
Efficient Viral Transduction in Fetal and Adult Human Inner Ear Explants with AAV9-PHP.B Vectors

 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. AAV9-PHP.B Vectors
2.2. Inner Ear Sample Collection, Dissection, and Viral Transduction
2.3. Immunostaining and Imaging
2.4. Image Acquisition and Processing
2.5. Quantification of eGFP Expression and Transduction Efficiency
3. Results
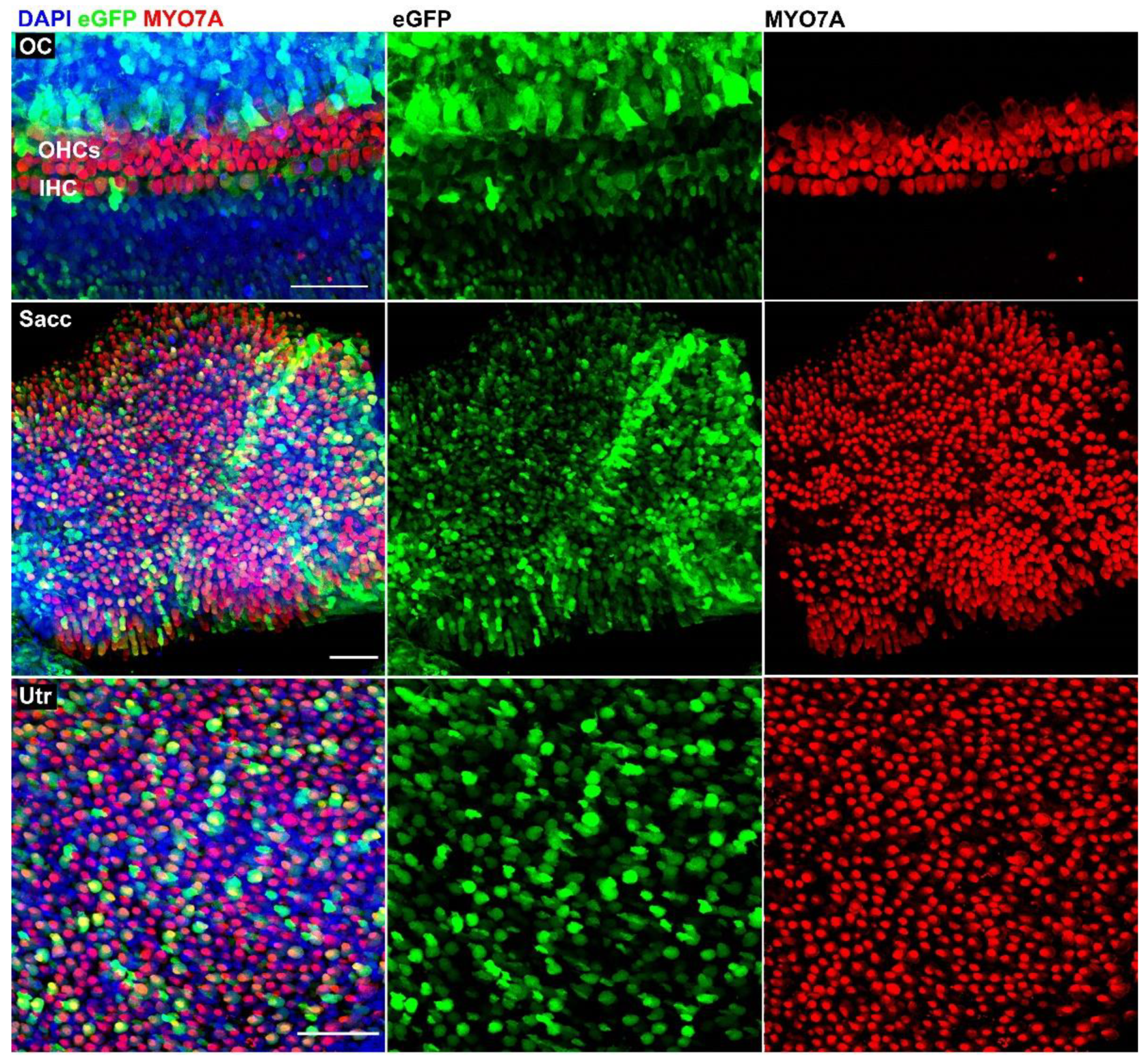
3.1. AAV9-PHP.B Vectors Transduce Cells in Adult human Vestibular Organs
3.2. AAV9-PHP.B Vectors Transduce Cells in the Fetal Human Inner Ear
3.3. Quantification of eGFP-Positive Hair Cells and Transduction Efficiency
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Neurology Collaborators. Global, regional, and national burden of neurological disorders, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef]
- Smith, R.J.; Bale, J.F., Jr.; White, K.R. Sensorineural hearing loss in children. Lancet 2005, 365, 879–890. [Google Scholar] [CrossRef]
- Kral, A. O’Donoghue GM. Profound deafness in childhood. N. Engl. J. Med. 2010, 363, 1438–1450. [Google Scholar] [CrossRef]
- Kesser, B.W.; Hashisaki, G.T.; Fletcher, K.; Eppard, H.; Holt, J.R. An in vitro model system to study gene therapy in the human inner ear. Gene Ther. 2007, 14, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Landegger, L.D.; Pan, B.; Askew, C.; Wassmer, S.J.; Gluck, S.D.; Galvin, A.; Taylor, R.; Forge, R.T.A.; Stankovic, K.M.; Holt, J.R. A synthetic AAV vector enables safe and efficient gene transfer to the mammalian inner ear. Nat. Biotechnol. 2017, 35, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.R.; Filia, A.; Paredes, U.; Asai, Y.; Holt, J.R.; Lovett, M.; Forge, A. Regenerating hair cells in vestibular sensory epithelia from humans. Elife 2018, 7, e34817. [Google Scholar] [CrossRef] [PubMed]
- Akil, O.; Seal, R.P.; Burke, K.; Wang, C.; Alemi, A.; During, M.; Edwards, R.H.; Lustig, L.R. Restoration of hearing in the VGLUT3 knockout mouse using virally mediated gene therapy. Neuron 2012, 75, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Askew, C.; Rochat, C.; Pan, B.; Asai, Y.; Ahmed, H.; Child, E.; Schneider, B.L.; Aebischer, P.; Holt, J.R. Tmc gene therapy restores auditory function in deaf mice. Sci. Transl. Med. 2015, 7, 295ra108. [Google Scholar] [CrossRef]
