
Effects of Palmitoylethanolamide on Neurodegenerative Diseases: A Review from Rodents to Humans


Abstract
:1. Introduction
1.1. PEA, an Anti-Inflammatory and Neuroprotective Substance
1.2. PEA Action in the Presence of Aging and Neurodegeneration
2. Mild Cognitive Impairment (MCI)
3. Alzheimer’s Disease (AD)
4. Parkinson’s Disease (PD)
5. Huntington’s Disease (HD)
6. Frontotemporal Dementia (FTD)
7. Amyotrophic Lateral Sclerosis (ALS)
8. Multiple Sclerosis (MS)
9. Other Diseases (Vascular Dementia, Myasthenia Gravis)
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peritore, A.F.; Siracusa, R.; Crupi, R.; Cuzzocrea, S. Therapeutic Efficacy of Palmitoylethanolamide and Its New Formulations in Synergy with Different Antioxidant Molecules Present in Diets. Nutrients 2019, 11, 2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiurchiù, V.; Leuti, A.; Smoum, R.; Mechoulam, R.; Maccarrone, M. Bioactive lipids ALIAmides differentially modulate inflammatory responses of distinct subsets of primary human T lymphocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 5716–5723. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.A.; Tsuboi, K.; Uyama, T.; Ueda, N. New players in the fatty acyl ethanolamide metabolism. Pharmacol. Res. 2014, 86, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.S.; Barrett, D.A.; Randall, M.D. ‘Entourage’ effects of N-palmitoylethanolamide and N-oleoylethanolamide on vasorelaxation to anandamide occur through TRPV1 receptors. Br. J. Pharmacol. 2008, 155, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Musella, A.; Fresegna, D.; Rizzo, F.R.; Gentile, A.; Bullitta, S.; De Vito, F.; Guadalupi, L.; Centonze, D.; Mandolesi, G. A novel crosstalk within the endocannabinoid system controls GABA transmission in the striatum. Sci. Rep. 2017, 7, 7363. [Google Scholar] [CrossRef] [Green Version]
- Costa, B.; Conti, S.; Giagnoni, G.; Colleoni, M. Therapeutic effect of the endogenous fatty acid amide, palmitoylethanolamide, in rat acute inflammation: Inhibition of nitric oxide and cyclo–oxygenase systems. Br. J. Pharmacol. 2002, 137, 413–420. [Google Scholar] [CrossRef]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [Green Version]
- Keppel Hesselink, J.M.; de Boer, T.; Witkamp, R.F. Palmitoylethanolamide: A Natural Body–Own Anti–Inflammatory Agent, Effective and Safe against Influenza and Common Cold. Int. J. Inflam. 2013, 2013, 151028. [Google Scholar] [CrossRef] [Green Version]
- Nestmann, E.R. Safety of micronized palmitoylethanolamide (microPEA): Lack of toxicity and genotoxic potential. Food Sci. Nutr. 2016, 5, 292–309. [Google Scholar] [CrossRef]
- Gabrielsson, L.; Mattsson, S.; Fowler, C.J. Palmitoylethanolamide for the treatment of pain: Pharmacokinetics, safety and efficacy. Br. J. Clin. Pharmacol. 2016, 82, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Petrosino, S.; Di Marzo, V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Schiano Moriello, A. Palmitoylethanolamide: A Nutritional Approach to Keep Neuroinflammation within Physiological Boundaries–A Systematic Review. Int. J. Mol. Sci. 2020, 21, 9526. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Cristino, L.; Karsak, M.; Gaffal, E.; Ueda, N.; Tüting, T.; Bisogno, T.; De Filippis, D.; D’Amico, A.; Saturnino, C.; et al. Protective role of palmitoylethanolamide in contact allergic dermatitis. Allergy 2010, 65, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and related N-acylethanolamines: Biological activities and metabolism. Inflamm. Regener. 2018, 38, 28. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. Palmitoylethanolamide is a new possible pharmacological treatment for the inflammation associated with trauma. Mini Rev. Med. Chem. 2013, 13, 237–255. [Google Scholar]
- Mattace Raso, G.; Russo, R.; Calignano, A.; Meli, R. Palmitoylethanolamide in CNS health and disease. Pharmacol. Res. 2014, 86, 32–41. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Zusso, M.; Giusti, P. An Inflammation–Centric View of Neurological Disease: Beyond the Neuron. Front. Cell. Neurosci. 2018, 12, 72. [Google Scholar] [CrossRef]
- Solorzano, C.; Zhu, C.; Battista, N.; Astarita, G.; Lodola, A.; Rivara, S.; Mor, M.; Russo, R.; Maccarrone, M.; Antonietti, F.; et al. Selective N-acylethanolamine-hydrolyzing acid amidase inhibition reveals a key role for endogenous palmitoylethanolamide in inflammation. Proc. Natl. Acad. Sci. USA 2009, 106, 20966–20971. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.; Ikematsu, N.; Uyama, T.; Deutsch, D.G.; Tokumura, A.; Ueda, N. Biosynthetic pathways of bioactive N–acylethanolamines in brain. CNS Neurol. Disord. 2013, 12, 7–16. [Google Scholar] [CrossRef]
