
Novel Approaches to an Integrated Route for Trisomy 21 Evaluation

 , , , and
, , , and
Abstract
1. Introduction
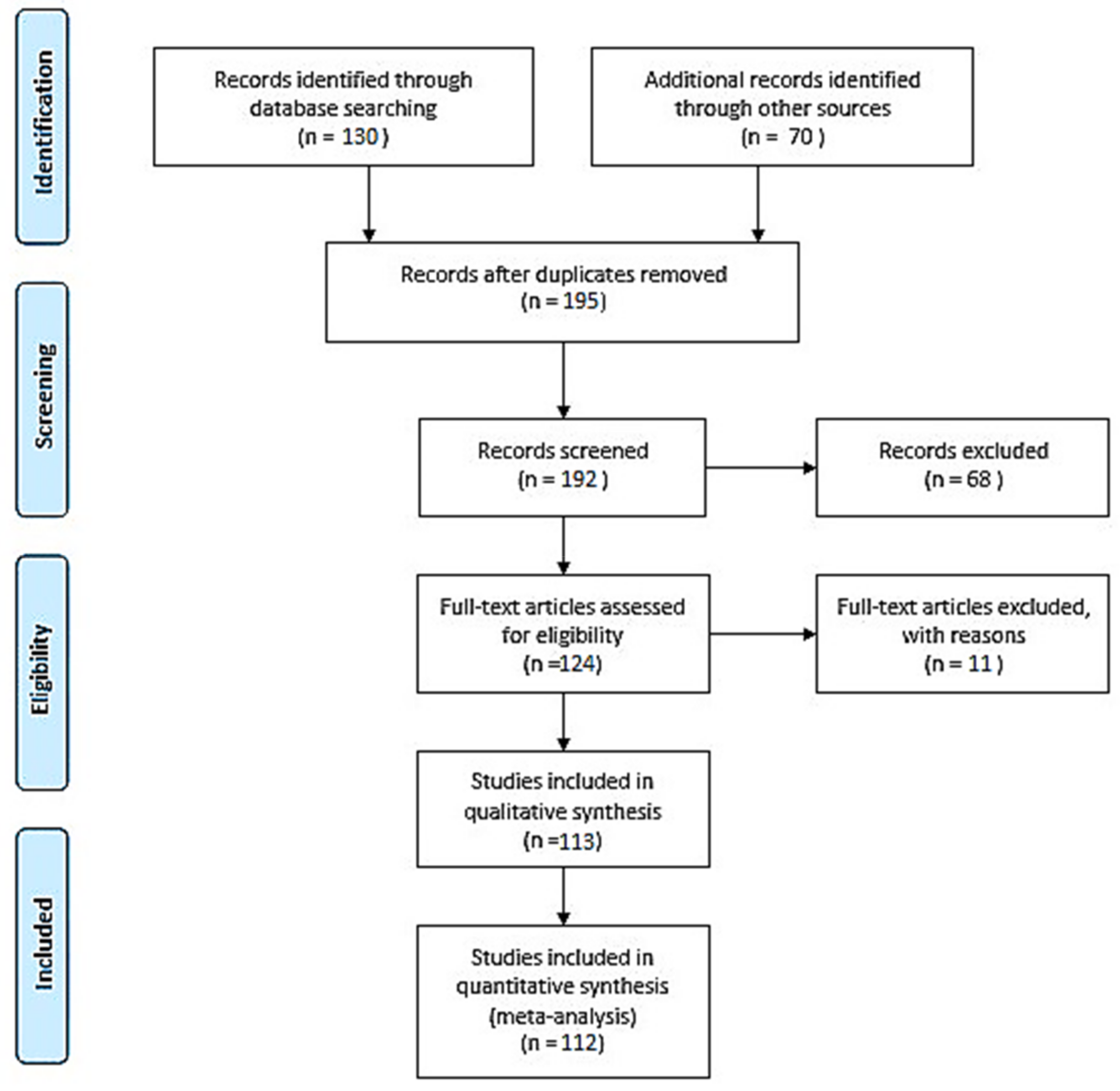
2. Materials and Methods
3. Current Recommendation for Down Syndrome Screening
4. Prenatal Genetic Diagnosis of Down Syndrome
5. Metabolomic Profiles as Down Syndrome Markers
6. Proteomics and Down Syndrome Screening
7. Single-Protein Determination
8. Discussion
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sherman, S.L.; Allen, E.G.; Bean, L.H.; Freeman, S.B. Epidemiology of Down syndrome. Ment. Retard. Dev. Disabil. Res. Rev. 2007, 13, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Malone, F. First-Trimester Sonographic Screening for Down Syndrome*1. Obstet. Gynecol. 2003, 102, 1066–1079. [Google Scholar] [CrossRef]
- Asim, A.; Kumar, A.; Muthuswamy, S.; Jain, S.; Agarwal, S. Down syndrome: An insight of the disease. J. Biomed. Sci. 2015, 22, 41. [Google Scholar] [CrossRef]
- Silverman, W. Down syndrome: Cognitive phenotype. Ment. Retard. Dev. Disabil. Res. Rev. 2007, 13, 228–236. [Google Scholar] [CrossRef]
- Foley, C.; Killeen, O.G. Musculoskeletal anomalies in children with Down syndrome: An observational study. Arch. Dis. Child. 2019, 104, 482–487. [Google Scholar] [CrossRef]
- Zbucka-Kretowska, M.; Charkiewicz, K.; Goscik, J.; Wolczynski, S.; Laudanski, P. Maternal plasma angiogenic and inflammatory factor profiling in foetal Down syndrome. PLoS ONE 2017, 12, e0189762. [Google Scholar] [CrossRef]
- Alldred, S.K.; Deeks, J.J.; Guo, B.; Neilson, J.P.; Alfirevic, Z. Second trimester serum tests for Down’s Syndrome screening. Cochrane Database Syst. Rev. 2012, 2012, cd009925. [Google Scholar] [CrossRef]
- Bianchi, D.W. Gene expression analysis of amniotic fluid: New biomarkers and novel antenatal treatments. Clin. Biochem. 2011, 44, 448–450. [Google Scholar] [CrossRef]
- Buczyńska, A.; Sidorkiewicz, I.; Ławicki, S.; Krętowski, A.; Zbucka-Krętowska, M. The Significance of Apolipoprotein E Measurement in the Screening of Fetal Down Syndrome. J. Clin. Med. 2020, 9, 3995. [Google Scholar] [CrossRef]
- Hutton, B.; Salanti, G.; Caldwell, D.M.; Chaimani, A.; Schmid, C.H.; Cameron, C.; Ioannidis, J.P.A.; Straus, S.; Thorlund, K.; Jansen, J.P.; et al. The PRISMA extension statement for reporting of systematic reviews incorporating network meta-analyses of health care interventions: Checklist and explanations. Ann. Intern. Med. 2015, 162, 777–784. [Google Scholar] [CrossRef]
- Calvert, M.; Blazeby, J.; Altman, D.G.; Revicki, D.A.; Moher, D.; Brundage, M.D. Reporting of patient-reported outcomes in randomized trials: The CONSORT PRO extension. JAMA—J. Am. Med. Assoc. 2013, 309, 814–822. [Google Scholar] [CrossRef]
- The EQUATOR Network|Enhancing the QUAlity and Transparency of Health Research. Available online: https://www.equator-network.org/ (accessed on 15 October 2020).
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Receiver operating characteristic (ROC) curve analysis for medical diagnostic test evaluation. Casp. J. Intern. Med. 2013, 4, 627–635. [Google Scholar]
- Nicolaides, K.H. Screening for fetal aneuploidies at 11 to 13 weeks. Prenat. Diagn. 2011, 31, 7–15. [Google Scholar] [CrossRef]
