, J.; Sun, H.; Wang, L.; Hu, Y.J.; Sun, Q.; Fu, L.C.; Sheikh, M.S.; et al. Dihydroartemisinin upregulates death receptor 5 expression and cooperates with TRAIL to induce apoptosis in human prostate cancer cells. Cancer Biol. Ther. 2010, 9, 819–824. [Google Scholar] [CrossRef]
- Rabi, T.; Shukla, S.; Gupta, S. Betulinic acid suppresses constitutive and TNFalpha-induced NF-kappaB activation and induces apoptosis in human prostate carcinoma PC-3 cells. Mol. Carcinog. 2008, 47, 964–973. [Google Scholar] [CrossRef]
- Reiner, T.; Parrondo, R.; de Las Pozas, A.; Palenzuela, D.; Perez-Stable, C. Betulinic acid selectively increases protein degradation and enhances prostate cancer-specific apoptosis: Possible role for inhibition of deubiquitinase activity. PLoS ONE 2013, 8, e56234. [Google Scholar] [CrossRef]
- de Las Pozas, A.; Reiner, T.; De Cesare, V.; Trost, M.; Perez-Stable, C. Inhibiting Multiple Deubiquitinases to Reduce Androgen Receptor Expression in Prostate Cancer Cells. Sci. Rep. 2018, 8, 13146. [Google Scholar] [CrossRef]
- Yu, Z.; Xu, J.; Shao, M.; Zou, J. Germacrone Induces Apoptosis as Well as Protective Autophagy in Human Prostate Cancer Cells. Cancer Manag. Res. 2020, 12, 4009–4016. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.Y.; Guo, H.; Han, J.; Hao, F.; An, Y.; Xu, Y.; Xiaokaiti, Y.; Pan, Y.; Li, X.J. Ginsenoside Rg3 attenuates cell migration via inhibition of aquaporin 1 expression in PC-3M prostate cancer cells. Eur. J. Pharm. 2012, 683, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, E.H.; Ko, S.R.; Choi, K.J.; Park, J.H.; Im, D.S. Effects of ginsenosides Rg3 and Rh2 on the proliferation of prostate cancer cells. Arch. Pharm. Res. 2004, 27, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Musende, A.G.; Eberding, A.; Wood, C.; Adomat, H.; Fazli, L.; Hurtado-Coll, A.; Jia, W.; Bally, M.B.; Guns, E.T. Pre-clinical evaluation of Rh2 in PC-3 human xenograft model for prostate cancer in vivo: Formulation, pharmacokinetics, biodistribution and efficacy. Cancer Chemother. Pharm. 2009, 64, 1085–1095. [Google Scholar] [CrossRef]
- Gioti, K.; Papachristodoulou, A.; Benaki, D.; Beloukas, A.; Vontzalidou, A.; Aligiannis, N.; Skaltsounis, A.L.; Mikros, E.; Tenta, R. Glycyrrhiza glabra-Enhanced Extract and Adriamycin Antiproliferative Effect on PC-3 Prostate Cancer Cells. Nutr. Cancer 2020, 72, 320–332. [Google Scholar] [CrossRef]
- Rafi, M.M.; Kanakasabai, S.; Reyes, M.D.; Bright, J.J. Lycopene modulates growth and survival associated genes in prostate cancer. J. Nutr. Biochem. 2013, 24, 1724–1734. [Google Scholar] [CrossRef]
- Yang, C.M.; Lu, I.H.; Chen, H.Y.; Hu, M.L. Lycopene inhibits the proliferation of androgen-dependent human prostate tumor cells through activation of PPARgamma-LXRalpha-ABCA1 pathway. J. Nutr. Biochem. 2012, 23, 8–17. [Google Scholar] [CrossRef]
- Yang, C.M.; Yen, Y.T.; Huang, C.S.; Hu, M.L. Growth inhibitory efficacy of lycopene and beta-carotene against androgen-independent prostate tumor cells xenografted in nude mice. Mol. Nutr. Food Res. 2011, 55, 606–612. [Google Scholar] [CrossRef]
- Konijeti, R.; Henning, S.; Moro, A.; Sheikh, A.; Elashoff, D.; Shapiro, A.; Ku, M.; Said, J.W.; Heber, D.; Cohen, P.; et al. Chemoprevention of prostate cancer with lycopene in the TRAMP model. Prostate 2010, 70, 1547–1554. [Google Scholar] [CrossRef]
- Chen, S.; Gao, J.; Halicka, H.D.; Huang, X.; Traganos, F.; Darzynkiewicz, Z. The cytostatic and cytotoxic effects of oridonin (Rubescenin), a diterpenoid from Rabdosia rubescens, on tumor cells of different lineage. Int. J. Oncol. 2005, 26, 579–588. [Google Scholar] [CrossRef]
- Li, X.; Li, X.; Wang, J.; Ye, Z.; Li, J.C. Oridonin up-regulates expression of P21 and induces autophagy and apoptosis in human prostate cancer cells. Int. J. Biol. Sci. 2012, 8, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Kaseb, A.O.; Chinnakannu, K.; Chen, D.; Sivanandam, A.; Tejwani, S.; Menon, M.; Dou, Q.P.; Reddy, G.P. Androgen receptor and E2F-1 targeted thymoquinone therapy for hormone-refractory prostate cancer. Cancer Res. 2007, 67, 7782–7788. [Google Scholar] [CrossRef] [PubMed]
