
Structure of New Ferroverdins Recruiting Unconventional Ferrous Iron Chelating Agents


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Compound Identification
3. Results

{kind=link}
{kind=link}
{kind=link}
| # | Ferroverdin | Molecular Formula | m/z (Exp) | Δm(ppm) | Fe2+ Chelators | Reference |
|---|---|---|---|---|---|---|
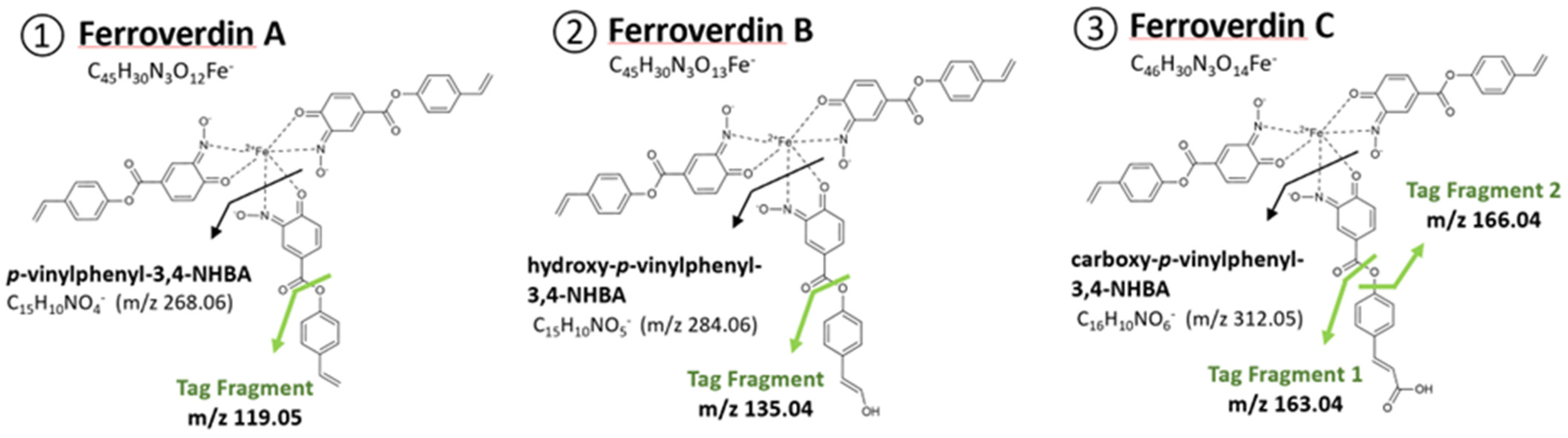
| 1 | A | C45H30N3O12Fe− | 860.1199 | 1.7 | AAA | [8,9] |
| 2 | B | C45H30N3O13Fe− | 876.1141 | 0.9 | AAB | [8,9,10] |
| 3 | C | C46H30N3O14Fe− | 904.1010 | 1.2 | AAC | [8,9,10] |
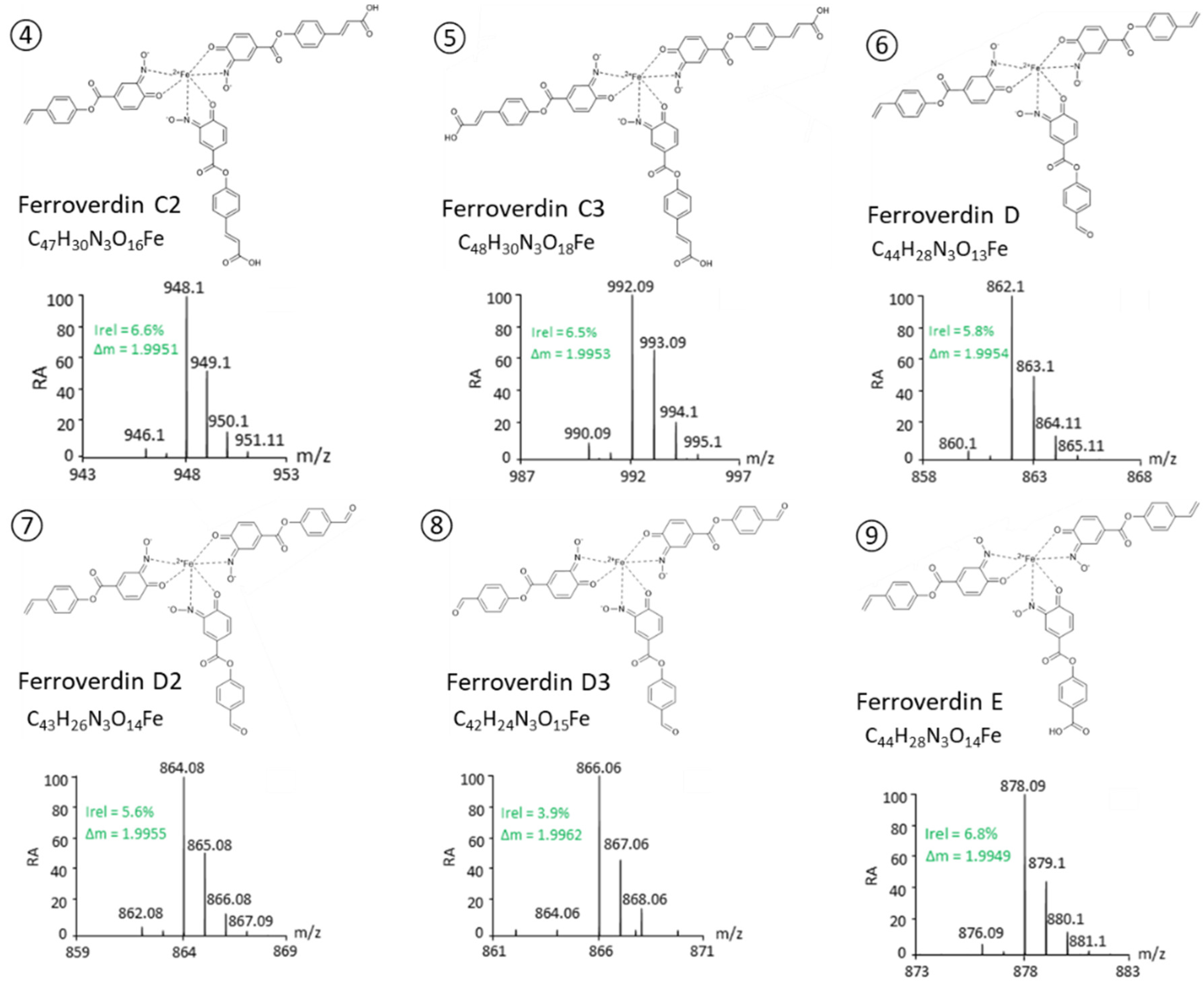
| 4 | C2 | C47H30N3O16Fe− | 948.0990 | 1 | ACC | This study |
| 5 | C3 | C48H30N3O18Fe− | 992.0868 | 1 | CCC | This study |
| 6 | D | C44H28N3O13Fe− | 862.0991 | 1.6 | AAD | This study |
| 7 | D2 | C43H26N3O14Fe− | 864.0784 | 1.7 | ADD | This study |
| 8 | D3 | C42H24N3O15Fe− | 866.0579 | 1.9 | DDD | This study |
| 9 | E | C44H28N3O14Fe− | 878.0933 | 0.7 | AAE | This study |
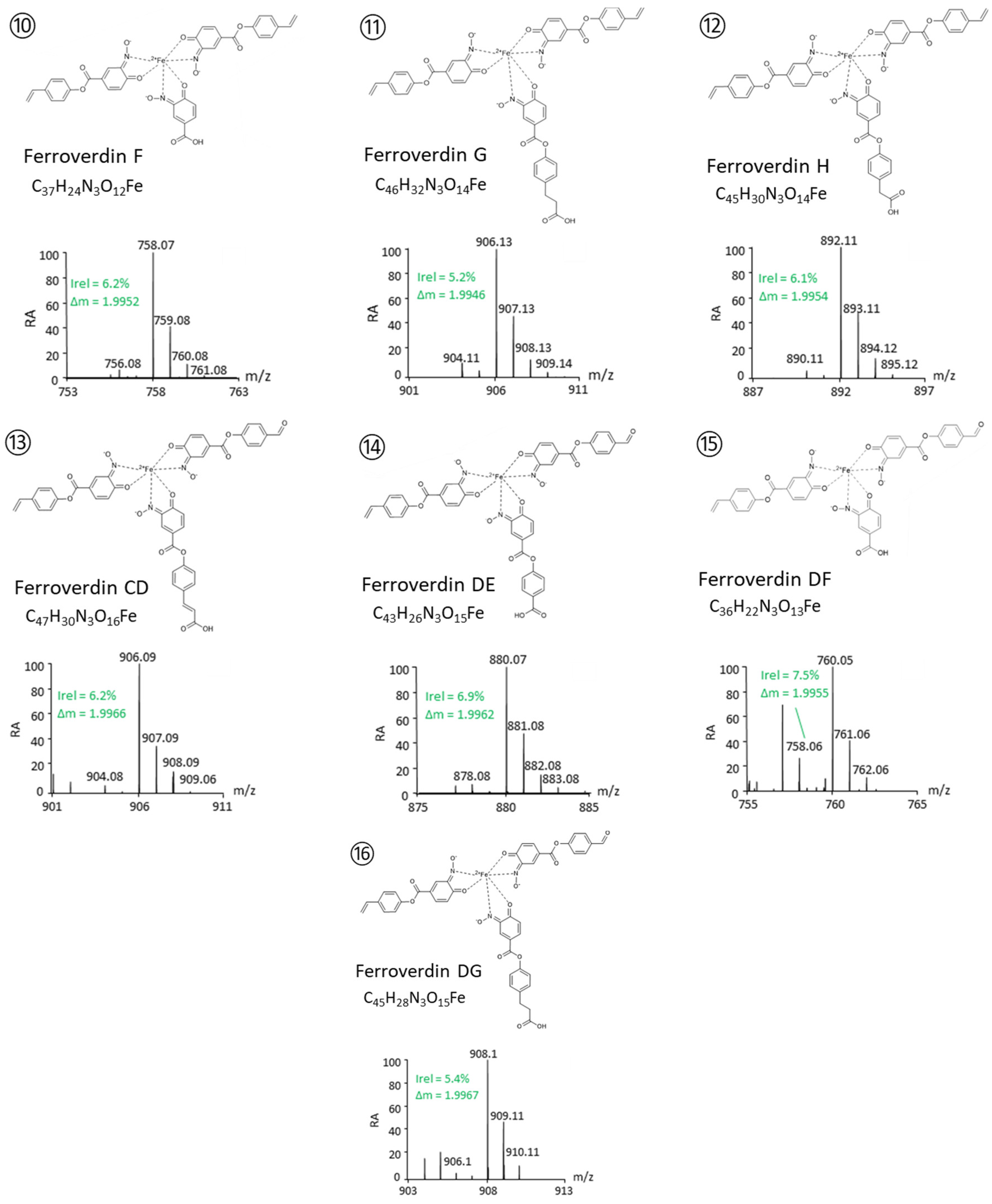
| 10 | F | C37H24N3O12Fe− | 758.0726 | 1.4 | AAF | This study |
| 11 | G | C46H32 N3O14Fe− | 906.1253 | 1.5 | AAG | This study |
| 12 | H | C44H30N3O14Fe− | 892.1101 | 2.1 | AAH | This study |