- Kitchen, F.L.; Jack, B.W. Prenatal Screening; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Wald, N.J.; Watt, H.C.; Hackshaw, A.K. Integrated screening for Down’s syndrome based on tests performed during the first and second trimesters. N. Engl. J. Med. 1999, 341, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Sparks, A.B.; Wang, E.T.; Struble, C.A.; Barrett, W.; Stokowski, R.; Mcbride, C.; Zahn, J.; Lee, K.; Shen, N.; Doshi, J.; et al. Selective analysis of cell-free DNA in maternal blood for evaluation of fetal trisomy. Prenat. Diagn. 2012, 32, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Cuckle, H. Biochemical screening for Down syndrome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2000, 92, 97–101. [Google Scholar] [CrossRef]
- Wiseman, F.K.; Alford, K.A.; Tybulewicz, V.L.J.; Fisher, E.M.C. Down syndrome-Recent progress and future prospects. Hum. Mol. Genet. 2009, 18, R75–R83. [Google Scholar] [CrossRef]
- Salman, M.S. Systematic review of the effect of therapeutic dietary supplements and drugs on cognitive function in subjects with Down syndrome. Eur. J. Paediatr. Neurol. 2002, 6, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, K.J. Pharmacological approaches to improving cognitive function in down syndrome: Current status and considerations. Drug Des. Devel. Ther. 2014, 9, 103–125. [Google Scholar] [CrossRef] [PubMed]
- KH, N. Nuchal translucency and other first-trimester sonographic markers of chromosomal abnormalities. Am. J. Obstet. Gynecol. 2004, 191, 45–67. [Google Scholar] [CrossRef]
- KH, N. A model for a new pyramid of prenatal care based on the 11 to 13 weeks’ assessment. Prenat. Diagn. 2011, 31, 3–6. [Google Scholar] [CrossRef]
- Durković, J.; Ubavić, M.; Durković, M.; Kis, T. Prenatal screening markers for down syndrome: Sensitivity, specificity, positive and negative expected value method. J. Med. Biochem. 2018, 37, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Norton, M.E.; Jacobsson, B.; Swamy, G.K.; Laurent, L.C.; Ranzini, A.C.; Brar, H.; Tomlinson, M.W.; Pereira, L.; Spitz, J.L.; Hollemon, D.; et al. Cell-free DNA analysis for noninvasive examination of trisomy. N. Engl. J. Med. 2015, 372, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Badeau, M.; Lindsay, C.; Blais, J.; Nshimyumukiza, L.; Takwoingi, Y.; Langlois, S.; Légaré, F.; Giguère, Y.; Turgeon, A.F.; Witteman, W.; et al. Genomics-based non-invasive prenatal testing for detection of fetal chromosomal aneuploidy in pregnant women. Cochrane Database Syst. Rev. 2017, 2017, CD011767. [Google Scholar] [CrossRef]
- De Castro, M. Johann gregor mendel: Paragon of experimental science. Mol. Genet. Genomic Med. 2016, 4, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A. The Human Genome; Wiley-Liss: New York, NY, USA, 2002. [Google Scholar]
- Jackson, M.; Marks, L.; May, G.H.W.; Wilson, J.B. The genetic basis of disease. Essays Biochem. 2018, 62, 643–723. [Google Scholar] [CrossRef]
- Steele, M.W.; Breg, W.R. Chromosome analysis of human amniotic-fluid cells. Lancet 1966, 1, 383–385. [Google Scholar] [CrossRef]
- Miron, P.M. Preparation, Culture, and Analysis of Amniotic Fluid Samples. Curr. Protoc. Hum. Genet. 2018, 98, e62. [Google Scholar] [CrossRef]
- Sánchez, O.; Domínguez, C.; Ruiz, A.; Ribera, I.; Alijotas, J.; Cabero, L.; Carreras, E.; Llurba, E. Angiogenic Gene Expression in Down Syndrome Fetal Hearts. Fetal Diagn. Ther. 2016, 40, 21–27. [Google Scholar] [CrossRef]
