Abstract
There have been magnificent advancements in the understanding of molecular mechanisms of chronic diseases over the past several years, but these diseases continue to be a considerable cause of death worldwide. Most of the approved medications available for the prevention and treatment of these diseases target only a single gene/protein/pathway and are known to cause severe side effects and are less effective than they are anticipated. Consequently, the development of finer therapeutics that outshine the existing ones is far-reaching. Natural compounds have enormous applications in curbing several disastrous and fatal diseases. Oroxylin A (OA) is a flavonoid obtained from the plants Oroxylum indicum, Scutellaria baicalensis, and S. lateriflora, which have distinctive pharmacological properties. OA modulates the important signaling pathways, including NF-κB, MAPK, ERK1/2, Wnt/β-catenin, PTEN/PI3K/Akt, and signaling molecules, such as TNF-α, TGF-β, MMPs, VEGF, interleukins, Bcl-2, caspases, HIF-1α, EMT proteins, Nrf-2, etc., which play a pivotal role in the molecular mechanism of chronic diseases. Overwhelming pieces of evidence expound on the anti-inflammatory, anti-bacterial, anti-viral, and anti-cancer potentials of this flavonoid, which makes it an engrossing compound for research. Numerous preclinical and clinical studies also displayed the promising potential of OA against cancer, cardiovascular diseases, inflammation, neurological disorders, rheumatoid arthritis, osteoarthritis, etc. Therefore, the current review focuses on delineating the role of OA in combating different chronic diseases and highlighting the intrinsic molecular mechanisms of its action.
1. Introduction
Chronic disease or non-communicable disease is an umbrella term that is used to define a large number of ailments, including cancer, cardiovascular diseases (CVDs), chronic respiratory diseases, and diabetes [1,2]. Despite remarkable improvements in the prevention and therapy of multigenic chronic diseases, their prevalence among patients has not reduced [1]. Lack of physical activity, tobacco and alcohol intake, poor or unhealthy diet, etc., are the foremost risk factors that catalyze the development and progression of these ailments [2,3]. Most of these diseases are complex and occur through alterations in multiple signaling pathways; hence, targeting a single pathway could not necessarily control disease development [4,5,6,7]. Therefore, drug combinations addressing many molecular abnormalities or disease hallmarks may be used to treat these diseases [8,9,10]. However, these drug combinations are known to induce severe adverse side effects in patients and affect their quality of life.
Consequently, there exists an increasing need for the development of safer, efficacious, multi-targeted, and affordable therapeutic regimens to supersede the extant toxic and less efficient treatment strategies [11,12,13]. It is well acknowledged that medicinal plants have immense potential for prophylaxis of multiple chronic diseases, including cancer [14,15,16,17,18,19,20,21,22]. A mounting number of preclinical and clinical evidence suggest that natural compounds extracted from various plants are plausible candidates against a multitude of life-threatening chronic diseases and different formulations can be used to increase their bioavailabilities [2,23,24,25,26,27,28,29,30,31,32,33,34,35]. OA is one such compound, which has gathered attention among scientific communities due to its remarkable multi-targeted properties in the prophylaxis and regimen of various non-communicable diseases. OA is an O-methylated flavone found mainly in Oroxylum indicum Scutellaria baicalensis and S. lateriflora. [36,37,38].
O. indicum has been an inevitable component in Asian ethnomedicinal systems since time immemorial for the treatment of various disorders, which include arthritic and rheumatic problems, diabetes, diarrhea, dysentery, gastric ulcers, jaundice, respiratory diseases, and tumors [39,40]. The tonic of this plant is used against anorexia, asthma, bronchitis, cough, dysentery, dyspepsia, fever, gout, leucoderma, neuralgia, rheumatoid arthritis, vomiting, and wounds. The root bark is used for cancer, stomatitis, and tuberculosis [40,41,42]. O. indicum is an important constituent in various Ayurvedic preparations, such as Narayana Taila, Dasamularistha, Dhanawantara Ghrita, Syonaka putapaka, Dantyadyarista, Syonaka sidda ghrta, Amartarista, Brahma Rasayana, Brhatpancamulyadi kvatha, and Chyavanaprasa [43,44,45].
Similarly, S. baicalensis and S. lateriflora have immense therapeutic potential and had been in medicinal use since ancient times. The genus Scutellaria is widely employed in Traditional Chinese Medicine (TCM) for treating an array of diseases, including diarrhea, dysentery, hepatitis, high blood pressure, and vomiting [46]. S. lateriflora possesses anxiolytic and anti-convulsant potential and was used as a nervine tonic traditionally [47,48]. The herb also has anti-oxidative and DNA-protecting effects [49]. The major flavonoids seen in the above plants, namely OA, baicalein, chrysin, and wogonin, have potential alleviatory effects against several life-threatening chronic diseases [50,51].
2. Isolation and Purification of OA
OA is mainly found in the root-bark O. indicum, S. baicalensis (radix), S. lateriflora, Anchietea pyrifolia, and Aster himalaicus, which are used extensively in Ayurveda and TCM [40]. The crude extract of OA was first isolated from O. indicum using alcohol percolation and distillation [52]. Later, OA was isolated by acetone extraction followed by crystallization, which gave rise to the yield of only 0.86. More recently, Li and Chen isolated and purified OA from S. radix using ethyl ether and hexane extraction and subsequently performed high speed counter current chromatography and high-performance liquid chromatography (HPLC) to obtain 93.2% purity [37]. This is currently the most widely used method for the isolation and purification of OA. Moreover, the isolation of OA of purity of more than 99% was shown to be achievable with the use of modern chromatography technologies, such as HPLC, thin-layer high performance liquid chromatography, and silica gel chromatography [53,54,55].
3. Structural Analysis
Structurally, OA is a 5, 7-dihydroxy-6-methoxy-2-phenylchromen-4-one and its molecular weight is about 284.26 g/mol. It is a monomethoxy and dihydroxy flavone in which two -OH groups are positioned at carbon-5 and carbon-7, and one methoxy group is at carbon-6 (Figure 1) (PubChem CID: 5320315). The two metabolites of OA are oroxylin A 7-O-β-D-glucuronide or oroxyloside (OAG) and oroxylin A sodium sulfonate (OS) (Figure 1). OAG (PubChem CID 14655552) is a monomethoxy or monohydroxy flavone derived from OA. A plethora of studies revealed the anti-bacterial, anti-viral, anti-oxidant, anti-inflammatory, anti-invasive, neuroprotective, hepatoprotective, and pro-apoptotic properties of OA, which buttresses its promising potential in the treatment of diseases [56,57,58,59,60,61,62]. Therefore, the current study recapitulates the prospects of OA for the prevention and therapy of multiple chronic diseases.
Figure 1.
Role of oroxylin A and its metabolites in treating different chronic diseases.
4. Molecular Targets of OA
It has been well established that natural products modulate multiple signaling pathways involved in the development of chronic diseases, which leads to their efficacy in preclinical and clinical studies [63,64,65,66,67]. Accumulating evidence has shown that the exceptional potential of OA in the prevention and treatment of severe chronic diseases, such as CVDs, diabetes, neurological diseases, inflammatory diseases, cancer, etc., by the modulation of multiple pathways [40,68]. Copious pre-clinical studies explicated the tremendous potential of OA as an anti-inflammatory agent. For instance, OA inhibited the expression of several pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukins (IL-1β, IL-4, IL-6, IL-13) [69,70]. Apart from these, OA was also shown to reduce the expression of enzymes, including cyclooxygenase 2 (cox-2), inducible nitric oxide synthase (iNOS), glycogen synthase kinase 3-β (GSK-β), lactate dehydrogenase (LDH), pyruvate kinases isozymes (PKM1/PKM2), etc., which envisages its anti-inflammatory properties [71,72].
Further, the immense antioxidant properties of OA are also well documented. For example, Wang and colleagues demonstrated that OA and OAG prevented FeSO4-induced lipid peroxidation in liver homogenate and these compounds have substantial cytoprotective effects against H2O2-induced oxidative damage in human umbilical vein endothelial cells [73]. In another study, PC12 cells pre-treated with OA were exposed to H2O2, which resulted in a notable depletion in the intracellular calcium and ROS levels and an increase in the mRNA level of Mn/SOD. Hence, it could be inferred that OA pre-treatment hampered H2O2-induced oxidative stress [74]. Apparently, OA regulates the expression of multiple proteins, such as transforming growth factor beta (TGF-β), nuclear factor E2-related factor 2 (Nrf2), mitofusin 2 (Mfn2), angiopoietin 2 (Ang-2), vascular endothelial growth factor (VEGF), glutathione (GSH), B-cell lymphoma 2 (Bcl-2), Bcl-2-associated X protein (Bax), caspase -3, -8 and -9, sirtuin 3 (SIRT3), alpha-smooth muscle actin (α-SMA), etc., [75,76,77,78,79]. The proteomic screening of cancer cells treated with OA has revealed that it downregulates several factors, such as mitochondrial uncoupling protein 2 (UCP2), MMP-2, MMP-9, PKM2, superoxide dismutase 2 (SOD2), hypoxia inducible factor 1 alpha (HIF-1α), and PROX1 [62,80,81].
Subsequently, OA could downregulate different signaling pathways, including the nuclear factor kappa B (NF-κB) pathway, the signal transducer, and the activator of transcription 3 (STAT3) pathway; Wnt/β-catenin pathway and Notch-1 that have been implicated in tumorigenesis [82,83,84,85,86,87,88,89,90,91]. Likewise, OA also regulates other pathways, such as endoplasmic reticulum (ER) stress-mediated pathway, phosphatase, and tensin homolog deleted on the chromosome 10/phosphatidylinositol 3-kinase/a serine threonine-protein kinase (PTEN/PI3K/Akt) pathway, the extracellular signal-regulated kinase (ERK 1/2) pathway, etc., [62,83,92,93,94,95,96].
5. OA for Cancer
Cancer is one of the leading causes of illness and fatalities with about 19.3 million new cases being diagnosed annually worldwide and resulting in approximately 10 million deaths [97]. Several innovative therapeutics, including targeted therapy, therapeutic repurposing, oncolytic virotherapy, immunotherapy, etc., have been employed in the treatment of this disease; however, their long-term uses are not devoid of life-threatening side effects [98,99,100,101,102,103,104,105,106,107,108]. Recently, a very large body of literature has emerged on the promising anti-cancer effects of the natural compound OA. The therapeutic effects of OA on various cancers (both in vitro and in vivo studies) are listed in Table 1. Breast cancer is the foremost cause of mortalities due to cancer among women with over 2 million cases diagnosed each year worldwide [109,110,111,112,113]. Many studies have augmented the potential of OA as a candidate for breast cancer treatment. For instance, a study found that OA inhibits the proliferation of human breast cancer cells and reduced the tumor mass and volume in breast cancer xenograft models, indicating that it has anti-cancer properties. Further, under hypoxia, OA lowered cellular oxidative stress via upregulating SIRT3, which leads to HIF-α destabilization and increased prolyl hydroxylase activity. Furthermore, OA elevated SOD2 gene expression and activity through SIRT3-mediated de-acetylation [76]. Another study found that OA suppressed cell proliferation, cell cycle progression, migration, and epithelial-mesenchymal transition (EMT) in breast cancer cells by downregulating the NF-κB pathway [114].
The incidence and progression of colon cancer tend to be deleterious to human health and well-being [115,116]. The timely diagnosis of cancer development and metastasis is very important for appropriate therapy and prognosis [117]. Accumulating number of studies has proved the promising effects of OA against colon cancer. For example, in a study it was found that OA elevated the expression of caspases 3 and 9, which are crucial mediators of apoptosis along with inhibition of the regulator of apoptosis, Bcl-2. In addition, the ROS levels and the Nrf2 expression were increased by OA [56]. In another study, OA and 5-fluorouracil (5-FU) synergistically resulted in the suppression of Bcl-2 and activation of p53, Bax, and procaspase-3 in human colon cancer cells, which showed an antitumor effect of OA in combination with 5-FU [118]. Additionally, in vivo studies in colon cancer xenograft suggested that a high-fat diet accelerated tumor development in the colon and OA decreased intracellular fatty acid levels, and hence caused fatty acid oxidation by inactivating HIF-1α. Therefore, OA caused the reprogramming of fatty acid metabolism of HCT 116 cells and can be a promising agent in the prevention of colon cancer [82]. OA had shown intriguing antitumor effects against hepatocellular carcinoma cells. For instance, a study revealed that OA in combination with 5-FU showed a higher inhibitory rate in H22 murine solid tumors than the 5-FU alone. OA also decreased the expression of cox-2, Bcl-2, and procaspase-3 and increased the expression of p53 [119]. In another study, OA showed inhibitory effects on TGF-β1/SMAD-induced EMT in HCC cells and elevated non-steroidal anti-inflammatory drug activated gene-1 (NAG1) [120].
Gliomas are the most common primary tumors of the central nervous system with characteristic genetic and epigenetic profiles [121,122]. Despite the advances in therapy, tumor stem cells (TSCs) develop chemo- and radio-resistance, resulting in disease recurrence [123,124,125]. However, a growing body of research suggests that OA could be a potential therapeutic agent for glioma. For example, OA-reduced Notch-1 and myeloid cell leukemia 1 expression (Mcl-1) and inhibited Akt and ERK activation in these cells. Moreover, this compound increased the expression of Beclin 1, a crucial autophagy-related protein, resulting in autophagy [126]. In another study, OA was shown to suppress IP3R1 Akt/β-catenin pathway remarkably, which resulted in sensitizing glioma cells to temozolomide (TMZ) [94].
Hematological malignancies, such as leukemia, lymphoma, and multiple myeloma are among the life-threatening cancers worldwide [127,128]. However, OA appears to be a likely agent for the treatment of these malignancies. For instance, OA sensitized acute myeloid leukemia (AML) cells to TNF-α [83]. Moreover, OA inhibited the PI3K/Akt pathway and tRXRα in NB4 and HL-60 cells [83]. Similarly, OA improved CD11b/CD14 expression of AML/ETO- positive cells but downregulated histone de-acetylase 1 (HDAC-1) protein levels in t (8i21)-positive AML cells. Further, OA enhanced C/EBPα and p21 expression. Taken together, this study proved that OA as a promising candidate for AML1/ETO-positive AML differentiation therapy [129]. In addition, studies have proved the efficacy of OA against chronic myeloid leukemia (CML). For example, Li and his co-workers (2017) proved that OA could reverse imatinib resistance and induce apoptosis in CML via suppressing the CXCL12/CXCR7 pathway and the expression of p-ERK [130].
Lung cancer is a major reason of death in both men and women globally [131,132,133]. Many studies explored the potential of OA in lung cancer therapeutics. For example, OA suppressed regulatory T-cells (Tregs) generation in lung cancer cells by inhibiting the secretion of TGF-β1 and downregulating NF-κB signaling in H460 cells [134]. Another study revealed that OA in combination with cisplatin reversed hypoxia-induced cisplatin resistance in lung cancer cell lines [135].
Esophageal squamous cell carcinoma (ESCC) is estimated to be the sixth primary cause of cancer deaths globally with a high rate of fatality [97]. Intriguingly, several studies have reported that OA could enhance the susceptibility of ESCC cells to X-ray radiation and hence OA could be an effective radiosensitizer [136]. Besides, OA has shown promising effects against skin cancer progression. In a recent study, OA was shown to decrease the inflammatory factors and hyperplasia via the suppression of NF-κB signaling and SHC SH2 domain-binding protein 1 (SHCBP1) in skin cancer in vitro and in vivo models [137]. In addition, fruitful results have been obtained from studies using OA against cervical cancer, which is one of the major causes of cancer deaths in women around the world. For instance, in a study, OA induced apoptosis of cervical tumor cells and suppressed Bcl-2 thereby decreasing tumorigenesis [75]. Further, another study investigated the effects of OA against Kaposi’s sarcoma and found that OA inhibited the invasion and neovascularization of lymphatic phenotype endothelial cell line generated by the infection of Kaposi’s sarcoma-associated herpes virus (KSHV vIL-6) [80]. These studies collectively endorse the anti-cancer properties of OA.
6. OA for Cardiovascular Diseases (CVDs)
CVDs such as acute coronary artery syndrome, atherosclerosis, cardiac arrest and arrythmias are a major health concern worldwide [138,139,140]. Hence, the development of intervention strategies with low cost and high efficacy is of paramount importance. OA has been reported to have prolific effects on various chronic cardiac ailments (Table 1). Doxorubicin (DOX) is a quinone-bearing anthracycline used to treat various hematological malignancies, but its use has deleterious effects on the heart resulting in a decreased number of cardiomyocytes and congestive heart failure, limiting DOX’s therapeutic application [141,142]. Fortuitously, OA was shown to have cardioprotective effects against the damages caused by DOX. For instance, a study that investigated the potential cardioprotective activity of OA revealed that it activated sirtuin 1 via the cAMP/protein kinase A pathway. As a result, OA prevented DOX-induced reduction in cardiac function, heart weight loss, and myocardial apoptosis and prevented heart injury [143].