| 13 | CD | C45H28N3O15Fe− | 906.0888 | 1.3 | ACD | This study |
| 14 | DE | C43H26N3O15Fe− | 880.0732 | 1.5 | ADE | This study |
| 15 | DF | C36H22 N3O13Fe− | 760.0518 | 1.4 | ADF | This study |
| 16 | DG | C46H28N3O15Fe− | 908.1043 | 1.2 | ADG | This study |
| From compounds 17 to 46, MS/MS fragmentation did not allow us to identify the third chelating molecule | ||||||
| 17 | NA | C47H32N3O15Fe− | 934.1196 | 0.79 | AAX | This study |
| 18 | NA | C39H30N3O10Fe− | 756.1295 | 1.2 | AAX | This study |
| 19 | NA | C38H31N3O16Fe− | 841.1068 | 1 | AAX | This study |
| 20 | NA | C45H32N3O14Fe− | 894.1246 | 0.8 | AAX | This study |
| 21 | NA | C47H32N3O13Fe− | 902.1298 | 0.8 | AAX | This study |
| 22 | NA | C54H56N6O19Fe− | 1148.2958 | 0.5 | AAX | This study |
| 23 | NA | C48H34N3O15Fe− | 948,1361 | 1.3 | AAX | This study |
| 24 | NA | C38H21N3O17Fe− | 857.1021 | 1.5 | AAX | This study |
| 25 | NA | C40H25N4O13Fe− | 825.0794 | 2.5 | AAX | This study |
| 26 | NA | C34H31N3O16Fe− | 793.0000 | 1.5 | AAX | This study |
| 27 | NA | C35H33N3O16Fe− | 807.0000 | 1.9 | AAX | This study |
| 28 | NA | C33H26N3O10Fe− | 680.0979 | 0.9 | AAX | This study |