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nat. Rev. Dis. Prim. 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Olson, L.E.; Richtsmeier, J.T.; Leszl, J.; Reeves, R.H. A chromosome 21 critical region does not cause specific down syndrome phenotypes. Science 2004, 306, 687–690. [Google Scholar] [CrossRef]
- Samura, O. Update on noninvasive prenatal testing: A review based on current worldwide research. J. Obstet. Gynaecol. Res. 2020, 46, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.C.K.; Lo, Y.M.D. Prenatal diagnosis innovation: Genome sequencing of maternal plasma. Annu. Rev. Med. 2016, 67, 419–432. [Google Scholar] [CrossRef]
- Bianchi, D.W.; Wilkins-Haug, L. Integration of noninvasive DNA testing for aneuploidy into prenatal care: What has happened since the rubber met the road? Clin. Chem. 2014, 60, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Hatt, L.; Ravn, K.; Vogel, I.; Petersen, O.B.; Uldbjerg, N.; Schelde, P. Fetal cells in maternal blood for prenatal diagnosis: A love story rekindled. Biomark. Med. 2017, 11, 705–710. [Google Scholar] [CrossRef]
- Fiddler, M. Fetal Cell Based Prenatal Diagnosis: Perspectives on the Present and Future. J. Clin. Med. 2014, 3, 972. [Google Scholar] [CrossRef] [PubMed]
- Kølvraa, S.; Singh, R.; Normand, E.A.; Qdaisat, S.; van den Veyver, I.B.; Jackson, L.; Hatt, L.; Schelde, P.; Uldbjerg, N.; Vestergaard, E.M.; et al. Genome-wide copy number analysis on DNA from fetal cells isolated from the blood of pregnant women. Prenat. Diagn. 2016, 36, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Breman, A.M.; Chow, J.C.; U’Ren, L.; Normand, E.A.; Qdaisat, S.; Zhao, L.; Henke, D.M.; Chen, R.; Shaw, C.A.; Jackson, L.; et al. Evidence for feasibility of fetal trophoblastic cell-based noninvasive prenatal testing. Prenat. Diagn. 2016, 36, 1009–1019. [Google Scholar] [CrossRef]
- Vossaert, L.; Wang, Q.; Salman, R.; McCombs, A.K.; Patel, V.; Qu, C.; Mancini, M.A.; Edwards, D.P.; Malovannaya, A.; Liu, P.; et al. Validation Studies for Single Circulating Trophoblast Genetic Testing as a Form of Noninvasive Prenatal Diagnosis. Am. J. Hum. Genet. 2019, 105, 1262–1273. [Google Scholar] [CrossRef]
- Rezaei, M.; Winter, M.; Zander-Fox, D.; Whitehead, C.; Liebelt, J.; Warkiani, M.E.; Hardy, T.; Thierry, B. A Reappraisal of Circulating Fetal Cell Noninvasive Prenatal Testing. Trends Biotechnol. 2019, 37, 632–644. [Google Scholar] [CrossRef]
- Ma, G.-C.; Lin, W.-H.; Huang, C.-E.; Chang, T.-Y.; Liu, J.-Y.; Yang, Y.-J.; Lee, M.-H.; Wu, W.-J.; Chang, Y.-S.; Chen, M. A Silicon-based Coral-like Nanostructured Microfluidics to Isolate Rare Cells in Human Circulation: Validation by SK-BR-3 Cancer Cell Line and Its Utility in Circulating Fetal Nucleated Red Blood Cells. Micromachines 2019, 10, 132. [Google Scholar] [CrossRef]
- Huang, C.-E.; Ma, G.-C.; Jou, H.-J.; Lin, W.-H.; Lee, D.-J.; Lin, Y.-S.; Ginsberg, N.A.; Chen, H.-F.; Chang, F.M.-C.; Chen, M. Noninvasive prenatal diagnosis of fetal aneuploidy by circulating fetal nucleated red blood cells and extravillous trophoblasts using silicon-based nanostructured microfluidics. Mol. Cytogenet. 2017, 10, 44. [Google Scholar] [CrossRef]
- Cayrefourcq, L.; Vincent, M.-C.; Pierredon, S.; Moutou, C.; Imbert-Bouteille, M.; Haquet, E.; Puechberty, J.; Willems, M.; Liautard-Haag, C.; Molinari, N.; et al. Single Circulating Fetal Trophoblastic Cells Eligible for Non Invasive Prenatal Diagnosis: The Exception Rather than the Rule. Sci. Rep. 2020, 10, 9861. [Google Scholar] [CrossRef] [PubMed]