7. Endotoxemia
Metabolic endotoxemia is caused by an increased level of plasma lipopolysaccharide (LPS), which ultimately results in metabolic disorders [144]. Metabolic endotoxemia also causes low-grade inflammation, which ultimately leads to chronic diseases, such as non-alcoholic fatty liver disease (NAFLD), type 2 diabetes mellitus (T2DM), chronic kidney disorders, and atherosclerosis [144,145,146,147,148,149]. Interestingly, OA treatment was shown to elevate coronary flow and cardiac function in LPS-induced endotoxemia mice. Therefore, it can be concluded that OA is a promising candidate for the treatment of myocardial dysfunction [55].
8. Hind Limb Ischemia (HLI)
Peripheral artery disease results in ischemia due to artery obstruction; hence, increasing angiogenesis is a crucial mechanism for revitalizing blood flow to the limb in response to ischemia [150,151,152]. OA has shown beneficial effects in angiogenesis and blood flow recovery by elevating VEGFA, angiopoetin-2 (Ang-2), fibroblast growth factor (FGF-2), platelet-derived growth factor (PDGF-BB) levels and promoting endothelial cell (EC) proliferation and migration. Further, OA has also been shown to downregulate macrophages and neutrophils, thereby opening new possibilities in the treatment of HLI [153].
9. OA for Chronic Liver Diseases
Liver diseases pose a major threat to health and are considered among common non-cancerous related deaths worldwide [154,155]. The major types of chronic liver diseases include alcoholic liver disease (ALD), non-alcoholic fatty liver disease (NAFLD), liver cirrhosis, and hepatocellular carcinoma (HCC) [155]. Nowadays, natural compounds are a valuable source of anti-fibrotic therapeutics and OA has also been proved as an effective candidate against chronic liver diseases (Table 1) [148,156,157,158]. For instance, the hepatoprotective effect of OA was investigated by administering OA to mice with CCl4-induced liver injury [69]. The expression of IL-1Rα, which acts as acute-phase protein (APP) in the initial events of liver regeneration, was found to be increased, but the mRNA levels of IL-6 and TNF-α were found to decline rapidly after treatment with OA [69]. Similarly, another study was conducted to identify the effects of OA against LPS and/or D-galactosamine-induced acute liver injury in mice. In this study, OA decreased the levels of TNF-α, alanine amino transferase (ALT), aspartate amino transferase (AST), and hepatic malondialdehyde content, which are markers of hepatic oxidative stress. Further, OA downregulated NF-κB and toll-like receptor (TLR4) pathway and upregulated Nrf 2 and heme oxygenase (HO-1), which undoubtedly proved that OA reversed the effects of acute liver injury [159].
9.1. Liver Fibrosis
Hepatic fibrosis or simply liver fibrosis is a chronic disease resulting from the long-term activation of physical, biochemical, or microbial stimuli in liver cells. The disease is marked by abnormal fibroblast accumulation and excessive extracellular matrix (ECM) deposition, as well as visible inflammatory lesions and structural changes [154,160]. Hepatic fibrosis ultimately results in liver cirrhosis and hepatocellular carcinoma. However, many studies have revealed that hepatic fibrosis is a compensatory repair mechanism in chronic liver diseases, and hepatic stellate cells (HSCs) cause the initiation and development of hepatic fibrosis [154,161]. A lot of evidence suggests that OA has a significant impact on liver fibrosis and associated inflammation. For instance, in a study, the anti-inflammatory effect of OA was investigated in activated HSCs. OA downregulated PI3/Akt/mTOR pathway by scavenging ROS. Further, OA also hinders the secretion of pro-inflammatory cytokines and caused autophagy in activated HSCs [92]. In another study, the effect of OA was studied in a carbon tetrachloride (CCl4)-induced liver fibrosis mice model. As a result, OA markedly repressed alkaline phosphatase (ALP), AST, and ALT, which are liver injury markers. Besides, OA inhibited the expression of α-1 collagen, fibronectin, α-SMA, platelet-derived growth factor beta receptor (PDGF-βR) and TGF- β R1 in the murine model of liver fibrosis induced by CCl4. Further, OA increased the expression of the autophagy markers, including Atg3, Atg6, Atg7, Atg4, Atg5, Atg9, Atg12, Atg14, and microtubule-associated proteins 1A/1B light chain 3B (LC3-B), along with Beclin 1 in both CCL4-induced murine model and HSCs. This study also proved the potential anti-fibrosis effect of OA and unveiled that autophagy is required for OA to eliminate hepatic fibrosis [162]. In another study, the effect of OA on the contraction of HSCs was explored and the results showed that OA hindered HSC contraction by blocking the aerobic glycolytic pathway. OA was shown to considerably reduce glucose uptake and lactate production, hexokinase 2 (HKII), phosphofructokinase 1 (PFK1) and PKM2 levels and the mRNA expression of lactate dehydrogenase-A (LDH-A) [163]. Therefore, it can be concluded that OA could be a potential therapeutic candidate against liver fibrosis and injury.
Loss of lipid droplets (LDs) is an important feature of liver fibrosis [164]. Therefore, the effect of OA on the disappearance of lipid droplets was examined in a study. Intriguingly, OA treatment considerably declined the expression of adipose triglyceride lipase (ATGL), which catalyzes lipolysis. Further, the effect of OA was accelerated by ROS-specific scavenger N-acetyl cysteine (NAC). This study portrayed the anti-fibrosis effect of OA [79]. In another study, OA treatment reduced cell proliferation and fibrogenesis but induced caspases and endoplasmic reticulum stress (ERS)-related proteins, resulting in the cell cycle arrest of HSCs. It was thus proved that ERS pathway activation was required for OA to induce apoptosis in HSC. Therefore, it can be concluded that OA has a therapeutic role in hepatic fibrosis via ERS activation [165].
9.2. Alcohol Liver Disease (ALD)
ALD is a complex disease caused by overconsumption of alcohol and is marked by a varied range of liver disorders, including liver cirrhosis, steatosis, and HCC [166,167]. Abstaining from alcohol must be the major objective of patients suffering from ALD, so that the condition would not progress into severe cirrhosis and ensure a longer survival rate [168]. The inhibitory effects of OA against ALD and its associated mechanisms have been studied profoundly. For example, in a study, OA was found to reduce the number of SA-β-gal-positive LO2 cells and inhibited cellular senescence in ethanol-treated hepatocytes via the activation of the YAP pathway and decreasing the expression of p16, p12, and HMGA1, which are important senescence markers [169]. In a different study, this compound suppressed pyroptosis, a type of programmed cell death seen in ALD through the NLR inflammasome dependent-canonical caspase 1 pathway. Further, OA also improved proliferator activator receptor gamma co-activator 1 alpha (PGC-1α), which is a major mitochondrial regulator and promotes the transcription of Mfn2. Taken together, this study proved that OA can prevent ALD via PGC-1α/Mfn2 signaling [77]. In addition, Jin and his co-workers (2018) elucidated the effect of OA against alcohol-induced hepatic steatosis where human hepatocyte LO2 cells were cultured and stimulated with ethanol to induce damage and the treatment with OA lowered lipid droplet accumulation and nuclear translocation of HIF-1α. However, the activation of HIF-1α reduced the effect of OA on lipid droplets accumulation in this model [170].
10. OA for Eye Diseases
Retinal ganglion cells are located in the inner retina and their axons comprise the optic nerve, which transports visual information to the brain. Several disorders of the visual system cause functional and/or anatomical changes in retinal ganglion cells (RGCs) (i.e., ischemic optic neuritis, demyelinating optic neuritis, diabetic retinopathy, glaucoma) [171,172,173,174,175]. Several studies have investigated the neuroprotective functions of OA and their effect on the survival of RGCs. For instance, OA decreased the number of ED1 positive cells at the lesion site in the rat optic nerve crush model. In addition, the expression of the glial fibrillary acidic protein (GFAP) was also decreased substantially in the OA-treated group. Further, OA also reduced iNOS and cox-2 expression in retinas. Taken together, this study proved the neuroprotective effect of OA on retinal ganglion cells [176].
A common cause of non-glaucomatous optic neuropathy in middle-aged and older persons is non-arteritic anterior ischemic optic neuropathy (NAION), which causes irreversible vision loss [177,178]. In a very recent study, OA was found to be effective against ischemic injury. OA markedly decreased the apoptosis of RGCs and optic disc edema and upregulated the Nrf2 signaling pathway and its downstream antioxidant enzymes NAD(P)H: quinone oxidoreductase (NQO-1) and HO-1 in the retina. Hence, OA can be efficiently used as a therapeutic drug candidate in the NAION [179].
11. OA for Inflammatory Diseases
11.1. Allergic Asthma
Asthma is a chronic disease of the airways, which results in chest tightness, wheezing, and coughing as a result of occasional airflow restriction and airway inflammation. Thickening and constriction of bronchi, as well as increased mucus production and edema, which occurs due to inflammatory and structural changes throughout the airway wall, contribute to episodes of obstruction of airflow during asthma [180,181]. Airway smooth muscle (ASM) thickening through hyper-responsiveness and remodeling, poor relaxation, and persistent airflow blockage may also lead to asthma [182]. Many studies have delineated the therapeutic effects of OA against allergic asthma. For instance, in a study, OA was administered by oral gavage in an ovalbumin (OVA)-induced allergic asthma model (BABL/c mice). OA elevated the number of inflammatory cells and airway hyper responsiveness but suppressed OVA-induced NF-κB activation. The study envisages OA as a therapeutic drug for the treatment of allergic asthma [183]. In addition, the anti-allergic and anti-inflammatory effects of OA were studied in vitro in rat RBL-2H3 mast cells and in vivo in a murine-ovalbumin-induced allergic asthma model and the β-hexosaminidase activity was measured in vitro and the results showed that OA reduced the expression of IFNγ, IL-4, and IL-13 and suppressed inflammation and mucin production in lungs. Hence, this study proved the promising anti-allergic effects of OA [70].
11.2. Inflammatory Bowel Disease (IBD)
IBD is a chronic inflammatory disease that includes two types of diseases, including Crohn’s disease and ulcerative colitis, and is manifested by prolonged stomach pain and diarrhea [184,185,186]. These characteristic symptoms are caused due to the reduced efficacy of the epithelial barrier and the colossal infiltration of immune cells into the intestinal tract and due to the disrupted immune response to commensal flora (gut microflora that is resident inside the human intestine) [186,187,188]. Congregate evidence suggests that OA can be an alternative therapy for IBD. For instance, Bai and his colleagues investigated the inhibitory effects of OA on low-grade colonic inflammation caused by fiber deficiency in the diet. OA allayed colitis and inhibited colitis-associated colon cancer development in mice. OA increased the amount Eubacterium coprostanoligenes (a probiotic gut bacteria), thereby resulting in an anti-inflammatory effect [189]. In another study, OA has been reported to ameliorate IBD via inhibiting pro-inflammatory cytokines, such as IL-1β, IL-6, TNF-α, and the activation of NLR family pyrin domain containing 3 (NLRP3) inflammasome in dextran sodium sulfate (DSS)-induced murine model [190]. Furthermore, the effect of OAG was also investigated in DSS-induced colitis and analyzed its anti-inflammatory effects. OAG was observed to reduce myeloperoxidase (MPO) and iNOS activities and decreased inflammatory cell infiltration. Moreover, OAG downregulated NF-κB via the activation of PPARγ and reduced the expression of IL-1β, IL-6 and TNF-α in bone marrow-derived macrophages (BMDM) and mouse macrophage cell line RAW 264.7 [191]. These studies open up the possibilities of using OA as a potential therapeutic agent against IBD.
11.3. Osteoarthritis
Osteoarthritis is the most common chronic joint disease, which is prevalent in old age and affects the majority of those over 65 years of age [192,193]. It mainly affects joints of the knees, hips, and hands and results in mobility impairment [194,195]. Many studies throw light on the attenuating effects of OA in the development and progression of osteoarthritis. For instance, OA was found to maintain the homeostasis of ECM of chondrocytes via the stimulation of IL-1β and inhibition of NF-κB and Wnt/β-catenin signaling [196]. In another study, the chondroprotective activities of OA were investigated on IL-1β-induced chondrocytes inflammatory reactions. The results revealed that OA markedly suppressed the upregulation of cox-2 and NOS by IL-1β. Besides, OA attenuated IL-1β-stimulated upregulation of MMP-3 and MMP-13 expression, disintegrin, and matrix metalloproteinase with thrombospondin motifs, ADAMTS-4 and ADAMTS-5 expression. Furthermore, OA suppressed the activation of ERK 1/2 and PI3K/Akt signaling pathways and caused the reversal of IL-1β-induced type II collagen and aggrecan degradation [72]. Both studies suggest that OA could be a potential therapeutic agent for osteoarthritis.
11.4. Rheumatoid Arthritis (RA)
RA is a chronic inflammatory autoimmune disease that mostly affects the joints and has a detrimental effect on the health and quality of life of the patients [197]. Non-steroidal anti-inflammatory medicines, anti-rheumatism drugs, and glucocorticoid drugs are the most common therapeutics used for RA [198,199]. A plethora of studies has revealed that natural plant extracts and compounds considerably reduced the symptoms of RA in preclinical and clinical settings [200,201]. The effect of OA was investigated in collagen-induced arthritis (CIA) and human RA fibroblast-like synoviocytes (FLS). OA was shown to markedly reduce serum anti-collagen II antibodies, IL-1β, IL-6, IL-17, TNF-α, and the number of Th17 cells but increase the number of Tregs. Further, OA suppressed p38, MAPK, ERK1/2, and NF-κB signaling, and hence decreased inflammation to a large extent [202].
12. OA for Neurological Disorders
12.1. Attention-Deficit/Hyperactivity Disorder (ADHD)
Affecting about 5.29 percent of children and adolescents around the world, ADHD is a common neurodevelopmental disease in childhood resulting in impairments in personal, social, or vocational function, leading to isolation, worse grades, and a higher risk of depression and antisocial behavior. Inattention, impulsivity, and hyperactivity are the hallmarks of ADHD [203,204,205,206]. The psychostimulant drugs used for the treatment of ADHD, such as methylphenidate, has worse side effects, such as loss of appetite, insomnia, nausea, and dry mouth, and these medications are associated with the risk of substance use disorder [207,208]. Therefore, there has been an increased interest in alternative therapeutics, including plant-based compounds. Researchers have investigated the potential of OA for the treatment of ADHD. For instance, in a study, OA alleviated ADHD-like behavior in a spontaneously hypertensive rat (SHR) model. Given that, the GABAergic system has an inevitable role in ADHD, it was hypothesized that OA modulated GABA-A receptors, but the results showed that OA influenced other systems, such as DAergic, etc. Further, OA inhibited dopamine (DA) uptake just like methylphenidate, a dopamine transporter blocker drug. In conclusion, the above study proved that OA enhances ADHD-like behaviors by improving DA neurotransmission and not by the GABA pathway as reported earlier [209].
12.2. Alzheimer’s Disease
Alzheimer’s disease (AD) is a neurodegenerative disorder that progresses with age and is marked by cognitive impairment [210,211,212]. It is the most common type of dementia, and the symptoms usually start with moderate memory loss and progress to cognitive impairment, dysfunctions in day-to-day activities, and a variety of other issues [213,214,215,216]. Severe neuronal loss and lesions occur even before the clinical diagnosis of the disease. Therefore, the timely delivery of neuroprotective medications is crucial [217]. In a study, OA was found to prevent neuronal apoptosis, which is an important hallmark in neurodegenerative diseases. The main bioactive flavones in S. baicalensis, including OA, were evaluated for neuroprotective effects against amyeloid β-protein fragment (Aβ 25–35) (involved in the pathogenesis of AD) induced neuronal damage. All the compounds inhibited Aβ 25–35-induced ROS generation and resulted in cell cycle arrest. Further, the compounds reduced the expression of iNOS and cox-2, which resulted in the suppression of inflammatory cytokines, including TNF-α, NO, and PGE2. In addition, the compounds downregulated the NF-κB/MAPK pathway and relieved the Aβ 25–35-stimulated neuronal apoptosis [71].
12.3. Memory Impairment
Numerous studies have reported the neuroprotective effects of OA. For instance, in a study, OA was investigated against memory impairment induced by transient bilateral common carotid artery occlusion (2VO) in mice [218]. The number of brain-derived neurotrophic factor (BDNF) positive cells and cAMP response element-binding protein (CREBP) was shown to be significantly increased by OA. Besides, OA elevated Nissl bodies and OX-42 positive cells of the dentate gyrus and hippocampal CA1 areas. These results suggest that OA suppressed memory impairment and could be a plausible candidate for the treatment of memory loss [218]. Another study examined the effect of OA on drug-induced memory impairment using mice treated with scopolamine or diazepam. Intriguingly, OA restored cognitive impairments in mice and prevented GABA-induced Cl− influx in a single cortical neuron. These results opened novel avenues for using OA as a potential drug for the treatment of memory impairment [219].
13. OA for Obesity
Obesity is determined using the body mass index (BMI), which is calculated by weight in kilograms divided by height in square meters. Adults with a BMI of 25.0 to 29.9 kg/m2 are considered overweight, while those with a BMI of 30 kg/m2 or greater are considered obese [220]. The exact cause of obesity is numerous but microorganisms, epigenetics, higher fecundity, lack of sleep, endocrine disruptors, pharmacological iatrogenesis (illness caused by medical examination or treatment), and intrauterine and intergenerational impacts have all been related to obesity [221,222]. Obesity increases the chances of various protracted and fatal diseases, including cancer and many studies are ongoing to combine fundamental science with clinical research for better prevention and treatment strategies for this disease [223,224]. Accumulating data has revealed that OA has a beneficial effect on obesity (Table 1). For instance, a study investigated the anti-obesity effect of OA in mature adipocytes. OA was found to repress intracellular lipid accumulation. The adipogenic assay in 3T3-L1 pre-adipocytes and pancreatic lipase assay showed that OA prevented lipid accumulation in 3T3-L1 pre-adipocytes. Further, OA also inhibited PPARγ and C/EBP α, the major adipogenic transcription factor [225]. However, further investigations are required to establish the definite role of OA in the treatment and prevention of obesity.
