
Role of Natural Killer Cells during Pregnancy and Related Complications

 , ,
, , 

Abstract
1. Introduction
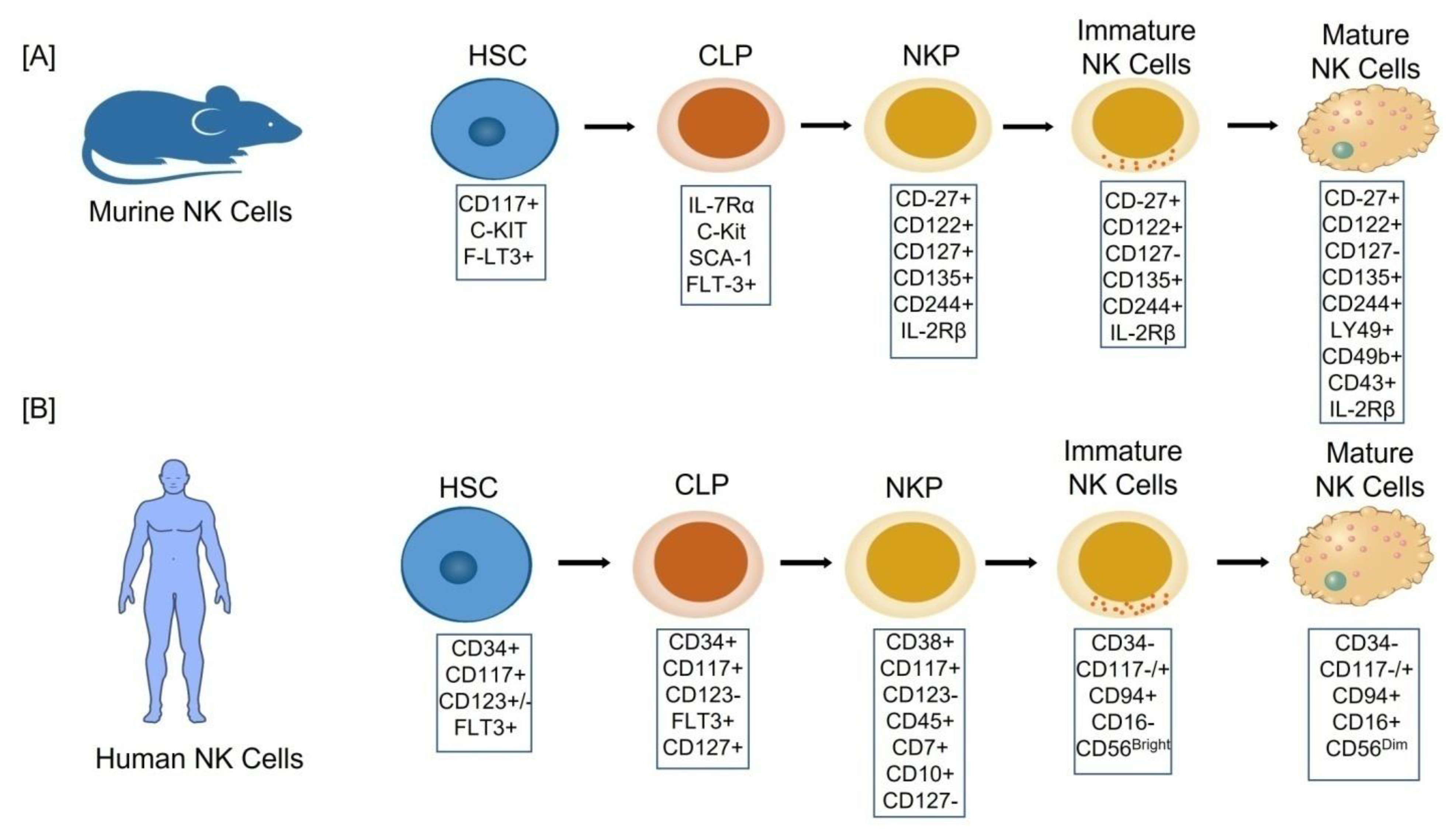
2. Development and Origin of NK Cells
2.1. Cellular Stages of NK Cell Development
- Stage 1: CD34+CD117+CD123+/−FLT3+,
- Stage 2: CD34+CD117+CD123−FLT3+CD127+,
- Stage 3: CD38+CD117+CD123− CD45+CD7+CD10+CD127−,
- Stage 4: (CD34−CD117−/+CD94+CD16−) also named as CD56bright,
- Stage 5: (CD34−CD117−/+CD94+CD16+) also named as CD56dim [14]
2.2. The Origin of uNK Cells
3. Phenotype and Subsets of dNK Cells
3.1. Surface Receptors of dNK Cells
3.2. Subsets of dNK Cells
4. Non-Pregnant Female Reproductive Tract NK Cells
5. NK Cells in Pregnant Female Reproductive Tract
5.1. NK Cells during Key Early Events of Pregnancy
5.2. An Overview of the Maternal-Fetal Interface Development
5.3. Role of dNK Cells at Maternal-Fetal Interface

{kind=link}
{kind=link}
| Maternal Phenotype | Fetal Phenotype | Effects | References |
|---|---|---|---|
| KIR2DS1+ | HLA-C2+ | Increase trophoblast cell invastion potential. Boost regulatory T cells. Create IDO producing monocytes. Induce effector T cell apoptosis | [71,81,82,83,84] |
| KIR AA | Homozygous HLA-C2 or Heterozygous HLA-C1C2 | Increase the risk of preeclampsia | [86] |
| KIR2DL1 | HLA-C2 | Repress dNK cells and enable to produce cytokines Mediate strong inhibitory signals leads to the placental abnormalities. Affect the ability of dNK cells to regulate trophoblastic cell invasion. | [86,87] |
| NKG2A | HLA-E | Inhibit NK cell activation. | [89] |
| ILT2 | HLA-G | Promotes the secretion inflammatory & angiogenic cytokines by dNK cells. | [89] |
6. NK Cell Dysfunction in Pregnancy Pathology
6.1. Preeclampsia (PE)
6.2. Recurrent Implantation Failure (RIF)
6.3. Recurrent Miscarriage (RM)
7. Immunomodulatory Strategies for Treatment of Pregnancy Complications
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Billingham, R.E.; Brent, L.; Medawar, P.B. Actively acquired tolerance of foreign cells. Nature 1953, 172, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Jabrane-Ferrat, N.; Siewiera, J. The up side of decidual natural killer cells: New developments in immunology of pregnancy. Immunology 2014, 141, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Mold, J.E.; Michaëlsson, J.; Burt, T.D.; Muench, M.O.; Beckerman, K.P.; Busch, M.P.; Lee, T.H.; Nixon, D.F.; McCune, J.M. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science 2008, 322, 1562–1565. [Google Scholar] [CrossRef]
- Sojka, D.K.; Yang, L.; Yokoyama, W.M. Uterine Natural Killer Cells. Front. Immunol. 2019, 10, 960. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef]
- Sojka, D.K.; Plougastel-Douglas, B.; Yang, L.; Pak-Wittel, M.A.; Artyomov, M.N.; Ivanova, Y.; Zhong, C.; Chase, J.M.; Rothman, P.B.; Yokoyama, W.M.; et al. Tissue-resident natural killer (NK) cells are cell lineages distinct from thymic and conventional splenic NK cells. Elife 2014, 3, e01659. [Google Scholar] [CrossRef]
- Yang, S.L.; Wang, H.Y.; Li, D.J.; Li, M.Q. Role of DecidualNatural Killer Cells at the Maternal–Fetal Interface during Pregnancy. Reprod. Dev. Med. 2019, 3, 165–169. [Google Scholar]
- Acar, N.; Ustunel, I.; Demir, R. Uterine natural killer (uNK) cells and their missions during pregnancy: A review. Actahistochemica 2011, 113, 82–91. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Cerwenka, A.; Lanier, L.L. Natural killer cell memory in infection, inflammation and cancer. Nat. Rev. Immunol. 2016, 16, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, E.; Malarkannan, S. Tissue-Resident NK Cells: Development, Maturation, and Clinical Relevance. Cancers 2020, 12, 1553. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Freud, A.G.; Caligiuri, M.A. Location and cellular stages of natural killer cell development. Trends Immunol. 2013, 34, 573–582. [Google Scholar] [CrossRef]