- Muskens, I.S.; Li, S.; Jackson, T.; Elliot, N.; Hansen, H.M.; Myint, S.S.; Pandey, P.; Schraw, J.M.; Roy, R.; Anguiano, J.; et al. The genome-wide impact of trisomy 21 on DNA methylation and its implications for hematopoiesis. Nat. Commun. 2021, 12, 821. [Google Scholar] [CrossRef]
- Lim, J.H.; Kang, Y.J.; Lee, B.Y.; Han, Y.J.; Chung, J.H.; Kim, M.Y.; Kim, M.H.; Kim, J.W.; Cho, Y.H.; Ryu, H.M. Epigenome-wide base-resolution profiling of DNA methylation in chorionic villi of fetuses with down syndrome by methyl-capture sequencing. Clin. Epigenet. 2019, 11, 180. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Shee, V. Genetic and Epigenetic Mechanisms in Down Syndrome Brain. In Down Syndrome; InTech: London, UK, 2013. [Google Scholar]
- Bacalini, M.G.; Gentilini, D.; Boattini, A.; Giampieri, E.; Pirazzini, C.; Giuliani, C.; Fontanesi, E.; Scurti, M.; Remondini, D.; Capri, M.; et al. Identification of a DNA methylation signature in blood cells from persons with down syndrome. Aging 2015, 7, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Zbucka-Kretowska, M.; Niemira, M.; Paczkowska-Abdulsalam, M.; Bielska, A.; Szalkowska, A.; Parfieniuk, E.; Ciborowski, M.; Wolczynski, S.; Kretowski, A. Prenatal circulating microRNA signatures of foetal Down syndrome. Sci. Rep. 2019, 9, 2394. [Google Scholar] [CrossRef] [PubMed]
- Burke, W. Genetic tests: Clinical validity and clinical utility. Curr. Protoc. Hum. Genet. 2014, 81, 9–15. [Google Scholar] [CrossRef]
- Song, K.; Musci, T.J.; Caughey, A.B. Clinical utility and cost of non-invasive prenatal testing with cfDNA analysis in high-risk women based on a US population. J. Matern. Neonatal Med. 2013, 26, 1180–1185. [Google Scholar] [CrossRef]
- Alexandrov, P.N.; Percy, M.E.; Lukiw, W.J. Chromosome 21-Encoded microRNAs (mRNAs): Impact on Down’s Syndrome and Trisomy-21 Linked Disease. Cell. Mol. Neurobiol. 2018, 38, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Gowda, G.A.N.; Zhang, S.; Gu, H.; Asiago, V.; Shanaiah, N.; Raftery, D. Metabolomics-based methods for early disease diagnostics. Expert Rev. Mol. Diagn. 2008, 8, 617–633. [Google Scholar] [CrossRef] [PubMed]
- Amberg, A.; Riefke, B.; Schlotterbeck, G.; Ross, A.; Senn, H.; Dieterle, F.; Keck, M. NMR and MS methods for metabolomics. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1641, pp. 229–258. [Google Scholar]
- Parfieniuk, E.; Zbucka-Kretowska, M.; Ciborowski, M.; Kretowski, A.; Barbas, C. Untargeted metabolomics: An overview of its usefulness and future potential in prenatal diagnosis. Expert Rev. Proteom. 2018, 15, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Parfieniuk, E.; Samczuk, P.; Kowalczyk, T.; Pietrowska, K.; Niemira, M.; Paczkowska-Abdulsalam, M.; Wolczynski, S.; Kretowski, A.; Ciborowski, M.; Zbucka-Kretowska, M. Maternal plasma metabolic fingerprint indicative for fetal Down syndrome. Prenat. Diagn. 2018, 38, 876–882. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Iles, R.K. Shotgun metabolomic profiles in maternal urine identify potential mass spectral markers of abnormal fetal biochemistry–dihydrouracil and progesterone in the metabolism of Down syndrome. Biomed. Chromatogr. 2015, 29, 1173–1183. [Google Scholar] [CrossRef]
- Charkiewicz, K.; Blachnio-Zabielska, A.; Zbucka-Kretowska, M.; Wolczynski, S.; Laudanski, P. Maternal plasma and amniotic fluid sphingolipids profiling in fetal down syndrome. PLoS ONE 2015, 10, e0127732. [Google Scholar] [CrossRef]