- Chien, W.W.; Isgrig, K.; Roy, S.; Belyantseva, I.A.; Drummond, M.C.; May, L.A.; Fitzgerald, T.S.; Friedman, T.B.; Cunningham, L. Gene Therapy Restores Hair Cell Stereocilia Morphology in Inner Ears of Deaf Whirler Mice. Mol. Ther. 2016, 24, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Emptoz, A.; Michel, V.; Lelli, A.; Akil, O.; de Monvel, J.B.; Lahlou, G.; Meyer, A.; Dupont, T.; Nouaille, S.; Ey, E.; et al. Local gene therapy durably restores vestibular function in a mouse model of Usher syndrome type 1G. Proc. Natl. Acad. Sci. USA 2017, 114, 9695–9700. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Askew, C.; Galvin, A.; Heman-Ackah, S.; Asai, Y.; Indzhykulian, A.; Jodelka, F.M.; Hastings, M.; Lentz, J.J.; Vandenberghe, L.H.; et al. Gene therapy restores auditory and vestibular function in a mouse model of Usher syndrome type 1c. Nat. Biotechnol. 2017, 35, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Dulon, D.; Papal, S.; Patni, P.; Cortese, M.; Vincent, M.; Tertrais, M.; Emptoz, A.; Tlili, A.; Bouleau, Y.; Michel, V.; et al. Clarin-1 gene transfer rescues auditory synaptopathy in model of Usher syndrome. J. Clin. Investig. 2018, 128, 3382–3401. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Meijer, E.J.; Ivanchenko, M.; Tenneson, K.; Emond, F.; Hanlon, K.S.; Indzhykulian, A.; Volak, A.; Karavitaki, K.D.; Tamvakologos, P.I.; et al. Gene Transfer with AAV9-PHP.B Rescues Hearing in a Mouse Model of Usher Syndrome 3A and Transduces Hair Cells in a Non-human Primate. Mol. Ther. Methods Clin. Dev. 2018, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Nist-Lund, C.; Solanes, P.; Goldberg, H.; Wu, J.; Pan, B.; Schneider, B.L.; Holt, J.R. Efficient viral transduction in mouse inner ear hair cells with utricle injection and AAV9-PHP.B. Hear. Res. 2020, 394, 107882. [Google Scholar] [CrossRef] [PubMed]
- Delmaghani, S.; El-Amraoui, A. Inner Ear Gene Therapies Take Off: Current Promises and Future Challenges. J. Clin. Med. 2020, 9, 2309. [Google Scholar] [CrossRef] [PubMed]