| 29 | NA | C45H30N3O15Fe− | 908.0593 | 4.3 | AAX | This study |
| 30 | NA | C36H32N3O10Fe− | 722.1453 | 1.5 | AAX | This study |
| 31 | NA | C40H32N4O16Fe− | 880.11792 | 1.3 | AAX | This study |
| 32 | NA | C32H27N3O16Fe− | 765.0757 | 1.5 | AAX | This study |
| 33 | NA | C46H30N3O15Fe− | 920.1043 | 1.2 | AAX | This study |
| 34 | NA | C47H34N3O14Fe− | 920.1412 | 1.2 | AAX | This study |
| For compounds 35 to 44, MS/MS fragmentation only identified p-vinylphenyl-3,4-NHBA as one of the three chelating molecules. | ||||||
| 35 | NA | C47H32N3O16Fe− | 950.1150 | 1.3 | AXX | This study |
| 36 | NA | C44H30N3O15Fe− | 896.1037 | 0.6 | AXX | This study |
| 37 | NA | C39H28N4O10Fe− | 768.1158 | 0.3 | AXX | This study |
| 38 | NA | C37H32N3O12Fe− | 766.1348 | 1 | AXX | This study |
| 39 | NA | C40H30N3O12Fe− | 800.1192 | 1 | AXX | This study |
| 40 | NA | C41H31N4O10Fe− | 795.1405 | 1.3 | AXX | This study |
| 41 | NA | C36H30N3O12Fe− | 752.1190 | 0.77 | AXX | This study |
| 42 | NA | C53H54N6O20Fe− | 1150.2754 | 0.2 | AXX | This study |
| 43 | NA | C43H26N3O15Fe− | 880.0732 | 1.5 | AXX | This study |
| 44 | NA | C40H30N5O16Fe− | 892.1035 | 0.1 | AXX | This study |
| 45 | NA | C35H30N3O10Fe− | 708.1296 | 1.4 | ? | This study |
