
Apparent Absence of BMAL1-Dependent Skeletal Muscle–Kidney Cross Talk in Mice

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
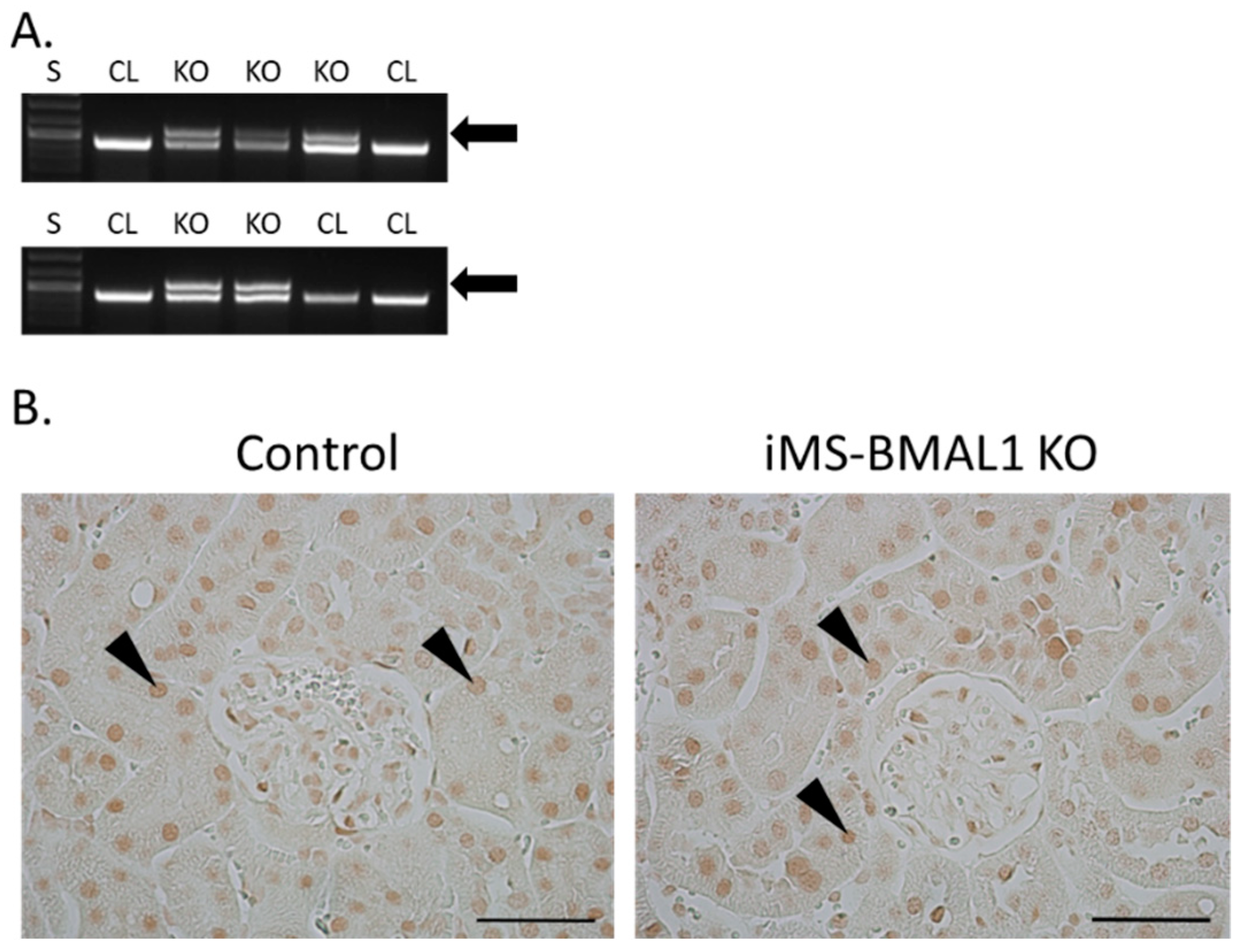
3.1. Verification of the iMS-BMAL1 KO Model
3.2. Fluid and Solute Handling under Basal Conditions and Following Potassium Depletion
3.3. Mitochondrial Function Assessment
3.4. Inflammatory Markers in the Kidney
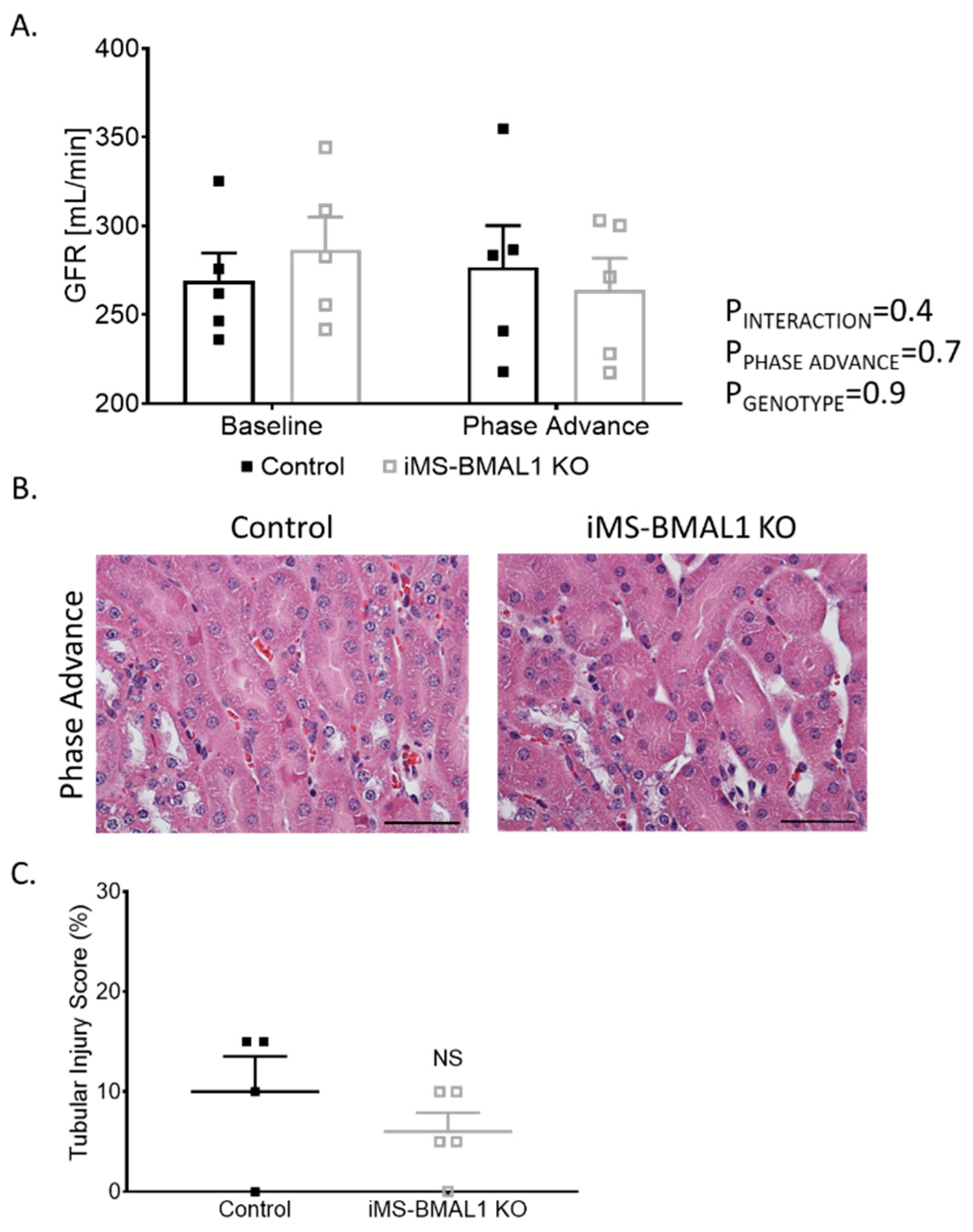
3.5. Renal Injury and Function
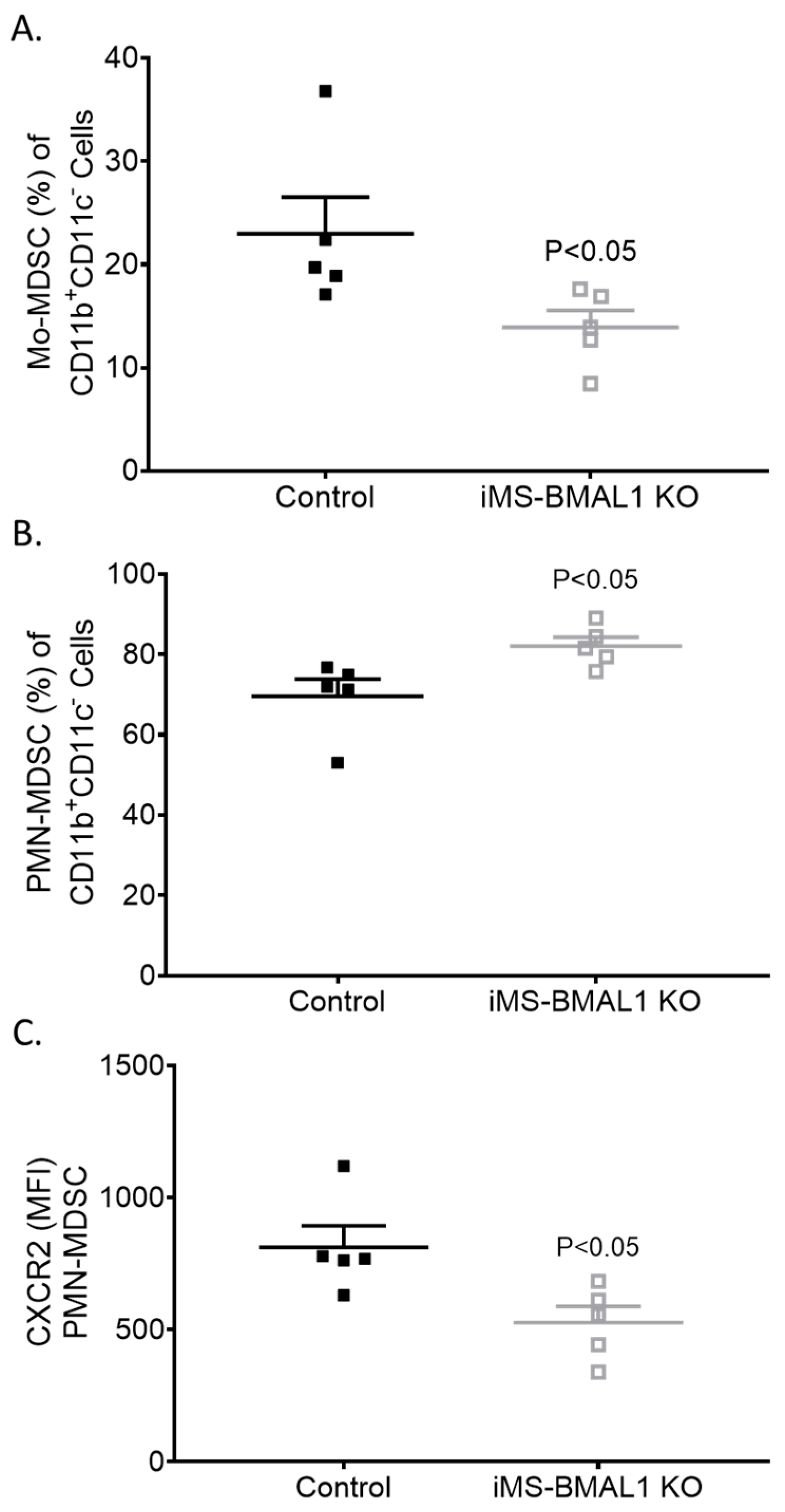
3.6. Extra-Renal Effects of iMS-BMAL1 KO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoo, S.H.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.K.; Oh, W.J.; Yoo, O.J.; et al. Period2: Luciferase real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumz, M.L. Molecular basis of circadian rhythmicity in renal physiology and pathophysiology. Exp. Physiol. 2016, 101, 1025–1029. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.; Gumz, M.L. Mechanism of the circadian clock in physiology. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R1053–R1064. [Google Scholar] [CrossRef] [PubMed]
- Egstrand, S.; Mace, M.L.; Olgaard, K.; Lewin, E. The vascular circadian clock in chronic kidney disease. Cells 2021, 10, 1769. [Google Scholar] [CrossRef] [PubMed]
- Schroder, E.A.; Harfmann, B.D.; Zhang, X.; Srikuea, R.; England, J.H.; Hodge, B.A.; Wen, Y.; Riley, L.A.; Yu, Q.; Christie, A.; et al. Intrinsic muscle clock is necessary for musculoskeletal health. J. Physiol. 2015, 593, 5387–5404. [Google Scholar] [CrossRef]
- Harfmann, B.D.; Schroder, E.A.; Kachman, M.T.; Hodge, B.A.; Zhang, X.; Esser, K.A. Muscle-specific loss of bmal1 leads to disrupted tissue glucose metabolism and systemic glucose homeostasis. Skelet. Muscle 2016, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, E.D.; Hughes, J.; Ferenbach, D.A. Renal aging: Causes and consequences. J. Am. Soc. Nephrol. 