- Koka, P.S.; Mondal, D.; Schultz, M.; Abdel-Mageed, A.B.; Agrawal, K.C. Studies on molecular mechanisms of growth inhibitory effects of thymoquinone against prostate cancer cells: Role of reactive oxygen species. Exp. Biol. Med. 2010, 235, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Cho, S.G.; Yi, Z.; Pang, X.; Rodriguez, M.; Wang, Y.; Sethi, G.; Aggarwal, B.B.; Liu, M. Thymoquinone inhibits tumor angiogenesis and tumor growth through suppressing AKT and extracellular signal-regulated kinase signaling pathways. Mol. Cancer Ther. 2008, 7, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kong, C.; Zeng, Y.; Wang, L.; Li, Z.; Wang, H.; Xu, C.; Sun, Y. Ursolic acid induces PC-3 cell apoptosis via activation of JNK and inhibition of Akt pathways in vitro. Mol. Carcinog. 2010, 49, 374–385. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Kong, C.Z.; Wang, L.H.; Li, J.Y.; Liu, X.K.; Xu, B.; Xu, C.L.; Sun, Y.H. Ursolic acid overcomes Bcl-2-mediated resistance to apoptosis in prostate cancer cells involving activation of JNK-induced Bcl-2 phosphorylation and degradation. J. Cell Biochem. 2010, 109, 764–773. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Manu, K.A.; Ong, T.H.; Ramachandran, L.; Surana, R.; Bist, P.; Lim, L.H.; Kumar, A.P.; Hui, K.M.; Sethi, G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer 2011, 129, 1552–1563. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Ong, T.H.; Kumar, A.P.; Lun, C.K.; Ho, P.C.; Wong, P.T.; Hui, K.M.; Sethi, G. Ursolic acid inhibits the initiation, progression of prostate cancer and prolongs the survival of TRAMP mice by modulating pro-inflammatory pathways. PLoS ONE 2012, 7, e32476. [Google Scholar] [CrossRef]
- Bao, X.; Zhu, J.; Ren, C.; Zhao, A.; Zhang, M.; Zhu, Z.; Lu, X.; Zhang, Y.; Li, X.; Sima, X.; et al. beta-elemonic acid inhibits growth and triggers apoptosis in human castration-resistant prostate cancer cells through the suppression of JAK2/STAT3/MCL-1 and NF-kB signal pathways. Chem. Biol. Interact. 2021, 342, 109477. [Google Scholar] [CrossRef]
- Mellado, B.; Jimenez, N.; Marin-Aguilera, M.; Reig, O. Diving Into Cabazitaxel’s Mode of Action: More Than a Taxane for the Treatment of Castration-Resistant Prostate Cancer Patients. Clin. Genitourin Cancer 2016, 14, 265–270. [Google Scholar] [CrossRef]
- Gan, L.; Wang, J.; Xu, H.; Yang, X. Resistance to docetaxel-induced apoptosis in prostate cancer cells by p38/p53/p21 signaling. Prostate 2011, 71, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Chintapalli, J.; Croce, C.M. Taxol induces bcl-2 phosphorylation and death of prostate cancer cells. Cancer Res. 1996, 56, 1253–1255. [Google Scholar] [PubMed]
- Gan, L.; Chen, S.; Wang, Y.; Watahiki, A.; Bohrer, L.; Sun, Z.; Wang, Y.; Huang, H. Inhibition of the androgen receptor as a novel mechanism of taxol chemotherapy in prostate cancer. Cancer Res. 2009, 69, 8386–8394. [Google Scholar] [CrossRef] [PubMed]
- Haney, K.M.; Zhang, F.; Arnatt, C.K.; Yuan, Y.; Li, G.; Ware, J.L.; Gewirtz, D.A.; Zhang, Y. The natural product CCR5 antagonist anibamine and its analogs as anti-prostate cancer agents. Bioorganic Med. Chem. Lett. 2011, 21, 5159–5163. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Liu, Z.; Li, B.; Sun, Z.; Zhou, H.; Zhang, X.; Gong, Y.; Shao, C. Berberine, a genotoxic alkaloid, induces ATM-Chk1 mediated G2 arrest in prostate cancer cells. Mutagenesis Res. 2012, 734, 20–29. [Google Scholar] [CrossRef]
- Li, J.; Cao, B.; Liu, X.; Fu, X.; Xiong, Z.; Chen, L.; Sartor, O.; Dong, Y.; Zhang, H. Berberine suppresses androgen receptor signaling in prostate cancer. Mol. Cancer Ther. 2011, 10, 1346–1356. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, C.; Yang, X.; Yang, B.; Wang, J.; Kang, Y.; Wang, Z.; Li, D.; Huang, G.; Ma, Z.; et al. Berberine inhibits the expression of hypoxia induction factor-1alpha and increases the radiosensitivity of prostate cancer. Diagn. Pathol. 2014, 9, 98. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Malagarie-Cazenave, S.; Olea, N.; Vara, D.; Chiloeches, A.; Diaz-Laviada, I. Apoptosis induced by capsaicin in prostate PC-3 cells involves ceramide accumulation, neutral sphingomyelinase, and JNK activation. Apoptosis 2007, 12, 2013–2024. [Google Scholar] [CrossRef]
