
Lyso-IP: Uncovering Pathogenic Mechanisms of Lysosomal Dysfunction



Abstract
:1. Introduction
2. Traditional Methods for Lysosome Isolation
3. The Development of Lyso-IP and Similar Lysosomal Isolation Techniques
4. Novel Insights into the Pathogenic Mechanisms of Niemann-Pick Type C Disease Revealed by the Lyso-IP Approach
5. The Lyso-IP Approach Yields Insights into Other Lysosomal Functions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luzio, J.P.; Pryor, P.R.; Bright, N.A. Lysosomes: Fusion and function. Nat. Rev. Mol. Cell Biol. 2007, 8, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.M.; Zoncu, R. The lysosome as a regulatory hub. Annu. Rev. Cell Dev. Biol. 2016, 32, 223–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonam, S.R.; Wang, F.; Muller, S. Lysosomes as a therapeutic target. Nat. Rev. Drug Discov. 2019, 18, 923–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballabio, A.; Bonifacino, J.S. Lysosomes as dynamic regulators of cell and organismal homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 101–118. [Google Scholar] [CrossRef]
- Saftig, P.; Klumperman, J. Lysosome biogenesis and lysosomal membrane proteins: Trafficking meets function. Nat. Rev. Mol. Cell Biol. 2009, 10, 623–635. [Google Scholar] [CrossRef]
- Eskelinen, E.-L.; Tanaka, Y.; Saftig, P. At the acidic edge: Emerging functions for lysosomal membrane proteins. Trends Cell Biol. 2003, 13, 137–145. [Google Scholar] [CrossRef]
- Platt, F.M.; D’Azzo, A.; Davidson, B.L.; Neufeld, E.F.; Tifft, C.J. Lysosomal storage diseases. Nat. Rev. Dis. Primers 2018, 4, 27. [Google Scholar] [CrossRef]
- Sun, A. Lysosomal storage disease overview. Ann. Transl. Med. 2018, 6, 476. [Google Scholar] [CrossRef]
- Parenti, G.; Medina, D.L.; Ballabio, A. The rapidly evolving view of lysosomal storage diseases. EMBO Mol. Med. 2021, 13, e12836. [Google Scholar] [CrossRef]
- Tharkeshwar, A.K.; Gevaert, K.; Annaert, W. Organellar omics—A reviving strategy to untangle the biomolecular complexity of the cell. Proteomics 2018, 18, e1700113. [Google Scholar] [CrossRef]
- De Duve, C.; Pressman, B.C.; Gianetto, R.; Wattieaux, R.; Appelmans, F. Tissue fractionation studies. 6. Intracellular distribution patterns of enzymes in rat-liver tissue. Biochem. J. 1955, 60, 604–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udayar, V.; Chen, Y.; Sidransky, E.; Jagasia, R. Lysosomal dysfunction in neurodegeneration: Emerging concepts and methods. Trends Neurosci. 2022, 45, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, Z.P.; Bretou, M.; Annaert, W. Endo-lysosomal dysregulations and late-onset Alzheimer’s disease: Impact of genetic risk factors. Mol. Neurodegener. 2019, 14, 20. [Google Scholar] [CrossRef] [Green Version]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef]