- Askew, C.; Chien, W.W. Adeno-associated virus gene replacement for recessive inner ear dysfunction: Progress and challenges. Hear. Res. 2020, 394, 107947. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Solanes, P.; Nist-Lund, C.; Spataro, S.; Shubina-Oleinik, O.; Marcovich, I.; Goldberg, H.; Schneider, B.L.; Holt, J.R. Single and Dual Vector Gene Therapy with AAV9-PHP.B Rescues Hearing in Tmc1 Mutant Mice. Mol. Ther. 2021, 29, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Hızlı, Ö.; Cureoglu, S.; Kaya, S.; Schachern, P.A.; Paparella, M.M.; Adams, M.E. Quantitative Vestibular Labyrinthine Otopathology in Temporal Bones with Vestibular Schwannoma. Otolaryngol. Head Neck Surg. 2016, 154, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Roosli, C.; Linthicum, F.H., Jr.; Cureoglu, S.; Merchant, S.N. Dysfunction of the cochlea contributing to hearing loss in acoustic neuromas: An underappreciated entity. Otol. Neurotol. 2012, 33, 473–480. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Beelen, E.S.A.; van der Valk, W.H.; Verhagen, T.O.; de Groot, J.C.M.J.; Madison, M.A.; Shadmanfar, W.; Hensen, E.F.; Jansen, J.C.; van Benthem, P.P.G.; Holt, J.R.; et al. Efficient Viral Transduction in Fetal and Adult Human Inner Ear Explants with AAV9-PHP.B Vectors. Biomolecules 2022, 12, 816. https://doi.org/10.3390/biom12060816
van Beelen ESA, van der Valk WH, Verhagen TO, de Groot JCMJ, Madison MA, Shadmanfar W, Hensen EF, Jansen JC, van Benthem PPG, Holt JR, et al. Efficient Viral Transduction in Fetal and Adult Human Inner Ear Explants with AAV9-PHP.B Vectors. Biomolecules. 2022; 12(6):816. https://doi.org/10.3390/biom12060816
Chicago/Turabian Stylevan Beelen, Edward S. A., Wouter H. van der Valk, Thijs O. Verhagen, John C. M. J. de Groot, Margot A. Madison, Wijs Shadmanfar, Erik F. Hensen, Jeroen C. Jansen, Peter Paul G. van Benthem, Jeffrey R. Holt, and et al. 2022. "Efficient Viral Transduction in Fetal and Adult Human Inner Ear Explants with AAV9-PHP.B Vectors" Biomolecules 12, no. 6: 816. https://doi.org/10.3390/biom12060816
APA Stylevan Beelen, E. S. A., van der Valk, W. H., Verhagen, T. O., de Groot, J. C. M. J., Madison, M. A., Shadmanfar, W., Hensen, E. F., Jansen, J. C., van Benthem, P. P. G., Holt, J. R., & Locher, H. (2022). Efficient Viral Transduction in Fetal and Adult Human Inner Ear Explants with AAV9-PHP.B Vectors. Biomolecules, 12(6), 816. https://doi.org/10.3390/biom12060816