- Rankin, L.; Fowler, C.J. The Basal Pharmacology of Palmitoylethanolamide. Int. J. Mol. Sci. 2020, 21, 7942. [Google Scholar] [CrossRef]
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A proposed autacoid mechanism controlling mastocyte behaviour. Agents Actions 1993, 39, C145–C147. [Google Scholar] [CrossRef] [PubMed]
- Paladini, A.; Varrassi, G.; Bentivegna, G.; Carletti, S.; Piroli, A.; Coaccioli, S. Palmitoylethanolamide in the Treatment of Failed Back Surgery Syndrome. Pain Res. Treat. 2017, 2017, 1486010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roviezzo, F.; Rossi, A.; Caiazzo, E.; Orlando, P.; Riemma, M.A.; Iacono, V.M.; Guarino, A.; Ialenti, A.; Cicala, C.; Peritore, A.; et al. Palmitoylethanolamide Supplementation during Sensitization Prevents Airway Allergic Symptoms in the Mouse. Front. Pharmacol. 2017, 8, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, S.; Costa, B.; Colleoni, M.; Parolaro, D.; Giagnoni, G. Antiinflammatory action of endocannabinoid palmitoylethanolamide and the synthetic cannabinoid nabilone in a model of acute inflammation in the rat. Br. J. Pharmacol. 2002, 135, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saturnino, C.; Popolo, A.; Ramunno, A.; Adesso, S.; Pecoraro, M.; Plutino, M.R.; Rizzato, S.; Albinati, A.; Marzocco, S.; Sala, M.; et al. Anti-Inflammatory, Antioxidant and Crystallographic Studies of N-Palmitoyl-ethanol Amine (PEA) Derivatives. Molecules 2017, 22, 616. [Google Scholar] [CrossRef] [Green Version]
- Warden, A.; Truitt, J.; Merriman, M.; Ponomareva, O.; Jameson, K.; Ferguson, L.B.; Mayfield, R.D.; Harris, R.A. Localization of PPAR isotypes in the adult mouse and human brain. Sci. Rep. 2016, 6, 27618. [Google Scholar] [CrossRef]
- Shi, Q.X.; Yang, L.K.; Shi, W.L.; Wang, L.; Zhou, S.M.; Guan, S.Y.; Zhao, M.G.; Yang, Q. The novel cannabinoid receptor GPR55 mediates anxiolytic-like effects in the medial orbital cortex of mice with acute stress. Mol. Brain 2017, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. Int. J. Mol. Sci. 2020, 21, 5330. [Google Scholar] [CrossRef]
- Beggiato, S.; Tomasini, M.C.; Ferraro, L. Palmitoylethanolamide (PEA) as a Potential Therapeutic Agent in Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 821. [Google Scholar] [CrossRef] [Green Version]
- Ardizzone, A.; Fusco, R.; Casili, G.; Lanza, M.; Impellizzeri, D.; Esposito, E.; Cuzzocrea, S. Effect of Ultra-Micronized-Palmitoylethanolamide and Acetyl-l-Carnitine on Experimental Model of Inflammatory Pain. Int. J. Mol. Sci. 2021, 22, 1967. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Micronized/ultramicronized palmitoylethanolamide displays superior oral efficacy compared to nonmicronized palmitoylethanolamide in a rat model of inflammatory pain. J. Neuroinflammation 2014, 11, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noce, A.; Albanese, M.; Marrone, G.; Di Lauro, M.; Pietroboni Zaitseva, A.; Palazzetti, D.; Guerriero, C.; Paolino, A.; Pizzenti, G.; Di Daniele, F.; et al. Ultramicronized Palmitoylethanolamide (um-PEA): A New Possible Adjuvant Treatment in COVID-19 patients. Pharmaceuticals 2021, 14, 336. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Cuzzocrea, S.; Crupi, R. An Update of Palmitoylethanolamide and Luteolin Effects in Preclinical and Clinical Studies of Neuroinflammatory Events. Antioxidants 2020, 9, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashaari, Z.; Hadjzadeh, M.A.; Hassanzadeh, G.; Alizamir, T.; Yousefi, B.; Keshavarzi, Z.; Mokhtari, T. The flavone luteolin improves central nervous system disorders by different mechanisms: A review. J. Mol. Neurosci. 2018, 65, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Adami, R.; Liparoti, S.; Di Capua, A.; Scogliamiglio, M.; Reverchon, E. Production of PEA composite microparticles with polyvinylpyrrolidone and luteolin using supercritical assisted atomization. J. Supercrit. Fluids. 2019, 143, 82–89. [Google Scholar] [CrossRef]
- Power, R.; Prado-Cabrero, A.; Mulcahy, R.; Howard, A.; Nolan, J.M. The Role of Nutrition for the Aging Population: Implications for Cognition and Alzheimer’s Disease. Annu. Rev. Food Sci. Technol. 2019, 10, 619–639. [Google Scholar] [CrossRef] [Green Version]