14. OA for Other Diseases
Coxsackievirus B (CVB) is a human pathogen that causes diseases such as myocarditis, pericarditis, meningitis, and pancreatitis [226,227]. In a study, the effects of OA on pancreatitis were investigated in the CVB3-infected mice model and it was found that OA attenuated the changes in body weight and blood glucose levels induced by CVB3 infection and lowered the pancreatic lesions and inflammatory factors IL-6 and TNF-α but increased phospho-eIF2 α levels [228]. Therefore, OA could be used as a potential pharmacological agent against CVB3-induced pancreatic injury.
OA is also effective against osteoporosis, which has been proved by various studies. For instance, Xian and his colleagues (2021) established that OA reduced the formation and function of osteoclasts, the multi-nucleated cells responsible for bone resorption by lowering intracellular ROS levels and suppressing the activity of nuclear factor of activated T cells 1 (NFATc1), the master transcriptional regulator of osteoclastogenesis. This study explained the anti-osteoclast effect of OA, and it can be a promising agent that aids in the treatment of osteolytic diseases [229]. Figure 2 briefly summarizes the mechanisms of action of OA against various chronic diseases.
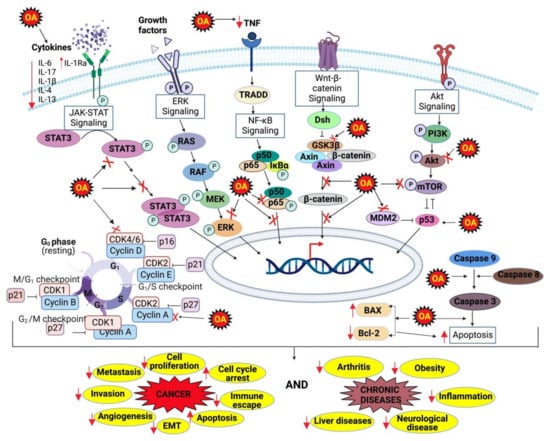
Figure 2.
Mechanisms of action of OA against various chronic diseases.
15. Pharmacokinetic Studies of OA
To date, a considerable amount of studies have been published on the pharmacokinetics and pharmacodynamics of OA. For instance, a study examined the pharmacokinetics and excretion and tissue distribution of OA in rats using the sensitive and rapid UPLC-MS/MS methodologies for the quantification of OA and its two metabolites OAG and OS. All three compounds were distributed throughout the rat tissues, with OA being more widely distributed in the liver and its metabolites being distributed more in the kidneys [230] Additionally, ultra-high-performance liquid chromatography-tandem mass spectrometry methods were used to identify OA, OAG, and OS in beagle dog plasma [231]. Another study developed and validated solid-phase extraction-liquid chromatography-tandem mass spectrometry (SPE-LC/MS/MS) for the simultaneous detection of OA and OAG from S. baicalensis following oral treatment in rats. The occurrence of these compounds in rat brains and plasma was demonstrated in this investigation, imply that they have the ability to cross the blood–brain barrier [232]. In another study, the in vitro cell pharmacokinetic profiles of OA and OAG were reported in tumor cell lines via a highly selective and sensitive solid-phase extraction (SPE)-UPLC-MS/MS method. This study showed that both OA and OAG were found to be largely distributed in the nuclei of HepG2 tumor cells [233].
16. Safety and Toxicities of OA
A plethora of studies has shown the anti-cancer and anti-inflammatory activities of OA in both in vitro and in vivo settings. A study conducted by Mu et al. showed that OA of up to 100 μM did not significantly kill the normal human cells, i.e., HUVECs and L-02 cells, whereas the same dose killed the cancer cells -HepG2, K-562, and MDAMB-435 significantly [234]. Moreover, dose increments up to 400 μM showed a detrimental effect on these cancer cells, whereas only around 40% cell death was shown in normal cells [234]. Another study showed that OA of 50 mg/kg administered orally along with 200 mg/kg of imatinib significantly inhibited tumor growth in K-562 xenograft models without affecting the body weight and vital organs, such as heart, kidney, liver, and spleen [235]. More recently, Wei et al., (2019) showed that OA of 300 mg/kg when treated alone or in combination with TMZ (50 mg/kg) effectively inhibited the glioma growth in BALB/c nude mice. The toxicity studies showed that OA did not induce changes in body weight and toxicity to peripheral blood cells and vital organs [236]. This study also showed that OA abrogated TMZ induced reduction in body weight, leukocyte count, and lung injury, indicating the safety of OA in normal cells/organs [236]. However, dose standardization for different diseases, and safe and toxic doses of OA for humans are currently unknown due to the lack of clinical studies.
17. Discussion and Conclusions
It is now well established that flavonoids are polyphenolic secondary metabolites found in plants and a variety of foods. Apart from their biological roles, flavonoids have a broad spectrum of pharmacological activities, including anti-atherosclerotic, anti-inflammatory, anti-cancer, anti-thrombotic, anti-viral, and anti-osteoporotic actions [237,238,239,240]. We have an immense amount of preclinical and clinical evidence in support of natural compounds as therapeutic drugs for a wide variety of chronic diseases. In the current review, we attempted to accentuate the pharmacological properties of the flavonoid, OA, against various chronic diseases, such as cancer, CVDs, liver diseases, eye diseases, neurological, and inflammatory diseases. OA is manifested to modulate different targets and pathways that lead to the development of these chronic diseases. Considering its multi-targeting properties and high capability in regulating various signaling pathways, OA is a credible candidate for forthcoming drug development with minimal side effects. Overwhelming pieces of evidence have spelled out the significant anti-oxidant, anti-analgesic, and anti-neoplastic effects of OA. It is widely known that inflammation and oxidative stress are the two most critical factors in the development of a wide range of chronic diseases. Numerous studies have recognized OA as a multifaceted drug due to its prominent anti-inflammatory, anti-oxidant, and anti-tumorigenic effects. This compound has shown a significant reduction in oxidative stress and enhanced antioxidant enzyme activity in various models. These remarkable anti-oxidant and anti-inflammatory qualities make OA one of the greatest therapeutic possibilities in the future.
According to studies, OA reduced the expression of inflammatory markers, such as TNF-α and interleukins (IL-1, IL-4, IL-6, IL-13, etc.), which are produced in response to deleterious external stimuli and have been associated with a variety of human diseases, such as arthritis, cancer, and liver injury. It is well known that cellular oxidative stress brings about inflammation and results in the development of various chronic diseases. This inflammation is caused by the generation of free radicals as a result of infection or injury [241]. Multiple lines of evidence strongly suggested that OA protects cells from oxidative damage by elevating the levels of anti-oxidant enzymes, such as SOD. Moreover, a large body of literature has extensively documented the abilities of OA to modulate various signaling pathways, such as NF-κB, STAT-3, ERK/MAPK, Hh, PI3K/Akt, etc., and many genes and proteins, such as cox-2, MMP-9, NAG-1, HIF-1α, VEGF, cyclin B1, survivin, p21, p27, p53, PARP, caspases, etc., [54,120,242,243,244,245,246,247,248,249,250].
Accumulating number of studies have explained the chemosensitizing and radiosensitizing properties of OA against various cancers. For instance, the synergistic effects of OA with cisplatin were evaluated in a study where OA significantly reduced NSCLC cell resistance to cisplatin by binding to HIF-1α, and thereby inhibiting xeroderma pigmentosum group C transcription (XPC) [135]. Various studies also reported that OA can potentially reverse imatinib drug resistance in CML cells [251]. Similarly, OA in combination with 5-FU, reversed the multidrug resistance by decreasing the expression of the multidrug resistance gene (MDR1) [252]. OA also possesses radiosensitizing properties, as evident in the studies conducted by Tan and his colleagues (2017) where OA exhibited radiosensitization of ESCC cells by arresting the tumor cells in the G2/M phase and inducing apoptosis [136].
Most crude medications or compound formulations in Ayurveda and TCM are made as decoctions and administered orally. Pharmacokinetic studies in rats after intragastrical administration of OA or its source plants revealed that OA may be absorbed in its native form from the gastrointestinal tract and that the concentration of OA in the plasma increased over time. When OA, OAG, and OS were administered, OA was more broadly distributed in tissue than its metabolites after oral administration, and the tissue concentration level of OA was the highest [230,253,254].
Even though OA has shown therapeutic potential in many in vitro investigations, it has a very low oral bioavailability due to its significant first-pass metabolism and primary glucuronidation in the guts [255]. Furthermore, the metabolites of OA, such as OAG and OS, also proved effective against a few chronic diseases. In a study, several derivatives of OA were synthesized and screened for antitumor activities in HepG2 cell lines. Intriguingly, some derivatives showed higher tumor inhibitory and apoptotic properties than OA. This study opened up new possibilities for synthesizing more efficient derivatives of OA as promising anti-cancer agents [256].
Comprehensively, OA embodies various biological roles and stands to be one of the most efficacious compounds in the prophylaxis and therapy of different chronic diseases with boundless potential in drug discovery. However, more clinical research is needed to back up the aforementioned findings. Furthermore, as stated earlier, more potent analogs and formulations of OA could aid in the advancement of safer and more effective drugs for a variety of chronic conditions. However, detailed clinical evaluation and trials are mandatory to examine the efficiency and toxicity of the compound and its formulations, thereby making OA an invaluable therapeutic agent.

Table 1.
Preventive and therapeutic properties of OA against various chronic diseases.
Table 1.
Preventive and therapeutic properties of OA against various chronic diseases.
| Disease | In Vitro/ In Vivo | Dose/Conc. | Model | Mechanism of Action or Outcome | References |
|---|---|---|---|---|---|
| Cancer | |||||
| Breast cancer | In vitro | 50, 100, 200 μM | MDA-MB-231, MCF-7 | ↑SIRT3, SOD2, PHD activity, ↓glycolysis, HIF-1α, mitochondrial ROS | [76] |
| In vivo | 100 mg/kg | MDA-MB-231 xenograft | ↑SIRT3, SOD2, ↓tumor volume and mass, glycolysis, HIF-1α, hexokinase II, | [76] | |
| In vitro | 10, 20, 40 μM | MDA-MB-231 | ↑E-cadherin, p27, ↓cell proliferation, CDK2, cyclin E, vimentin, N-cadherin, EMT, migration, invasion, COX-2, NF-κB, IL-6, IL-8, TNF-α | [114] | |
| Cervical cancer | In vitro | 5, 20, 80 μM | HeLa | ↑Procaspase-3, procaspase-8, procaspase-9, cleaved PARP, apoptosis ↓Bcl-2, cell growth | [75] |
| In vivo | 40, 80 mg/kg | HeLa xenograft | ↑Cleaved PARP, ↓tumor growth, Bcl-2, procaspase-3, procaspase-8, procaspase-9 | [75] | |
| Colon cancer | In vitro | 200 μM/L | HT-29 cells | ↑Bax, p53, PARP, procaspase-3, ROS, ↓COX-2, Bcl-2, PGE2 | [118] |
| In vivo | 100 mg/kg | HT-29 | ↓Tumor, COX-2 | [118] | |
| In vitro | 100 μM/L | HCT-116 | ↑Caspase-3, caspase-9, Bax, ROS, Nrf2, HO-1, NQO1, ↓Bcl-2, | [56] | |
| In vivo | 50, 100, 200 mg/kg | HCT-116 xenograft | ↑Nrf-2, apoptosis, ↓tumor growth | [56] | |
| ESCC | In vitro | 10, 50 μM | TE13, ECA109 | ↑Apoptosis, G2/M arrest, radiosensitization, ↓cyclin B1, cdc2 | [136] |
| Glioma | In vitro | 25, 50, 75, 100, 125, 150, 175, 200 μM | U251, U118, U87 | ↑Autophagy, Beclin, ↓Akt, ERK Notch-1, Mcl-1 | [126] |
| In vitro | 50 μM | C6, U251 | ↑Apoptosis, ↓p-Akt, β-catenin, IP3R1, p-GSK-3β | [94] | |
| In vivo | 150 mg/kg | C6 xenograft | ↑Apoptosis, ↓Akt/β-catenin, IP3R1, p-GSK-3β, | [94] | |
| Hematological malignancies | In vitro | 60 μM | K562, KU812 with M2-10B4 | ↑Apoptosis, ↓CXCL12/CXCR7, p-ERK, p-BAD, survivin | [130] |
| In vivo | 200 mg/kg | K562 xenograft | ↑Apoptosis, ↓CXCR7, p-ERK, CD13+ cells | [130] | |
| In vitro | 20 μM | HL-60, NB4 | ↑TNF-α sensitivity, ↓tRXRα, PI3K/Akt, NF-κB | [83] | |
| In vivo | 80 mg/kg | AML cell xenograft | ↑Survival, ↓NF-κB, AML cell population | [83] | |
| In vitro | 10–160 μM | t (8i21)-positive kasumi-l, primary AML cells | ↑C/EBPα, p21, CD11b/CD14, ↓AML 1/ETO, HDAC-1 | [129] | |
| In vivo | 200 mg/kg | NOD/SCID mice | ↑Survival, ↓HDAC-1, AML1/ETO, CD45+ cells, | [129] | |