- Stabile, H.; Fionda, C.; Santoni, A.; Gismondi, A. Impact of bone marrow-derived signals on NK cell development and functional maturation. Cytokinegrowth Factor Rev. 2018, 42, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Vosshenrich, C.A.; Di Santo, J.P. Developmental programming of natural killer and innate lymphoid cells. Curr. Opin. Immunol. 2013, 25, 130–138. [Google Scholar] [CrossRef]
- Rosmaraki, E.E.; Douagi, I.; Roth, C.; Colucci, F.; Cumano, A.; Di Santo, J.P. Identification of committed NK cell progenitors in adult murine bone marrow. Eur. J. Immunol. 2001, 31, 1900–1909. [Google Scholar] [CrossRef]
- Kim, S.; Iizuka, K.; Kang, H.S.; Dokun, A.; French, A.R.; Greco, S.; Yokoyama, W.M. In vivo developmental stages in murine natural killer cell maturation. Nat. Immunol. 2002, 3, 523–528. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Smyth, M.J. CD27 dissects mature NK cells into two subsets with distinct responsiveness and migratory capacity. J. Immunol. 2006, 176, 1517–1524. [Google Scholar] [CrossRef]
- Renoux, V.M.; Zriwil, A.; Peitzsch, C.; Michaëlsson, J.; Friberg, D.; Soneji, S.; Sitnicka, E. Identification of a Human Natural Killer Cell Lineage-Restricted Progenitor in Fetal and Adult Tissues. Immunity 2015, 43, 394–407. [Google Scholar] [CrossRef]
- Bonanni, V.; Sciumè, G.; Santoni, A.; Bernardini, G. Bone Marrow NK Cells: Origin, Distinctive Features, and Requirements for Tissue Localization. Front. Immunol. 2019, 10, 1569. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56(bright) subset. Blood 2001, 97, 3146–3151. [Google Scholar] [CrossRef]
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Stabile, H.; Nisti, P.; Morrone, S.; Pagliara, D.; Bertaina, A.; Locatelli, F.; Santoni, A.; Gismondi, A. Multifunctional human CD56 low CD16 low natural killer cells are the prominent subset in bone marrow of both healthy pediatric donors and leukemic patients. Haematologica 2015, 100, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.K.; Yang, L.; Yokoyama, W.M. Uterine natural killer cells: To protect and to nurture. Birth Defects Res. 2018, 110, 1531–1538. [Google Scholar] [CrossRef]
- Chantakru, S.; Miller, C.; Roach, L.E.; Kuziel, W.A.; Maeda, N.; Wang, W.C.; Evans, S.S.; Croy, B.A. Contributions from self-renewal and trafficking to the uterine NK cell population of early pregnancy. J. Immunol. 2002, 168, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Croy, B.A.; Esadeg, S.; Chantakru, S.; van den Heuvel, M.; Paffaro, V.A.; He, H.; Black, G.P.; Ashkar, A.A.; Kiso, Y.; Zhang, J. Update on pathways regulating the activation of uterine Natural Killer cells, their interactions with decidual spiral arteries and homing of their precursors to the uterus. J. Reprod. Immunol. 2003, 59, 175–191. [Google Scholar] [CrossRef]
- Borzychowski, A.M.; Chantakru, S.; Minhas, K.; Paffaro, V.A.; Yamada, A.T.; He, H.; Korach, K.S.; Croy, B.A. Functional analysis of murine uterine natural killer cells genetically devoid of oestrogenreceptors. Placenta 2003, 24, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Ashkar, A.A.; Black, G.P.; Wei, Q.; He, H.; Liang, L.; Head, J.R.; Croy, B.A. Assessment of requirements for IL-15 and IFN regulatory factors in uterine NK cell differentiation and function during pregnancy. J. Immunol. 2003, 171, 2937–2944. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.K.; Yang, L.; Plougastel-Douglas, B.; Higuchi, D.A.; Croy, B.A.; Yokoyama, W.M. Cutting Edge: Local Proliferation of Uterine Tissue-Resident NK Cells during Decidualization in Mice. J. Immunol. 2018, 201, 2551–2556. [Google Scholar] [CrossRef] [PubMed]
- Boulenouar, S.; Doisne, J.M.; Sferruzzi-Perri, A.; Gaynor, L.M.; Kieckbusch, J.; Balmas, E.; Yung, H.W.; Javadzadeh, S.; Volmer, L.; Hawkes, D.A.; et al. The Residual Innate Lymphoid Cells in NFIL3-Deficient Mice Support Suboptimal Maternal Adaptations to Pregnancy. Front. Immunol. 2016, 7, 43. [Google Scholar] [CrossRef]
- Redhead, M.L.; Portilho, N.A.; Felker, A.M.; Mohammad, S.; Mara, D.L.; Croy, B.A. The Transcription Factor NFIL3 Is Essential for Normal Placental and Embryonic Development but Not for Uterine Natural Killer (UNK) Cell Differentiation in Mice. Biol. Reprod. 2016, 94, 101. [Google Scholar] [CrossRef]