- Mori, A.; Lehmann, S.; O’Kelly, J.; Kumagai, T.; Desmond, J.C.; Pervan, M.; McBride, W.H.; Kizaki, M.; Koeffler, H.P. Capsaicin, a component of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Res. 2006, 66, 3222–3229. [Google Scholar] [CrossRef]
- Erdogan, S.; Turkekul, K. Neferine inhibits proliferation and migration of human prostate cancer stem cells through p38 MAPK/JNK activation. J. Food Biochem. 2020, 44, e13253. [Google Scholar] [CrossRef]
- Samykutty, A.; Shetty, A.V.; Dakshinamoorthy, G.; Bartik, M.M.; Johnson, G.L.; Webb, B.; Zheng, G.; Chen, A.; Kalyanasundaram, R.; Munirathinam, G. Piperine, a Bioactive Component of Pepper Spice Exerts Therapeutic Effects on Androgen Dependent and Androgen Independent Prostate Cancer Cells. PLoS ONE 2013, 8, e65889. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, D.Y.; Zeng, L.H.; Pan, H.; Xu, L.L.H.; Wang, Y.; Liu, K.P.; He, X.H. Piperine inhibits the proliferation of human prostate cancer cells via induction of cell cycle arrest and autophagy. Food Chem. Toxicol. 2013, 60, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Makhov, P.; Golovine, K.; Canter, D.; Kutikov, A.; Simhan, J.; Corlew, M.M.; Uzzo, R.G.; Kolenko, V.M. Co-administration of piperine and docetaxel results in improved anti-tumor efficacy via inhibition of CYP3A4 activity. Prostate 2012, 72, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, C.; Nadiminty, N.; Lou, W.; Zhu, Y.; Yang, J.; Evans, C.P.; Zhou, Q.; Gao, A.C. Inhibition of Stat3 activation by sanguinarine suppresses prostate cancer cell growth and invasion. Prostate 2012, 72, 82–89. [Google Scholar] [CrossRef]
- Sun, M.; Lou, W.; Chun, J.Y.; Cho, D.S.; Nadiminty, N.; Evans, C.P.; Chen, J.; Yue, J.; Zhou, Q.; Gao, A.C. Sanguinarine suppresses prostate tumor growth and inhibits survivin expression. Genes Cancer 2010, 1, 283–292. [Google Scholar] [CrossRef]
- Bodle, C.R.; Mackie, D.I.; Hayes, M.P.; Schamp, J.H.; Miller, M.R.; Henry, M.D.; Doorn, J.A.; Houtman, J.C.D.; James, M.A.; Roman, D.L. Natural Products Discovered in a High-Throughput Screen Identified as Inhibitors of RGS17 and as Cytostatic and Cytotoxic Agents for Lung and Prostate Cancer Cell Lines. J. Nat. Prod. 2017, 80, 1992–2000. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Tang, D.; Wang, L.; Huang, L.Q.; Jiang, G.S.; Xiao, X.Y.; Zeng, F.Q. Gambogic acid inhibits TNF-alpha-induced invasion of human prostate cancer PC3 cells in vitro through PI3K/Akt and NF-kappaB signaling pathways. Acta Pharmacol. Sin. 2012, 33, 531–541. [Google Scholar] [CrossRef]
- Yi, T.; Yi, Z.; Cho, S.G.; Luo, J.; Pandey, M.K.; Aggarwal, B.B.; Liu, M. Gambogic acid inhibits angiogenesis and prostate tumor growth by suppressing vascular endothelial growth factor receptor 2 signaling. Cancer Res. 2008, 68, 1843–1850. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, G.; Tian, H.; Hu, Y.; Wu, S.; Geng, Y.; Lin, K.; Wu, W. Sulforaphane metabolites cause apoptosis via microtubule disruption in cancer. Endocr. -Relat. Cancer 2018, 25, 255–268. [Google Scholar] [CrossRef]
- Ferreira, P.M.P.; Rodrigues, L.; de Alencar Carnib, L.P.; de Lima Sousa, P.V.; Nolasco Lugo, L.M.; Nunes, N.M.F.; do Nascimento Silva, J.; da Silva Araujo, L.; de Macedo Goncalves Frota, K. Cruciferous Vegetables as Antioxidative, Chemopreventive and Antineoplasic Functional Foods: Preclinical and Clinical Evidences of Sulforaphane Against Prostate Cancers. Curr. Pharm. Des. 2018, 24, 4779–4793. [Google Scholar] [CrossRef]
- Gibbs, A.; Schwartzman, J.; Deng, V.; Alumkal, J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc. Natl. Acad. Sci. USA 2009, 106, 16663–16668. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.D.; Hsu, A.; Yu, Z.; Dashwood, R.H.; Ho, E. Differential effects of sulforaphane on histone deacetylases, cell cycle arrest and apoptosis in normal prostate cells versus hyperplastic and cancerous prostate cells. Mol. Nutr. Food Res. 2011, 55, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Kim, S.H.; Hahm, E.R.; Pore, S.K.; Jacobs, B.L.; Singh, S.V. Prostate cancer chemoprevention by sulforaphane in a preclinical mouse model is associated with inhibition of fatty acid metabolism. Carcinogenesis 2018, 39, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Beaver, L.M.; Lhr, C.V.; Clarke, J.D.; Glasser, S.T.; Watson, G.W.; Wong, C.P.; Zhang, Z.; Williams, D.E.; Dashwood, R.H.; Shannon, J.; et al. Broccoli Sprouts Delay Prostate Cancer Formation and Decrease Prostate Cancer Severity with a Concurrent Decrease in HDAC3 Protein Expression in Transgenic Adenocarcinoma of the Mouse Prostate (TRAMP) Mice. Curr. Dev. Nutr. 2018, 2, nzy002. [Google Scholar] [CrossRef]