| Hepatocellular carcinoma | In vitro | 50 μM | HepG2 cells | ↑Apoptosis, p53, cleaved PARP, ↓Cell viability, TS and DPD mRNA, COX-2, Bcl-2, procaspase-3 | [119] |
| In vivo | 1000 mg/kg/day | H22 xenograft | ↓Tumor growth, tumor weight | [119] | |
| In vitro | 12.5, 25, 50 μM | SMMC-7721 | ↑NAG1, acetylation of C/EBPβ, ↓migration, invasion, EMT, p-SMAD2/3, TGF-β1/SMAD axis | [120] | |
| In vivo | 200 mg/kg | SMMC-7721 | ↑E-cadherin, ↓pulmonary metastasis, vimentin, twist1 | [120] | |
| Kaposi’s sarcoma | In vitro | 20–1000 μM | KSHVvIL-6 | ↑Apoptosis, PPARγ, invasion, neovascularization, ↓Prox1, VEGFR3, LYVE-1, podoplanin | [80] |
| Lung cancer | In vitro | 40 μM | H460 | ↓Tregs, TGF-β, NF-κB | [134] |
| In vivo | 60 mg/kg | H460 xenograft | ↓Tumor, Tregs, FOXP3, | [134] | |
| In vitro | 50 μM/L | H460, A549, 95D, PC9, HCC827, H1975 | ↑Apoptosis, ↓tumor, XPC transcription | [135] | |
| In vivo | 50 mg/kg | H460 xenografts | ↑Cisplatin sensitivity, ↓tumor growth, Ki67, PCNA, XPC expression | [135] | |
| Skin cancer | In vitro | 20 μM | JB6P+ | ↓Transformation, inflammation, SHCBP1, NF-κB p65, IL-1β, IL-6, IL-18, TNF-α, COX-2, iNOS, | [137] |
| In vivo | 40 mg/kg | DMBA/TPA mice | ↓SHCBP1, IL-1β, IL-4, IL-6, IL-18, TNF-α, NLRP3, PCNA tumorigenesis, incidence rate, tumor multiplicity, epidermal thickness | [137] | |
| Cardiovascular diseases | |||||
| Cardioprotective effects | In vivo | 40 mg/kg | C57BL/6 mice | ↑Body weight, SIRT1, cAMP/protein kinase A, improved contractile function, Nrf2, HO-1, NQO1, Bcl-2, ↓plasma and cardiac CK-MB, LDH, LVEDP, 4-HNE, nitrotyrosine, gp91phox, NADPH oxidase 4, p47phox, p67phox, IL-6, IL-1β, MMP-2, MMP-9, p-IκBα, caspase 3/7 activity, PARP activity, apoptosis | [143] |
| Endotoxemia | In vivo | 10, 20 μM | Sprague-Dawley rats | ↑Coronary flow, LVDP ↓CPP | [55] |
| Hind limb ischemia | In vivo | 10 mg/kg/day | C57BL/6 mice | ↑VEGFA, Ang-2, FGF-2, PDGF-BB, angiogenesis, perfusion recovery, regeneration of myocytes ↓IL-1β, tissue injury, ischemia, apoptosis of myocytes | [153] |
| Chronic liver diseases | |||||
| Acute liver injury | In vivo | 60 mg/kg | C57 BL/6 mice | ↑IL-1Ra, HGF, EGF, PCNA positive cells, survival, ↓IL-1β, IL-6, TNF-α, necrotic areas | [69] |
| In vivo | 15, 30, 60 mg/kg | BALB/c mice | ↑Nrf 2, HO-1, ↓AST, ALT, TNF-α, MDA, MPO activity, NF-κB, TLR4, necrosis | [159] | |
| ALD | In vitro | 10–100 μM | LO2 cells | ↑YAP, ↓AST, ALT, LDH, p21, p16 and HMGA1 | [169] |
| In vivo | 30 mg/kg | ICR mice | ↑YAP, ↓AST, ALT, ALP, lipid vacuolation | [169] | |
| In vitro | 10, 20, 40 μM | LO2 cells | ↑Mfn2, PGC-1α, ↓LDH, IL-1β, IL-18, caspase-1, NF-κB, ROS, NLRP3 inflammasome | [77] | |
| In vivo | 40 mg/kg | ICR mice | ↓Inflammation, lipid accumulation, ALT, ALP, AST | [77] | |
| Hepatic steatosis | In vitro | 10, 20, 40 μM | LO2 cells | ↑CPT1, PPARα, PPARγ, ↓lipid droplet accumulation, HIF-1α, apoptosis, SREBP1, FAS, SCD1 | [59] |
| In vivo | 30 mg/kg | ICR mice | ↓ Apoptosis, ALT, AST, ALP, IL-6, IL-8, TNF-α | [59] | |
| Hepatic fibrosis | In vitro | 20, 30, 40 μM | HSCs | ↑LC3-B, Atg3, Atg4, Atg5, Atg7, Atg9, beclin, Atg12, Atg14, ↓α-SMA, desmin, α1collagen, fibronectin, TGF-β, TNF-α, p62 | [162] |
| In vivo | 20, 30, 40 mg/kg | ICR mice | ↑LC3-B, Atg5, beclin1, ↓AST, ALT, ALP, α-SMA, α1collagen, fibronectin, PDGF-βR, TGF-βR1, p62, fibrosis lesions, necrosis, inflammation | [162] | |
| In vitro | 20, 30, 40 μM | HSCs | ↓Hexokinase II, LDH-A, PFK1, PKM2, actin stress fibers, p-MLC2, contraction | [163] | |
| In vivo | 40 mg/kg | ICR mice | ↓Liver injury, glycolysis, α-SMA, α1collagen, fibronectin, ALT, AST, TBIL, IBIL, p-MLC2 | [163] | |
| In vitro | 20, 30, 40 μM | HSC | ↑SLC7A11, GSH, lipid droplet content, retinol, cholesterol, triglyceride ↓ATGL, α-SMA, α1collagen, fibronectin, desmin, ROS | [79] | |
| In vivo | 20 mg/kg | C57BL/6 | ↑Antioxidant activity, ↓liver fibrosis, collagen deposition lipid droplet content, retinol, cholesterol, triglyceride, α-SMA, collagen I | [79] | |
| In vitro | 20, 30, 40 μM | HSC | ↑Autophagy, Atg5, Atg12, beclin, LC3B, ↓ NF-κB, NLRP3, TNF-α, IL-1β, p-PI3K, p-Akt p-mTOR, ROS, p62, IL-1β, IL-4, IL-6, IL18, TNF-α, IFNγ | [92] | |
| In vivo | 20, 30, 40 mg/kg | C57BL/6 | ↓ NF-κB, α-SMA, IL-1β, IL-4, IL-6, IL18, TNF-α, IFNγ | [92] | |
| In vitro | 20, 30, 40 μM | LSECs | ↓VEGF-A, angiogenesis | [256] | |
| In vivo | 40 mg/kg | ICR mice | ↓VEGF-A, Ang-2, CD31, HIF-1α | [256] | |
| In vitro | 20, 30, 40 μM | HSC | ↑Cleaved caspase-9, cleaved caspase-3, cleaved PARP, p51, p21, p27, S-phase arrest, Bax, collagen degradation, MMP-9, ATF4, p-PERK, cleaved ATF6, ↓Fibrogenesis, PDGF-β, TGF-β, EGFR, cyclin A, cyclin E, CDK-2, Bcl-2, collagen synthesis, TIMP-2, α-SMA, collagen I | [165] | |
| In vivo | 20, 30, 40 mg/kg | ICR mice | ↑ERS pathway, IL-6, IL18, TNF-α, AST, ALT | [165] | |
| Inflammatory diseases | |||||
| Allergic asthma | In vitro | 0.1, 0.3, 1, 3, 10, 30 μM | RBL-2H3 mast cells | ↓β-Hexosaminidase release, antigen-induced degranulation | [70] |
| In vivo | 5 mg/kg | Female BALB/c mice | ↓IFNγ, IL-2, IL-4, IL-5, IL-13, eosinophils, inflammation score, mucin | [70] | |
| In vivo | 15, 30, 60 mg/kg | BALB/c mice | ↓ IgE, p-IκB, p-NF-κB, IL-4, IL-5, IL-13, airway hyporesponsiveness, inflammatory cells infiltration, thickening of alveolar wall | [183] | |
| Inflammatory bowel disease | In vivo | 50 mg/kg | BALB/c mice | ↓Inflammation, IL-1β, IL-6, IL-17, TNF- α, Muc2, IFNγ | [189] |
| In vivo | 100, 200 Mg/kg | BALB/c mice | ↓Distribution of CD11b+ inflammatory cells and F4/80+ macrophages, MPO, iNOS, NLRP3, IL-1β, IL-6, TNF- α, p65 | [190] | |
| Lung inflammation | In vitro | 50, 100, 150 μM | BEAS-2B and RAW 2647 cells | ↑Nrf2, GSH, HO-1, ↓TNF-α, IL-1β | [143] |
| In vivo | 15, 30, 60 mg/kg | C57BL/6 | ↑GR activity, GSH, ↓interstitial edema, infiltrated immune cells, alveolar wall thickness, TNF α, IL-1β, MCP-1, 3-nitrotyrosine, 8-OHdG, 8-isoprostane | [143] | |
| Rheumatoid arthritis | In vitro | 1, 4, 16 μM | FLS cells | ↑IL-10, ↓IL-1β, IL-6, p-ERK, p-MAPK, p65 | [202] |
| In vivo | 10 mg/kg | DBA/1 mice with CIA | ↑Tregs ↓total IgG, IgG1, IgG2a, IgG2b, IL-1β, IL-6, IL-17, TNF-α, arthritis score, swelling, joint inflammation, Th17 cells | [202] | |
| Osteoarthritis | In vitro | 2.5, 5, 10, 20, 50 μM | Chondrocytes | ↓IL-1β, MMP-13, ADAMTS-5, NO, PGE2, ALP IL-6, TNF-α, NF-κB, RUNX-2, collagen X, β-catenin | [196] |
| In vivo | 10 mg/kg | OA-induced mice model | ↓OARSI score | [196] | |
| In vitro | 2–128 μM | Chondrocytes | ↓NOS, cox-2, MMP-3, MMP-13, ERK1/2, PI3K/Akt | [72] | |
| Obesity | In vitro | 25, 50, 75 μM | 3T3-L1 pre-adipocytes | ↓Lipid accumulation, adipogenesis, PPARγ, C/EBPα | [225] |
| Neurological diseases | |||||
| ADHD | In vivo | 5, 10 mg/kg | SHR, WKY | ↓Drinking attempts, drinking frequency, dopamine reuptake | [209] |
| Alzheimer’s disease | In vitro | 10, 50, 100 μM | PC12 cells | ↓Ca2+, Bax, iNOS, cleaved caspase-8, cleaved PARP-1, TNF-α, NO, PGE2, p-IκBα, cox-2, p-NF-κB, p-p38, p-JNK, ROS, apoptosis, cell cycle arrest, | [71] |
| Memory impairment | In vivo | 5 mg/kg | ICR mice | ↑ChAT, ↓Nissl bodies, OX-42 positive cells, GFAP positive cells, iNOS, spontaneous alteration behavior, micro glial cell activation, lipid peroxidation | [218] |
↑—Increase/Upregulation; ↓—Decrease/Downregulation.
Author Contributions
A.B.K., J.-T.C. and G.S. contributed to this study design, conceptualization, supervision, and review editing. A.S. performed bibliographic search, contributed to table preparation, and original manuscript writing. M.H. performed bibliographic search and contributed to table preparation, artwork, review and editing. T.N.D. contributed to table preparation. M.S.A., M.A., S.K.S. and S.G. contributed to review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by BT/556/NE/U-Excel/2016 grant awarded to Ajaikumar B. Kunnumakkara by Department of Biotechnology (DBT), Government of India. The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University (KKU) for funding this work through the Research Group Program under research project number (RGP.2/248/43).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Mangala Hegde acknowledges Science and Engineering Research Board (SERB), Govt. of India for providing the prestigious National Post-Doctoral Fellowship (PDF/2021/004053).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
4-HNE—4-Hydroxynonenal; 8-OHdG—8-Hydroxy-2′-deoxyguanosine; α-SMA-alpha smooth muscle actin; ADAMTS5—A disintegrin and metalloproteinase with thrombospondin motifs 5; AML—Acute myeloid leukemia; ALD—Alcohol liver disease; ALL—acute lymphocytic leukemia; ALP—Alkaline phosphatase; ALT—Alanine aminotransferase; cAMP—Cyclic adenosine monophosphate; Ang-2—Angiopoetin-2; AST—Aspartate amino transferase; ATF4—Activating transcription factor 4; Atg—Autophagy related; ATGL—adipose triglyceride lipase; BAD—Bcl-2 associated agonist of cell death; BAX—Bcl-2 associated X protein; Bcl-2—B-cell lymphoma 2; C/EBPα—CCAAT binding protein alpha; C/EBPβ—CCAAT binding protein beta; cdc2—Cell division cycle 2; CD—Cluster of differentiation; CDK—Cyclin dependent kinase; ChAT—Choline acetyl transferase; CK-MB—Creatinine kinase MB; CPP—Coronary perfusion pressure; DPD—Dihydropyrimidine dehydrogenase; COX-2—Cyclooxygenase-2; CXCL2—Chemokine C-X-C motif ligand 2; CXCR4—Chemokine receptor type 4; EGF—Epidermal growth factor; EGFR—Epidermal growth factor receptor; EMT—Epithelial-mesenchymal transition; ERK—Extracellular signal-regulated kinase; ERS—Endoplasmic reticulum stress; FAS—Fatty acid synthase; FGF-2—Fibroblast growth factor 2, FLS—Fibroblast-like synoviocytes; FOXP3—Forkhead box P3; GFAP—Glial fibrillary acidic protein; GR—Glutathione reductase; GSK-3β—Glycogen synthase kinase 3 beta; GSH—Glutathione; HDAC—Histone deacetylase; HGF—Hepatocyte growth factor; HIF-1α—Hypoxia inducible factor 1 alpha; HKII—Hexokinase II; HNF—Hepatocyte nuclear factor; HMGA1—High mobility group AT-hook 1; HO-1—Hemeoxygenase-1; IBL—Indirect bilirubin; IκBα—IkappaB kinase alpha; IFNγ—Interferon gamma; IL—Interleukin; iNOS—Inducible nitric oxide synthase; IP3R1—Inositol 1,4,5-triphospshate receptor, type 1; JNK—Jun N-terminal kinase; LC3-B—Microtubule-associated proteins 1A/1B light chain 3B; LDH—Lactate dehydrogenase; LSEC—Liver sinusoidal endothelial cells; LVDP—Left ventricular developed pressure; LYVE-1—Lymphatic vessel endothelial hyaluronan receptor 1; Mcl-1—Myeloid cell leukemia 1; MCP-1—Monocyte chemoattractant protein 1; MDA—Malondialdehyde; MDM—Mouse double minute; MDR—Multi drug resistant gene; Mfn2—Mitofusin 2; MLC2—Myocin light chain 2; MMP—Matrix metalloproteinase; MMPo—Mitochondrial membrane potential; MPO—Myeloperoxidase; mTOR—Mammalian target of rapamycin; NADPH—Nicotinamide adenine dinucleotide phosphate; NAG1—Non-steroidal anti-inflammatory drug activated gene-1; NF-κB—nuclear factor-kappa B; NQO1—NAD(P)H quinone dehydrogenase 1; NOS—nitric oxide synthase; Nrf2—Nuclear factor erythroid-2-related factor-2; OARSI—Osteoarthritis research society international; OSSC—Oral squamous cell carcinoma; PARP—Poly (ADP-ribose) polymerase; PCNA—Proliferating cell nuclear antigen; PDGF-BB—platelet-derived growth factor; PDGF-βR—platelet-derived growth factor beta receptor; PERK—Protein kinase RNA-like endoplasmic reticulum kinase; PFK—Phosphofructokinase; PGE2—Prostaglandin E2; PHD—Prolyl hydroxylase; PI3K—Phosphoinositide 3 kinase; PKM1/M2—Pyruvate kinase isozymes M1/M2; PPARα—Peroxisome proliferator-activated receptor alpha; PPARγ—Peroxisome proliferator-activated receptor; Prox1—Prospero homeobox 1; ROS—Reactive oxygen species; RUNX2—Runt related transcription factor 2; SCD1—Stearoyl-CoA desaturase 1; SIRT3—Sirtuin 3; SHCBP1—SHC binding and spindle associated 1; SLC7A11—Solute carrier family 7 member 11; SOD2—Superoxide dismutase 2; SREBP1—Sterol regulatory element binding protein 1; STAT3—Signal transducer and activator of transcription; TBIL—Total bilirubin; TGF-β—Transforming growth factor-beta; TGF-βR1—Transforming growth factor-beta receptor 1; TIMP-2—Tissue inhibitor of metalloproteinases 2; TLR4—Toll like receptor 4; TNF-α—tumor necrosis factor alpha; Tregs—regulatory T-cell; tRXRα—Truncated retinoid X receptor alpha; TS—Thymidine synthetase; VEGFA—Vascular endothelial growth factor A; VEGFR3—Vascular endothelial growth factor receptor 3; XPC—Xenoderma pigmentosum group C protein; YAP—Yes1 associated transcriptional regulator.