2017, 28, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Su, W.; Liu, S.; Zhao, G.; Esser, K.; Schroder, E.A.; Lefta, M.; Stauss, H.M.; Guo, Z.; Gong, M.C. Smooth-muscle bmal1 participates in blood pressure circadian rhythm regulation. J. Clin. Investig. 2015, 125, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Crislip, G.R.; Douma, L.G.; Masten, S.H.; Cheng, K.Y.; Lynch, I.J.; Johnston, J.G.; Barral, D.; Glasford, K.B.; Holzworth, M.R.; Verlander, J.W.; et al. Differences in renal bmal1 contribution to na. Am. J. Physiol. Ren. Physiol. 2020, 318, F1463–F1477. [Google Scholar] [CrossRef]
- Morla, L.; Shore, O.; Lynch, I.J.; Merritt, M.E.; Wingo, C.S. A noninvasive method to study the evolution of extracellular fluid volume in mice using time-domain nuclear magnetic resonance. Am. J. Physiol. Ren. Physiol. 2020, 319, F115–F124. [Google Scholar] [CrossRef] [PubMed]
- Hodge, B.A.; Wen, Y.; Riley, L.A.; Zhang, X.; England, J.H.; Harfmann, B.D.; Schroder, E.A.; Esser, K.A. The endogenous molecular clock orchestrates the temporal separation of substrate metabolism in skeletal muscle. Skelet. Muscle 2015, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, D.K.; Fasching, M.; Fontana-Ayoub, M.; Gnaiger, E. Cytochrome redox states and respiratory control in mouse and beef heart mitochondria at steady-state levels of hypoxia. J. Appl. Physiol. (1985) 2015, 119, 1210–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.C.; Fu, C.; Lu, Y.; Williams, M.A.; Pi, L.; Brantly, M.L.; Ventetuolo, C.E.; Raizada, M.K.; Mehrad, B.; Scott, E.W.; et al. Chemokine signaling axis between endothelial and myeloid cells regulates development of pulmonary hypertension associated with pulmonary fibrosis and hypoxia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 317, L434–L444. [Google Scholar] [CrossRef]