- Vauzour, D.; Camprubi-Robles, M.; Miquel-Kergoat, S.; Andres-Lacueva, C.; Bánáti, D.; Barberger-Gateau, P.; Bowman, G.L.; Caberlotto, L.; Clarke, R.; Hogervorst, E.; et al. Nutrition for the ageing brain: Towards evidence for an optimal diet. Ageing Res. Rev. 2017, 35, 222–240. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Azam, S.; Haque, M.E.; Balakrishnan, R.; Kim, I.S.; Choi, D.K. The Ageing Brain: Molecular and Cellular Basis of Neurodegeneration. Front. Cell. Dev. Biol. 2021, 9, 683459. [Google Scholar] [CrossRef]
- Skaper, S.D.; Barbierato, M.; Facci, L.; Borri, M.; Contarini, G.; Zusso, M.; Giusti, P. Co–Ultramicronized Palmitoylethanolamide/Luteolin Facilitates the Development of Differentiating and Undifferentiated Rat Oligodendrocyte Progenitor Cells. Mol. Neurobiol. 2018, 55, 103–114. [Google Scholar] [CrossRef]
- Jakel, S.; Dimou, L. Glial Cells and Their Function in the Adult Brain: A Journey through the History of Their Ablation. Front. Cell. Neurosci. 2017, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Hussain, R.; Zubair, H.; Pursell, S.; Shahab, M. Neurodegenerative Diseases: Regenerative Mechanisms and Novel Therapeutic Approaches. Brain Sci. 2018, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.K.; Grossberg, G.T. Diagnosis and treatment of Alzheimer’s disease. Neurology 2005, 64, S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Schapira, A.H.V. Non-motor symptoms of Parkinson’s disease: Dopaminergic pathophysiology and treatment. Lancet Neurol. 2009, 8, 464–474. [Google Scholar] [CrossRef]
- Rees, K.; Stowe, R.; Patel, S.; Ives, N.; Breen, K.; Clarke, C.E.; Ben-Shlomo, Y. Non-steroidal anti-inflammatory drugs as disease-modifying agents for Parkinson’s disease: Evidence from observational studies. Cochrane Database Syst Rev. 2011, Cd008454. [Google Scholar] [CrossRef] [PubMed]
- Calabrò, R.S.; Naro, A.; De Luca, R.; Leonardi, S.; Russo, M.; Marra, A.; Bramanti, P. PEALut efficacy in mild cognitive impairment: Evidence from a SPECT case study! Aging Clin. Exp. Res. 2016, 28, 1279–1282. [Google Scholar] [CrossRef]
- Brotini, S.; Schievano, C.; Guidi, L. Ultra–micronized Palmitoylethanolamide: An Efficacious Adjuvant Therapy for Parkinson’s Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 705–713. [Google Scholar] [CrossRef]
- Brotini, S. Palmitoylethanolamide/Luteolin as Adjuvant Therapy to Improve an Unusual Case of Camptocormia in a Patient with Parkinson’s Disease: A Case Report. Innov. Clin. Neurosci. 2021, 18, 12–14. [Google Scholar]
- Assogna, M.; Casula, E.P.; Borghi, I.; Bonnì, S.; Samà, D.; Motta, C.; Di Lorenzo, F.; D’Acunto, A.; Porrazzini, F.; Minei, M.; et al. Effects of Palmitoylethanolamide Combined with Luteoline on Frontal Lobe Functions, High Frequency Oscillations, and GABAergic Transmission in Patients with Frontotemporal Dementia. J. Alzheimers Dis. 2020, 76, 1297–1308. [Google Scholar] [CrossRef]
- Clemente, S. Amyotrophic lateral sclerosis treatment with ultramicronized palmitoylethanolamide: A case report. CNS Neurol. Disord. Drug Targets 2012, 11, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Reyes–Ruiz, J.M.; Lopergolo, D.; Roseti, C.; Bertollini, C.; Ruffolo, G.; Cifelli, P.; Onesti, E.; Limatola, C.; Miledi, R.; et al. Acetylcholine receptors from human muscle as pharmacological targets for ALS therapy. Proc. Natl. Acad. Sci. USA. 2016, 113, 3060–3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean-Gilles, L.; Feng, S.; Tench, C.R.; Chapman, V.; Kendall, D.A.; Barrett, D.A.; Constantinescu, C.S. Plasma endocannabinoid levels in multiple sclerosis. J. Neurol. Sci. 2009, 287, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Kopsky, D.J.; Hesselink, J.M. Multimodal stepped care approach with acupuncture and PPAR–α agonist palmitoylethanolamide in the treatment of a patient with multiple sclerosis and central neuropathic pain. Acupunct. Med. 2012, 30, 53–55. [Google Scholar] [CrossRef]
- Orefice, N.S.; Alhouayek, M.; Carotenuto, A.; Montella, S.; Barbato, F.; Comelli, A.; Calignano, A.; Muccioli, G.G.; Orefice, G. Oral Palmitoylethanolamide Treatment Is Associated with Reduced Cutaneous Adverse Effects of Interferon–β1a and Circulating Proinflammatory Cytokines in Relapsing–Remitting Multiple Sclerosis. Neurotherapeutics 2016, 13, 428–438. [Google Scholar] [CrossRef]