- Yin, P.; Kawamura, T.; He, M.; Vanaja, D.K.; Young, C.Y. Phenethyl isothiocyanate induces cell cycle arrest and reduction of alpha- and beta-tubulin isotypes in human prostate cancer cells. Cell Biol. Int. 2009, 33, 57–64. [Google Scholar] [CrossRef]
- Sakao, K.; Desineni, S.; Hahm, E.R.; Singh, S.V. Phenethyl isothiocyanate suppresses inhibitor of apoptosis family protein expression in prostate cancer cells in culture and in vivo. Prostate 2012, 72, 1104–1116. [Google Scholar] [CrossRef]
- Kim, S.H.; Sehrawat, A.; Sakao, K.; Hahm, E.R.; Singh, S.V. Notch activation by phenethyl isothiocyanate attenuates its inhibitory effect on prostate cancer cell migration. PLoS ONE 2011, 6, e26615. [Google Scholar] [CrossRef]
- Hudson, T.S.; Perkins, S.N.; Hursting, S.D.; Young, H.A.; Kim, Y.S.; Wang, T.C.; Wang, T.T. Inhibition of androgen-responsive LNCaP prostate cancer cell tumor xenograft growth by dietary phenethyl isothiocyanate correlates with decreased angiogenesis and inhibition of cell attachment. Int. J. Oncol. 2012, 40, 1113–1121. [Google Scholar] [CrossRef]
- Powolny, A.A.; Bommareddy, A.; Hahm, E.R.; Normolle, D.P.; Beumer, J.H.; Nelson, J.B.; Singh, S.V. Chemopreventative potential of the cruciferous vegetable constituent phenethyl isothiocyanate in a mouse model of prostate cancer. J. Natl. Cancer Inst. 2011, 103, 571–584. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Ichikawa, H. Molecular targets and anticancer potential of indole-3-carbinol and its derivatives. Cell Cycle 2005, 4, 1201–1215. [Google Scholar] [CrossRef]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharm. Rev. 2011, 5, 1–12. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, D.H.; Jeong, J.H.; Guo, Z.S.; Lee, Y.J. Quercetin augments TRAIL-induced apoptotic death: Involvement of the ERK signal transduction pathway. Biochem. Pharm. 2008, 75, 1946–1958. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Budhraja, A.; Son, Y.O.; Wang, X.; Zhang, Z.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.C.; Xu, M.; et al. Quercetin inhibits angiogenesis mediated human prostate tumor growth by targeting VEGFR- 2 regulated AKT/mTOR/P70S6K signaling pathways. PLoS ONE 2012, 7, e47516. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Molecular targets for apigenin-induced cell cycle arrest and apoptosis in prostate cancer cell xenograft. Mol. Cancer Ther. 2006, 5, 843–852. [Google Scholar] [CrossRef]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; Maclennan, G.T.; Gupta, S. Apigenin inhibits prostate cancer progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef]
- Hafeez, B.B.; Siddiqui, I.A.; Asim, M.; Malik, A.; Afaq, F.; Adhami, V.M.; Saleem, M.; Din, M.; Mukhtar, H. A dietary anthocyanidin delphinidin induces apoptosis of human prostate cancer PC3 cells in vitro and in vivo: Involvement of nuclear factor-kappaB signaling. Cancer Res. 2008, 68, 8564–8572. [Google Scholar] [CrossRef]
- Henning, S.M.; Wang, P.; Heber, D. Chemopreventive effects of tea in prostate cancer: Green tea versus black tea. Mol. Nutr. Food Res. 2011, 55, 905–920. [Google Scholar] [CrossRef]
- Wang, P.; Henning, S.M.; Heber, D.; Vadgama, J.V. Sensitization to docetaxel in prostate cancer cells by green tea and quercetin. J. Nutr. Biochem. 2015, 26, 408–415. [Google Scholar] [CrossRef]
- Wang, P.; Henning, S.M.; Magyar, C.E.; Elshimali, Y.; Heber, D.; Vadgama, J.V. Green tea and quercetin sensitize PC-3 xenograft prostate tumors to docetaxel chemotherapy. J. Exp. Clin. Cancer Res. 2016, 35, 73. [Google Scholar] [CrossRef]