- Yang, W.; Chen, Y.H.; Liu, H.; Qu, H.D. Neuroprotective effects of piperine on the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson’s disease mouse model. Int. J. Mol. Med. 2015, 36, 1369–1376. [Google Scholar] [CrossRef]
- Parfieniuk, E.; Pietrowska, K.; Samczuk, P.; Kretowski, A.; Ciborowski, M.; Zbucka-Kretowska, M. Amniotic fluid metabolic fingerprinting indicated metabolites which may play a role in the pathogenesis of foetal Down syndrome—A preliminary report. Ginekol. Pol. 2021, 92, 188–194. [Google Scholar] [CrossRef]
- Nemutlu, E.; Orgul, G.; Recber, T.; Aydin, E.; Ozkan, E.; Turgal, M.; Alikasifoglu, M.; Kir, S.; Beksac, M.S. Metabolic Infrastructure of Pregnant Women with Trisomy 21 Fetuses; Metabolomic Analysis. Z. Geburtshilfe Neonatol. 2019, 223, 297–303. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Yan, G.; Wang, P.; Wang, X. Metabolomics for Biomarker Discovery: Moving to the Clinic. Biomed. Res. Int. 2015, 2015, 354671. [Google Scholar] [CrossRef]
- Shao, Y.; Le, W. Recent advances and perspectives of metabolomics-based investigations in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 3. [Google Scholar] [CrossRef]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and their applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef]
- Kang, Y.; Dong, X.; Zhou, Q.; Zhang, Y.; Cheng, Y.; Hu, R.; Su, C.; Jin, H.; Liu, X.; Ma, D.; et al. Identification of novel candidate maternal serum protein markers for Down syndrome by integrated proteomic and bioinformatic analysis. Prenat. Diagn. 2012, 32, 284–292. [Google Scholar] [CrossRef] [PubMed]
- López Uriarte, G.A.; Burciaga Flores, C.H.; Torres de la Cruz, V.M.; Medina Aguado, M.M.; Gómez Puente, V.M.; Romero Gutiérrez, L.N.; Martínez de Villarreal, L.E. Proteomic profile of serum of pregnant women carring a fetus with Down syndrome using nano uplc Q-tof ms/ms technology. J. Matern. Neonatal Med. 2018, 31, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Charkiewicz, K.; Zbucka-Kretowska, M.; Goscik, J.; Wolczynski, S.; Lemancewicz, A.; Laudanski, P. Brief communication: Maternal plasma autoantibodies screening in fetal down syndrome. J. Immunol. Res. 2016, 2016, 9362169. [Google Scholar] [CrossRef] [PubMed]
- Laudanski, P.; Zbucka-Kretowska, M.; Charkiewicz, K.; Wolczynski, S.; Wojcik, D.; Charkiewicz, R. Maternal plasma and amniotic fluid chemokines screening in fetal down syndrome. Mediat. Inflamm. 2014, 2014, 835837. [Google Scholar] [CrossRef] [PubMed]
- Zbucka-Kretowska, M.; Charkiewicz, K.; Czerniecki, J.; Goscik, J.; Wolczynski, S.; Laudanski, P. Amniotic Fluid Angiogenic and Inflammatory Factor Profiling in Foetal Down Syndrome. Fetal Diagn. Ther. 2018, 44, 44–50. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Karaca, E.; Aykut, A.; Ertürk, B.; Durmaz, B.; Güler, A.; Büke, B.; Yeniel, A.Ö.; Ergenoğlu, A.M.; Özkınay, F.; Özeren, M.; et al. Microrna expression profile in the prenatal amniotic fluid samples of pregnant women with down syndrome. Balkan Med. J. 2018, 35, 163–166. [Google Scholar] [CrossRef]
- Ahlfors, H.; Anyanwu, N.; Pakanavicius, E.; Dinischiotu, N.; Lana-Elola, E.; Watson-Scales, S.; Tosh, J.; Wiseman, F.; Briscoe, J.; Page, K.; et al. Gene expression dysregulation domains are not a specific feature of Down syndrome. Nat. Commun. 2019, 10, 2489. [Google Scholar] [CrossRef]