- Graham, J.M. Purification of a crude mitochondrial fraction by density-gradient centrifugation. Curr Protoc Cell Biol 2001, 4, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Leighton, F.; Poole, B.; Beaufay, H.; Baudhuin, P.; Coffey, J.W.; Fowler, S.; De Duve, C. The large-scale separation of peroxisomes, mitochondria, and lysosomes from the livers of rats injected with triton wr-1339. Improved isolation procedures, automated analysis, biochemical and morphological properties of fractions. J. Cell Biol. 1968, 37, 482–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagshaw, R.D.; Mahuran, D.J.; Callahan, J.W. A proteomic analysis of lysosomal integral membrane proteins reveals the diverse composition of the organelle. Mol. Cell Proteom. 2005, 4, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado, C.; Perez-Jimenez, E.; Lahuerta, M.; Knecht, E. Isolation of lysosomes from mammalian tissues and cultured cells. Methods Mol. Biol. 2016, 1449, 299–311. [Google Scholar] [CrossRef]
- Gauthier, D.J.; Sobota, J.A.; Ferraro, F.; Mains, R.E.; Lazure, C. Flow cytometry-assisted purification and proteomic analysis of the corticotropes dense-core secretory granules. Proteomics 2008, 8, 3848–3861. [Google Scholar] [CrossRef] [Green Version]
- Tharkeshwar, A.K.; Demedts, D.; Annaert, W. Superparamagnetic nanoparticles for lysosome isolation to identify spatial alterations in lysosomal protein and lipid composition. STAR Protoc. 2020, 1, 100122. [Google Scholar] [CrossRef]
- Nylandsted, J.; Becker, A.C.; Bunkenborg, J.; Andersen, J.S.; Dengjel, J.; Jaattela, M. ErbB2-associated changes in the lysosomal proteome. Proteomics 2011, 11, 2830–2838. [Google Scholar] [CrossRef] [PubMed]
- Castellano, B.M.; Thelen, A.M.; Moldavski, O.; Feltes, M.; Van Der Welle, R.E.; Mydock-McGrane, L.; Jiang, X.; Van Eijkeren, R.J.; Davis, O.B.; Louie, S.M.; et al. Lysosomal cholesterol activates mTORC1 via an SLC38A9-Niemann-Pick C1 signaling complex. Science 2017, 355, 1306–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, O.B.; Shin, H.R.; Lim, C.Y.; Wu, E.Y.; Kukurugya, M.; Maher, C.F.; Perera, R.M.; Ordonez, M.P.; Zoncu, R. NPC1-mTORC1 signaling couples cholesterol sensing to organelle homeostasis and is a targetable pathway in Niemann-Pick type C. Dev. Cell 2021, 56, 260–276. [Google Scholar] [CrossRef] [PubMed]