| 46 | NA | C35H28N3O10Fe− | 706.1140 | 1.4 | ? | This study |
- A total of 5 novel ferroverdins (D(6), E(9), F(10), G(11), and H(12), Figure 2), as well as ferroverdin B(2) and ferroverdin C(3), also use 2 molecules of p-vinylphenyl-3,4-NHBA to bind to the ferrous iron, but the third molecule is an unconventional chelator: p-formylphenyl-3,4-NHBA for ferroverdin D(6), p-benzoic acid-3,4-NHBA for ferroverdin E(9), 3,4-NHBA for ferroverdin F(10), p-phenylpropionate-3,4-NHBA for ferroverdin G(11), and p-phenylacetate-3,4-NHBA(12) for ferroverdin H (see Figure 2 and Table 2).
- In 6 new ferroverdins, p-vinylphenyl-3,4-NHBA is only one of the 3 molecules used for ferrous iron binding: ferroverdins C2(4), D2(7), CD(13), DE(14), DF(15), and DG(16) (Figure 2). In the previously known ferroverdins, at least two molecules of p-vinylphenyl-3,4-NHBA were used for Fe2+ binding.
- Ferroverdins C3(5) and D3(8) (Figure 2) are remarkable as they are the first ferroverdins reported for which p-vinylphenyl-3,4-NHBA is never used for chelating Fe2+, but which are, instead, composed of 3 carboxy-p-vinylphenyl-3,4-NHBA, or 3 p-formylphenyl-3,4-NHBA, respectively.