- Lana-Elola, E.; Watson-Scales, S.D.; Fisher, E.M.C.; Tybulewicz, V.L.J. Down syndrome: Searching for the genetic culprits. DMM Dis. Model. Mech. 2011, 4, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.; Fisher, E.M.C. Down syndrome genetics: Unravelling a multifactorial disorder. Hum. Mol. Genet. 1996, 5, 1411–1416. [Google Scholar] [CrossRef]
- Galambos, C.; Minic, A.D.; Bush, D.; Nguyen, D.; Dodson, B.; Seedorf, G.; Abman, S.H. Increased lung expression of anti-angiogenic factors in Down syndrome: Potential role in abnormal lung vascular growth and the risk for pulmonary hypertension. PLoS ONE 2016, 11, e0159005. [Google Scholar] [CrossRef] [PubMed]
- Kolialexi, A.; Tsangaris, G.T.; Papantoniou, N.; Anagnostopoulos, A.K.; Vougas, K.K.; Bagiokos, V.; Antsaklis, A.; Mavrou, A. Application of proteomics for the identification of differentially expressed protein markers for Down syndrome in maternal plasma. Prenat. Diagn. 2008, 28, 691–698. [Google Scholar] [CrossRef]
- Anagnostopoulos, A.; Th Tsangaris, G. Serum amyloid-p (SAP), a potential biomarker for Down syndrome fetuses prevention in maternal plasma. EPMA J. 2014, 5, A98. [Google Scholar] [CrossRef]
- Stoltzner, S.E.; Grenfell, T.J.; Mori, C.; Wisniewski, K.E.; Wisniewski, T.M.; Selkoe, D.J.; Lemere, C.A. Temporal accrual of complement proteins in amyloid plaques in Down’s syndrome with Alzheimer’s disease. Am. J. Pathol. 2000, 156, 489–499. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The Role of Apolipoprotein E in Alzheimer’s Disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Sui, W.; Gan, Q.; Gong, W.W.; Wei, X.; Ou, M.; Tang, D.; Jing, H.; Lin, H.; Zhang, Y.; Dai, Y. Verification of foetal Down syndrome biomarker proteins in maternal plasma and applications in prenatal screening for Down syndrome. Transl. Med. Commun. 2018, 3, 9. [Google Scholar] [CrossRef]
- Strohmeyer, R.; Shen, Y.; Rogers, J. Detection of complement alternative pathway mRNA and proteins in the Alzheimer’s disease brain. Mol. Brain Res. 2000, 81, 7–18. [Google Scholar] [CrossRef]
- Tyers, M.; Mann, M. From genomics to proteomics. Nature 2003, 422, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Betzen, C.; Alhamdani, M.S.S.; Lueong, S.; Schröder, C.; Stang, A.; Hoheisel, J.D. Clinical proteomics: Promises, challenges and limitations of affinity arrays. PROTEOMICS—Clin. Appl. 2015, 9, 342–347. [Google Scholar] [CrossRef]
- Alhajj, M.; Farhana, A. Enzyme Linked Immunosorbent Assay; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Sakamoto, S.; Putalun, W.; Vimolmangkang, S.; Phoolcharoen, W.; Shoyama, Y.; Tanaka, H.; Morimoto, S. Enzyme-linked immunosorbent assay for the quantitative/qualitative analysis of plant secondary metabolites. J. Nat. Med. 2018, 72, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zheng, Y.; Kang, X.; Zhang, X.; Hao, H.; Chen, W.; Liu, L.; Li, X.; Li, L.; Yuan, Q.; et al. A multiplex ELISA-based protein array for screening diagnostic antigens and diagnosis of Flaviviridae infection. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Van Gool, A.; Corrales, F.; Čolović, M.; Krstić, D.; Oliver-Martos, B.; Martínez-Cáceres, E.; Jakasa, I.; Gajski, G.; Brun, V.; Kyriacou, K.; et al. Analytical techniques for multiplex analysis of protein biomarkers. Expert Rev. Proteom. 2020, 17, 257–273. [Google Scholar] [CrossRef]