- Vodnik, V.V.; Mojic, M.; Stamenovic, U.; Otonicar, M.; Ajdzanovic, V.; Maksimovic-Ivanic, D.; Mijatovic, S.; Markovic, M.M.; Barudzija, T.; Filipovic, B.; et al. Development of genistein-loaded gold nanoparticles and their antitumor potential against prostate cancer cell lines. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 124, 112078. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Jiao, M.; Wu, D.; Wu, K.; Li, X.; Zhu, G.; Yang, L.; Wang, X.; Hsieh, J.T.; et al. Genistein inhibits the stemness properties of prostate cancer cells through targeting Hedgehog-Gli1 pathway. Cancer Lett. 2012, 323, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Adjakly, M.; Bosviel, R.; Rabiau, N.; Boiteux, J.P.; Bignon, Y.J.; Guy, L.; Bernard-Gallon, D. DNA methylation and soy phytoestrogens: Quantitative study in DU-145 and PC-3 human prostate cancer cell lines. Epigenomics 2011, 3, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Mentor-Marcel, R.; Lamartiniere, C.A.; Eltoum, I.E.; Greenberg, N.M.; Elgavish, A. Genistein in the diet reduces the incidence of poorly differentiated prostatic adenocarcinoma in transgenic mice (TRAMP). Cancer Res. 2001, 61, 6777–6782. [Google Scholar] [PubMed]
- Yo, Y.T.; Shieh, G.S.; Hsu, K.F.; Wu, C.L.; Shiau, A.L. Licorice and licochalcone-A induce autophagy in LNCaP prostate cancer cells by suppression of Bcl-2 expression and the mTOR pathway. J. Agric. Food Chem. 2009, 57, 8266–8273. [Google Scholar] [CrossRef]
- Shoulars, K.; Rodriguez, M.A.; Thompson, T.; Markaverich, B.M. Regulation of cell cycle and RNA transcription genes identified by microarray analysis of PC-3 human prostate cancer cells treated with luteolin. J. Steroid Biochem. Mol. Biol. 2010, 118, 41–50. [Google Scholar] [CrossRef]
- Kavitha, C.V.; Deep, G.; Gangar, S.C.; Jain, A.K.; Agarwal, C.; Agarwal, R. Silibinin inhibits prostate cancer cells- and RANKL-induced osteoclastogenesis by targeting NFATc1, NF-kappaB, and AP-1 activation in RAW264.7 cells. Mol. Carcinog. 2014, 53, 169–180. [Google Scholar] [CrossRef]
- Singh, R.P.; Deep, G.; Blouin, M.J.; Pollak, M.N.; Agarwal, R. Silibinin suppresses in vivo growth of human prostate carcinoma PC-3 tumor xenograft. Carcinogenesis 2007, 28, 2567–2574. [Google Scholar] [CrossRef]
- Shah, S.; Prasad, S.; Knudsen, K.E. Targeting pioneering factor and hormone receptor cooperative pathways to suppress tumor progression. Cancer Res. 2012, 72, 1248–1259. [Google Scholar] [CrossRef]
- Li, W.; Su, Z.Y.; Guo, Y.; Zhang, C.; Wu, R.; Gao, L.; Zheng, X.; Du, Z.Y.; Zhang, K.; Kong, A.N. Curcumin Derivative Epigenetically Reactivates Nrf2 Antioxidative Stress Signaling in Mouse Prostate Cancer TRAMP C1 Cells. Chem. Res. Toxicol. 2018, 31, 88–96. [Google Scholar] [CrossRef]
- Applegate, C.C.; Rowles, J.L., 3rd; Erdman, J.W., Jr. Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies. Nutrients 2019, 11, 633. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, M. Chemotherapy in Prostate Cancer. Curr. Oncol. Rep. 2015, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, R.; Berman-Booty, L.D.; Schiewer, M.J.; Ciment, S.J.; Den, R.B.; Dicker, A.P.; Kelly, W.K.; Trabulsi, E.J.; Lallas, C.D.; Gomella, L.G.; et al. Novel actions of next-generation taxanes benefit advanced stages of prostate cancer. Clin. Cancer Res. 2015, 21, 795–807. [Google Scholar] [CrossRef]
- Vrignaud, P.; Semiond, D.; Lejeune, P.; Bouchard, H.; Calvet, L.; Combeau, C.; Riou, J.F.; Commercon, A.; Lavelle, F.; Bissery, M.C. Preclinical antitumor activity of cabazitaxel, a semisynthetic taxane active in taxane-resistant tumors. Clin. Cancer Res. 2013, 19, 2973–2983. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Sanchez, M.G.; Malagarie-Cazenave, S.; Olea, N.; Diaz-Laviada, I. Induction of apoptosis in prostate tumor PC-3 cells and inhibition of xenograft prostate tumor growth by the vanilloid capsaicin. Apoptosis 2006, 11, 89–99. [Google Scholar] [CrossRef]
- Maina, S.; Misinzo, G.; Bakari, G.; Kim, H.Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules 2020, 25, 3682. [Google Scholar] [CrossRef]
- Livingstone, T.L.; Beasy, G.; Mills, R.D.; Plumb, J.; Needs, P.W.; Mithen, R.; Traka, M.H. Plant Bioactives and the Prevention of Prostate Cancer: Evidence from Human Studies. Nutrients 2019, 11, 2245. [Google Scholar] [CrossRef]