- Remarkably, the 2 ferroverdins F(10) and DF(15) (Figure 2) recruit 3,4-NHBA for iron chelation; therefore, they are the only ferroverdins using a molecule not resulting from the activity of the FevW/BagE enzyme for the condensation of 3,4-NHBA with p-vinylphenol (see the proposed biosynthetic pathway for ferroverdin biosynthesis in [12]).
| Ferroverdins | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecule involved in Fe2+ chelation | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
| A | B | C | C2 | C3 | D | D2 | D3 | E | F | G | H | CD | DE | DF | DG | |
| p-vinylphenyl-3,4-NHBA | 3 | 2 | 2 | 1 | - | 2 | 1 | - | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
| Hydroxy-p-vinylphenyl-3,4-NHBA | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Carboxy-p-vinylphenyl-3,4-NHBA | - | - | 1 | 2 | 3 | - | - | - | - | - | - | - | 1 | - | - | - |
| p-formylphenyl-3,4-NHBA | - | - | - | - | - | 1 | 2 | 3 | - | - | - | - | 1 | 1 | 1 | 1 |
| p-benzoic acid-3,4-NHBA | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | - | - |
| 3,4-NHBA | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | - |
| p-phenylpropionate-3,4-NHBA | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 |
| p-phenyacetate-3,4-NHBA | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicholls, A.J.; Barber, T.; Baxendale, I.R. The Synthesis and Utility of Metal-Nitrosophenolato Compounds—Highlighting the Baudisch Reaction. Molecules 2019, 24, 4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chain, E.B.; Tonolo, A.; Carilli, A. Ferroverdin, a Green Pigment Containing Iron Produced by a Streptomycete. Nature 1955, 176, 645. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, M.; Pessi, I.S.; Arguelles-Arias, A.; Noirfalise, P.; Luis, G.; Ongena, M.; Barton, H.; Carnol, M.; Rigali, S. Streptomyces lunaelactis sp. Nov., a Novel Ferroverdin A-Producing Streptomyces Species Isolated from a Moonmilk Speleothem. Antonie Van Leeuwenhoek 2015, 107, 519–531. [Google Scholar] [CrossRef]
- Martinet, L.; Naômé, A.; Baiwir, D.; De Pauw, E.; Mazzucchelli, G.; Rigali, S. On the Risks of Phylogeny-Based Strain Prioritization for Drug Discovery: Streptomyces lunaelactis as a Case Study. Biomolecules 2020, 10, 1027. [Google Scholar] [CrossRef]
- Adam, D.; Maciejewska, M.; Naômé, A.; Martinet, L.; Coppieters, W.; Karim, L.; Baurain, D.; Rigali, S. Isolation, Characterization, and Antibacterial Activity of Hard-to-Culture Actinobacteria from Cave Moonmilk Deposits. Antibiotics 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska, M.; Adam, D.; Martinet, L.; Naômé, A.; Całusińska, M.; Delfosse, P.; Carnol, M.; Barton, H.A.; Hayette, M.-P.; Smargiasso, N.; et al. A Phenotypic and Genotypic Analysis of the Antimicrobial Potential of Cultivable Streptomyces Isolated from Cave Moonmilk Deposits. Front. Microbiol. 2016, 7, 1455. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska, M.; Całusińska, M.; Cornet, L.; Adam, D.; Pessi, I.S.; Malchair, S.; Delfosse, P.; Baurain, D.; Barton, H.A.; Carnol, M.; et al. High-Throughput Sequencing Analysis of the Actinobacterial Spatial Diversity in Moonmilk Deposits. Antibiotics 2018, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Candeloro, S.; Grdenic, D.; Taylor, N.; Thompson, B.; Viswamitra, M.; Hodgkin, D.C. Structure of Ferroverdin. Nature 1969, 224, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Bertholdt, H.; Chain, E.B.; Di Vittorio, V. Structure of Ferroverdin. Nature 1962, 194, 769–770. [Google Scholar] [CrossRef]