- Taylor, E.B.; Sasser, J.M. Natural killer cells and T lymphocytes in pregnancy and pre-eclampsia. Clin. Sci. 2017, 131, 2911–2917. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, C.; Juelke, K.; Falco, M.; Morandi, B.; D’Agostino, A.; Costa, R.; Ratto, G.; Forte, G.; Carrega, P.; Lui, G.; et al. CD56brightCD16- killer Ig-like receptor- NK cells display longer telomeres and acquire features of CD56dim NK cells upon activation. J. Immunol. 2007, 178, 4947–4955. [Google Scholar] [CrossRef] [PubMed]
- Montaldo, E.; Vacca, P.; Chiossone, L.; Croxatto, D.; Loiacono, F.; Martini, S.; Ferrero, S.; Walzer, T.; Moretta, L.; Mingari, M.C. Unique Eomes(+) NK Cell Subsets Are Present in Uterus and Decidua During Early Pregnancy. Front. Immunol. 2016, 6, 646. [Google Scholar] [CrossRef] [PubMed]
- Cerdeira, A.S.; Rajakumar, A.; Royle, C.M.; Lo, A.; Husain, Z.; Thadhani, R.I.; Sukhatme, V.P.; Karumanchi, S.A.; Kopcow, H.D. Conversion of peripheral blood NK cells to a decidual NK-like phenotype by a cocktail of defined factors. J. Immunol. 2013, 190, 3939–3948. [Google Scholar] [CrossRef]
- Vacca, P.; Moretta, L.; Moretta, A.; Mingari, M.C. Origin, phenotype and function of human natural killer cells in pregnancy. Trends Immunol. 2011, 32, 517–523. [Google Scholar] [CrossRef]
- Chazara, O.; Xiong, S.; Moffett, A. Maternal KIR and fetal HLA-C: A fine balance. J. Leukoc. Biol. 2011, 90, 703–716. [Google Scholar] [CrossRef]
- Santoni, A.; Carlino, C.; Stabile, H.; Gismondi, A. Mechanisms underlying recruitment and accumulation of decidual NK cells in uterus during pregnancy. Am. J. Reprod. Immunol. 2008, 59, 417–424. [Google Scholar] [CrossRef]
- Santoni, A.; Zingoni, A.; Cerboni, C.; Gismondi, A. Natural killer (NK) cells from killers to regulators: Distinct features between peripheral blood and decidual NK cells. Am. J. Reprod. Immunol. 2007, 58, 280–288. [Google Scholar] [CrossRef]
- Chantakru, S.; Kuziel, W.A.; Maeda, N.; Croy, B.A. A study on the density and distribution of uterine Natural Killer cells at mid pregnancy in mice genetically-ablated for CCR2, CCR 5 and the CCR5 receptor ligand, MIP-1 alpha. J. Reprod. Immunol. 2001, 49, 33–47. [Google Scholar] [CrossRef]
- Bulmer, J.N.; Lash, G.E. The Role of Uterine NK Cells in Normal Reproduction and Reproductive Disorders. Adv. Exp. Med. Biol. 2015, 868, 95–126. [Google Scholar] [PubMed]
- Kopcow, H.D.; Eriksson, M.; Mselle, T.F.; Damrauer, S.M.; Wira, C.R.; Sentman, C.L.; Strominger, J.L. Human decidual NK cells from gravid uteri and NK cells from cycling endometrium are distinct NK cell subsets. Placenta 2010, 31, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.E.; Stephenson, E.; Polański, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Jabrane-Ferrat, N. Features of Human Decidual NK Cells in Healthy Pregnancy and During Viral Infection. Front. Immunol. 2019, 10, 1397. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, H.; Morozov, P.; Straus, A.; Sahasrabudhe, N.; Max, K.; Garzia, A.; Kustagi, M.; Tuschl, T.; Williams, Z. A single-cell survey of the human first-trimester placenta and decidua. Sci. Adv. 2018, 4, eaau4788. [Google Scholar] [CrossRef]
- Fu, B.; Zhou, Y.; Ni, X.; Tong, X.; Xu, X.; Dong, Z.; Sun, R.; Tian, Z.; Wei, H. Natural Killer Cells Promote Fetal Development through the Secretion of Growth-Promoting Factors. Immunity 2017, 47, 1100–1113.e6. [Google Scholar] [CrossRef]
- Sentman, C.L.; Wira, C.R.; Eriksson, M. NK cell function in the human female reproductive tract. Am. J. Reprod. Immunol. 2017, 57, 108–115. [Google Scholar] [CrossRef]
- Givan, A.L.; White, H.D.; Stern, J.E.; Colby, E.; Gosselin, E.J.; Guyre, P.M.; Wira, C.R. Flow cytometric analysis of leukocytes in the human female reproductive tract: Comparison of fallopian tube, uterus, cervix, and vagina. Am. J. Reprod. Immunol. 1997, 38, 350–359. [Google Scholar] [CrossRef]
- Bulmer, J.N.; Lash, G.E. Human uterine natural killer cells: A reappraisal. Mol. Immunol. 2005, 42, 511–521. [Google Scholar] [CrossRef]