- Onesti, E.; Frasca, V.; Ceccanti, M.; Tartaglia, G.; Gori, M.C.; Cambieri, C.; Libonati, L.; Palma, E.; Inghilleri, M. Short–Term Ultramicronized Palmitoylethanolamide Therapy in Patients with Myasthenia Gravis: A Pilot Study to Possible Future Implications of Treatment. CNS Neurol. Disord. Drug Targets 2019, 18, 232–238. [Google Scholar] [CrossRef]
- Scuderi, C.; Stecca, C.; Valenza, M.; Ratano, P.; Bronzuoli, M.R.; Bartoli, S.; Steardo, L.; Pompili, E.; Fumagalli, L.; Campolongo, P.; et al. Palmitoylethanolamide controls reactive gliosis and exerts neuroprotective functions in a rat model of Alzheimer’s disease. Cell Death Dis. 2014, 5, e1419. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, G.; Russo, R.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Palmitoylethanolamide protects against the amyloid–β25–35–induced learning and memory impairment in mice, an experimental model of Alzheimer disease. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2012, 37, 1784–1792. [Google Scholar] [CrossRef] [Green Version]
- Bronzuoli, M.R.; Facchinetti, R.; Steardo, L., Jr.; Romano, A.; Stecca, C.; Passarella, S.; Steardo, L.; Cassano, T.; Scuderi, C. Palmitoylethanolamide Dampens Reactive Astrogliosis and Improves Neuronal Trophic Support in a Triple Transgenic Model of Alzheimer’s Disease: In vitro and In vivo Evidence. Oxid. Med. Cell. Longev. 2018, 2018, 4720532. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Bronzuoli, M.R.; Facchinetti, R.; Pace, L.; Ferraro, L.; Broad, K.D.; Serviddio, G.; Bellanti, F.; Palombelli, G.; Carpinelli, G.; et al. Ultramicronized palmitoylethanolamide rescues learning and memory impairments in a triple transgenic mouse model of Alzheimer’s disease by exerting anti–inflammatory and neuroprotective effects. Transl. Psychiatry 2018, 8, 32. [Google Scholar] [CrossRef]
- Beggiato, S.; Tomasini, M.C.; Cassano, T.; Ferraro, L. Chronic Oral Palmitoylethanolamide Administration Rescues Cognitive Deficit and Reduces Neuroinflammation, Oxidative Stress, and Glutamate Levels in A Transgenic Murine Model of Alzheimer’s Disease. J. Clin. Med. 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, E.; Impellizzeri, D.; Mazzon, E.; Paterniti, I.; Cuzzocrea, S. Neuroprotective activity of palmitoylethanolamide in an animal model of Parkinson’s disease. PLoS ONE 2012, 7, e41880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crupi, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Casili, G.; Evangelista, M.; Cuzzocrea, S. N–palmitoylethanolamide prevents parkinsonian phenotypes in aged mice. Mol. Neurobiol. 2018, 55, 8455–8472. [Google Scholar] [CrossRef] [PubMed]
- Avagliano, C.; Russo, R.; De Caro, C.; Cristiano, C.; La Rana, G.; Piegari, G.; Paciello, O.; Citraro, R.; Russo, E.; De Sarro, G.; et al. Palmitoylethanolamide protects mice against 6–OHDA–induced neurotoxicity and endoplasmic reticulum stress: In vivo and in vitro evidence. Pharmacol. Res. 2016, 113 Pt A, 276–289. [Google Scholar] [CrossRef]
- Siracusa, R.; Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Crupi, R.; Navarra, M.; Cuzzocrea, S.; Esposito, E. The Association of Palmitoylethanolamide with Luteolin Decreases Neuroinflammation and Stimulates Autophagy in Parkinson’s Disease Model. CNS Neurol. Disord. Drug Targets 2015, 14, 1350–1365. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Di Paola, R.; Cuzzocrea, S. 2–Pentadecyl–2–Oxazoline Reduces Neuroinflammatory Environment in the MPTP Model of Parkinson Disease. Mol. Neurobiol. 2018, 55, 9251–9266. [Google Scholar] [CrossRef]
- Bisogno, T.; Martire, A.; Petrosino, S.; Popoli, P.; Di Marzo, V. Symptom-related changes of endocannabinoid and palmitoylethanolamide levels in brain areas of R6/2 mice, a transgenic model of Huntington’s disease. Neurochem. Int. 2008, 52, 307–313. [Google Scholar] [CrossRef]
- Baker, D.; Pryce, G.; Croxford, J.L.; Brown, P.; Pertwee, R.G.; Makriyannis, A.; Khanolkar, A.; Layward, L.; Fezza, F.; Bisogno, T.; et al. Endocannabinoids control spasticity in a multiple sclerosis model. FASEB J. 2001, 15, 300–302. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, A.; Faizi, M.; Talebi, F.; Noorbakhsh, F.; Kahrizi, F.; Naderi, N. Interaction between the protective effects of cannabidiol and palmitoylethanolamide in experimental model of multiple sclerosis in C57BL/6 mice. Neurosci. 2015, 290, 279–287. [Google Scholar] [CrossRef]