- Rabe, M.; Schaefer, F. Non-transgenic mouse models of kidney disease. Nephron 2016, 133, 53–61. [Google Scholar] [CrossRef]
- Dyar, K.A.; Ciciliot, S.; Wright, L.E.; Biensø, R.S.; Tagliazucchi, G.M.; Patel, V.R.; Forcato, M.; Paz, M.I.; Gudiksen, A.; Solagna, F.; et al. Muscle insulin sensitivity and glucose metabolism are controlled by the intrinsic muscle clock. Mol. Metab. 2014, 3, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.L.; Zhang, X.; McCarthy, J.J.; McDearmon, E.L.; Hornberger, T.A.; Russell, B.; Campbell, K.S.; Arbogast, S.; Reid, M.B.; Walker, J.R.; et al. Clock and bmal1 regulate myod and are necessary for maintenance of skeletal muscle phenotype and function. Proc. Natl. Acad. Sci. USA 2010, 107, 19090–19095. [Google Scholar] [CrossRef] [Green Version]
- Castanon-Cervantes, O.; Wu, M.; Ehlen, J.C.; Paul, K.; Gamble, K.L.; Johnson, R.L.; Besing, R.C.; Menaker, M.; Gewirtz, A.T.; Davidson, A.J. Dysregulation of inflammatory responses by chronic circadian disruption. J. Immunol. 2010, 185, 5796–5805. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Barrientos, A.; López-Romero, P.; Vivas, D.; Castro-Ferreira, F.; Núñez-Gil, I.; Franco, E.; Ruiz-Mateos, B.; García-Rubira, J.C.; Fernández-Ortiz, A.; Macaya, C.; et al. Circadian variations of infarct size in acute myocardial infarction. Heart 2011, 97, 970–976. [Google Scholar] [CrossRef] [Green Version]
- Wolff, G.; Duncan, M.J.; Esser, K.A. Chronic phase advance alters circadian physiological rhythms and peripheral molecular clocks. J. Appl. Physiol. (1985) 2013, 115, 373–382. [Google Scholar] [CrossRef]
- Davidson, A.J.; Sellix, M.T.; Daniel, J.; Yamazaki, S.; Menaker, M.; Block, G.D. Chronic jet-lag increases mortality in aged mice. Curr. Biol. 2006, 16, R914–R916. [Google Scholar] [CrossRef] [Green Version]