- Loke, Y.W.; King, A. Human Implantation: Cell Biology and Immunology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Moffett-King, A. Natural killer cells and pregnancy. Nat. Rev. Immunol. 2002, 2, 656–663. [Google Scholar] [CrossRef]
- Le Bouteiller, P.; Piccinni, M.P. Human NK cells in pregnant uterus: Why there? Am. J. Reprod. Immunol. 2008, 59, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, T.; Konno, R.; Okamoto, S.; Moriya, T.; Satoh, S.; Yajima, A. Involvement of granule-mediated apoptosis in the cyclic changes of the normal human endometrium. Tohoku J. Exp. Med. 2001, 193, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Trundley, A.; Moffett, A. Human uterine leukocytes and pregnancy. Tissue Antigens 2004, 63, 1–12. [Google Scholar] [CrossRef]
- Jones, R.K.; Bulmer, J.N.; Searle, R.F. Phenotypic and functional studies of leukocytes in human endometrium and endometriosis. Hum. Reprod. Update 1998, 4, 702–709. [Google Scholar] [CrossRef]
- Kodama, T.; Hara, T.; Okamoto, E.; Kusunoki, Y.; Ohama, K. Characteristic changes of large granular lymphocytes that strongly express CD56 in endometrium during the menstrual cycle and early pregnancy. Hum. Reprod. 1998, 13, 1036–1043. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, X.F.; Charnock-Jones, D.S.; Zhang, E.; Hiby, S.; Malik, S.; Day, K.; Licence, D.; Bowen, J.M.; Gardner, L.; King, A.; et al. Angiogenic growth factor messenger ribonucleic acids in uterine natural killer cells. J. Clin. Endocrinol. Metab. 2001, 86, 1823–1834. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, M.; Lee, S.K. Role of endometrial immune cells in implantation. Clin. Exp. Reprod. Med. 2011, 38, 119–125. [Google Scholar] [CrossRef]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Croy, B.A. Interferon-gamma contributes to the normalcy of murine pregnancy. Biol. Reprod. 1999, 61, 493–502. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Di Santo, J.P.; Croy, B.A. Interferon gamma contributes to initiation of uterine vascular modification, decidual integrity, and uterine natural killer cell maturation during normal murine pregnancy. J. Exp. Med. 2000, 192, 259–270. [Google Scholar] [CrossRef]
- Norwitz, E.R. Defective implantation and placentation: Laying the blueprint for pregnancy complications. Reprod. Biomed. Online 2006, 13, 591–599. [Google Scholar] [CrossRef]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Kirkley, M.; De Oliveira, L.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Interferon-gamma inhibits extravillous trophoblast cell invasion by a mechanism that involves both changes in apoptosis and protease levels. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 2512–2518. [Google Scholar]
- Wallace, A.E.; Fraser, R.; Cartwright, J.E. Extravillous trophoblast and decidual natural killer cells: A remodellingpartnership. Hum. Reprod. Update 2012, 18, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef]
- Morelli, S.; Mandal, M.; Goldsmith, L.T.; Kashani, B.N.; Ponzio, N.M. The maternal immune system during pregnancy and its influence on fetal development. Res. Rep. Biol. 2015, 6, 171–189. [Google Scholar] [CrossRef]
- Guimond, M.J.; Luross, J.A.; Wang, B.; Terhorst, C.; Danial, S.; Croy, B.A. Absence of natural killer cells during murine pregnancy is associated with reproductive compromise in TgE26 mice. Biol. Reprod. 1997, 56, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Guimond, M.J.; Wang, B.; Croy, B.A. Engraftment of bone marrow from severe combined immunodeficient (SCID) mice reverses the reproductive deficits in natural killer cell-deficient tg epsilon 26 mice. J. Exp. Med. 1998, 187, 217–223. [Google Scholar] [CrossRef]
- Red-Horse, K.; Zhou, Y.; Genbacev, O.; Prakobphol, A.; Foulk, R.; McMaster, M.; Fisher, S.J. Trophoblast differentiation during embryo implantation and formation of the maternal-fetal interface. J. Clin. Investig. 2004, 114, 744–754. [Google Scholar] [CrossRef]