- Contarini, G.; Franceschini, D.; Facci, L.; Barbierato, M.; Giusti, P.; Zusso, M. A co–ultramicronized palmitoylethanolamide/luteolin composite mitigates clinical score and disease–relevant molecular markers in a mouse model of experimental autoimmune encephalomyelitis. J. Neuroinflammation 2019, 16, 126. [Google Scholar] [CrossRef] [Green Version]
- Loría, F.; Petrosino, S.; Mestre, L.; Spagnolo, A.; Correa, F.; Hernangómez, M.; Guaza, C.; Di Marzo, V.; Docagne, F. Study of the regulation of the endocannabinoid system in a virus model of multiple sclerosis reveals a therapeutic effect of palmitoylethanolamide. Eur. J. Neurosci. 2008, 28, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Crupi, R.; Peritore, A.F.; Gugliandolo, E.; D’Amico, R.; Petrosino, S.; Evangelista, M.; Di Paola, R.; et al. N–Palmitoylethanolamine–oxazoline (PEA–OXA): A new therapeutic strategy to reduce neuroinflammation, oxidative stress associated to vascular dementia in an experimental model of repeated bilateral common carotid arteries occlusion. Neurobiol. Dis. 2019, 125, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; Reisberg, B.; Zaudig, M.; Petersen, R.C.; Ritchie, K.; Broich, K.; Belleville, S.; Brodaty, H.; Bennett, D.; Chertkow, H.; et al. International Psychogeriatric Association Expert Conference on mild cognitive impairment. Mild cognitive impairment. Lancet 2006, 367, 1262–1270. [Google Scholar] [CrossRef]
- Ellison, J.M.; Harper, D.G.; Berlow, Y.; Zeranski, L. Beyond the “C” in MCI: Noncognitive symptoms in amnestic and non–amnestic mild cognitive impairment. CNS Spectr. 2008, 13, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C.; Caracciolo, B.; Brayne, C.; Gauthier, S.; Jelic, V.; Fratiglioni, L. Mild cognitive impairment: A concept in evolution. Int. J. Med. 2014, 275, 214–228. [Google Scholar] [CrossRef]
- Lombardi, G.; Crescioli, G.; Cavedo, E.; Lucenteforte, E.; Casazza, G.; Bellatorre, A.G.; Lista, C.; Costantino, G.; Frisoni, G.; Virgili, G.; et al. Structural magnetic resonance imaging for the early diagnosis of dementia due to Alzheimer’s disease in people with mild cognitive impairment. Cochrane Database Syst. Rev. 2020, 3, CD009628. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological alterations in Alzheimer disease. Cold Spring Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Andrews, K.A.; Modat, M.; Macdonald, K.E.; Yeatman, T.; Cardoso, M.J.; Leung, K.K.; Barnes, J.; Villemagne, V.L.; Rowe, C.C.; Fox, N.C.; et al. Acceleration of hippocampal atrophy rates in asymptomatic amyloidosis. Neurobiol. Aging 2016, 39, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the cholinergic system. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Esposito, G.; Blasio, A.; Valenza, M.; Arietti, P.; Steardo, L., Jr.; Carnuccio, R.; De Filippis, D.; Petrosino, S.; Iuvone, T.; et al. Palmitoylethanolamide counteracts reactive astrogliosis induced by β–amyloid peptide. J. Cell. Mol. Med. 2011, 15, 2664–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scuderi, C.; Valenza, M.; Stecca, C.; Esposito, G.; Carratù, M.R.; Steardo, L. Palmitoylethanolamide exerts neuroprotective effects in mixed neuroglial cultures and organotypic hippocampal slices via peroxisome proliferator–activated receptor–α. J. Neuroinflammation 2012, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scuderi, C.; Steardo, L. Neuroglial roots of neurodegenerative diseases: Therapeutic potential of palmitoylethanolamide in models of Alzheimer’s disease. CNS Neurol. Disord. Drug Targets 2013, 12, 62–69. [Google Scholar] [CrossRef]
- Beggiato, S.; Borelli, A.C.; Ferraro, L.; Tanganelli, S.; Antonelli, T.; Tomasini, M.C. Palmitoylethanolamide blunts amyloid-β42-induced astrocyte activation and improves neuronal survival in primary mouse cortical astrocyte–neuron co-cultures. J. Alzheimers Dis. 2018, 61, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Tomasini, M.C.; Borelli, A.C.; Beggiato, S.; Ferraro, L.; Cassano, T.; Tanganelli, S.; Antonelli, T. Differential Effects of Palmitoylethanolamide against Amyloid–β Induced Toxicity in Cortical Neuronal and Astrocytic Primary Cultures from Wild–Type and 3xTg–AD Mice. J. Alzheimer’s Dis. 2015, 46, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, S.; Cassano, T.; Ferraro, L.; Tomasini, M.C. Astrocytic palmitoylethanolamide pre–exposure exerts neuroprotective effects in astrocyte–neuron co–cultures from a triple transgenic mouse model of Alzheimer’s disease. Life Sci. 2020, 257, 118037. [Google Scholar] [CrossRef]