References
- Kumar, A.; Harsha, C.; Parama, D.; Girisa, S.; Daimary, U.D.; Mao, X.; Kunnumakkara, A.B. Current clinical developments in curcumin-based therapeutics for cancer and chronic diseases. Phytother. Res. 2021, 35, 6768–6801. [Google Scholar] [CrossRef] [PubMed]
- Parama, D.; Girisa, S.; Khatoon, E.; Kumar, A.; Alqahtani, M.S.; Abbas, M.; Sethi, G.; Kunnumakkara, A.B. An overview of the pharmacological activities of scopoletin against different chronic diseases. Pharmacol. Res. 2022, 179, 106202. [Google Scholar] [CrossRef] [PubMed]
- Heymach, J.V.; Shackleford, T.J.; Tran, H.T.; Yoo, S.Y.; Do, K.A.; Wergin, M.; Saintigny, P.; Vollmer, R.T.; Polascik, T.J.; Snyder, D.C.; et al. Effect of low-fat diets on plasma levels of NF-kappaB-regulated inflammatory cytokines and angiogenic factors in men with prostate cancer. Cancer Prev. Res. 2011, 4, 1590–1598. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Sailo, B.L.; Roy, N.K.; Thakur, K.K.; Banik, K.; Shakibaei, M.; Gupta, S.C.; Aggarwal, B.B. Cancer drug development: The missing links. Exp. Biol. Med. 2019, 244, 663–689. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Yang, S.F.; Sethi, G.; Hu, D.N. Natural bioactives in cancer treatment and prevention. Biomed. Res. Int. 2015, 2015, 182835. [Google Scholar] [CrossRef]
- Kirtonia, A.; Gala, K.; Fernandes, S.G.; Pandya, G.; Pandey, A.K.; Sethi, G.; Khattar, E.; Garg, M. Repurposing of drugs: An attractive pharmacological strategy for cancer therapeutics. Semin. Cancer Biol. 2021, 68, 258–278. [Google Scholar] [CrossRef]
- Deldar Abad Paskeh, M.; Asadi, S.; Zabolian, A.; Saleki, H.; Khoshbakht, M.A.; Sabet, S.; Naghdi, M.J.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; et al. Targeting cancer stem cells by dietary agents: An important therapeutic strategy against human malignancies. Int. J. Mol. Sci. 2021, 22, 11669. [Google Scholar] [CrossRef]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef]
- Manu, K.A.; Shanmugam, M.K.; Li, F.; Chen, L.; Siveen, K.S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J. Mol. Med. 2014, 92, 267–276. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Kim, S.H.; Sethi, G.; Ahn, K.S. Farnesol inhibits tumor growth and enhances the anticancer effects of bortezomib in multiple myeloma xenograft mouse model through the modulation of STAT3 signaling pathway. Cancer Lett. 2015, 360, 280–293. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Banik, K.; Girisa, S.; Shabnam, B.; Shakibaei, M.; Fan, L.; Arfuso, F.; Monisha, J.; Wang, H.; Mao, X.; et al. The vital role of ATP citrate lyase in chronic diseases. J. Mol. Med. 2020, 98, 71–95. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Parama, D.; Daimari, E.; Girisa, S.; Banik, K.; Harsha, C.; Dutta, U.; Kunnumakkara, A.B. Rationalizing the therapeutic potential of apigenin against cancer. Life Sci. 2021, 267, 118814. [Google Scholar] [CrossRef]
- Bordoloi, D.; Banik, K.; Shabnam, B.; Padmavathi, G.; Monisha, J.; Arfuso, F.; Dharmarajan, A.; Mao, X.; Lim, L.H.K.; Wang, L.; et al. TIPE family of proteins and its implications in different chronic diseases. Int. J. Mol. Sci. 2018, 19, 2974. [Google Scholar] [CrossRef]
- Padmavathi, G.; Roy, N.K.; Bordoloi, D.; Arfuso, F.; Mishra, S.; Sethi, G.; Bishayee, A.; Kunnumakkara, A.B. Butein in health and disease: A comprehensive review. Phytomedicine 2017, 25, 118–127. [Google Scholar] [CrossRef]
- Shabnam, B.; Padmavathi, G.; Banik, K.; Girisa, S.; Monisha, J.; Sethi, G.; Fan, L.; Wang, L.; Mao, X.; Kunnumakkara, A.B. Sorcin a potential molecular target for cancer therapy. Transl. Oncol. 2018, 11, 1379–1389. [Google Scholar] [CrossRef]
- Buhrmann, C.; Kunnumakkara, A.B.; Kumar, A.; Samec, M.; Kubatka, P.; Aggarwal, B.B.; Shakibaei, M. Multitargeting effects of Calebin A on malignancy of CRC cells in multicellular tumor microenvironment. Front. Oncol. 2021, 11, 650603. [Google Scholar] [CrossRef]
- Babu, B.H.; Jayram, H.N.; Nair, M.G.; Ajaikumar, K.B.; Padikkala, J. Free radical scavenging, antitumor and anticarcinogenic activity of gossypin. J. Exp. Clin. Cancer Res. 2003, 22, 581–589. [Google Scholar]
- Mastron, J.K.; Siveen, K.S.; Sethi, G.; Bishayee, A. Silymarin and hepatocellular carcinoma: A systematic, comprehensive, and critical review. Anticancer Drugs 2015, 26, 475–486. [Google Scholar] [CrossRef]
- Patel, S.M.; Nagulapalli Venkata, K.C.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Zhang, J.; Ahn, K.S.; Kim, C.; Shanmugam, M.K.; Siveen, K.S.; Arfuso, F.; Samym, R.P.; Deivasigamanim, A.; Lim, L.H.; Wang, L.; et al. Nimbolide-induced oxidative stress abrogates STAT3 signaling cascade and inhibits tumor growth in transgenic adenocarcinoma of mouse prostate model. Antioxid. Redox Signal 2016, 24, 575–589. [Google Scholar] [CrossRef]
- Lee, J.H.; Chiang, S.Y.; Nam, D.; Chung, W.S.; Lee, J.; Na, Y.S.; Sethi, G.; Ahn, K.S. Capillarisin inhibits constitutive and inducible STAT3 activation through induction of SHP-1 and SHP-2 tyrosine phosphatases. Cancer Lett. 2014, 345, 140–148. [Google Scholar] [CrossRef]
- Roy, N.K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A.K.; Thakur, K.K.; Padmavathi, G.; Shakibaei, M.; Fan, L.; Sethi, G.; et al. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. Int. J. Mol. Sci. 2019, 20, 74101. [Google Scholar] [CrossRef]
- Shishodia, S.; Sethi, G.; Ahn, K.S.; Aggarwal, B.B. Guggulsterone inhibits tumor cell proliferation, induces S-phase arrest, and promotes apoptosis through activation of c-Jun N-terminal kinase, suppression of Akt pathway, and downregulation of antiapoptotic gene products. Biochem. Pharmacol. 2007, 74, 118–130. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Banik, K.; Bordoloi, D.; Harsha, C.; Sailo, B.L.; Padmavathi, G.; Roy, N.K.; Gupta, S.C.; Aggarwal, B.B. Googling the guggul (commiphora and boswellia) for prevention of chronic diseases. Front. Pharm. 2018, 9, 686. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Bordoloi, D.; Thakur, K.K.; Monisha, J.; Arfuso, F.; Sethi, G.; Mishra, S.; Kumar, A.P.; Kunnumakkara, A.B. Possible use of Punica granatum (Pomegranate) in cancer therapy. Pharmacol. Res. 2018, 133, 53–64. [Google Scholar] [CrossRef]
- Moballegh Nasery, M.; Abadi, B.; Poormoghadam, D.; Zarrabi, A.; Keyhanvar, P.; Khanbabaei, H.; Ashrafizadeh, M.; Mohammadinejad, R.; Tavakol, S.; Sethi, G. Curcumin Delivery Mediated by Bio-Based Nanoparticles: A Review. Molecules 2020, 25, 689. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Sung, B.; Ravindran, J.; Diagaradjane, P.; Deorukhkar, A.; Dey, S.; Koca, C.; Tong, Z.; Gelovani, J.G.; Guha, S.; et al. Zyflamend suppresses growth and sensitizes human pancreatic tumors to gemcitabine in an orthotopic mouse model through modulation of multiple targets. Int. J. Cancer 2012, 131, E292–E303. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Nair, A.S.; Ahn, K.S.; Pandey, M.K.; Yi, Z.; Liu, M.; Aggarwal, B.B. Gossypin, a pentahydroxy glucosyl flavone, inhibits the transforming growth factor beta-activated kinase-1-mediated NF-kappaB activation pathway, leading to potentiation of apoptosis, suppression of invasion, and abrogation of osteoclastogenesis. Blood 2007, 109, 5112–5121. [Google Scholar] [CrossRef]
- Girisa, S.; Kumar, A.; Rana, V.; Parama, D.; Daimary, U.D.; Warnakulasuriya, S.; Kumar, A.P.; Kunnumakkara, A.B. From simple mouth cavities to complex oral mucosal disorders—curcuminoids as a promising therapeutic approach. ACS Pharmacol. Transl. Sci. 2021, 4, 647–665. [Google Scholar] [CrossRef]
- Daimary, U.D.; Parama, D.; Rana, V.; Banik, K.; Kumar, A.; Harsha, C.; Kunnumakkara, A.B. Emerging roles of cardamonin, a multitargeted nutraceutical in the prevention and treatment of chronic diseases. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100008. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Damayenti, Y.D.; Deka, A.; Monisha, J.; Roy, N.K.; Padmavathi, G.; Kunnumakkara, A.B. Acorus calamus: A bio-reserve of medicinal values. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Devi Daimary, U.; Girisa, S.; Parama, D.; Verma, E.; Kumar, A.; Kunnumakkara, A.B. Embelin: A novel XIAP inhibitor for the prevention and treatment of chronic diseases. J. Biochem. Mol. Toxicol. 2022, 36, e22950. [Google Scholar] [CrossRef] [PubMed]
- Jude, S.; Amalraj, A.; Kunnumakkara, A.B.; Divya, C.; Löffler, B.M.; Gopi, S. Development of validated methods and quantification of curcuminoids and curcumin metabolites and their pharmacokinetic study of oral administration of complete natural turmeric formulation (Cureit™) in human plasma via UPLC/ESI-Q-TOF-MS spectrometry. Molecules 2018, 23, 2415. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, J.H.; Sethi, G.; Kim, C.; Baek, S.H.; Nam, D.; Chung, W.S.; Kim, S.H.; Shim, B.S.; Ahn, K.S. Bergamottin, a natural furanocoumarin obtained from grapefruit juice induces chemosensitization and apoptosis through the inhibition of STAT3 signaling pathway in tumor cells. Cancer Lett. 2014, 354, 153–163. [Google Scholar] [CrossRef]
- Gopi, S.; Jacob, J.; Varma, K.; Jude, S.; Amalraj, A.; Arundhathy, C.; George, R.; Sreeraj, T.; Divya, C.; Kunnumakkara, A.B. Comparative oral absorption of curcumin in a natural turmeric matrix with two other curcumin formulations: An open-label parallel-arm study. Phytother. Res. 2017, 31, 1883–1891. [Google Scholar] [CrossRef]
- Olennikov, D.; Chirikova, N.; Tankhaeva, L. Phenolic compounds of Scutellaria baicalensis Georgi. Russ. J. Bioorganic Chem. 2010, 36, 816–824. [Google Scholar] [CrossRef]
- Li, H.-B.; Chen, F. Isolation and purification of baicalein, wogonin and oroxylin A from the medicinal plant Scutellaria baicalensis by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1074, 107–110. [Google Scholar] [CrossRef]
- Li, J.; Ding, Y.; Li, X.C.; Ferreira, D.; Khan, S.; Smillie, T.; Khan, I.A. Scuteflorins A and B, dihydropyranocoumarins from Scutellaria lateriflora. J. Nat. Prod. 2009, 72, 983–987. [Google Scholar] [CrossRef]
- Dinda, B.; SilSarma, I.; Dinda, M.; Rudrapaul, P. Oroxylum indicum (L.) Kurz, an important Asian traditional medicine: From traditional uses to scientific data for its commercial exploitation. J. Ethnopharmacol. 2015, 161, 255–278. [Google Scholar] [CrossRef]
- Sajeev, A.; Hegde, M.; Daimary, U.D.; Kumar, A.; Girisa, S.; Sethi, G.; Kunnumakkara, A.B. Modulation of diverse oncogenic signaling pathways by oroxylin A: An important strategy for both cancer prevention and treatment. Phytomedicine 2022, 105, 154369. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Herbal Remedies: Rational Western Therapy, Ayurvedic, and Other Traditional Usage, Botany; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Bhattacharjee, S.K. Use Of Flavours And Fragrances. In Handbook Of Aromatic Plants, 2nd ed.; Pointer Publishers: Jaipur, India, 2005. [Google Scholar]
- Zaveri, M.N.; Khandhar, A.P.; Jain, S.M. Quantification of baicalein, chrysin, biochanin-A and ellagic acid in root bark of Oroxylum indicum by RP-HPLC with UV detection. Eurasian J. Anal. Chem. 2008, 3, 245–257. [Google Scholar]
- Jabbar, S.; Khan, M.T.; Choudhuri, M.S.; Sil, B.K. Bioactivity studies of the individual ingredients of the Dashamularishta. Pak. J. Pharm. Sci. 2004, 17, 9–17. [Google Scholar]
- Singh, V.; Chaudhary, A.K. A review on the taxonomy, ethnobotany, chemistry and pharmacology of Oroxylum indicum vent. Indian J. Pharm. Sci. 2011, 73, 483–490. [Google Scholar] [CrossRef]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharm. 2019, 71, 1353–1369. [Google Scholar] [CrossRef]
- Awad, R.; Arnason, J.T.; Trudeau, V.; Bergeron, C.; Budzinski, J.W.; Foster, B.C.; Merali, Z. Phytochemical and biological analysis of skullcap (Scutellaria lateriflora L.): A medicinal plant with anxiolytic properties. Phytomedicine 2003, 10, 640–649. [Google Scholar] [CrossRef]
- Zhang, Z.; Lian, X.Y.; Li, S.; Stringer, J.L. Characterization of chemical ingredients and anticonvulsant activity of American skullcap (Scutellaria lateriflora). Phytomedicine 2009, 16, 485–493. [Google Scholar] [CrossRef]
- Lohani, M.; Ahuja, M.; Buabeid, M.A.; Dean, S.; Dennnis, S.; Suppiramaniam, V.; Kemppainen, B.; Dhanasekaran, M. Anti-oxidative and DNA protecting effects of flavonoids-rich Scutellaria lateriflora. Nat. Prod. Commun. 2013, 8, 1415–1418. [Google Scholar] [CrossRef]
- Rojsanga, P.; Bunsupa, S.; Sithisarn, P. Flavones contents in extracts from Oroxylum indicum seeds and plant tissue cultures. Molecules 2020, 25, 1545. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharm. 2021, 133, 110917. [Google Scholar] [CrossRef]
- Shah, R.; Mehta, C.; Wheeler, T. 131. The constitution of oroxylin-A, a yellow colouring matter from the root-bark of Oroxylum indicum, vent. J. Chem. Soc. 1936, 591–593. [Google Scholar] [CrossRef]
- Pal-Bhadra, M.; Ramaiah, M.J.; Reddy, T.L.; Krishnan, A.; Pushpavalli, S.; Babu, K.S.; Tiwari, A.K.; Rao, J.M.; Yadav, J.S.; Bhadra, U. Plant HDAC inhibitor chrysin arrest cell growth and induce p21 WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer 2012, 12, 180. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, Y.; Li, Z.; Zhang, H.; Yang, Y.; Qin, H.; Xu, Q.; Zhao, L. Oroxylin A reversed Fibronectin-induced glioma insensitivity to Temozolomide by suppressing IP3R1/AKT/β-catenin pathway. Life Sci. 2020, 260, 118411. [Google Scholar] [CrossRef]
- Liu, C.H.; Chen, M.F.; Tseng, T.L.; Chen, L.G.; Kuo, J.S.; Lee, T.J. Oroxylin a, but not vasopressin, ameliorates cardiac dysfunction of endotoxemic rats. Evid. Based Complementary Altern. Med. 2012, 2012, 408187. [Google Scholar] [CrossRef]
- Hu, R.; Chen, N.; Yao, J.; Zhao, Q.; Zhang, F.; Li, Z.Y.; You, Q.D.; Guo, Q.L. The role of Nrf2 and apoptotic signaling pathways in oroxylin A-mediated responses in HCT-116 colorectal adenocarcinoma cells and xenograft tumors. Anti-Cancer Drugs 2012, 23, 651–658. [Google Scholar] [CrossRef]
- Jeon, S.J.; Bak, H.; Seo, J.; Han, S.M.; Lee, S.H.; Han, S.H.; Kwon, K.J.; Ryu, J.H.; Cheong, J.H.; Ko, K.H.; et al. Oroxylin A induces BDNF expression on cortical neurons through adenosine A2A receptor stimulation: A possible role in neuroprotection. Biomol. Ther. 2012, 20, 27–35. [Google Scholar] [CrossRef]
- Jin, J.; Chen, S.; Wang, D.; Chen, Y.; Wang, Y.; Guo, M.; Zhou, C.; Dou, J. Oroxylin A suppresses influenza A virus replication correlating with neuraminidase inhibition and induction of IFNs. Biomed. Pharm. 2018, 97, 385–394. [Google Scholar] [CrossRef]
- Jin, H.; Lian, N.; Bian, M.; Zhang, C.; Chen, X.; Shao, J.; Wu, L.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A prevents alcohol-induced hepatic steatosis through inhibition of hypoxia inducible factor 1alpha. Chem. Biol. Interact. 2018, 285, 14–20. [Google Scholar] [CrossRef]