- Wyant, G.A.; Abu-Remaileh, M.; Wolfson, R.L.; Chen, W.W.; Freinkman, E.; Danai, L.V.; Vander Heiden, M.G.; Sabatini, D.M. MTORC1 activator SLC38A9 is required to efflux essential amino acids from lysosomes and use protein as a nutrient. Cell 2017, 171, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.; Bras, J.; Deas, E.; O’Sullivan, S.S.; Parkkinen, L.; Lachmann, R.H.; Li, A.; Holton, J.; Guerreiro, R.; Paudel, R.; et al. Glucocerebrosidase mutations in clinical and pathologically proven Parkinson’s disease. Brain 2009, 132, 1783–1794. [Google Scholar] [CrossRef]
- Wyant, G.A.; Abu-Remaileh, M.; Frenkel, E.M.; Laqtom, N.N.; Dharamdasani, V.; Lewis, C.A.; Chan, S.H.; Heinze, I.; Ori, A.; Sabatini, D.M. NUFIP1 is a ribosome receptor for starvation-induced ribophagy. Science 2018, 360, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Abu-Remaileh, M.; Wyant, G.A.; Kim, C.; Laqtom, N.N.; Abbasi, M.; Chan, S.H.; Freinkman, E.; Sabatini, D.M. Lysosomal metabolomics reveals V-ATPase- and mTOR-dependent regulation of amino acid efflux from lysosomes. Science 2017, 358, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Shields, C.W.T.; Reyes, C.D.; Lopez, G.P. Microfluidic cell sorting: A review of the advances in the separation of cells from debulking to rare cell isolation. Lab. Chip 2015, 15, 1230–1249. [Google Scholar] [CrossRef] [Green Version]
- Rajotte, D.; Stearns, C.D.; Kabcenell, A.K. Isolation of mast cell secretory lysosomes using flow cytometry. Cytom. A 2003, 55, 94–101. [Google Scholar] [CrossRef]
- Diettrich, O.; Mills, K.; Johnson, A.W.; Hasilik, A.; Bryan, G. Winchester. Application of magnetic chromatography to the isolation of lysosomes from fibroblasts of patients with lysosomal storage disorders. FEBS Lett. 1998, 441, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Tharkeshwar, A.K.; Trekker, J.; Vermeire, W.; Pauwels, J.; Sannerud, R.; Priestman, D.A.; Te Vruchte, D.; Vints, K.; Baatsen, P.; Decuypere, J.P.; et al. A novel approach to analyze lysosomal dysfunctions through subcellular proteomics and lipidomics: The case of NPC1 deficiency. Sci. Rep. 2017, 7, 41408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoncu, R.; Bar-Peled, L.; Efeyan, A.; Wang, S.; Sancak, Y.; Sabatini, D.M. MTORC1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar H(+)-ATPase. Science 2011, 334, 678–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.W.; Freinkman, E.; Wang, T.; Birsoy, K.; Sabatini, D.M. Absolute quantification of matrix metabolites reveals the dynamics of mitochondrial metabolism. Cell 2016, 166, 1324–1337. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Freinkman, E.; Sabatini, D.M. Rapid immunopurification of mitochondria for metabolite profiling and absolute quantification of matrix metabolites. Nat. Protoc. 2017, 12, 2215–2231. [Google Scholar] [CrossRef]
- Corcelli, A.; Saponetti, M.S.; Zaccagnino, P.; Lopalco, P.; Mastrodonato, M.; Liquori, G.E.; Lorusso, M. Mitochondria isolated in nearly isotonic KCl buffer: Focus on cardiolipin and organelle morphology. Biochim. Biophys. Acta 2010, 1798, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; He, J.; Xie, W.P.; Hinojosa, E.; Ambati, C.S.R.; Putluri, N.; Kim, H.E.; Zhu, M.X.; Du, G. Rapid affinity purification of intracellular organelles using a twin strep tag. J. Cell Sci. 2019, 132, jcs235390. [Google Scholar] [CrossRef]
- Delgadillo, R.F.; Mueser, T.C.; Zaleta-Rivera, K.; Carnes, K.A.; Gonzalez-Valdez, J.; Parkhurst, L.J. Detailed characterization of the solution kinetics and thermodynamics of biotin, biocytin and HABA binding to avidin and streptavidin. PLoS ONE 2019, 14, e0204194. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Kaade, E.; Muntel, J.; Bruderer, R.; Reiter, L.; Thelen, M.; Winter, D. Systematic comparison of strategies for the enrichment of lysosomes by data independent acquisition. J. Proteome Res. 2020, 19, 371–381. [Google Scholar] [CrossRef]
- Vivas, O.; Tiscione, S.A.; Dixon, R.E.; Ory, D.S.; Dickson, E.J. Niemann-Pick type C disease reveals a link between lysosomal cholesterol and PtdIns (4,5) P2 that regulates neuronal excitability. Cell Rep. 2019, 27, 2636–2648. [Google Scholar] [CrossRef] [Green Version]
- Vanier, M.T. Complex lipid trafficking in Niemann-Pick disease type C. J. Inherit. Metab. Dis. 2015, 38, 187–199. [Google Scholar] [CrossRef]
- Lim, C.Y.; Davis, O.B.; Shin, H.R.; Zhang, J.; Berdan, C.A.; Jiang, X.; Counihan, J.L.; Ory, D.S.; Nomura, D.K.; Zoncu, R. ER-lysosome contacts enable cholesterol sensing by mTORC1 and drive aberrant growth signalling in Niemann-Pick type C. Nat. Cell Biol. 2019, 21, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Drizyte-Miller, K.; Chen, J.; Cao, H.; Schott, M.B.; McNiven, M.A. The small GTPase Rab32 resides on lysosomes to regulate mTORC1 signaling. J. Cell Sci. 2020, 133, jcs236661. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, E.C.; Baudrier, L.; Ozerdem, C.; Lewis, C.A.; Chan, S.H.; Kunchok, T.; Abu-Remaileh, M.; Cangelosi, A.L.; Sabatini, D.M.; Birsoy, K.; et al. MITO-Tag Mice enable rapid isolation and multimodal profiling of mitochondria from specific cell types in vivo. Proc. Natl. Acad. Sci. USA 2019, 116, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