- Carré, J.E.; Affourtit, C. Mitochondrial activity and skeletal muscle insulin resistance in kidney disease. Int. J. Mol. Sci. 2019, 20, 2751. [Google Scholar] [CrossRef] [Green Version]
- Workeneh, B.T.; Mitch, W.E. Review of muscle wasting associated with chronic kidney disease. Am. J. Clin. Nutr. 2010, 91, 1128S–1132S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferenbach, D.A.; Bonventre, J.V. Mechanisms of maladaptive repair after aki leading to accelerated kidney ageing and ckd. Nat. Rev. Nephrol. 2015, 11, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, G.G.; Kapoor, S.C. Potassium depletion exacerbates essential hypertension. Ann. Intern. Med. 1991, 115, 77–83. [Google Scholar] [CrossRef]
- Gallen, I.W.; Rosa, R.M.; Esparaz, D.Y.; Young, J.B.; Robertson, G.L.; Batlle, D.; Epstein, F.H.; Landsberg, L. On the mechanism of the effects of potassium restriction on blood pressure and renal sodium retention. Am. J. Kidney Dis. 1998, 31, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Terker, A.S.; Zhang, C.; McCormick, J.A.; Lazelle, R.A.; Meermeier, N.P.; Siler, D.A.; Park, H.J.; Fu, Y.; Cohen, D.M.; Weinstein, A.M.; et al. Potassium modulates electrolyte balance and blood pressure through effects on distal cell voltage and chloride. Cell Metab. 2015, 21, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, I.S.; Larsen, C.K.; Leipziger, J.; Sørensen, M.V. Na(+) dependence of k(+) -induced natriuresis, kaliuresis and na(+) /cl(-) cotransporter dephosphorylation. Acta Physiol. (Oxf.) 2016, 218, 49–61. [Google Scholar]