- Tomoda, H.; Tabata, N.; Shinose, M.; Takahashi, Y.; Woodruff, H.B.; Omura, S. Ferroverdins, Inhibitors of Cholesteryl Ester Transfer Protein Produced by Streptomyces sp. WK-5344. I. Production, Isolation and Biological Properties. J. Antibiot. 1999, 52, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.C.; Leong, J. Production of Deferriferrioxamines B and E from a Ferroverdin-Producing Streptomyces Species. J. Bacteriol. 1982, 149, 381–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinet, L.; Naômé, A.; Deflandre, B.; Maciejewska, M.; Tellatin, D.; Tenconi, E.; Smargiasso, N.; de Pauw, E.; van Wezel, G.P.; Rigali, S. A Single Biosynthetic Gene Cluster Is Responsible for the Production of Bagremycin Antibiotics and Ferroverdin Iron Chelators. mBio 2019, 10, e01230-19. [Google Scholar] [CrossRef] [Green Version]
- Traxler, M.F.; Seyedsayamdost, M.R.; Clardy, J.; Kolter, R. Interspecies Modulation of Bacterial Development through Iron Competition and Siderophore Piracy. Mol. Microbiol. 2012, 86, 628–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.; Traxler, M.F.; Craig, M.; Maciejewska, M.; Ongena, M.; van Wezel, G.P.; Kolter, R.; Rigali, S. Altered Desferrioxamine-Mediated Iron Utilization Is a Common Trait of Bald Mutants of Streptomyces coelicolor. Metallomics 2014, 6, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Oikawa, H.; Ogawa, H.; Hosono, K.; Shinmachi, F.; Takano, H.; Sakuda, S.; Beppu, T.; Ueda, K. Desferrioxamine E Produced by Streptomyces Griseus Stimulates Growth and Development of Streptomyces tanashiensis. Microbiology 2005, 151, 2899–2905. [Google Scholar] [CrossRef]
- Craig, M.; Lambert, S.; Jourdan, S.; Tenconi, E.; Colson, S.; Maciejewska, M.; Ongena, M.; Martin, J.F.; van Wezel, G.; Rigali, S. Unsuspected Control of Siderophore Production by N-Acetylglucosamine in Streptomycetes. Environ. Microbiol. Rep. 2012, 4, 512–521. [Google Scholar] [CrossRef]
- Tierrafría, V.H.; Ramos-Aboites, H.E.; Gosset, G.; Barona-Gómez, F. Disruption of the Siderophore-Binding DesE Receptor Gene in Streptomyces coelicolor A3(2) Results in Impaired Growth in Spite of Multiple Iron-Siderophore Transport Systems. Microb. Biotechnol. 2011, 4, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Traxler, M.F.; Watrous, J.D.; Alexandrov, T.; Dorrestein, P.C.; Kolter, R. Interspecies Interactions Stimulate Diversification of the Streptomyces coelicolor Secreted Metabolome. mBio 2013, 4, e00459-13. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Yang, R.; Lyu, M.; Wang, S.; Liu, X.; Wen, Y.; Song, Y.; Li, J.; Chen, Z. IdeR, a DtxR Family Iron Response Regulator, Controls Iron Homeostasis, Morphological Differentiation, Secondary Metabolism, and the Oxidative Stress Response in Streptomyces avermitilis. Appl. Environ. Microbiol. 2018, 84, e01503-18. [Google Scholar] [CrossRef] [Green Version]
- Tenconi, E.; Traxler, M.F.; Hoebreck, C.; van Wezel, G.P.; Rigali, S. Production of Prodiginines Is Part of a Programmed Cell Death Process in Streptomyces coelicolor. Front. Microbiol. 2018, 9, 1742. [Google Scholar] [CrossRef]
- Naômé, A.; Maciejewska, M.; Calusinska, M.; Martinet, L.; Anderssen, S.; Adam, D.; Tenconi, E.; Deflandre, B.; Coppieters, W.; Karim, L.; et al. Complete Genome Sequence of Streptomyces lunaelactis MM109T, Isolated from Cave Moonmilk Deposits. Genome Announc. 2018, 6, e00435-18. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhu, Y.; Hou, B.; Wu, H.; Zhang, H. Characterization of the Bagremycin Biosynthetic Gene Cluster in Streptomyces sp. Tü 4128. Biosci. Biotechnol. Biochem. 2019, 83, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Tabata, N.; Tomoda, H.; Omura, S. Ferroverdins, Inhibitors of Cholesteryl Ester Transfer Protein Produced by Streptomyces sp. WK-5344. II. Structure Elucidation. J. Antibiot. 1999, 52, 1108–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omura, S.; Tomoda, H.; Takahashi, Y. Substances Wk-5344a and Wk-5344b and Process for Producing the Same. U.S. Patent 6,512,008, 2 January 2003. [Google Scholar]