- Atwell, L.L.; Beaver, L.M.; Shannon, J.; Williams, D.E.; Dashwood, R.H.; Ho, E. Epigenetic Regulation by Sulforaphane: Opportunities for Breast and Prostate Cancer Chemoprevention. Curr. Pharm. Rep. 2015, 1, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Saw, C.L.; Cintron, M.; Wu, T.Y.; Guo, Y.; Huang, Y.; Jeong, W.S.; Kong, A.N. Pharmacodynamics of dietary phytochemical indoles I3C and DIM: Induction of Nrf2-mediated phase II drug metabolizing and antioxidant genes and synergism with isothiocyanates. Biopharm. Drug Dispos. 2011, 32, 289–300. [Google Scholar] [CrossRef]
- Hahm, E.R.; Chandra-Kuntal, K.; Desai, D.; Amin, S.; Singh, S.V. Notch activation is dispensable for D, L-sulforaphane-mediated inhibition of human prostate cancer cell migration. PLoS ONE 2012, 7, e44957. [Google Scholar] [CrossRef]
- Tang, N.Y.; Huang, Y.T.; Yu, C.S.; Ko, Y.C.; Wu, S.H.; Ji, B.C.; Yang, J.S.; Yang, J.L.; Hsia, T.C.; Chen, Y.Y.; et al. Phenethyl isothiocyanate (PEITC) promotes G2/M phase arrest via p53 expression and induces apoptosis through caspase- and mitochondria-dependent signaling pathways in human prostate cancer DU 145 cells. Anticancer Res. 2011, 31, 1691–1702. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Natural Compound | Constituent | Plant Source | Chemical Structure | Mechanism | Molecular Pathway (Reference) | In Vitro | In Vivo |
|---|---|---|---|---|---|---|---|
| 1. Polyphenol | |||||||
| 1. 1 Polyphenol Flavonols | |||||||
| 1.1.1 Quercetin | Flavonol | Apple and onion |  | Induction of apoptosis. | PI3K/Akt and MAPK/ERK [6]; p21, FA, Hsp90 [7]. | LNCaP, DU 145, PC-3, Prostate cancer stem cells. | |
| 1.1.2 Apigenin | Flavone | Anthemis sp. |  | Induction of apoptosis and cell cycly arrest. | IGF-IR; PI3k/Akt [8]; HDACs [9]; ROS [10]; VEGF [11]. | PC-3 and 22Rv1. | 22Rv1 and PC-3 xenografts. |
| 1.1.3 Baicalin | Flavone | Genus Scutellaria |  | Inhibition of proliferation and induction of apoptosis. | Caveolin-1/AKT/mTOR [12]; AR target genes [13]; Ezrin [14]. | DU 145 and PC-3. | PC-3 xenografts. |
| 1.1.4 Cyanidin-3-glucoside | Flavonoid anthocyanin | Berries, red cabbages, teas, and coloured grains. |  | Inhibition of EMT. | Snail/E-cadherin [15]. | PC-3. | |
| 1.1.5 Daidzein | Isoflavone | Soybeans |  | Induction of cell cycle arrest; CpG island demethylation. | CDK [16]; GSTP1 and EPHB2 [17]; ERβ [18]. | DU 145, LNCaP and PC-3. | |
| 1.1.6 Delphinidin | Flavonoid anthocyanin | Viola sp. |  | Inhibition of cell growth; Induction of apoptosis. | NF-κB [19]; β-catenin [20]; DR5 [21]; HDAC [22]. | PC-3 and LNCaP. | PC-3 xenografts. |
| 1.1.7 Epigallocatechin-3-gallate (EGCG) | Catechin | Green tea |  | Suppression of the proliferation, invasion and migration, and vasculogenic mimicry; Induction of apoptosis and autophagy. | ERK1/2 [23], VEGF, uPA, angiopoietin 1/2, MMP-2, and MMP-9 [24], PSA [25], Caspase 9 [26]; Twist/VE-Cadherin/AKT [27]; AR [28]; CHOP and p-eIF2α [29]. | LNCaP, PC-3, 22Rv1. | PC-3 xenografts. |
| 1.1.8 Fisetin | Flavonol | Acacia greggii |  | Inhibition of adhesion, migration, and metastasis; Induction of autophagy. | NF-κB [30]; mTOR and PI3K/Akt [31]; AR [32]; NF-κB [33]; Nudc [34]. | DU 145, LNCaP, PC-3. | CWR22 Rupsilon1 cells xenograft. |
| 1.1.9 Formononetin (FN) | Omethylated isoflavone | Trifolium pretense. |  | Induction of apoptosis. | ERK1/2, MAPK-Bax [35]; IGF-1/IGF-1R [36]; p38/Akt [37]; RASD1 [38]. | LNCaP, PC-3, and DU 145. | |
| 1.1.10 Genistein | Flavanone | Glycine max |  | Inhibition of the proliferation; Suppression of cancer stem cells; Activation of epigenetic modification; Induction of apoptosis. | IGF-1/IGF-1R [39]; miR-1260b [40]; miR-34a and HOTAIR [41]. | LNCaP, PC-3, and DU 145. | TRAMP mice. |
| 1.1.11 Glycyrrhiza | |||||||
| 1.1.11.1 Isoangustone A | Flavonoid | Glycyrrhiza uralensis |  | Induction of apoptosis; G1 phase arrest. | Caspase [42]; CDK2/4, cyclin A [43]. | DU 145. | |
| 1.1.11.2 Licoricidin | Flavonoid | Glycyrrhiza uralensis |  | Inhibition of metastasis and invasion. | MMP, TIMP-1, VEGF [44]. | DU 145. | |