- Paterniti, I.; Cordaro, M.; Campolo, M.; Siracusa, R.; Cornelius, C.; Navarra, M.; Cuzzocrea, S.; Esposito, E. Neuroprotection by association of palmitoylethanolamide with luteolin in experimental Alzheimer’s disease models: The control of neuroinflammation. CNS Neurol. Disord. Drug Targets 2014, 13, 1530–1541. [Google Scholar] [CrossRef]
- Surguchov, A. Biomarkers in Parkinson’s Disease. In Neurodegenerative Diseases Biomarkers; Peplow, P.V., Martinez, B., Gennarelli, T.A., Eds.; Humana: New York, NY, USA, 2022; Volume 173, pp. 155–180. [Google Scholar]
- Alexander, G.E. Biology of Parkinson’s disease: Pathogenesis and pathophysiology of a multisystem neurodegenerative disorder. Dialogues Clin. Neurosci. 2004, 6, 259–280. [Google Scholar] [CrossRef]
- DeMaagd, G.; Philip, A. Parkinson’s Disease and Its Management: Part 1: Disease Entity, Risk Factors, Pathophysiology, Clinical Presentation, and Diagnosis. Pharm. Ther. 2015, 40, 504–532. [Google Scholar]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef]
- Michel, P.P.; Hirsch, E.C.; Hunot, S. Understanding Dopaminergic Cell Death Pathways in Parkinson Disease. Neuron 2016, 90, 675–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirdefeldt, K.; Adami, H.O.; Cole, P.; Trichopoulos, D.; Mandel, J. Epidemiology and etiology of Parkinson’s disease: A review of the evidence. Eur. J. Epidemiol. 2011, 26 (Suppl. 1), S1–S58. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef] [PubMed]
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenner, P.; Marsden, C.D. The actions of 1–methyl–4–phenyl–1,2,3,6–tetrahydropyridine in animals as a model of Parkinson’s disease. J. Neural Transm. Suppl. 1986, 20, 11–39. [Google Scholar] [PubMed]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef]
- Brinkman, R.R.; Mezei, M.M.; Theilmann, J.; Almqvist, E.; Hayden, M.R. The likelihood of being affected with Huntington disease by a particular age, for a specific CAG size. Am. J. Hum. Genet. 1997, 60, 1202–1210. [Google Scholar]
- Gatto, E.M.; Rojas, N.G.; Persi, G.; Etcheverry, J.L.; Cesarini, M.E.; Perandones, C. Huntington disease: Advances in the understanding of its mechanisms. Clin. Park. Relat. Disord. 2020, 3, 100056. [Google Scholar] [CrossRef]
- Glass, M. The role of cannabinoids in neurodegenerative diseases. Prog. Neuropsychopharmacol. Biol. Psychiatry 2001, 25, 743–765. [Google Scholar] [CrossRef]
- Finger, E.C. Frontotemporal Dementias. Continuum 2016, 22, 464–489. [Google Scholar] [CrossRef] [Green Version]
- Young, J.J.; Lavakumar, M.; Tampi, D.; Balachandran, S.; Tampi, R.R. Frontotemporal dementia: Latest evidence and clinical implications. Ther. Adv. Psychopharmacol. 2018, 8, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Zarei, S.; Carr, K.; Reiley, L.; Diaz, K.; Guerra, O.; Altamirano, P.F.; Pagani, W.; Lodin, D.; Orozco, G.; Chinea, A. A comprehensive review of amyotrophic lateral sclerosis. Surg. Neurol. Int. 2015, 6, 171. [Google Scholar] [CrossRef]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; van den Berg, L.H. Amyotrophic lateral sclerosis. Nat. Rev. Dis. Primers 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed]
- van Es, M.A.; Hardiman, O.; Chio, A.; Al-Chalabi, A.; Pasterkamp, R.J.; Veldink, J.H.; van den Berg, L.H. Amyotrophic lateral sclerosis. Lancet 2017, 390, 2084–2098. [Google Scholar] [CrossRef]
- Chang, J.L.; Lomen-Hoerth, C.; Murphy, J.; Henry, R.G.; Kramer, J.H.; Miller, B.L.; Gorno-Tempini, M.L. A voxel-based morphometry study of patterns of brain atrophy in ALS and ALS/FTLD. Neurology 2005, 65, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Correale, J.; Marrodan, M.; Ysrraelit, M.C. Mechanisms of Neurodegeneration and Axonal Dysfunction in Progressive Multiple Sclerosis. Biomedicines 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Rahn, K.; Slusher, B.; Kaplin, A. Cognitive impairment in multiple sclerosis: A forgotten disability remembered. Cerebrum 2012, 2012, 14. [Google Scholar]
- Miljković, D.; Spasojević, I. Multiple sclerosis: Molecular mechanisms and therapeutic opportunities. Antioxid. Redox Signal. 2013, 19, 2286–2334. [Google Scholar] [CrossRef]
- Peschl, P.; Bradl, M.; Höftberger, R.; Berger, T.; Reindl, M. Myelin Oligodendrocyte Glycoprotein: Deciphering a Target in Inflammatory Demyelinating Diseases. Front. Immunol. 2017, 8, 529. [Google Scholar] [CrossRef]