- Snoj Tratnik, J.; Falnoga, I.; Trdin, A.; Mazej, D.; Fajon, V.; Miklavčič, A.; Kobal, A.B.; Osredkar, J.; Sešek Briški, A.; Krsnik, M.; et al. Prenatal mercury exposure, neurodevelopment and apolipoprotein E genetic polymorphism. Environ. Res. 2017, 152, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, P.; Ghosh, P.; Ghosh, S.; Feingold, E.; Ozbek, U.; Sarkar, B.; Dey, S.K. Combined association of presenilin-1 and apolipoprotein E polymorphisms with maternal meiosis II error in down syndrome births. Genet. Mol. Biol. 2017, 40, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Day, R.J.; McCarty, K.L.; Ockerse, K.E.; Head, E.; Rohn, T.T. Proteolytic cleavage of apolipoprotein e in the down syndrome brain. Aging Dis. 2016, 7, 267–277. [Google Scholar] [CrossRef]
- D’Souza, H.; Mason, L.; Mok, K.Y.; Startin, C.M.; Hamburg, S.; Hithersay, R.; Baksh, R.A.; Hardy, J.; Strydom, A.; Thomas, M.S.C. Differential Associations of Apolipoprotein E ε4 Genotype with Attentional Abilities Across the Life Span of Individuals With Down Syndrome. JAMA Netw. Open 2020, 3, e2018221. [Google Scholar] [CrossRef]
- Perluigi, M.; Butterfield, D.A. The identification of protein biomarkers for oxidative stress in Down syndrome. Expert Rev. Proteom. 2011, 8, 427–429. [Google Scholar] [CrossRef][Green Version]
- Barone, E.; Head, E.; Butterfield, D.A.; Perluigi, M. HNE-modified proteins in Down syndrome: Involvement in development of Alzheimer disease neuropathology. Free Radic. Biol. Med. 2017, 111, 262–269. [Google Scholar] [CrossRef]
- Barone, E.; Arena, A.; Head, E.; Butterfield, D.A.; Perluigi, M. Disturbance of redox homeostasis in Down Syndrome: Role of iron dysmetabolism. Free Radic. Biol. Med. 2018, 114, 84–93. [Google Scholar] [CrossRef]
- Perrone, S.; Longini, M.; Bellieni, C.V.; Centini, G.; Kenanidis, A.; De Marco, L.; Petraglia, F.; Buonocore, G. Early oxidative stress in amniotic fluid of pregnancies with Down syndrome. Clin. Biochem. 2007, 40, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Muchová, J.; Žitňanová, I.; Ďuračková, Z. Oxidative stress and Down syndrome. do antioxidants play a role in therapy? Physiol. Res. 2014, 63, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Yin, Y.X.; Ding, J.; Yuan, H.; Yang, L.; Xu, J.J.; Hu, L.Q. Alpha-1-Antitrypsin suppresses oxidative stress in preeclampsia by inhibiting the p38MAPK signaling pathway: An in vivo and in vitro study. PLoS ONE 2017, 12, e0173711. [Google Scholar] [CrossRef]
- Buczyńska, A.; Sidorkiewicz, I.; Ławicki, S.; Krętowski, A.J.; Zbucka-Krętowska, M. Prenatal Screening of Trisomy 21: Could Oxidative Stress Markers Play a Role? J. Clin. Med. 2021, 10, 2382. [Google Scholar] [CrossRef]
- Van den Veyver, I.B. Recent advances in prenatal genetic screening and testing. F1000Research 2016, 5, 2591. [Google Scholar] [CrossRef] [PubMed]
- Pös, O.; Budiš, J.; Szemes, T. Recent trends in prenatal genetic screening and testing. F1000Research 2019, 8, 764. [Google Scholar] [CrossRef]
- Kazemi, M.; Salehi, M.; Kheirollahi, M. Down syndrome: Current status, challenges and future perspectives. Int. J. Mol. Cell. Med. 2016, 5, 125–133. [Google Scholar] [PubMed]