- Nicholls, S.J.; Bubb, K. The Mystery of Evacetrapib-Why Are CETP Inhibitors Failing? Expert Rev. Cardiovasc. Ther. 2020, 18, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Ference, B.A.; Kastelein, J.J.P.; Ginsberg, H.N.; Chapman, M.J.; Nicholls, S.J.; Ray, K.K.; Packard, C.J.; Laufs, U.; Brook, R.D.; Oliver-Williams, C.; et al. Association of Genetic Variants Related to CETP Inhibitors and Statins With Lipoprotein Levels and Cardiovascular Risk. JAMA 2017, 318, 947–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosmas, C.E.; DeJesus, E.; Rosario, D.; Vittorio, T.J. CETP Inhibition: Past Failures and Future Hopes. Clin. Med. Insights Cardiol. 2016, 10, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tall, A.R.; Rader, D.J. The Trials and Tribulations of CETP Inhibitors. Circ. Res. 2018, 122, 106–112. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Baars, O.; Morel, F.M.M.; Perlman, D.H. ChelomEx: Isotope-Assisted Discovery of Metal Chelates in Complex Media Using High-Resolution LC-MS. Anal. Chem. 2014, 86, 11298–11305. [Google Scholar] [CrossRef]
- Baars, O.; Zhang, X.; Morel, F.M.M.; Seyedsayamdost, M.R. The Siderophore Metabolome of Azotobacter vinelandii. Appl. Environ. Microbiol. 2016, 82, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Deicke, M.; Mohr, J.F.; Bellenger, J.-P.; Wichard, T. Metallophore Mapping in Complex Matrices by Metal Isotope Coded Profiling of Organic Ligands. Analyst 2014, 139, 6096–6099. [Google Scholar] [CrossRef] [Green Version]
- Lehner, S.M.; Atanasova, L.; Neumann, N.K.N.; Krska, R.; Lemmens, M.; Druzhinina, I.S.; Schuhmacher, R. Isotope-Assisted Screening for Iron-Containing Metabolites Reveals a High Degree of Diversity among Known and Unknown Siderophores Produced by Trichoderma spp. Appl. Environ. Microbiol. 2013, 79, 18–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdali, H.; Lebrihi, A.; Monje, M.C.; Benharref, A.; Hafidi, M.; Ouhdouch, Y.; Virolle, M.J. A Molecule of the Viridomycin Family Originating from a Streptomyces griseus-Related Strain Has the Ability to Solubilize Rock Phosphate and to Inhibit Microbial Growth. Antibiotics 2021, 10, 72. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinet, L.; Baiwir, D.; Mazzucchelli, G.; Rigali, S. Structure of New Ferroverdins Recruiting Unconventional Ferrous Iron Chelating Agents. Biomolecules 2022, 12, 752. https://doi.org/10.3390/biom12060752
Martinet L, Baiwir D, Mazzucchelli G, Rigali S. Structure of New Ferroverdins Recruiting Unconventional Ferrous Iron Chelating Agents. Biomolecules. 2022; 12(6):752. https://doi.org/10.3390/biom12060752
Chicago/Turabian StyleMartinet, Loïc, Dominique Baiwir, Gabriel Mazzucchelli, and Sébastien Rigali. 2022. "Structure of New Ferroverdins Recruiting Unconventional Ferrous Iron Chelating Agents" Biomolecules 12, no. 6: 752. https://doi.org/10.3390/biom12060752
APA StyleMartinet, L., Baiwir, D., Mazzucchelli, G., & Rigali, S. (2022). Structure of New Ferroverdins Recruiting Unconventional Ferrous Iron Chelating Agents. Biomolecules, 12(6), 752. https://doi.org/10.3390/biom12060752