- Chang, A.; Tourtellotte, W.W.; Rudick, R.; Trapp, B.D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N. Engl. J. Med. 2002, 346, 165–173. [Google Scholar] [CrossRef]
- Barbierato, M.; Facci, L.; Marinelli, C.; Zusso, M.; Argentini, C.; Skaper, S.D.; Giusti, P. Co-ultramicronized Palmitoylethanolamide/Luteolin Promotes the Maturation of Oligodendrocyte Precursor Cells. Sci. Rep. 2015, 5, 16676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facci, L.; Barbierato, M.; Fusco, M.; Giusti, P.; Zusso, M. Co-Ultramicronized Palmitoylethanolamide/Luteolin-Induced Oligodendrocyte Precursor Cell Differentiation is Associated with Tyro3 Receptor Upregulation. Front. Pharmacol. 2021, 12, 698133. [Google Scholar] [CrossRef] [PubMed]
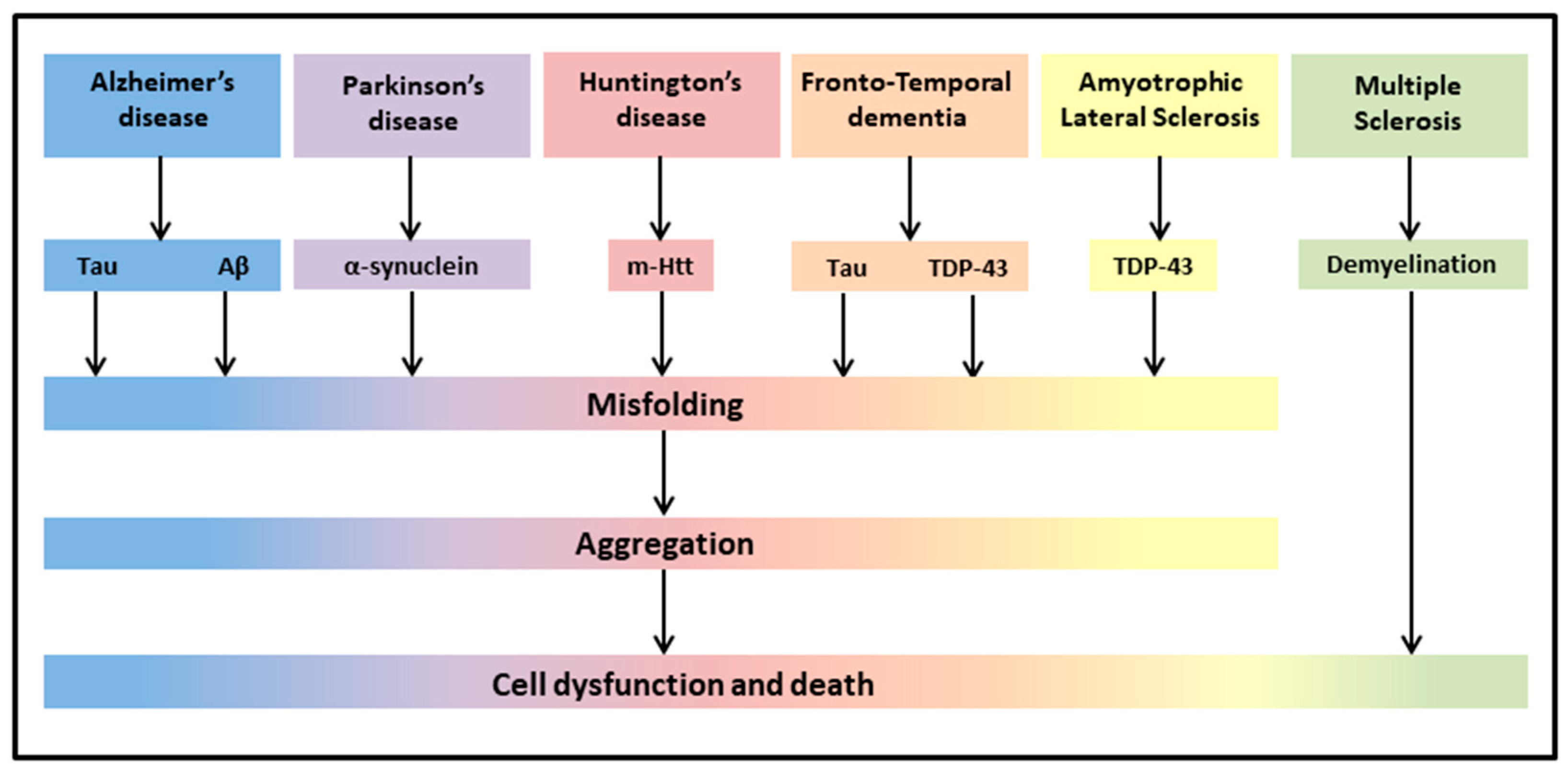

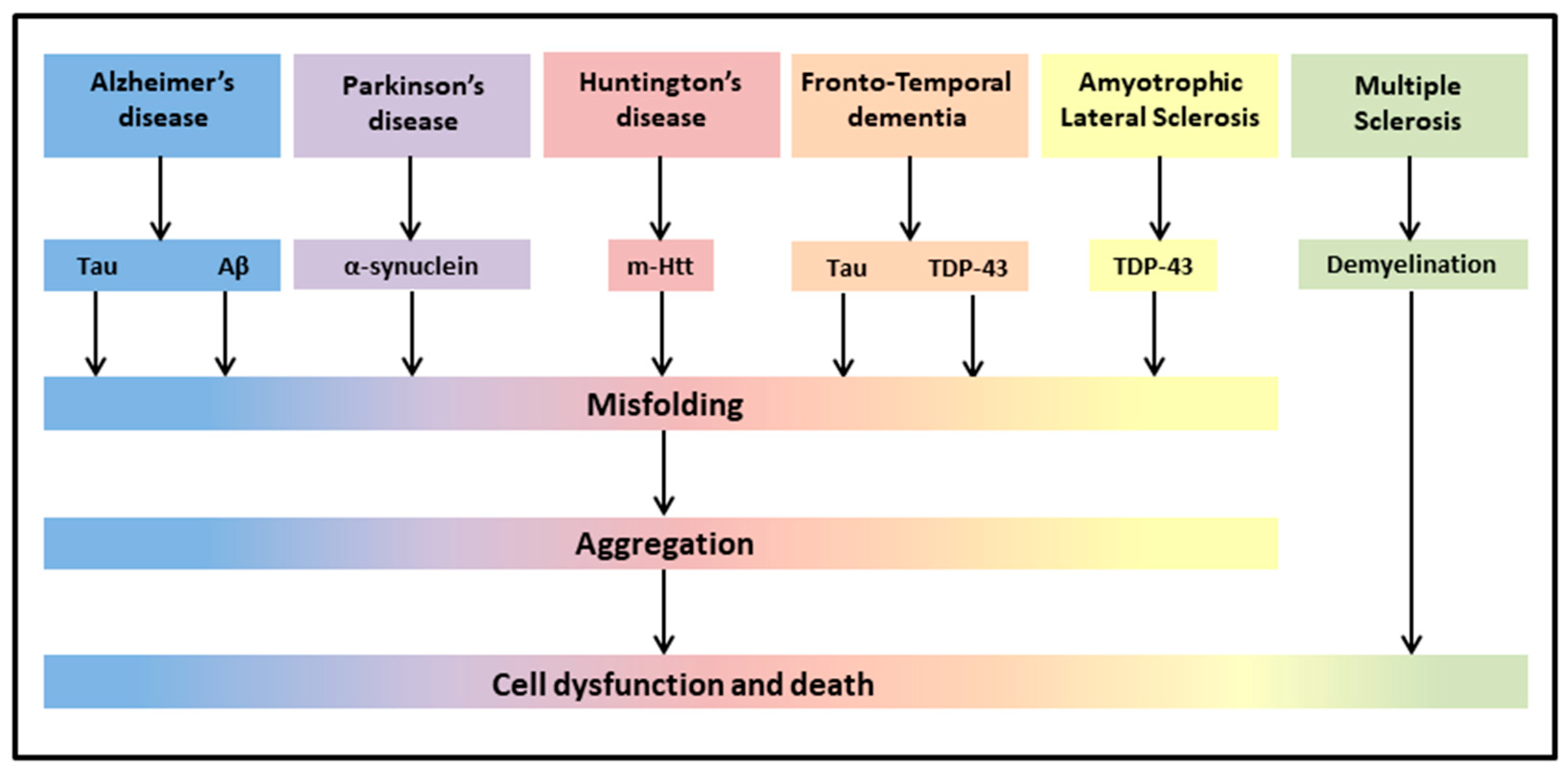{kind=link}
| Study | Disease | Sample | um PEA (Alone or In Combination) | Dosage | Duration | Main Outcomes of PEA Treatment |
|---|---|---|---|---|---|---|
| [47] | MCI | 1 patient | co-um-PEALut | 700/70 mg daily | T3: 3 months treatment T9: 9 months follow-up | T3: mild (though not significant) cognitive improvement; T9: near-normal neuropsychological assessment; improvement in test scores; brain SPECT near-normal. |
| [48] | PD | 30 patients | PEA added to regular levodopa | 600 mg daily | 12 months | Progressive reduction in the total MDS-UPDRS score; reduction in most nonmotor and motor symptoms. |
| [49] | PD | 1 patient | co-um-PEALut added to regular carbidopa/levodopa | 700/70 mg daily | 4 months | Complete resolution of leg and trunk dyskinesia and marked reduction in the onset of camptocormia during the “off” state. |
| [50] | FTD | 17 patients | co-um-PEALut | 700 mg/2 daily | 4 weeks | Improvement in test scores and neurophysiological evaluation; increase in TMS-evoked frontal lobe activity and of high-frequency oscillations in the beta/gamma range. |
| [51] | ALS | 1 patient | PEA | 600 mg/2 daily | ∼40 days | Improvement in clinical picture. |
| [52] | ALS | 28 treated and 36 untreated patients | PEA + 50 mg riluzole or 50 mg riluzole only | 600 mg/2 daily | 6 months | Lower decrease in forced vital capacity over time as compared with untreated ALS patients. |
| [53] | MS | 24 patients 17 healthy controls | eCBs levels in blood | _ | _ | eCB system is altered in MS. |
| [54] | MS | 1 patient | PEA | 600 mg/2 daily | ∼9 months | Pain reduction; increased interval between acupuncture sessions. |
| [55] | MS | 29 patients | PEA added to IFN-β1a or placebo | 600 mg daily | 12 months | Improvement in pain sensation, no reduction of erythema at the injection site, improved evaluation of quality of life, increase in PEA, AEA and OEA plasma levels, reduction of interferon-γ, tumor necrosis factor-α, and interleukin-17 serum profile. |
| [56] | Myasthenia gravis | 22 patients | PEA | 600 mg/2 daily | 1 week | Reduced level of disability and decremental muscle response. |
| Study | Disease | Sample | um PEA (Alone or In Combination) | Dosage | Duration | Main Outcomes of PEA Treatment |
|---|---|---|---|---|---|---|
| [57] | AD model (Aβ 1–42 intra-hippocampal injection) | Male adult Sprague-Dawley rats (9–12/group) | i.p. PEA PEA added to GW6471 | PEA:10 mg/kg; GW647: 2 mg/kg | 7 days | Restoration of Aβ 1–42-induced alterations; reduced mnestic deficits. |
| [58] | AD model (Aβ 25–35 i.c.v. injection) | Male PPAR-α/(B6.129S4-SvJaePparatm 1Gonz) and WT mice (9–10/group) | s.c. PEA and GW7647 | PEA: 3–30 mg/kg daily, GW7647: 5 mg/kg daily | 1–2 weeks or a single dose | Reduction (10 mg/kg) or prevention (30 mg/kg) of behavioral impairments. No rescue of memory deficits. PEA acute treatment was ineffective. |
| [59] | AD model | 3-month-old male 3 × Tg-AD and WT mice (9–10/group) | s.c. PEA or vehicle | 10 mg/kg daily | 90 days | Counteraction of disease progression, improvement of trophic support to neurons, in the absence of astrocytes and neuronal toxicity. |
| [60] | AD model | 3-month-old or 9-month-old male 3 × Tg-AD or WT mice (7–11/group) | s.c. PEA or vehicle | 10 mg/kg daily | 90 days | Improvement of learning and memory, amelioration of depressive and anhedonia-like symptoms, reduced Aβ formation, tau protein phosphorylation, promotion of hippocampal neuronal survival and astrocytic function, rebalancing of glutamatergic transmission, restraint of neuroinflammation. |