- Pagliarini, D.J.; Calvo, S.E.; Chang, B.; Sheth, S.A.; Vafai, S.B.; Ong, S.E.; Walford, G.A.; Sugiana, C.; Boneh, A.; Chen, W.K.; et al. A mitochondrial protein compendium elucidates complex i disease biology. Cell 2008, 134, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Dong, Z. Mitochondria in kidney injury: When the power plant fails. J. Am. Soc. Nephrol. 2016, 27, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.E.; Chen, Y.; Priver, T.S.; Ralph, D.L.; Mercado, A.; Gamba, G.; Madhur, M.S.; McDonough, A.A. Coordinate adaptations of skeletal muscle and kidney to maintain extracellular [K+] during K+-deficient diet. Am. J. Physiol. Cell Physiol. 2020, 319, C757–C770. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Hong, Q.; Lin, H.; Zhu, H.; Liu, Q.; Wang, J.; Xie, Y.; Shang, X.; Shi, S.; et al. Timp-1 promotes age-related renal fibrosis through upregulating icam-1 in human timp-1 transgenic mice. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 1130–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.A.; Fraser, D.J.; Fielding, C.A.; Jones, G.W. Interleukin-6 in renal disease and therapy. Nephrol. Dial. Transplant. 2015, 30, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Liu, Y.; Wang, Q.; Chen, Y.; Liu, M.; Li, X.; Xiang, R.; Wei, Y.; Duan, Y.; Han, J. Tamoxifen induces the development of hernia in mice by activating mmp-2 and mmp-13 expression. Biochim. Biophys. Acta 2015, 1852, 1038–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.S.; Kim, I.J.; Choi, J.S.; Bae, E.H.; Ma, S.K.; Kim, S.W. Tamoxifen ameliorates obstructive nephropathy through src and the pi3k/akt/mtor pathway. Biol. Cell 2019, 111, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte (pg/mg) | Kidney Region | Control | iMS-BMAL1 KO |
|---|---|---|---|
| Interleukin 6 | Cortex | 1.8 ± 0.9 (4) | 13.2 ± 4.0 (4) * |
| Medulla | 1.7 ± 0.1 (3) | 6.2 ± 1.7 (4) | |
| Fibroblast Growth Factor 23 | Cortex | 7.0 ± 1.2 (4) | 6.2 ± 0.4 (5) |
| Medulla | 5.6 ± 3.4 (4) | 7.4 ± 1.3 (5) | |
| Interleukin 5 | Cortex | 0.8 ± 0.1 (4) | 0.7 ± 0.2 (5) |
| Interleukin 10 | Cortex | 2.4 ± 0.3 (4) | 1.9 ± 0.2 (5) |