- Lu, Z.; Lu, N.; Li, C.; Li, F.; Zhao, K.; Lin, B.; Guo, Q. Oroxylin A inhibits matrix metalloproteinase-2/9 expression and activation by up-regulating tissue inhibitor of metalloproteinase-2 and suppressing the ERK1/2 signaling pathway. Toxicol. Lett. 2012, 209, 211–220. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, D.; Zhang, Y.; Dong, J.; Wang, J.; Niu, X. Molecular modeling reveals the novel inhibition mechanism and binding mode of three natural compounds to staphylococcal α-hemolysin. PLoS ONE 2013, 8, e80197. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, N.; Ling, Y.; Gao, Y.; Chen, Y.; Wang, L.; Hu, R.; Qi, Q.; Liu, W.; Yang, Y.; et al. Oroxylin A suppresses invasion through down-regulating the expression of matrix metalloproteinase-2/9 in MDA-MB-435 human breast cancer cells. Eur. J. Pharm. 2009, 603, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Banik, K.; Kunnumakkara, A.B.; Kubatka, P.; Koklesova, L.; Shakibaei, M. Targeting NF-κB signaling by calebin a, a compound of turmeric, in multicellular tumor microenvironment: Potential role of apoptosis induction in CRC cells. Biomedicines 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Harsha, C.; Banik, K.; Ang, H.L.; Girisa, S.; Vikkurthi, R.; Parama, D.; Rana, V.; Shabnam, B.; Khatoon, E.; Kumar, A.P.; et al. Targeting AKT/mTOR in oral cancer: Mechanisms and advances in clinical trials. Int. J. Mol. Sci. 2020, 21, 3285. [Google Scholar] [CrossRef]
- Muralimanoharan, S.B.; Kunnumakkara, A.B.; Shylesh, B.; Kulkarni, K.H.; Haiyan, X.; Ming, H.; Aggarwal, B.B.; Rita, G.; Kumar, A.P. Butanol fraction containing berberine or related compound from Nexrutine® inhibits NFκB signaling and induces apoptosis in prostate cancer cells. Prostate 2009, 69, 494–504. [Google Scholar] [CrossRef]
- Bordoloi, D.; Banik, K.; Padmavathi, G.; Vikkurthi, R.; Harsha, C.; Roy, N.K.; Singh, A.K.; Monisha, J.; Wang, H.; Kumar, A.P.; et al. TIPE2 induced the proliferation, survival, and migration of lung cancer cells through modulation of Akt/mTOR/NF-κB signaling cascade. Biomolecules 2019, 9, 836. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kunnumakkara, A.B. Molecular Targets and Therapeutic Uses of Spices: Modern Uses for Ancient Medicine; World Scientific: Singapore, 2009. [Google Scholar]
- Lu, L.; Guo, Q.; Zhao, L. Overview of oroxylin A: A promising flavonoid compound. Phytother. Res. 2016, 30, 1765–1774. [Google Scholar] [CrossRef]
- Zhu, R.; Zeng, G.; Chen, Y.; Zhang, Q.; Liu, B.; Liu, J.; Chen, H.; Li, M. Oroxylin A accelerates liver regeneration in CCl4-induced acute liver injury mice. PLoS ONE 2013, 8, e71612. [Google Scholar] [CrossRef]
- Lee, A.Y.; Kang, S.; Park, S.J.; Huang, J.; Im, D.S. Anti-allergic effect of Oroxylin A from Oroxylum indicum using in vivo and in vitro experiments. Biomol. Ther. 2016, 24, 283–290. [Google Scholar] [CrossRef]
- Ji, Y.; Han, J.; Lee, N.; Yoon, J.H.; Youn, K.; Ha, H.J.; Yoon, E.; Kim, D.H.; Jun, M. Neuroprotective effects of Baicalein, Wogonin, and Oroxylin A on Amyloid Beta-Induced Toxicity via NF-κB/MAPK pathway modulation. Molecules 2020, 25, 5087. [Google Scholar] [CrossRef]
- Zhang, Y.; Weng, Q.; Chen, J.; Li, M.; Han, J. Oroxylin A attenuates IL-1β-induced inflammatory reaction via inhibiting the activation of the ERK and PI3K/AKT signaling pathways in osteoarthritis chondrocytes. Exp. Med. 2021, 21, 388. [Google Scholar] [CrossRef]
- Wang, M.H.; Li, L.Z.; Sun, J.B.; Wu, F.H.; Liang, J.Y. A new antioxidant flavone glycoside from Scutellaria baicalensis Georgi. Nat. Prod. Res. 2014, 28, 1772–1776. [Google Scholar] [CrossRef]
- Han, Q.; Wang, H.; Xiao, C.; Fu, B.D.; Du, C.T. Oroxylin A inhibits H(2)O(2)-induced oxidative stress in PC12 cells. Nat. Prod. Res. 2017, 31, 1339–1342. [Google Scholar] [CrossRef] [PubMed]
- Li, H.N.; Nie, F.F.; Liu, W.; Dai, Q.S.; Lu, N.; Qi, Q.; Li, Z.Y.; You, Q.D.; Guo, Q.L. Apoptosis induction of oroxylin A in human cervical cancer HeLa cell line in vitro and in vivo. Toxicology 2009, 257, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhou, Y.; Qiao, C.; Ni, T.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Oroxylin A inhibits glycolysis-dependent proliferation of human breast cancer via promoting SIRT3-mediated SOD2 transcription and HIF1alpha destabilization. Cell Death Dis. 2015, 6, e1714. [Google Scholar] [CrossRef]
- Kai, J.; Yang, X.; Wang, Z.; Wang, F.; Jia, Y.; Wang, S.; Tan, S.; Chen, A.; Shao, J.; Zhang, F.; et al. Oroxylin a promotes PGC-1α/Mfn2 signaling to attenuate hepatocyte pyroptosis via blocking mitochondrial ROS in alcoholic liver disease. Free Radic. Biol. Med. 2020, 153, 89–102. [Google Scholar] [CrossRef]
- Huang, X.E.; Wei, D.; Yang, Y.N.; Chen, S.Q.; Zhu, M.; Zhang, X.M.; Yu, J. MPTP related mitochondrial pathway in oroxylin A induced-apoptosis in HepG2 cancer cells. Int. J. Clin. Exp. Pathol. 2016, 9, 11139–11148. [Google Scholar]
- Zhang, Z.; Guo, M.; Shen, M.; Li, Y.; Tan, S.; Shao, J.; Zhang, F.; Chen, A.; Wang, S.; Zheng, S. Oroxylin A regulates the turnover of lipid droplet via downregulating adipose triglyceride lipase (ATGL) in hepatic stellate cells. Life Sci. 2019, 238, 116934. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y.; Zhu, W.; Ji, M.; Xu, J.; Guo, Y.; Gao, F.; Gu, W.; Yang, X.; Zhang, C. Oroxylin A inhibits Kaposi’s sarcoma-associated herpes virus (KSHV) vIL-6-mediated lymphatic reprogramming of vascular endothelial cells through modulating PPARgamma/Prox1 axis. J. Med. Virol. 2019, 91, 463–472. [Google Scholar] [CrossRef]
- Qiao, C.; Wei, L.; Dai, Q.; Zhou, Y.; Yin, Q.; Li, Z.; Xiao, Y.; Guo, Q.; Lu, N. UCP2-related mitochondrial pathway participates in oroxylin A-induced apoptosis in human colon cancer cells. J. Cell. Physiol. 2015, 230, 1054–1063. [Google Scholar] [CrossRef]
- Ni, T.; He, Z.; Dai, Y.; Yao, J.; Guo, Q.; Wei, L. Oroxylin A suppresses the development and growth of colorectal cancer through reprogram of HIF1α-modulated fatty acid metabolism. Cell Death Dis. 2017, 8, e2865. [Google Scholar] [CrossRef]
- Li, H.; Lu, N.; Yu, X.; Liu, X.; Hu, P.; Zhu, Y.; Shen, L.; Xu, J.; Li, Z.; Guo, Q.; et al. Oroxylin A, a natural compound, mitigates the negative effects of TNFalpha-treated acute myelogenous leukemia cells. Carcinogenesis 2018, 39, 1292–1303. [Google Scholar] [CrossRef]
- Liu, W.; Mu, R.; Nie, F.F.; Yang, Y.; Wang, J.; Dai, Q.S.; Lu, N.; Qi, Q.; Rong, J.J.; Hu, R.; et al. MAC-related mitochondrial pathway in oroxylin-A-induced apoptosis in human hepatocellular carcinoma HepG2 cells. Cancer Lett. 2009, 284, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, F.; Wang, Y.; Cai, M.; Wang, Q.; Guo, Q.; Li, Z.; Hu, R. Oroxylin A inhibits colitis-associated carcinogenesis through modulating the IL-6/STAT3 signaling pathway. Inflamm. Bowel. Dis. 2013, 19, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and chronic diseases: How are they linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Saghari, S.; Bassiri, F.; Raesi, R.; Zarrabi, A.; Hushmandi, K.; Sethi, G.; Tergaonkar, V. NF-κB as a regulator of cancer metastasis and therapy response: A focus on epithelial-mesenchymal transition. J. Cell. Physiol. 2022, 237, 2770–2795. [Google Scholar] [CrossRef]
- Morgan, D.; Garg, M.; Tergaonkar, V.; Tan, S.Y.; Sethi, G. Pharmacological significance of the non-canonical NF-κB pathway in tumorigenesis. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188449. [Google Scholar] [CrossRef]
- Kim, C.; Lee, S.G.; Yang, W.M.; Arfuso, F.; Um, J.Y.; Kumar, A.P.; Bian, J.; Sethi, G.; Ahn, K.S. Formononetin-induced oxidative stress abrogates the activation of STAT3/5 signaling axis and suppresses the tumor growth in multiple myeloma preclinical model. Cancer Lett. 2018, 431, 123–141. [Google Scholar] [CrossRef]
- Hiremath, I.S.; Goel, A.; Warrier, S.; Kumar, A.P.; Sethi, G.; Garg, M. The multidimensional role of the Wnt/β-catenin signaling pathway in human malignancies. J. Cell. Physiol. 2022, 237, 199–238. [Google Scholar] [CrossRef]
- Dai, X.; Ahn, K.S.; Kim, C.; Siveen, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Shi, J.; Kumar, A.P.; Wang, L.Z.; et al. Ascochlorin, an isoprenoid antibiotic inhibits growth and invasion of hepatocellular carcinoma by targeting STAT3 signaling cascade through the induction of PIAS3. Mol. Oncol. 2015, 9, 818–833. [Google Scholar] [CrossRef]
- Shen, M.; Guo, M.; Wang, Z.; Li, Y.; Kong, D.; Shao, J.; Tan, S.; Chen, A.; Zhang, F.; Zhang, Z.; et al. ROS-dependent inhibition of the PI3K/Akt/mTOR signaling is required for Oroxylin A to exert anti-inflammatory activity in liver fibrosis. Int. Immunopharmacol. 2020, 85, 106637. [Google Scholar] [CrossRef]
- Xuan, L.; Zhang, J.; Ji, J.; Hu, J.; Li, F. Oroxylin A exerts its antitumor effects in human gallbladder cancer via inhibition of the PTEN/PI3K/AKT signaling pathway. Biol. Pharm. Bull. 2020, 43, 1511–1518. [Google Scholar] [CrossRef]
- Wei, L.; Dai, Y.; Zhou, Y.; He, Z.; Yao, J.; Zhao, L.; Guo, Q.; Yang, L. Oroxylin A activates PKM1/HNF4 alpha to induce hepatoma differentiation and block cancer progression. Cell Death Dis. 2017, 8, e2944. [Google Scholar] [CrossRef]
- Yang, M.H.; Lee, J.H.; Ko, J.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Brassinin represses invasive potential of lung carcinoma cells through deactivation of PI3K/Akt/mTOR signaling cascade. Molecules 2019, 24, 1584. [Google Scholar] [CrossRef]
- Ong, P.S.; Wang, L.Z.; Dai, X.; Tseng, S.H.; Loo, S.J.; Sethi, G. Judicious toggling of mTOR activity to combat insulin resistance and cancer: Current evidence and perspectives. Front. Pharm. 2016, 7, 395. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef]
- Siegel, R.; DeSantis, C.; Virgo, K.; Stein, K.; Mariotto, A.; Smith, T.; Cooper, D.; Gansler, T.; Lerro, C.; Fedewa, S. Cancer treatment and survivorship statistics, 2012. CA A Cancer J. Clin. 2012, 62, 220–241. [Google Scholar] [CrossRef]
- Sleire, L.; Førde, H.E.; Netland, I.A.; Leiss, L.; Skeie, B.S.; Enger, P.Ø. Drug repurposing in cancer. Pharmacol. Res. 2017, 124, 74–91. [Google Scholar] [CrossRef]
- Mondal, M.; Guo, J.; He, P.; Zhou, D. Recent advances of oncolytic virus in cancer therapy. Hum. Vaccin. Immunother. 2020, 16, 2389–2402. [Google Scholar] [CrossRef]
- Roy, N.K.; Bordoloi, D.; Monisha, J.; Padmavathi, G.; Kotoky, J.; Golla, R.; Kunnumakkara, A.B. Specific targeting of Akt kinase isoforms: Taking the precise path for prevention and treatment of cancer. Curr. Drug Targets 2017, 18, 421–435. [Google Scholar] [CrossRef]
- Monisha, J.; Padmavathi, G.; Roy, N.K.; Deka, A.; Bordoloi, D.; Anip, A.; Kunnumakkara, A.B. NF-κB blockers gifted by mother nature: Prospectives in cancer cell chemosensitization. Curr. Pharm. Des. 2016, 22, 4173–4200. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Rana, V.; Parama, D.; Banik, K.; Girisa, S.; Henamayee, S.; Thakur, K.K.; Dutta, U.; Garodia, P.; Gupta, S.C.; et al. COVID-19, cytokines, inflammation, and spices: How are they related? Life Sci. 2021, 284, 119201. [Google Scholar] [CrossRef]
- Sethi, G.; Ahn, K.S.; Sung, B.; Kunnumakkara, A.B.; Chaturvedi, M.M.; Aggarwal, B.B. SH-5, an AKT inhibitor potentiates apoptosis and inhibits invasion through the suppression of anti-apoptotic, proliferative and metastatic gene products regulated by IκBα kinase activation. Biochem. Pharmacol. 2008, 76, 1404–1416. [Google Scholar] [CrossRef]
- Goyal, B.; Yadav, S.R.M.; Awasthee, N.; Gupta, S.; Kunnumakkara, A.B.; Gupta, S.C. Diagnostic, prognostic, and therapeutic significance of long non-coding RNA MALAT1 in cancer. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2021, 1875, 188502. [Google Scholar] [CrossRef]
- Khatoon, E.; Banik, K.; Harsha, C.; Sailo, B.L.; Thakur, K.K.; Khwairakpam, A.D.; Vikkurthi, R.; Devi, T.B.; Gupta, S.C.; Kunnumakkara, A.B. Phytochemicals in cancer cell chemosensitization: Current knowledge and future perspectives. Semin. Cancer Biol. 2022, 80, 306–339. [Google Scholar] [CrossRef]
- Maruthanila, V.L.; Elancheran, R.; Kunnumakkara, A.B.; Kabilan, S.; Kotoky, J. Recent development of targeted approaches for the treatment of breast cancer. Breast Cancer 2017, 24, 191–219. [Google Scholar] [CrossRef]
- Thakur, K.K.; Bordoloi, D.; Kunnumakkara, A.B. Alarming burden of triple-negative breast cancer in India. Clin. Breast Cancer 2018, 18, e393–e399. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Golani, A.; Kumar, L.D. EMT in breast cancer metastasis: An interplay of microRNAs, signaling pathways and circulating tumor cells. Front. Biosci. 2020, 25, 979–1010. [Google Scholar] [CrossRef] [PubMed]
- Thakur, K.K.; Kumar, A.; Banik, K.; Verma, E.; Khatoon, E.; Harsha, C.; Sethi, G.; Gupta, S.C.; Kunnumakkara, A.B. Long noncoding RNAs in triple-negative breast cancer: A new frontier in the regulation of tumorigenesis. J. Cell. Physiol. 2021, 236, 7938–7965. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.Y.; Shanmugam, M.K.; Sethi, G.; Bishayee, A. Potential role of targeted therapies in the treatment of triple-negative breast cancer. Anticancer Drugs 2016, 27, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Ahn, K.S.; Hsu, A.; Woo, C.C.; Yuan, Y.; Tan, K.H.B.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Koh, A.P.F.; et al. Thymoquinone inhibits bone metastasis of breast cancer cells through abrogation of the CXCR4 signaling axis. Front. Pharm. 2018, 9, 1294. [Google Scholar] [CrossRef]
- Sun, X.; Chang, X.; Wang, Y.; Xu, B.; Cao, X. Oroxylin A suppresses the cell proliferation, migration, and EMT via NF-kappaB signaling pathway in human breast cancer cells. Biomed Res. Int. 2019, 2019, 9241769. [Google Scholar] [CrossRef]
- Katoch, A.; Nayak, D.; Faheem, M.M.; Kumar, A.; Sahu, P.K.; Gupta, A.P.; Kumar, L.D.; Goswami, A. Natural podophyllotoxin analog 4DPG attenuates EMT and colorectal cancer progression via activation of checkpoint kinase 2. Cell Death Discov. 2021, 7, 25. [Google Scholar] [CrossRef]
- Chakraborty, S.; Kumar, A.; Faheem, M.M.; Katoch, A.; Kumar, A.; Jamwal, V.L.; Nayak, D.; Golani, A.; Rasool, R.U.; Ahmad, S.M.; et al. Vimentin activation in early apoptotic cancer cells errands survival pathways during DNA damage inducer CPT treatment in colon carcinoma model. Cell Death Dis. 2019, 10, 467. [Google Scholar] [CrossRef]
- Qi, L.; Ding, Y. Screening of differentiation-specific molecular biomarkers for colon cancer. Cell Physiol. Biochem. 2018, 46, 2543–2550. [Google Scholar] [CrossRef]
- Ha, J.; Zhao, L.; Zhao, Q.; Yao, J.; Zhu, B.B.; Lu, N.; Ke, X.; Yang, H.Y.; Li, Z.; You, Q.D.; et al. Oroxylin A improves the sensitivity of HT-29 human colon cancer cells to 5-FU through modulation of the COX-2 signaling pathway. Biochem. Cell Biol. 2012, 90, 521–531. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, Z.; Wang, J.; Yang, L.; Zhao, Q.; Wang, J.; Qi, Q.; Mu, R.; You, Q.D.; Guo, Q.L. Synergistic effect of 5-fluorouracil and the flavanoid oroxylin A on HepG2 human hepatocellular carcinoma and on H22 transplanted mice. Cancer Chemother. Pharm. 2010, 65, 481–489. [Google Scholar] [CrossRef]