- Arck, P.C.; Hecher, K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat. Med. 2013, 19, 548–556. [Google Scholar] [CrossRef]
- Fu, B.; Wei, H. Decidual natural killer cells and the immune microenvironment at the maternal-fetal interface. Sci. ChinaLife Sci. 2016, 59, 1224–1231. [Google Scholar] [CrossRef]
- Dey, S.K.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.C.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev. 2004, 25, 341–373. [Google Scholar] [CrossRef]
- Gellersen, B.; Brosens, I.A.; Brosens, J.J. Decidualization of the human endometrium: Mechanisms, functions, and clinical perspectives. Semin. Reprod. Med. 2007, 25, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Yoshimura, Y. Molecular and cellular mechanisms for differentiation and regeneration of the uterine endometrium. Endocr. J. 2008, 55, 795–810. [Google Scholar] [CrossRef]
- Ramathal, C.Y.; Bagchi, I.C.; Taylor, R.N.; Bagchi, M.K. Endometrial decidualization: Of mice and men. Semin. Reprod. Med. 2010, 28, 17–26. [Google Scholar] [CrossRef]
- Burton, G.J.; Watson, A.L.; Hempstock, J.; Skepper, J.N.; Jauniaux, E. Uterine glands provide histiotrophic nutrition for the human fetus during the first trimester of pregnancy. J. Clin. Endocrinol. Metab. 2002, 87, 2954–2959. [Google Scholar] [CrossRef]
- Hempstock, J.; Cindrova-Davies, T.; Jauniaux, E.; Burton, G.J. Endometrial glands as a source of nutrients, growth factors and cytokines during the first trimester of human pregnancy: A morphological and immunohistochemical study. Reprod. Biol. Endocrinol. 2004, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.W.; Norwitz, G.A.; Pavlicev, M.; Tilburgs, T.; Simón, C.; Norwitz, E.R. Endometrial Decidualization: The Primary Driver of Pregnancy Health. Int. J. Mol. Sci. 2020, 21, 4092. [Google Scholar] [CrossRef]
- Demir, R.; Kayisli, U.A.; Celik-Ozenci, C.; Korgun, E.T.; Demir-Weusten, A.Y.; Arici, A. Structural differentiation of human uterine luminal and glandular epithelium during early pregnancy: An ultrastructural and immunohistochemical study. Placenta 2002, 23, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.D.; Zhang, J.; Dunk, C.; Lye, S.J.; Croy, B.A. Leukocyte driven-decidual angiogenesis in early pregnancy. Cell. Mol. Immunol. 2014, 11, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Sharkey, A.M.; Kennedy, P.R.; Gardner, L.; Farrell, L.E.; Chazara, O.; Bauer, J.; Hiby, S.E.; Colucci, F.; Moffett, A. Maternal uterine NK cell-activating receptor KIR2DS1 enhances placentation. J. Clin. Investig. 2013, 123, 4264–4272. [Google Scholar] [CrossRef]
- Fu, B.; Li, X.; Sun, R.; Tong, X.; Ling, B.; Tian, Z.; Wei, H. Natural killer cells promote immune tolerance by regulating inflammatory TH17 cells at the human maternal-fetal interface. Proc. Natl. Acad. Sci. USA 2013, 110, E231–E240. [Google Scholar] [CrossRef]
- Vacca, P.; Cantoni, C.; Vitale, M.; Prato, C.; Canegallo, F.; Fenoglio, D.; Ragni, N.; Moretta, L.; Mingari, M.C. Crosstalk between decidual NK and CD14+ myelomonocytic cells results in induction of Tregs and immunosuppression. Proc. Natl. Acad. Sci. USA 2010, 107, 11918–11923. [Google Scholar] [CrossRef]
- Felker, A.M.; Croy, B.A. Uterine natural killer cell partnerships in early mouse decidua basalis. J. Leukoc. Biol. 2016, 100, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Male, V.; Sharkey, A.; Masters, L.; Kennedy, P.R.; Farrell, L.E.; Moffett, A. The effect of pregnancy on the uterine NK cell KIR repertoire. Eur. J. Immunol. 2011, 41, 3017–3027. [Google Scholar] [CrossRef] [PubMed]
- Parham, P.; Moffett, A. Variable NK cell receptors and their MHC class I ligands in immunity, reproduction and human evolution. Nat. Rev. Immunol. 2013, 13, 133–144. [Google Scholar] [CrossRef]
- Hiby, S.E.; Apps, R.; Sharkey, A.M.; Farrell, L.E.; Gardner, L.; Mulder, A.; Claas, F.H.; Walker, J.J.; Redman, C.W.; Morgan, L.; et al. Maternal activating KIRs protect against human reproductive failure mediated by fetal HLA-C2. J. Clin. Investig. 2010, 120, 4102–4110. [Google Scholar] [CrossRef] [PubMed]