| 1.1.12 Licochalcone | Flavonoid | Licorice root |  | Induction of G2/M phase arrest and apoptosis. | Cyclin B1 and cdc2 [45]. | LNCaP, PC-3. | |
| 1.1.13 Luteolin | Flavone | Terminalia chebula |  | Suppression of angiogenesis; Induction of cell cycle arrest. | IGF-1 [46]; VEGF-2R, AR [47]. | LNCaP and PC-3. | PC-3 xenografts. |
| 1.2 Polyphenol lignans | |||||||
| 1.2.1 Arctigenin | lignan | Arctium lappa |  | Inhibition of the proliferation; Induction of apoptosis and autophagy. | AR [48], FFA [49], PI3K/Akt/mTOR [50]. | LAPC-4 and LNCaP. | LAPC-4 xenograft. |
| 1.2.2 Honokiol | Lignin | Magnolia officinalis |  | Induction of G0-G1 phase arrest and apoptosis; Inhibition of angiogenesis. | Bax, Bak, Bad, Bcl-xL, and Mcl-1 [51,52]. | LNCaP, PC-3, and C4-2. | PC-3 xenografts. |
| 1.2.3 Magnolol | Hydroxylated biphenyl lignan | Magnolia officinalis |  | Induction of apoptosis and inhibition of the adhesion, invasion, and migration. | EGFR [53]. | PC-3. | |
| 1.2.4 Obovatol | Biphenyl ether lignan | Magnolia obovate |  | Induction of apoptosis. | NF-κB [54]. | LNCaP and PC-3. | |
| 1.2.5 Silibinin | Flavolignan | Silybum marianum |  | Induction of G1 phase arrest; Induction of apoptosis; Inhibition of EMT; Restriction of tumor microvessel density. | p21 and p27 [55]; NF-κB [56]; ZEB1 and SLUG [57]. | DU 145. | PC-3 xenografts and TRAMP mice [58]. |
| 1.3 Polyphenol Stilbenoids | |||||||
| 1.3.1 Resveratrol | Stilbenoid | Grape, raspberry, mulberry. |  | Induction of apoptosis; Cellular antioxidant defense. | FKHRL1 [59]; VEGF, MMPs [60]; CXCR4 [61]; ROS, RNS, HO-1 [62]; AR [63]. | TRAMP mouse [61]. | |
| 1.4 Other Polyphenols | |||||||
| 1.4.1 Curcumin | Polyphenols | Curcuma longa |  | Induction of apoptosis; Induction of autophagy. | EGF-R [64]; AIF [65]; Wnt/β-catenin [66]; Nrf-2 [67]; MMPs [68]. | DU 145, LNCaP and PC-3. | LNCaP xenograft [69]; DU 145 xenografts [68]. |
| 1.4.2 Ellagitannins | Polyphenols | Pomegranate fruit |  | Induction of apoptosis and S phase cell cycle arrest; Inhibition of metastasis and angiogenesis. | SIRT1, p21, AIF [70]; Eicosanoid [71]; cyclin B1 and D1 [72]. MMP-2 [73]; p53-MDM2 [74]. | LNCaP, 22RV1 and PC-3. | murine studies [75]. |
| 1.4.3 Gallic acid | Polyphenols | Grape seed |  | Induction of G2/M cell cycle arrest; Induction of apoptosis; Inhibition of invasion and migration. | cdc25A/C-cdc2 [76]; PARP [77]; p38, JNK, PKC, and PI3K/AKT [78]; Cdk, cyclin B1, and E [79]. | DU 145; PC-3. | TRAMP mice [79]; DU 145 and 22Rv1 xenografts [80]. |
| 1.4.4 Gossypol | Polyphenolic aldehyde | Cotton seed |  | Induction of G0/G1 phase arrest; Induction of apoptosis; Induction of autophagy; Inhibition of angiogenesis; Reduction of the microvessel density. | TGF-β1 and Akt [81]; Bcl-xL [82]; Beclin1 [83]. AP-1, NF-κB [84]. | MAT-LyLu; PC-3. | PC-3 xenografts [85]. |
| 2. Terpenoids | |||||||
| 2.1 Artemisinin | Sesquiterpene | Artemisia annua |  | Induction of G1 cell cycle arrest; Inhibition of viability. | CDK4 and Sp1 [86], caspases 8/9 [87]. | LNCaP, DU 145 and PC-3. | |
| 2.2 Betulinic Acid (BA) | Triterpene | Betula papyrifera |  | Induction of apoptosis; Inhibition of angiogenesis. | Bax/Bcl-2 [88], DUBs [89], AR and cyclin D [90]. | DU 145, LNCaP, and PC-3. | TRAMP mice [90]. |
| 2.3 Germacrone | Sesquiterpene | Geranium macrorrhizum. |  | Induction of apoptosis and autophagy. | Akt/mTOR [91]. | PC-3 and 22RV1. | |
| 2.4 Ginsenosides | Steroid glycosides and triterpene saponins | Genus Panax |  | Inhibition of proliferation; Inhibition of migration and metastasis; Induction of apoptosis. | p38 [92]; MAP [93]. | PC-3M, LNCaP and PC-3. | PC-3 xenograft [94]. |
| 2.5 Glycyrrhizin | Saponins | Fabaceae |  | Induction of apoptosis and autophagy. | Apoptosis and autophagy [95]. | PC-3. | |
| 2.6 Lycopene | Carotenoid | Tomatoes |  | Reduction in cholesterol synthesis; Induction of apoptosis. | CDK7, EGFR, IGF-1R, and BCL2 [96]; PPARγ-LXRα-ABCA1 [97]; CDK7,EGFR, IGF-1R, BCL2 [96]; IGF-I, IGF-IR, and IGFBP-3 [98]. | LNCaP, PC-3 and DU 145. | PC-3 xenograft [98]; TRAMP mice [99]. |