| Medulla | 3.3 ± 0.4 (4) | 3.6 ± 0.6 (5) | |
| Interleukin 17 | Cortex | 0.3 ± 0.08 (4) | 0.3 ± 0.04 (5) |
| Renin | Cortex | 2710 ± 465 (4) | 2620 ± 134 (5) |
| Medulla | 3082 ± 233 (4) | 3937 ± 626 (5) | |
| Kidney Injury Molecule 1 | Cortex | 112 ± 5.3 (4) | 115 ± 11.1 (5) |
| Medulla | 251 ± 43 (4) | 282 ± 29 (5) | |
| Tissue Inhibitor of Metalloproteinase 1 | Cortex | 122 ± 14 (4) | 193 ± 30 (5) |
| Medulla | 42 ± 5 (4) | 82 ± 12 (5) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crislip, G.R.; Wohlgemuth, S.E.; Wolff, C.A.; Gutierrez-Monreal, M.A.; Douglas, C.M.; Ebrahimi, E.; Cheng, K.-Y.; Masten, S.H.; Barral, D.; Bryant, A.J.; et al. Apparent Absence of BMAL1-Dependent Skeletal Muscle–Kidney Cross Talk in Mice. Biomolecules 2022, 12, 261. https://doi.org/10.3390/biom12020261
Crislip GR, Wohlgemuth SE, Wolff CA, Gutierrez-Monreal MA, Douglas CM, Ebrahimi E, Cheng K-Y, Masten SH, Barral D, Bryant AJ, et al. Apparent Absence of BMAL1-Dependent Skeletal Muscle–Kidney Cross Talk in Mice. Biomolecules. 2022; 12(2):261. https://doi.org/10.3390/biom12020261
Chicago/Turabian StyleCrislip, Gene Ryan, Stephanie E. Wohlgemuth, Christopher A. Wolff, Miguel A. Gutierrez-Monreal, Collin M. Douglas, Elnaz Ebrahimi, Kit-Yan Cheng, Sarah H. Masten, Dominique Barral, Andrew J. Bryant, and et al. 2022. "Apparent Absence of BMAL1-Dependent Skeletal Muscle–Kidney Cross Talk in Mice" Biomolecules 12, no. 2: 261. https://doi.org/10.3390/biom12020261
APA StyleCrislip, G. R., Wohlgemuth, S. E., Wolff, C. A., Gutierrez-Monreal, M. A., Douglas, C. M., Ebrahimi, E., Cheng, K.-Y., Masten, S. H., Barral, D., Bryant, A. J., Esser, K. A., & Gumz, M. L. (2022). Apparent Absence of BMAL1-Dependent Skeletal Muscle–Kidney Cross Talk in Mice. Biomolecules, 12(2), 261. https://doi.org/10.3390/biom12020261