- Hiby, S.E.; Apps, R.; Chazara, O.; Farrell, L.E.; Magnus, P.; Trogstad, L.; Gjessing, H.K.; Carrington, M.; Moffett, A. Maternal KIR in combination with paternal HLA-C2 regulate human birth weight. J. Immunol. 2014, 192, 5069–5073. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Bryceson, Y.T.; Kuppusamy, S.P.; Geraghty, D.E.; van der Meer, A.; Joosten, I.; Long, E.O. Activation of NK cells by an endocytosed receptor for soluble HLA-G. PLoS Biol. 2006, 4, e9. [Google Scholar] [CrossRef]
- Beaman, K.D.; Jaiswal, M.K.; Dambaeva, S.; Gilman-Sachs, A. Future directions of clinical laboratory evaluation of pregnancy. Cell. Mol. Immunol. 2014, 11, 582–588. [Google Scholar] [CrossRef]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef]
- Moffett, A.; Hiby, S.E. How Does the maternal immune system contribute to the development of pre-eclampsia? Placenta 2007, 28, S51–S56. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Loke, C. Immunology of placentation in eutherian mammals. Nat. Rev. Immunol. 2006, 6, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, T.; Hebisch, G.; Orban, P.; Lü, X. Aberrant positioning of trophoblast and lymphocytes in the feto-maternal interface with pre-eclampsia. Virchows Archiv. 1999, 434, 207–211. [Google Scholar] [CrossRef]
- Wilczyński, J.R.; Tchórzewski, H.; Banasik, M.; Głowacka, E.; Wieczorek, A.; Lewkowicz, P.; Malinowski, A.; Szpakowski, M.; Wilczyński, J. Lymphocyte subset distribution and cytokine secretion in third trimester decidua in normal pregnancy and preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 109, 8–15. [Google Scholar] [CrossRef]
- Bachmayer, N.; Rafik Hamad, R.; Liszka, L.; Bremme, K.; Sverremark-Ekström, E. Aberrant uterine natural killer (NK)-cell expression and altered placental and serum levels of the NK-cell promoting cytokine interleukin-12 in pre-eclampsia. Am. J. Reprod. Immunol. 2006, 56, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.J.; Bulmer, J.N.; Searle, R.F.; Innes, B.A.; Robson, S.C. Altered decidual leucocyte populations in the placental bed in pre-eclampsia and foetal growth restriction: A comparison with late normal pregnancy. Reproduction 2009, 138, 177–184. [Google Scholar] [CrossRef]
- Wallace, A.E.; Whitley, G.S.; Thilaganathan, B.; Cartwright, J.E. Decidual natural killer cell receptor expression is altered in pregnancies with impaired vascular remodeling and a higher risk of pre-eclampsia. J. Leukoc. Biol. 2015, 97, 79–86. [Google Scholar] [CrossRef]
- Shreeve, N.; Depierreux, D.; Hawkes, D.; Traherne, J.A.; Sovio, U.; Huhn, O.; Jayaraman, J.; Horowitz, A.; Ghadially, H.; Perry, J.; et al. The CD94/NKG2A inhibitory receptor educates uterine NK cells to optimize pregnancy outcomes in humans and mice. Immunity 2021, 54, 1231–1244. [Google Scholar] [CrossRef]
- Caniggia, I.; Grisaru-Gravnosky, S.; Kuliszewsky, M.; Post, M.; Lye, S.J. Inhibition of TGF-beta 3 restores the invasive capability of extravillous trophoblasts in preeclamptic pregnancies. J. Clin. Investig. 1999, 103, 1641–1650. [Google Scholar] [CrossRef]
- Zhang, J.; Dunk, C.E.; Shynlova, O.; Caniggia, I.; Lye, S.J. TGFb1 suppresses the activation of distinct dNK subpopulations in preeclampsia. EBioMedicine 2019, 39, 531–539. [Google Scholar] [CrossRef]
- Du, M.; Wang, W.; Huang, L.; Guan, X.; Lin, W.; Yao, J.; Li, L. Natural Killer Cells in the Pathogenesis of Preeclampsia: A Double-Edged Sword. J. Matern. Fetal Neonatal Med. 2020, 19, 1–8. [Google Scholar] [CrossRef]
- Milosevic-Stevanovic, J.; Krstic, M.; Radovic-Janosevic, D.; Popovic, J.; Tasic, M.; Stojnev, S. Number of Decidual Natural Killer Cells & Macrophages in Pre-Eclampsia. Indian J. Med. Res. 2016, 144, 823–830. [Google Scholar] [PubMed]
- Polanski, L.T.; Baumgarten, M.N.; Quenby, S.; Brosens, J.; Campbell, B.K.; Raine-Fenning, N.J. What exactly do we mean by ‘recurrent implantation failure’? A systematic review and opinion. Reprod. Biomed. Online 2014, 28, 409–423. [Google Scholar] [CrossRef]