- Huo, T.X.; Wang, X.P.; Yu, Z.; Kong, B.; He, Y.; Guo, Q.L.; Zhang, X.B.; Qiang, L. Oroxylin A inhibits the migration of hepatocellular carcinoma cells by inducing NAG-1 expression. Acta Pharm. Sin. 2022, 43, 724–734. [Google Scholar] [CrossRef]
- Weller, M.; Wick, W.; Aldape, K.; Brada, M.; Berger, M.; Pfister, S.M.; Nishikawa, R.; Rosenthal, M.; Wen, P.Y.; Stupp, R. Glioma. Nat. Rev. Dis. Primers 2015, 1, 15017. [Google Scholar] [CrossRef]
- Chen, R.; Smith-Cohn, M.; Cohen, A.L.; Colman, H. Glioma subclassifications and their clinical significance. Neurotherapeutics 2017, 14, 284–297. [Google Scholar] [CrossRef]
- Bradshaw, A.; Wickremsekera, A.; Tan, S.T.; Peng, L.; Davis, P.F.; Itinteang, T. Cancer stem cell hierarchy in glioblastoma multiforme. Front. Surg. 2016, 3, 21. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Gamit, N.; Patil, M.; Arfuso, F.; Sethi, G.; Dharmarajan, A.; Kumar, A.P.; Warrier, S. Stemness, pluripotentiality, and Wnt antagonism: sFRP4, a Wnt antagonist mediates pluripotency and stemness in glioblastoma. Cancers 2018, 11, 25. [Google Scholar] [CrossRef]
- Sun, T.; Xu, Y.J.; Jiang, S.Y.; Xu, Z.; Cao, B.Y.; Sethi, G.; Zeng, Y.Y.; Kong, Y.; Mao, X.L. Suppression of the USP10/CCND1 axis induces glioblastoma cell apoptosis. Acta Pharm. Sin. 2021, 42, 1338–1346. [Google Scholar] [CrossRef]
- Zou, M.; Hu, C.; You, Q.; Zhang, A.; Wang, X.; Guo, Q. Oroxylin A induces autophagy in human malignant glioma cells via the mTOR-STAT3-Notch signaling pathway. Mol. Carcinog. 2015, 54, 1363–1375. [Google Scholar] [CrossRef]
- Arora, L.; Kumar, A.P.; Arfuso, F.; Chng, W.J.; Sethi, G. The role of signal transducer and activator of transcription 3 (STAT3) and its targeted inhibition in hematological malignancies. Cancers 2018, 10, 327. [Google Scholar] [CrossRef]
- Kirtonia, A.; Pandya, G.; Sethi, G.; Pandey, A.K.; Das, B.C.; Garg, M. A comprehensive review of genetic alterations and molecular targeted therapies for the implementation of personalized medicine in acute myeloid leukemia. J. Mol. Med. 2020, 98, 1069–1091. [Google Scholar] [CrossRef] [PubMed]
- Hui, H.; Zhang, X.; Li, H.; Liu, X.; Shen, L.; Zhu, Y.; Xu, J.; Guo, Q.; Lu, N. Oroxylin A, a natural anticancer flavonoid compound, induces differentiation of t(8;21)-positive Kasumi-1 and primary acute myeloid leukemia cells. J. Cancer Res. Clin. Oncol. 2016, 142, 1449–1459. [Google Scholar] [CrossRef]
- Li, W.; Ding, Q.; Ding, Y.; Lu, L.; Wang, X.; Zhang, Y.; Zhang, X.; Guo, Q.; Zhao, L. Oroxylin A reverses the drug resistance of chronic myelogenous leukemia cells to imatinib through CXCL12/CXCR7 axis in bone marrow microenvironment. Mol. Carcinog. 2017, 56, 863–876. [Google Scholar] [CrossRef]
- Wang, L.; Syn, N.L.; Subhash, V.V.; Any, Y.; Thuya, W.L.; Cheow, E.S.H.; Kong, L.; Yu, F.; Peethala, P.C.; Wong, A.L.; et al. Pan-HDAC inhibition by panobinostat mediates chemosensitization to carboplatin in non-small cell lung cancer via attenuation of EGFR signaling. Cancer Lett. 2018, 417, 152–160. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Shanmugam, M.K.; Narula, A.S.; Kim, C.; Lee, J.H.; Namjoshi, O.A.; Blough, B.E.; Sethi, G.; Ahn, K.S. Oxymatrine attenuates tumor growth and deactivates STAT5 signaling in a lung cancer xenograft model. Cancers 2019, 11, 49. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Lee, S.G.; Sethi, G.; Ahn, K.S. Ophiopogonin D, a steroidal glycoside abrogates STAT3 signaling cascade and exhibits anti-cancer activity by causing GSH/GSSG imbalance in lung carcinoma. Cancers 2018, 10, 427. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, L.L.; Li, H.; Liu, X.; Yu, X.X.; Hu, P.; Hui, H.; Guo, Q.L.; Zhang, S. Oroxylin A inhibits the generation of Tregs in non-small cell lung cancer. Oncotarget 2017, 8, 49395–49408. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Li, W.; Xu, Y.; Zhuo, Y.; Li, M.; He, Y.; Wang, X.; Guo, Q.; Zhao, L.; et al. Oroxylin A reverses hypoxia-induced cisplatin resistance through inhibiting HIF-1alpha mediated XPC transcription. Oncogene 2020, 39, 6893–6905. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Qian, X.; Ge, Y.; Yang, B.; Wang, F.; Guan, Z.; Cai, J. Oroxylin a could be a promising radiosensitizer for esophageal squamous cell carcinoma by inducing G2/M arrest and activating apoptosis. Pathol. Oncol. Res. 2017, 23, 323–328. [Google Scholar] [CrossRef]
- Huang, H.; Cai, H.; Zhang, L.; Hua, Z.; Shi, J.; Wei, Y. Oroxylin A inhibits carcinogen-induced skin tumorigenesis through inhibition of inflammation by regulating SHCBP1 in mice. Int. Immunopharmacol. 2020, 80, 106123. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Zhao, D. Epidemiological features of cardiovascular disease in Asia. JACC Asia 2021, 1, 1–13. [Google Scholar] [CrossRef]
- Jagannathan, R.; Patel, S.A.; Ali, M.K.; Narayan, K. Global updates on cardiovascular disease mortality trends and attribution of traditional risk factors. Curr. Diabetes Rep. 2019, 19, 44. [Google Scholar] [CrossRef]
- Octavia, Y.; Tocchetti, C.G.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-induced cardiomyopathy: From molecular mechanisms to therapeutic strategies. J. Mol. Cell. Cardiol. 2012, 52, 1213–1225. [Google Scholar] [CrossRef]
- Takemura, G.; Fujiwara, H. Doxorubicin-induced cardiomyopathy from the cardiotoxic mechanisms to management. Prog. Cardiovasc. Dis. 2007, 49, 330–352. [Google Scholar] [CrossRef]
- Zhang, W.-B.; Zheng, Y.-F.; Wu, Y.-G. Protective effects of oroxylin a against doxorubicin-induced cardiotoxicity via the activation of Sirt1 in mice. Oxidative Med. Cell. Longev. 2021, 2021, 6610543. [Google Scholar] [CrossRef]
- Mohammad, S.; Thiemermann, C. Role of Metabolic Endotoxemia in Systemic Inflammation and Potential Interventions. Front. Immunol. 2020, 11, 594150. [Google Scholar] [CrossRef]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Kitabatake, H.; Tanaka, N.; Fujimori, N.; Komatsu, M.; Okubo, A.; Kakegawa, K.; Kimura, T.; Sugiura, A.; Yamazaki, T.; Shibata, S.; et al. Association between endotoxemia and histological features of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 712–722. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Chaturvedi, M.M.; Aggarwal, B.B. Simvastatin, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor, suppresses osteoclastogenesis induced by receptor activator of nuclear factor-kappaB ligand through modulation of NF-kappaB pathway. Int. J. Cancer 2008, 123, 1733–1740. [Google Scholar] [CrossRef]
- Glaros, T.G.; Chang, S.; Gilliam, E.A.; Maitra, U.; Deng, H.; Li, L. Causes and consequences of low grade endotoxemia and inflammatory diseases. Front. Biosci. -Sch. 2013, 5, 754–765. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M.; Kibbe, M.R. Improving Lower Extremity Functioning in Peripheral Artery Disease: Exercise, Endovascular Revascularization, or Both? JAMA 2017, 317, 689–690. [Google Scholar] [CrossRef]
- Cooke, J.P.; Losordo, D.W. Modulating the vascular response to limb ischemia: Angiogenic and cell therapies. Circ. Res. 2015, 116, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Lu, H.; Liang, W.; Garcia-Barrio, M.T.; Guo, Y.; Zhang, J.; Zhu, T.; Hao, Y.; Zhang, J.; Chen, Y.E. Endothelial TFEB (Transcription Factor EB) Positively Regulates Postischemic Angiogenesis. Circ. Res. 2018, 122, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, L.; Li, C.; Shi, H.; Wang, Q.; Yang, W.; Fang, L.; Leng, Y.; Sun, W.; Li, M.; et al. Oroxylin a attenuates limb ischemia by promoting angiogenesis via modulation of endothelial cell migration. Front. Pharm. 2021, 12, 705617. [Google Scholar] [CrossRef] [PubMed]
- Pinzani, M.; Rombouts, K. Liver fibrosis: From the bench to clinical targets. Dig. Liver Dis. 2004, 36, 231–242. [Google Scholar] [CrossRef]
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary epidemiology of chronic liver disease and cirrhosis. Clin. Gastroenterol. Hepatol. 2020, 18, 2650–2666. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef]
- Singh, Y.P.; Girisa, S.; Banik, K.; Ghosh, S.; Swathi, P.; Deka, M.; Padmavathi, G.; Kotoky, J.; Sethi, G.; Fan, L. Potential application of zerumbone in the prevention and therapy of chronic human diseases. J. Funct. Foods 2019, 53, 248–258. [Google Scholar] [CrossRef]
- Shu, Y.; Liu, X.; Huang, H.; Wen, Q.; Shu, J. Research progress of natural compounds in anti-liver fibrosis by affecting autophagy of hepatic stellate cells. Mol. Biol. Rep. 2021, 48, 1915–1924. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, X.; Li, J. Protective effect of oroxylin A against lipopolysaccharide and/or D-galactosamine-induced acute liver injury in mice. J. Surg. Res. 2015, 195, 522–528. [Google Scholar] [CrossRef]
- Bonder, A.; Tapper, E.B.; Afdhal, N.H. Contemporary assessment of hepatic fibrosis. Clin. Liver Dis. 2015, 19, 123–134. [Google Scholar] [CrossRef]
- Friedman, S.L. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Z.; Yao, Z.; Wang, L.; Zhang, F.; Shao, J.; Chen, A.; Zheng, S. Activation of autophagy is required for Oroxylin A to alleviate carbon tetrachloride-induced liver fibrosis and hepatic stellate cell activation. Int. Immunopharmacol. 2018, 56, 148–155. [Google Scholar] [CrossRef]
- Wang, F.; Jia, Y.; Li, M.; Wang, L.; Shao, J.; Guo, Q.; Tan, S.; Ding, H.; Chen, A.; Zhang, F.; et al. Blockade of glycolysis-dependent contraction by oroxylin a via inhibition of lactate dehydrogenase-a in hepatic stellate cells. Cell Commun. Signal. 2019, 17, 11. [Google Scholar] [CrossRef]
- Gandhi, C.R. Hepatic stellate cell activation and pro-fibrogenic signals. J. Hepatol. 2017, 67, 1104–1105. [Google Scholar] [CrossRef] [PubMed]
- Bian, M.; He, J.; Jin, H.; Lian, N.; Shao, J.; Guo, Q.; Wang, S.; Zhang, F.; Zheng, S. Oroxylin A induces apoptosis of activated hepatic stellate cells through endoplasmic reticulum stress. Apoptosis 2019, 24, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Pessione, F.; Ramond, M.J.; Peters, L.; Pham, B.N.; Batel, P.; Rueff, B.; Valla, D.C. Five-year survival predictive factors in patients with excessive alcohol intake and cirrhosis. Effect of alcoholic hepatitis, smoking and abstinence. Liver Int. 2003, 23, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lian, N.; Bian, M.; Zhang, C.; Chen, X.; Shao, J.; Wu, L.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A inhibits ethanol-induced hepatocyte senescence via YAP pathway. Cell Prolif. 2018, 51, e12431. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Lee, Y.; Gullen, E.A.; Cheng, Y.C. Impacts of baicalein analogs with modification of the 6th position of A ring on the activity toward NF-kappaB-, AP-1-, or CREB-mediated transcription. Bioorg. Med. Chem. Lett. 2008, 18, 5046–5049. [Google Scholar] [CrossRef]
- Parisi, V.; Oddone, F.; Ziccardi, L.; Roberti, G.; Coppola, G.; Manni, G. Citicoline and retinal ganglion cells: Effects on morphology and function. Curr. Neuropharmacol. 2018, 16, 919–932. [Google Scholar] [CrossRef]
- Shindler, K.S.; Ventura, E.; Dutt, M.; Rostami, A. Inflammatory demyelination induces axonal injury and retinal ganglion cell apoptosis in experimental optic neuritis. Exp. Eye Res. 2008, 87, 208–213. [Google Scholar] [CrossRef]
- Potilinski, M.C.; Lorenc, V.; Perisset, S.; Gallo, J.E. Mechanisms behind retinal ganglion cell loss in diabetes and therapeutic approach. Int. J. Mol. Sci. 2020, 21, 2351. [Google Scholar] [CrossRef]
- Levin, L.A.; Louhab, A. Apoptosis of retinal ganglion cells in anterior ischemic optic neuropathy. Arch. Ophthalmol. 1996, 114, 488–491. [Google Scholar] [CrossRef]
- Quigley, H.A. Ganglion cell death in glaucoma: Pathology recapitulates ontogeny. Aust. N. Z. J. Ophthalmol. 1995, 23, 85–91. [Google Scholar] [CrossRef]
- Lin, S.F.; Chien, J.Y.; Kapupara, K.; Huang, C.F.; Huang, S.P. Oroxylin A promotes retinal ganglion cell survival in a rat optic nerve crush model. PLoS ONE 2017, 12, e0178584. [Google Scholar] [CrossRef]
- Biousse, V.; Newman, N.J. Ischemic optic neuropathies. N. Engl. J. Med. 2015, 372, 2428–2436. [Google Scholar] [CrossRef]
- Luneau, K.; Newman, N.J.; Biousse, V. Ischemic optic neuropathies. Neurologist 2008, 14, 341–354. [Google Scholar] [CrossRef]
- Chien, J.Y.; Lin, S.F.; Chou, Y.Y.; Huang, C.F.; Huang, S.P. Protective effects of Oroxylin A on retinal ganglion cells in experimental model of anterior ischemic optic neuropathy. Antioxidants 2021, 10, 902. [Google Scholar] [CrossRef]
- Hanania, N.A. Targeting airway inflammation in asthma: Current and future therapies. Chest 2008, 133, 989–998. [Google Scholar] [CrossRef]
- Goh, F.Y.; Upton, N.; Guan, S.; Cheng, C.; Shanmugam, M.K.; Sethi, G.; Leung, B.P.; Wong, W.S. Fisetin, a bioactive flavonol, attenuates allergic airway inflammation through negative regulation of NF-κB. Eur. J. Pharm. 2012, 679, 109–116. [Google Scholar] [CrossRef]
- Doeing, D.C.; Solway, J. Airway smooth muscle in the pathophysiology and treatment of asthma. J. Appl. Physiol. 2013, 114, 834–843. [Google Scholar] [CrossRef]
- Zhou, D.G.; Diao, B.Z.; Zhou, W.; Feng, J.L. Oroxylin A inhibits allergic airway inflammation in ovalbumin (OVA)-induced asthma murine model. Inflammation 2016, 39, 867–872. [Google Scholar] [CrossRef]
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef]
- Ponder, A.; Long, M.D. A clinical review of recent findings in the epidemiology of inflammatory bowel disease. Clin. Epidemiol. 2013, 5, 237–247. [Google Scholar] [CrossRef]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef]
- Biasi, F.; Leonarduzzi, G.; Oteiza, P.I.; Poli, G. Inflammatory bowel disease: Mechanisms, redox considerations, and therapeutic targets. Antioxid. Redox Signal. 2013, 19, 1711–1747. [Google Scholar] [CrossRef]
- Zaki, M.H.; Lamkanfi, M.; Kanneganti, T.D. The Nlrp3 inflammasome: Contributions to intestinal homeostasis. Trends Immunol. 2011, 32, 171–179. [Google Scholar] [CrossRef]
- Bai, D.; Sun, T.; Zhao, J.; Du, J.; Bu, X.; Cao, W.; Zhao, Y.; Lu, N. Oroxylin A maintains the colonic mucus barrier to reduce disease susceptibility by reconstituting a dietary fiber-deprived gut microbiota. Cancer Lett. 2021, 515, 73–85. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, X.; Zhang, X.; Tang, J.; Li, Z.; Wang, Q.; Hu, R. Oroxylin A inhibits colitis by inactivating NLRP3 inflammasome. Oncotarget 2017, 8, 58903–58917. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Y.; Zhao, Y.; Ding, Y.; Zhang, X.; Kong, L.; Li, Z.; Guo, Q.; Zhao, L. Oroxyloside prevents dextran sulfate sodium-induced experimental colitis in mice by inhibiting NF-κB pathway through PPARγ activation. Biochem. Pharmacol. 2016, 106, 70–81. [Google Scholar] [CrossRef]
- Dahaghin, S.; Bierma-Zeinstra, S.M.; Ginai, A.Z.; Pols, H.A.; Hazes, J.M.; Koes, B.W. Prevalence and pattern of radiographic hand osteoarthritis and association with pain and disability (the Rotterdam study). Ann. Rheum. Dis. 2005, 64, 682–687. [Google Scholar] [CrossRef]