| 2.7 Oridonin | Isoprenoid | Rabdosia rubescens |  | Induction of apoptosis and G0/G1 cell cycle arrest; Induction of apoptosis, autophagy, and G2/M phase arrest. | p53 and Bax [100]; p21 [101]. | LNCaP and PC-3. | |
| 2.8 Thymoquinone (TQ) | Monoterpene | Nigella sativa |  | Induction of apoptosis; Inhibition of angiogenesis. | AR and E2F-1 [102]; ROS [103]; AKT, VEGF [104]. | LNCaP, PC-3, C4-B, and DU 145. | |
| 2.9 Ursolic acid (UA) | Pentacyclic triterpenoid | Cornus Officinalis |  | Induction of apoptosis. | Akt and MMP9 [105]; JNK [106]; CXCR4 [107]. NF-κB and STAT3 [108]; | PC-3, LNCaP, LNCaP-AI, and DU 145. | TRAMP mice [108]. |
| 2.10 β-Elemonic acid | Triterpene | Ganoderma tsugae, lucidum, and Boswellia. |  | Induction of apoptosis. | JAK2/STAT3/MCL-1 and NF-κB [109]. | 22RV1. | 22RV1xenograft [109]. |
| 3. Taxanes | |||||||
| 3.1 Cabazitaxel | Taxane | European yew tree |  | Inhibition of proliferation. | P-gp [110]. | C4-2. | |
| 3.2 Docetaxel | Taxane | European yew tree |  | Induction of apoptosis. | p38/p53/p21 [111]. | LNCaP, PC3 and DU 145. | |
| 3.3 Paclitaxel | Taxane | Pacific yew tree |  | Induction of apoptosis. | Bcl-2 [112]; AR [113]. | C4-2. | 22Rv1 xenografts [113]. |
| 4. Alkaloids | |||||||
| 4.1 Anibamine, | Pyridine quaternary alkaloid | Aniba sp. |  | Inhibition of proliferation. | CCR5 [114]. | LNCaP. | |
| 4.2 Berberine | Isoquinoline Alkaloid | Genus Berberis |  | Induction of G1 phase arrest; enhancement of the radiosensitivity. | p53-p21 [115], AR [116], MAPK/caspase-3 and ROS [117]. | RM-1. | LNCaP xenografts [116]. |
| 4.3 Capsaicin | Alkaloid | Red pepper |  | Induction of apoptosis. | JNK and ERK [118]. p53, p21, and Bax, AR [119]. | LNCaP, PC-3, and DU 145. | |
| 4.4 Neferine | Bisbenzylisoquinoline alkaloid | Nelumbo nucifera |  | Inhibition of proliferation and migration of prostate cancer stem cells. | p38 mapk/jnk [120]. | PC3. | |
| 4.5 Piperine | Alkaloid | Black pepper |  | Induction of apoptosis, autophagy, and G0/G1 phase arrest. | NF-kB, STAT-3 [121]. | LNCaP, PC-3, and DU 145. | Animal xeno-transplanted model [122]; CRPC xenograft model [123]. |
| 4.6 Sanguinarine | Alkaloid | Sanguinaria Canadensis |  | Inhibition of ubiquitin-proteasome system. | Stat3 [124], survivin [125], RGS17 [126]. | DU 145, C4-2B, and LNCaP. | DU 145 xenografts [125]. |
| 5. Other | |||||||
| 5.1 Gambogic acid | Xanthone | Garcinia hanburyi |  | Inhibition of angiogenesis. | PI3K/Akt and NF-κB [127]; VEGF-2R [128]. | PC-3. | PC-3 xenograft model [128]. |
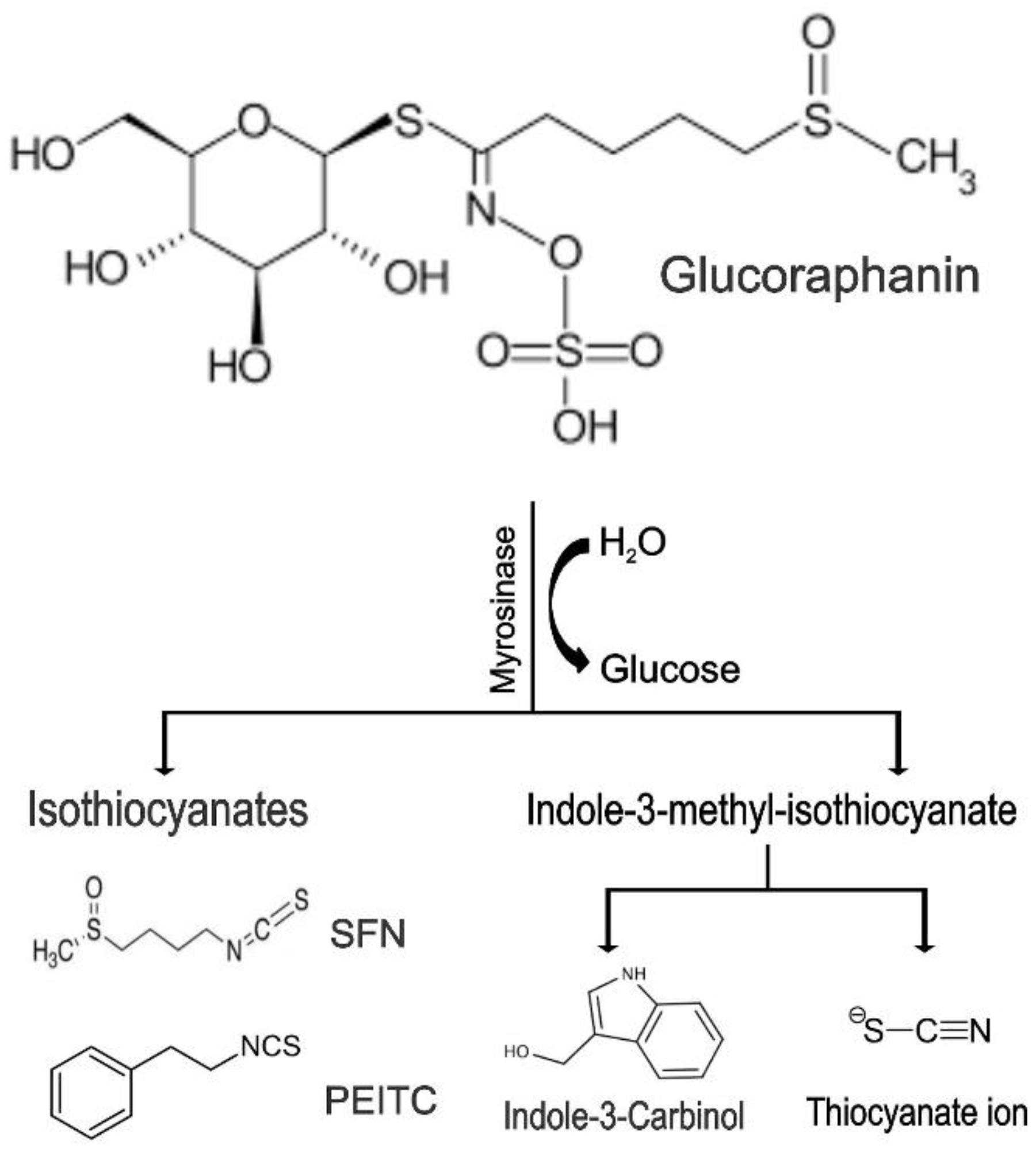
| 5.2 Glucoraphanin → Thiocyanates + Isothiocyanates + Indoles | |||||||
| 5.2.1 Sulforaphane (SFN) | Isothiocyanates (ITCs) | Cruciferous vegetables |  | Induction of apoptosis; Induction of G2/M cell cycle arrest; Inhibition of FA metabolism. | ERK1/2 [129]; Hsp90 [130]; AR [131]; ROS [132]; FA [133]; HDACs [134]. | LNCaP, DU 145 and PC-3. | TRAMP mice [134]. |
| 5.2.2 Phenethyl-Isothiocyan (PEITC) | Isothiocyanates (ITCs) | Cruciferous vegetables |  | Induction of G2/M cell cycle arrest; Induction of apoptosis; Inhibition of angiogenesis. | α- and β-tubulin [135]; Bax [136]; Akt [137]; PECAM1-CD31 [138]. | C4-2B, DU 145, PC-3, and LNCaP. | LNCaP xenograft model [138]; TRAMP mice [139]. |
| 5.2.3 Indole-3-Carbinol | Indoles | Cruciferous vegetables |  | Induction of apoptosis and cell cycle arrest; Modulation of epigenetic alterations of cancer stem cells. | Bax [140]; NF-ĸB, Nrf2 [140]. | LNCaP, and PC-3. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Q.; Wu, Y.; Vadgama, J.V.; Wang, P. Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies. Biomolecules 2022, 12, 1306. https://doi.org/10.3390/biom12091306
Hao Q, Wu Y, Vadgama JV, Wang P. Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies. Biomolecules. 2022; 12(9):1306. https://doi.org/10.3390/biom12091306
Chicago/Turabian StyleHao, Qiongyu, Yanyuan Wu, Jaydutt V. Vadgama, and Piwen Wang. 2022. "Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies" Biomolecules 12, no. 9: 1306. https://doi.org/10.3390/biom12091306
APA StyleHao, Q., Wu, Y., Vadgama, J. V., & Wang, P. (2022). Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies. Biomolecules, 12(9), 1306. https://doi.org/10.3390/biom12091306