- Moffett, A.; Colucci, F. Uterine NK cells: Active regulators at the maternal-fetal interface. J. Clin. Investig. 2014, 124, 1872–1879. [Google Scholar] [CrossRef]
- Matteo, M.G.; Greco, P.; Rosenberg, P.; Mestice, A.; Baldini, D.; Falagario, T.; Martino, V.; Santodirocco, M.; Massenzio, F.; Castellana, L.; et al. Normal percentage of CD56bright natural killer cells in young patients with a history of repeated unexplained implantation failure after in vitro fertilization cycles. Fertil. Steril. 2007, 88, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Tuckerman, E.; Mariee, N.; Prakash, A.; Li, T.C.; Laird, S. Uterine natural killer cells in peri-implantation endometrium from women with repeated implantation failure after IVF. J. Reprod. Immunol. 2010, 87, 60–66. [Google Scholar] [CrossRef]
- Mariee, N.; Li, T.C.; Laird, S.M. Expression of leukaemia inhibitory factor and interleukin 15 in endometrium of women with recurrent implantation failure after IVF; correlation with the number of endometrial natural killer cells. Hum. Reprod. 2012, 27, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Lédée-Bataille, N.; Dubanchet, S.; Coulomb-L’hermine, A.; Durand-Gasselin, I.; Frydman, R.; Chaouat, G. A new role for natural killer cells, interleukin (IL)-12, and IL-18 in repeated implantation failure after in vitro fertilization. Fertil. Steril. 2004, 81, 59–65. [Google Scholar] [CrossRef]
- Donoghue, J.; Paiva, P.; Teh, W.; Cann, L.; Nowell, C.; Rees, H.; Bittinger, S.; Obers, V.; Bulmer, J.; Stern, C.; et al. Endometrial uNK cell counts do not predict successful implantation in an IVF population. Hum. Reprod. 2019, 34, 1–11. [Google Scholar] [CrossRef]
- Alecsandru, D.; Garrido, N.; Vicario, J.; Barrio, A.; Aparicio, P.; Requena, A.; Garcia-Velasco, J. Maternal KIR haplotype influences live birth rate after double embryo transfer in IVF cycles in patients with recurrent miscarriages and implantation failure. Hum. Reprod. 2014, 29, 2637–2643. [Google Scholar] [CrossRef]
- Tang, A.W.; Alfirevic, Z.; Quenby, S. Natural killer cells and pregnancy outcomes in women with recurrent miscarriage and infertility: A systematic review. Hum. Reprod. 2011, 26, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Royal College of Obstetricians and Gynaecologists. The Investigation and Treatment of Couples with Recurrent First-Trimester and Second-Trimester Miscarriage; Green-top Guideline No. 17; RCOG: London, UK, 2011. [Google Scholar]
- American College of Obstetricians and Gynecologists. The American College of Obstetricians and Gynecologists Practice Bulletin no. 150. Early pregnancy loss. Obstet. Gynecol. 2015, 125, 1258–1267. [Google Scholar] [CrossRef]
- Chong, H.; Quenby, S. Natural killer cells and reproductive health. Obstet. Gynaecol. 2016, 18, 91–97. [Google Scholar] [CrossRef]
- Clifford, K.; Flanagan, A.M.; Regan, L. Endometrial CD56+ natural killer cells in women with recurrent miscarriage: A histomorphometricstudy. Hum. Reprod. 1999, 14, 2727–2730. [Google Scholar] [CrossRef]
- Quenby, S.; Bates, M.; Doig, T.; Brewster, J.; Lewis-Jones, D.I.; Johnson, P.M.; Vince, G. Pre-implantation endometrial leukocytes in women with recurrent miscarriage. Hum. Reprod. 1999, 14, 2386–2391. [Google Scholar] [CrossRef]
- Tuckerman, E.; Laird, S.M.; Prakash, A.; Li, T.C. Prognostic value of the measurement of uterine natural killer cells in the endometrium of women with recurrent miscarriage. Hum. Reprod. 2007, 22, 2208–2213. [Google Scholar] [CrossRef] [PubMed]
- Lachapelle, M.H.; Miron, P.; Hemmings, R.; Roy, D.C. Endometrial T, B, and NK cells in patients with recurrent spontaneous abortion. Altered profile and pregnancy outcome. J. Immunol. 1996, 156, 4027–4034. [Google Scholar]
- Shimada, S.; Kato, E.H.; Morikawa, M.; Iwabuchi, K.; Nishida, R.; Kishi, R.; Onoé, K.; Minakami, H.; Yamada, H. No difference in natural killer or natural killer T-cell population, but aberrant T-helper cell population in the endometrium of women with repeated miscarriage. Hum. Reprod. 2004, 19, 1018–1024. [Google Scholar] [CrossRef]