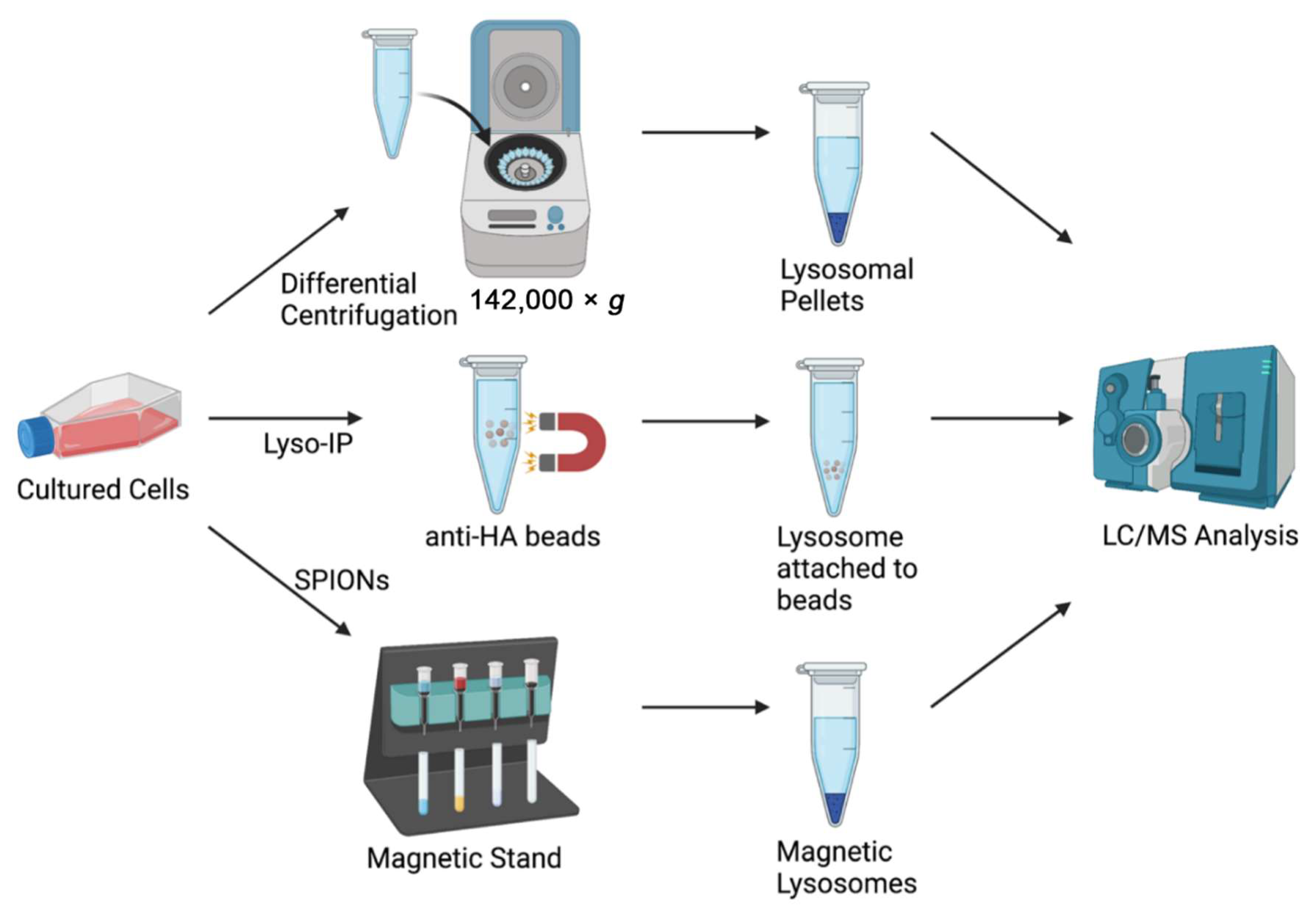

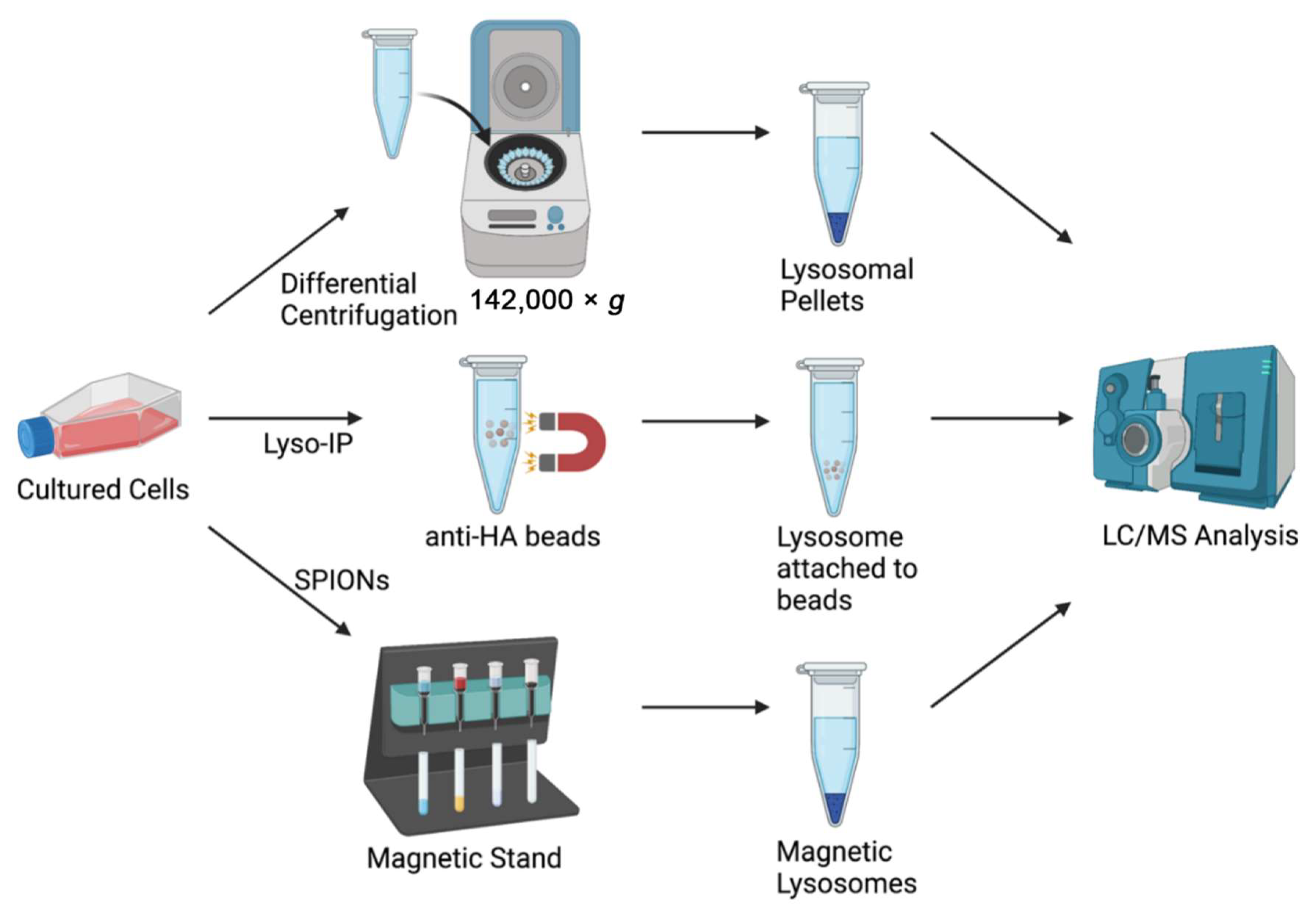{kind=link}
| Isolation Method | Advantages | Disadvantages |
|---|---|---|
| Density Gradient Differential Centrifugation [18] | Commonly used, well-established methods | High chance of contamination with other organelles (e.g., mitochondria and peroxisomes), long and laborious protocols, requires ultra-high-speed centrifuge |
| Fluorescence-Assisted Organelle Sorting (FAOS) [19] | High specificity and efficiency | Easy to break open organelles during sorting, difficult to sort small organelles (e.g., lysosomes), requires overexpression of fluorescently labeled lysosomal protein |
| Superparamagnetic Iron Oxide Nanoparticles (SPIONs) [20] | Higher yield, preserves integrity of lysosomes | Labor-intensive protocol to generate DMSA-coated SPIONs, requires use of dangerous chemicals, modifies native lysosomal environment |
| Affinity Purification and Immunoisolation [21] | No modifications of lysosome, efficient isolation | Long incubation times, requires large amounts of specific antibodies |
| Lyso-IP [22,23,24,25,26,27] | Rapid isolation of intact lysosomes (~10 min), uses LC/MS compatible buffer, preserves labile molecules, applicable to cellular and animal models, preserves integrity of lysosomes | Lower yield compared to SPIONs, higher variability in lysosomal proteins detected, requires overexpression of lysosomal transmembrane protein, not possible in human tissue samples |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Sidransky, E.; Chen, Y. Lyso-IP: Uncovering Pathogenic Mechanisms of Lysosomal Dysfunction. Biomolecules 2022, 12, 616. https://doi.org/10.3390/biom12050616
Chen C, Sidransky E, Chen Y. Lyso-IP: Uncovering Pathogenic Mechanisms of Lysosomal Dysfunction. Biomolecules. 2022; 12(5):616. https://doi.org/10.3390/biom12050616
Chicago/Turabian StyleChen, Chase, Ellen Sidransky, and Yu Chen. 2022. "Lyso-IP: Uncovering Pathogenic Mechanisms of Lysosomal Dysfunction" Biomolecules 12, no. 5: 616. https://doi.org/10.3390/biom12050616
APA StyleChen, C., Sidransky, E., & Chen, Y. (2022). Lyso-IP: Uncovering Pathogenic Mechanisms of Lysosomal Dysfunction. Biomolecules, 12(5), 616. https://doi.org/10.3390/biom12050616