- Oliveria, S.A.; Felson, D.T.; Reed, J.I.; Cirillo, P.A.; Walker, A.M. Incidence of symptomatic hand, hip, and knee osteoarthritis among patients in a health maintenance organization. Arthritis Rheum 1995, 38, 1134–1141. [Google Scholar] [CrossRef]
- Felson, D.T. Epidemiology of hip and knee osteoarthritis. Epidemiol. Rev. 1988, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.H.; Zheng, G.; Zhong, X.Y.; Lin, Z.H.; Yang, S.W.; Liu, H.X.; Shang, P. Oroxylin A attenuates osteoarthritis progression by dual inhibition of cell inflammation and hypertrophy. Food Funct. 2021, 12, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Ferro, F.; Elefante, E.; Luciano, N.; Talarico, R.; Todoerti, M. One year in review 2017: Novelties in the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 721–734. [Google Scholar]
- Bortoluzzi, A.; Furini, F.; Generali, E.; Silvagni, E.; Luciano, N.; Scirè, C.A. One year in review 2018: Novelties in the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2018, 36, 347–361. [Google Scholar]
- Xu, T.; Liu, S.; Zhao, J.; Feng, G.; Pi, Z.; Song, F.; Liu, Z. A study on the effective substance of the Wu-tou formula based on the metabonomic method using UPLC-Q-TOF-HDMS. Mol. Biosyst. 2015, 11, 3081–3091. [Google Scholar] [CrossRef]
- Jacob, J.; Amalraj, A.; Raj, K.K.J.; Divya, C.; Kunnumakkara, A.B.; Gopi, S. A novel bioavailable hydrogenated curcuminoids formulation (CuroWhite™) improves symptoms and diagnostic indicators in rheumatoid arthritis patients-A randomized, double blind and placebo controlled study. J. Tradit. Complementary Med. 2019, 9, 346–352. [Google Scholar] [CrossRef]
- Wang, Y.L.; Gao, J.M.; Xing, L.Z. Therapeutic potential of Oroxylin A in rheumatoid arthritis. Int. Immunopharmacol. 2016, 40, 294–299. [Google Scholar] [CrossRef]
- Polanczyk, G.; de Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The worldwide prevalence of ADHD: A systematic review and metaregression analysis. Am. J. Psychiatry 2007, 164, 942–948. [Google Scholar] [CrossRef]
- Faraone, S.V.; Asherson, P.; Banaschewski, T.; Biederman, J.; Buitelaar, J.K.; Ramos-Quiroga, J.A.; Rohde, L.A.; Sonuga-Barke, E.J.; Tannock, R.; Franke, B. Attention-deficit/hyperactivity disorder. Nat. Rev. Dis. Primers 2015, 1, 15020. [Google Scholar] [CrossRef]
- Thapar, A.; Van den Bree, M.; Fowler, T.; Langley, K.; Whittinger, N. Predictors of antisocial behaviour in children with attention deficit hyperactivity disorder. Eur. Child Adolesc. Psychiatry 2006, 15, 118–125. [Google Scholar] [CrossRef]
- Van den Berg, A.E.; Van den Berg, C. A comparison of children with ADHD in a natural and built setting. Child Care Health Dev. 2011, 37, 430–439. [Google Scholar] [CrossRef]
- Schachter, H.M.; Pham, B.; King, J.; Langford, S.; Moher, D. How efficacious and safe is short-acting methylphenidate for the treatment of attention-deficit disorder in children and adolescents? A meta-analysis. Can. Med. Assoc. J. 2001, 165, 1475–1488. [Google Scholar]
- Wilens, T.E.; Kwon, A.; Tanguay, S.; Chase, R.; Moore, H.; Faraone, S.V.; Biederman, J. Characteristics of adults with attention deficit hyperactivity disorder plus substance use disorder: The role of psychiatric comorbidity. Am. J. Addict. 2005, 14, 319–327. [Google Scholar] [CrossRef]
- Yoon, S.Y.; dela Peña, I.; Kim, S.M.; Woo, T.S.; Shin, C.Y.; Son, K.H.; Park, H.; Lee, Y.S.; Ryu, J.H.; Jin, M.; et al. Oroxylin A improves attention deficit hyperactivity disorder-like behaviors in the spontaneously hypertensive rat and inhibits reuptake of dopamine in vitro. Arch. Pharm. Res. 2013, 36, 134–140. [Google Scholar] [CrossRef]
- Selkoe, D.J. The origins of Alzheimer disease: A is for amyloid. Jama 2000, 283, 1615–1617. [Google Scholar] [CrossRef]
- Segaran, R.C.; Chan, L.Y.; Wang, H.; Sethi, G.; Tang, F.R. Neuronal development-related miRNAs as biomarkers for Alzheimer’s disease, depression, schizophrenia and ionizing radiation exposure. Curr. Med. Chem. 2021, 28, 19–52. [Google Scholar] [CrossRef]
- Chai, Y.L.; Chong, J.R.; Raquib, A.R.; Xu, X.; Hilal, S.; Venketasubramanian, N.; Tan, B.Y.; Kumar, A.P.; Sethi, G.; Chen, C.P.; et al. Plasma osteopontin as a biomarker of Alzheimer’s disease and vascular cognitive impairment. Sci. Rep. 2021, 11, 4010. [Google Scholar] [CrossRef]
- Kukull, W.A.; Bowen, J.D. Dementia epidemiology. Med. Clin. N. Am. 2002, 86, 573–590. [Google Scholar] [CrossRef]
- Teng, E.; Lu, P.H.; Cummings, J.L. Neuropsychiatric symptoms are associated with progression from mild cognitive impairment to Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2007, 24, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Doody, R.S.; Pavlik, V.; Massman, P.; Rountree, S.; Darby, E.; Chan, W. Predicting progression of Alzheimer’s disease. Alzheimer’s Res. Ther. 2010, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Ahadieh, S.; Corrigan, B.; French, J.; Fullerton, T.; Tensfeldt, T.; Group, A.s.D.W. Disease progression meta-analysis model in Alzheimer’s disease. Alzheimer’s Dement. 2010, 6, 39–53. [Google Scholar] [CrossRef] [PubMed]
- DeKosky, S.T.; Marek, K. Looking backward to move forward: Early detection of neurodegenerative disorders. Science 2003, 302, 830–834. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.; Jeon, S.J.; Son, K.H.; Lee, S.; Yoon, B.H.; Cheong, J.H.; Ko, K.H.; Ryu, J.H. The effects of acute and repeated oroxylin A treatments on Abeta(25-35)-induced memory impairment in mice. Neuropharmacology 2008, 55, 639–647. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeon, S.J.; Son, K.H.; Jung, J.W.; Lee, S.; Yoon, B.H.; Lee, J.J.; Cho, Y.W.; Cheong, J.H.; Ko, K.H.; et al. The ameliorating effect of oroxylin A on scopolamine-induced memory impairment in mice. Neurobiol. Learn. Mem. 2007, 87, 536–546. [Google Scholar] [CrossRef]
- Nuttall, F.Q. Body Mass Index: Obesity, BMI, and Health: A Critical Review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef]
- Skelton, J.A.; Irby, M.B.; Grzywacz, J.G.; Miller, G. Etiologies of obesity in children: Nature and nurture. Pediatr. Clin. N. Am. 2011, 58, 1333–1354. [Google Scholar] [CrossRef]
- McAllister, E.J.; Dhurandhar, N.V.; Keith, S.W.; Aronne, L.J.; Barger, J.; Baskin, M.; Benca, R.M.; Biggio, J.; Boggiano, M.M.; Eisenmann, J.C.; et al. Ten putative contributors to the obesity epidemic. Crit. Rev. Food Sci. Nutr. 2009, 49, 868–913. [Google Scholar] [CrossRef]
- Calle, E.E.; Kaaks, R. Overweight, obesity and cancer: Epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef]
- Skelton, J.A.; DeMattia, L.; Miller, L.; Olivier, M. Obesity and its therapy: From genes to community action. Pediatr. Clin. N. Am. 2006, 53, 777–794. [Google Scholar] [CrossRef][Green Version]
- Mangal, P.; Khare, P.; Jagtap, S.; Bishnoi, M.; Kondepudi, K.K.; Bhutani, K.K. Screening of six Ayurvedic medicinal plants for anti-obesity potential: An investigation on bioactive constituents from Oroxylum indicum (L.) Kurz bark. J. Ethnopharmacol. 2017, 197, 138–146. [Google Scholar] [CrossRef]
- Tebruegge, M.; Curtis, N. Enterovirus infections in neonates. Semin. Fetal. Neonatal. Med. 2009, 14, 222–227. [Google Scholar] [CrossRef]
- Garmaroudi, F.S.; Marchant, D.; Hendry, R.; Luo, H.; Yang, D.; Ye, X.; Shi, J.; McManus, B.M. Coxsackievirus B3 replication and pathogenesis. Future Microbiol. 2015, 10, 629–653. [Google Scholar] [CrossRef]
- Kwon, B.E.; Song, J.H.; Song, H.H.; Kang, J.W.; Hwang, S.N.; Rhee, K.J.; Shim, A.; Hong, E.H.; Kim, Y.J.; Jeon, S.M.; et al. Antiviral activity of Oroxylin A against Coxsackievirus B3 alleviates virus-induced acute pancreatic damage in mice. PLoS ONE 2016, 11, e0155784. [Google Scholar] [CrossRef]
- Xian, Y.; Su, Y.; Liang, J.; Long, F.; Feng, X.; Xiao, Y.; Lian, H.; Xu, J.; Zhao, J.; Liu, Q.; et al. Oroxylin A reduces osteoclast formation and bone resorption via suppressing RANKL-induced ROS and NFATc1 activation. Biochem. Pharm. 2021, 193, 114761. [Google Scholar] [CrossRef]
- Ren, G.; Chen, H.; Zhang, M.; Yang, N.; Yang, H.; Xu, C.; Li, J.; Ning, C.; Song, Z.; Zhou, S.; et al. Pharmacokinetics, tissue distribution and excretion study of Oroxylin A, Oroxylin A 7-O-glucuronide and Oroxylin A sodium sulfonate in rats after administration of Oroxylin A. Fitoterapia 2020, 142, 104480. [Google Scholar] [CrossRef]
- Ren, G.; Chen, H.; Zhang, M.; Yang, N.; Yang, H.; Xu, C.; Li, J.; Ning, C.; Song, Z.; Zhou, S.; et al. Determination of oroxylin A, oroxylin A 7-O-glucuronide, and oroxylin A sodium sulfonate in beagle dogs by using UHPLC MS/MS Application in a pharmacokinetic study. J. Sep. Sci. 2020, 43, 2290–2300. [Google Scholar] [CrossRef]
- Fong, S.Y.; Wong, Y.C.; Zuo, Z. Development of a SPE-LC/MS/MS method for simultaneous quantification of baicalein, wogonin, oroxylin A and their glucuronides baicalin, wogonoside and oroxyloside in rats and its application to brain uptake and plasma pharmacokinetic studies. J. Pharm. Biomed. Anal. 2014, 97, 9–23. [Google Scholar] [CrossRef]
- Zhang, Q.; Cong, D.; An, D.; Fan, A.; Liu, Q.; Yi, Y.; Song, Z.; Chen, X.; Lu, Y.; Zhao, D.; et al. Determination of oroxylin A and oroxylin A 7-O-d-glucuronide in HepG2 cell lysate and subcellular fractions with SPE-UPLC-MS/MS: Cellular pharmacokinetic study to indicate anti-cancer mechanisms. J. Pharm. Biomed. Anal. 2018, 154, 364–372. [Google Scholar] [CrossRef]
- Mu, R.; Qi, Q.; Gu, H.; Wang, J.; Yang, Y.; Rong, J.; Liu, W.; Lu, N.; You, Q.; Guo, Q. Involvement of p53 in oroxylin A-induced apoptosis in cancer cells. Mol. Carcinog. 2009, 48, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Miao, H.; Zhang, Y.; Li, W.; Li, Z.; Zhou, Y.; Zhao, L.; Guo, Q. Bone marrow microenvironment confers imatinib resistance to chronic myelogenous leukemia and oroxylin A reverses the resistance by suppressing Stat3 pathway. Arch. Toxicol. 2015, 89, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Ma, R.; Huang, S.; Liao, Y.; Ding, Y.; Li, Z.; Guo, Q.; Tan, R.; Zhang, L.; Zhao, L. Oroxylin A increases the sensitivity of temozolomide on glioma cells by hypoxia-inducible factor 1α/hedgehog pathway under hypoxia. J. Cell. Physiol. 2019, 234, 17392–17404. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S.; et al. Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study. Arch. Intern. Med. 1995, 155, 381–386. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Ferri, C. Flavonoids: Antioxidants against atherosclerosis. Nutrients 2010, 2, 889–902. [Google Scholar] [CrossRef]
- Arai, Y.; Watanabe, S.; Kimira, M.; Shimoi, K.; Mochizuki, R.; Kinae, N. Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 2000, 130, 2243–2250. [Google Scholar] [CrossRef]
- Rana, V.; Parama, D.; Girisa, S.; Harsha, C.; Kunnumakkara, A.B. Oxidative Stress And Inflammation. In Antioxidants and Functional Foods for Neurodegenerative Disorders; CRC Press: Boca Raton, FL, USA, 2021; pp. 21–36. [Google Scholar]
- Yang, Y.; Hu, Y.; Gu, H.Y.; Lu, N.; Liu, W.; Qi, Q.; Zhao, L.; Wang, X.T.; You, Q.D.; Guo, Q.L. Oroxylin A induces G2/M phase cell-cycle arrest via inhibiting Cdk7-mediated expression of Cdc2/p34 in human gastric carcinoma BGC-823 cells. J. Pharm. Pharmacol. 2008, 60, 1459–1463. [Google Scholar] [CrossRef]
- Yao, J.; Hu, R.; Sun, J.; Lin, B.; Zhao, L.; Sha, Y.; Zhu, B.; You, Q.D.; Yan, T.; Guo, Q.L. Oroxylin A prevents inflammation-related tumor through down-regulation of inflammatory gene expression by inhibiting NF-kappaB signaling. Mol. Carcinog. 2014, 53, 145–158. [Google Scholar] [CrossRef]
- Ku, W.T.; Tung, J.J.; Lee, T.J.; Lai, K.C. Long-Term Exposure to Oroxylin A Inhibits Metastasis by Suppressing CCL2 in Oral Squamous Cell Carcinoma Cells. Cancers 2019, 11, 353. [Google Scholar] [CrossRef]
- Zhao, K.; Li, X.; Lin, B.; Yang, D.; Zhou, Y.; Li, Z.; Guo, Q.; Lu, N. Oroxyloside inhibits angiogenesis through suppressing internalization of VEGFR2/Flk-1 in endothelial cells. J. Cell Physiol. 2018, 233, 3454–3464. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, Y.; You, Q.D.; Liu, W.; Gu, H.Y.; Zhao, L.; Zhang, K.; Wang, W.; Wang, X.T.; Guo, Q.L. Oroxylin A induced apoptosis of human hepatocellular carcinoma cell line HepG2 was involved in its antitumor activity. Biochem. Biophys. Res. Commun. 2006, 351, 521–527. [Google Scholar] [CrossRef]
- Zhu, B.; Zhao, L.; Zhu, L.; Wang, H.; Sha, Y.; Yao, J.; Li, Z.; You, Q.; Guo, Q. Oroxylin A reverses CAM-DR of HepG2 cells by suppressing Integrinbeta1 and its related pathway. Toxicol. Appl. Pharmacol. 2012, 259, 387–394. [Google Scholar] [CrossRef]
- Xu, Z.F.; Sun, X.K.; Chen, G.; Han, C.; Wang, F.; Zhang, Y.D. Oroxyloside inhibits human glioma progression by suppressing proliferation, metastasis and inducing apoptosis related pathways. Biomed. Pharmacother. 2018, 97, 1564–1574. [Google Scholar] [CrossRef]
- Zhang, C.; Bian, M.; Chen, X.; Jin, H.; Zhao, S.; Yang, X.; Shao, J.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A prevents angiogenesis of LSECs in liver fibrosis via inhibition of YAP/HIF-1α signaling. J. Cell. Biochem. 2018, 119, 2258–2268. [Google Scholar] [CrossRef]
- Li, J.; Tong, D.; Liu, J.; Chen, F.; Shen, Y. Oroxylin A attenuates cigarette smoke-induced lung inflammation by activating Nrf2. Int. Immunopharmacol. 2016, 40, 524–529. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, H.; Li, W.; Yao, J.; Sun, Y.; Li, Z.; Zhao, L.; Guo, Q. CXCL12/CXCR4 axis confers adriamycin resistance to human chronic myelogenous leukemia and oroxylin A improves the sensitivity of K562/ADM cells. Biochem. Pharm. 2014, 90, 212–225. [Google Scholar] [CrossRef]
- Yang, H.Y.; Zhao, L.; Yang, Z.; Zhao, Q.; Qiang, L.; Ha, J.; Li, Z.Y.; You, Q.D.; Guo, Q.L. Oroxylin A reverses multi-drug resistance of human hepatoma BEL7402/5-FU cells via downregulation of P-glycoprotein expression by inhibiting NF-kappaB signaling pathway. Mol. Carcinog. 2012, 51, 185–195. [Google Scholar] [CrossRef]
- Li, C.; Zhang, L.; Lin, G.; Zuo, Z. Identification and quantification of baicalein, wogonin, oroxylin A and their major glucuronide conjugated metabolites in rat plasma after oral administration of Radix scutellariae product. J. Pharm. Biomed. Anal. 2011, 54, 750–758. [Google Scholar] [CrossRef]
- Cai, Y.; Li, S.; Li, T.; Zhou, R.; Wai, A.T.; Yan, R. Oral pharmacokinetics of baicalin, wogonoside, oroxylin A 7-O-β-d-glucuronide and their aglycones from an aqueous extract of Scutellariae Radix in the rat. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1026, 124–133. [Google Scholar] [CrossRef]
- Tukey, R.H.; Strassburg, C.P. Human UDP-glucuronosyltransferases: Metabolism, expression, and disease. Annu. Rev. Pharm. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef]
- Fu, W.; Wang, J.; Yu, L.; Zhao, L.; Lu, N.; You, Q.; Guo, Q.; Li, Z. Synthesis and biological evaluation of 7-O-modified oroxylin A derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 1118–1121. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).