- Michimata, T.; Ogasawara, M.S.; Tsuda, H.; Suzumori, K.; Aoki, K.; Sakai, M.; Fujimura, M.; Nagata, K.; Nakamura, M.; Saito, S. Distributions of endometrial NK cells, B cells, T cells, and Th2/Tc2 cells fail to predict pregnancy outcome following recurrent abortion. Am. J. Reprod. Immunol. 2002, 47, 196–202. [Google Scholar] [CrossRef]
- Chao, K.H.; Yang, Y.S.; Ho, H.N.; Chen, S.U.; Chen, H.F.; Dai, H.J.; Huang, S.C.; Gill, T.J. Decidual natural killer cytotoxicity decreased in normal pregnancy but not in anembryonic pregnancy and recurrent spontaneous abortion. Am. J. Reprod. Immunol. 1995, 34, 274–280. [Google Scholar] [CrossRef]
- Quack, K.C.; Vassiliadou, N.; Pudney, J.; Anderson, D.J.; Hill, J.A. Leukocyte activation in the decidua of chromosomally normal and abnormal fetuses from women with recurrent abortion. Hum. Reprod. 2001, 16, 949–955. [Google Scholar] [CrossRef]
- Emmer, P.M.; Steegers, E.A.; Kerstens, H.M.; Bulten, J.; Nelen, W.L.; Boer, K.; Joosten, I. Altered phenotype of HLA-G expressing trophoblast and decidual natural killer cells in pathological pregnancies. Hum. Reprod. 2002, 17, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hou, Y.; Zhang, S.; Zhou, Y.; Wang, D.; Tao, S.; Ni, F. CD49a Regulates the Function of Human Decidual Natural Killer Cells. Am. J. Reprod. Immunol. 2019, 81, e13101. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Fang, L.; Li, B.; Xiao, X.; Chen, S.; Wang, J.; Yang, F.; Chen, L.; Wang, X. Decreased Human Leukocyte Antigen-G Expression by miR-133a Contributes to Impairment of Proinvasion and Proangiogenesis Functions of Decidual NK Cells. Front. Immunol. 2017, 8, 741. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A. Immunomodulation in recurrent miscarriage. J. Obstet. Gynaecol. India 2014, 64, 165–168. [Google Scholar] [CrossRef]
- Quenby, S.; Kalumbi, C.; Bates, M.; Farquharson, R.; Vince, G. Prednisolone reduces preconceptual endometrial natural killer cells in women with recurrent miscarriage. Fertil. Steril. 2005, 84, 980–984. [Google Scholar] [CrossRef]
- Kuon, R.; Müller, F.; Vomstein, K.; Weber, M.; Hudalla, H.; Rösner, S.; Strowitzki, T.; Markert, U.; Daniel, V.; Toth, B. Pre-Pregnancy Levels of Peripheral Natural Killer Cells as Markers for Immunomodulatory Treatment in Patients with Recurrent Miscarriage. Arch. Immunol. Ther. Exp. 2017, 65, 339–346. [Google Scholar] [CrossRef]
- Shokri, M.; Bozorgmehr, M.; Ghanavatinejad, A.; Falak, R.; Aleahmad, M.; Kazemnejad, S.; Shokri, F.; Zarnani, A. Human menstrual blood-derived stromal/stem cells modulate functional features of natural killer cells. Sci. Rep. 2019, 9, 10007. [Google Scholar] [CrossRef]
- Filardi, T.; Varì, R.; Ferretti, E.; Zicari, A.; Morano, S.; Santangelo, C. Curcumin: Could This Compound Be Useful in Pregnancy and Pregnancy-Related Complications? Nutrients 2020, 12, 3179. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahajan, D.; Sharma, N.R.; Kancharla, S.; Kolli, P.; Tripathy, A.; Sharma, A.K.; Singh, S.; Kumar, S.; Mohanty, A.K.; Jena, M.K. Role of Natural Killer Cells during Pregnancy and Related Complications. Biomolecules 2022, 12, 68. https://doi.org/10.3390/biom12010068
Mahajan D, Sharma NR, Kancharla S, Kolli P, Tripathy A, Sharma AK, Singh S, Kumar S, Mohanty AK, Jena MK. Role of Natural Killer Cells during Pregnancy and Related Complications. Biomolecules. 2022; 12(1):68. https://doi.org/10.3390/biom12010068
Chicago/Turabian StyleMahajan, Deviyani, Neeta Raj Sharma, Sudhakar Kancharla, Prachetha Kolli, Ashutosh Tripathy, Amarish Kumar Sharma, Sanjeev Singh, Sudarshan Kumar, Ashok Kumar Mohanty, and Manoj Kumar Jena. 2022. "Role of Natural Killer Cells during Pregnancy and Related Complications" Biomolecules 12, no. 1: 68. https://doi.org/10.3390/biom12010068
APA StyleMahajan, D., Sharma, N. R., Kancharla, S., Kolli, P., Tripathy, A., Sharma, A. K., Singh, S., Kumar, S., Mohanty, A. K., & Jena, M. K. (2022). Role of Natural Killer Cells during Pregnancy and Related Complications. Biomolecules, 12(1), 68. https://doi.org/10.3390/biom12010068