| [61] | AD model | 2-month-old male 3 × Tg-AD or WT mice (7–11/group) | oral PEA or vehicle | single dose/sub-chronic/chronic:100 mg/kg daily | 1–8–90 days | Rescue of cognitive deficit, restraint of neuroinflammation and oxidative stress, reduced increase in hippocampal glutamate levels. |
| [62] | PD model (MPTP) | 6–7-week-old male PPAR-αKO PPAR-αWT mice (10/group) | i.p. PEA | 10 mg/kg | 8 days | Reduction of MPTP-induced microglial activation, glial fibrillary acidic protein positive expression astrocyte numbers, overexpression of S100b; protection against alterations in microtubule-associated protein 2a,b, dopamine transporter, nNOS-positive cells in the substantia nigra. Reversal of motor deficits. |
| [63] | PD model (MPTP) | 3/21-month-old male CD1 mice (10/group) | oral PEA | 10 mg/kg | 60 days | Amelioration of behavioral deficits and of reduction of tyrosine hydroxylase and dopamine transporter in substantia nigra. Reduction of hippocampal proinflammatory cytokines and pro-neurogenic effects. |
| [64] | PD model (6-OHDA) | Ten-week-old male Swiss CD1 mice (6 × group) | s.c. PEA or GW7647 | PEA 3–30 mg/kg/day; GW7647 5 mg/kg/day | 28 days | Improvement of behavioral impairment. Increased tyrosine hydroxylase expression at striatal level. Reduction in the expression of pro-inflammatory enzymes, protective scavenging effect. |
| [65] | PD model (MPTP) | 8-week-old male C57BL/6 (10/group) | i.p. co-um-PEALut | 1 mg/kg daily | 8 days | Reduction of motor impairment, cataleptic response, immobility and anxiety levels. Reduction of neuronal degeneration and of specific PD markers, attenuation of inflammatory processes (activation of astrocytes, pro-inflammatory cytokines, and nitric oxide synthase), stimulation of autophagy. |
| [66] | PD model (MPTP) | 8-week-old male C57BL/6 (10/group) | oral PEA-OXA or vehicle | 10 mg/kg daily | 8 days | Prevention of MPTP-induced bradykinesia and anxiety, and neuronal degeneration of the dopaminergic tract, prevention of dopamine depletion, modulation of microglia and astrocyte activation. |
| [67] | HD model | ∼32-day-old-R6/2 10-week-old R6/2 mice and WT mice (4/group) | Measurement of PEA, AEA and 2-AG endogenous levels | _ | _ | Alteration of the eCB system, decreased levels of PEA in the striatum |
| [68] | MS model (EAE) | 12-week-old female C57BL/6 (8/group) | i.p. PEA or CBD or in combination | PEA 5 mg/kg CBD 5 mg/kg | 3 days | Reduced severity of EAE neurobehavioral scores, diminished inflammation, demyelination, axonal damage and inflammatory cytokine expression. |
| [69] | MS model (chronic relapsing EAE) | Biozzi ADH mice (>6/group) | i.v. or i.p. PEA | 1–10 mg/kg | Single injection | Amelioration of spasticity |
| [70] | MS model (EAE) | C57BL/6 mice (8/group) | i.p. co-um-PEALut or vehicle | 0.1, 1, and 5 mg/kg | 16 days | Dose-dependent improvement of clinical signs through anti-inflammatory signals and pro-resolving circuits. |
| [71] | MS model (TMEV-IDD) | Four-week female SJL/J mice | i.p. PEA or vehicle | 5 mg/kg | 10 days | Reduction of motor disability, anti-inflammatory effect. |
| [72] | Vascular dementia | CD1 mice | Oral PEA-OXA or vehicle | 10 mg/kg daily | 15 days | Improvement of behavioral deficits, reduction of histological alterations, decrease of markers of astrocyte and microglia activation and oxidative stress, modulation of antioxidant response, inhibition of apoptotic process. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landolfo, E.; Cutuli, D.; Petrosini, L.; Caltagirone, C. Effects of Palmitoylethanolamide on Neurodegenerative Diseases: A Review from Rodents to Humans. Biomolecules 2022, 12, 667. https://doi.org/10.3390/biom12050667
Landolfo E, Cutuli D, Petrosini L, Caltagirone C. Effects of Palmitoylethanolamide on Neurodegenerative Diseases: A Review from Rodents to Humans. Biomolecules. 2022; 12(5):667. https://doi.org/10.3390/biom12050667
Chicago/Turabian StyleLandolfo, Eugenia, Debora Cutuli, Laura Petrosini, and Carlo Caltagirone. 2022. "Effects of Palmitoylethanolamide on Neurodegenerative Diseases: A Review from Rodents to Humans" Biomolecules 12, no. 5: 667. https://doi.org/10.3390/biom12050667
APA StyleLandolfo, E., Cutuli, D., Petrosini, L., & Caltagirone, C. (2022). Effects of Palmitoylethanolamide on Neurodegenerative Diseases: A Review from Rodents to Humans. Biomolecules, 12(5), 667. https://doi.org/10.3390/biom12050667