- Corella, D.; Ordovas, J.M. The role of omics in precision nutrition: Strengths and weaknesses. Nutr. Hosp. 2018, 35, 10–18. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- O’Connell, E.; Hurley, F. A review of the strengths and weaknesses of quantitative methods used in health impact assessment. Public Health 2009, 123, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Sachs, M.C. Statistical principles for omics-based clinical trials. Chin. Clin. Oncol. 2015, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- JA, R. Down syndrome: A curative prospect? AIMS Neurosci. 2020, 7, 168–193. [Google Scholar] [CrossRef]
- Morello, G.; Salomone, S.; D’Agata, V.; Conforti, F.L.; Cavallaro, S. From Multi-Omics Approaches to Precision Medicine in Amyotrophic Lateral Sclerosis. Front. Neurosci. 2020, 14, 577755. [Google Scholar] [CrossRef]
- Guedj, F.; Bianchi, D.W.; Delabar, J.M. Prenatal treatment of Down syndrome: A reality? Curr. Opin. Obstet. Gynecol. 2014, 26, 92–103. [Google Scholar] [CrossRef]


{kind=link}
| Pregnancy Period | Ultrasound | Biochemical Test | Sensitivity | Specificity |
|---|---|---|---|---|
| First trimester (11–13 weeks) | + (NT) | PAPP-A and free βhCG | 85–90% | 82–87% |
| Second trimester (18–24 weeks) | + | βhCG + uE3 + AFP + inhibin A | 69–92% | 81–96% |
| First or second trimester | + | PAPP-A, AFP, uE3, total hCG | 88% | 90–95% |
| First or second trimester | + | PAPP-A, inhibin A, AFP, uE3, free βhCG/total hCG | 85% | 90–95% |
| Biological Sample | Significant Deregulated Metabolites in T21 Prenatal Screening |
|---|---|
| maternal blood | 2-hydroxybutyrate, alanine, citric acid, phenylalanine, 3-methyl histidine, proline, benzoic acid, glyceric acid, mannose, myristic acid, stearic acid |
| maternal serum | 2-hydroxybutyrate, 3- hydroxybutyrate, acetone, glycerol, glycine, isobutyrate, ornithine, phenylalanine, succinate, methylhistidine, arginine, 12-hydroxybutyrate, carnitine, lactate, pyruvate, dimethylamine, methionine |
| maternal plasma | butyryl-l-carnitine, palmitic amide, linoleamide, oleamide, piperine, proline, methanol, creatinine |
| maternal urine | dihydrouracil, methanol, β-hydroxybutyrate |
| amniotic fluid | methylhistidine, hexanoylcarnitine, diacetylspermine, and p-cresol sulfate |
| T21 Screening Panel | AUC Based on ROC Curves |
|---|---|
| βhCG + PAPP-A | 0.918 |
| PAPP-A + NT | 0.922 |
| PAPP-A + hCG-sLHCGR | 0.920 |
| βhCG + hCG-sLHCGR | 0.856 |
| βhCG + NT | 0.753 |
| hCG-sLHCGR + NT | 0.888 |
| NT + PAPP-A + βhCG | 0.940 |
| NT + PAPP-A + hCG-sLHCGR | 0.928 |
| hCGsLHCGR + NT + PAPP-A + βhCG | 0.966 |
| Marker | Unit | AUC | Cut-Off Value | Sensitivity | Specificity |
|---|---|---|---|---|---|
| ApoE | ng/mL | 0.978 | >1.37 | 80% | 100% |
| Asprosin PS | ng/mL | 0.970 | >12.70 | 100% | 85% |
| Asprosin AF | ng/mL | 0.830 | >12.91 | 95% | 65% |
| AGE PS | ng/mL | 0.850 | <11.00 | 81% | 80% |
| AGE AF | ng/mL | 0.960 | <4.184 | 95% | 90% |
| A1AT PS | mg/L | 0.530 | <2.341 | 81% | 33% |
| A1AT AF | mg/L | 0.870 | <0.3180 | 76% | 86% |
| DNA/RNA OSDP PS | pg/mL | 0.510 | <40.30 | 80% | 40% |
| DNA/RNA OSDP AF | pg/mL | 0.730 | >31.76 | 84% | 58% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buczyńska, A.; Sidorkiewicz, I.; Trochimiuk, A.; Ławicki, S.; Krętowski, A.J.; Zbucka-Krętowska, M. Novel Approaches to an Integrated Route for Trisomy 21 Evaluation. Biomolecules 2021, 11, 1328. https://doi.org/10.3390/biom11091328
Buczyńska A, Sidorkiewicz I, Trochimiuk A, Ławicki S, Krętowski AJ, Zbucka-Krętowska M. Novel Approaches to an Integrated Route for Trisomy 21 Evaluation. Biomolecules. 2021; 11(9):1328. https://doi.org/10.3390/biom11091328
Chicago/Turabian StyleBuczyńska, Angelika, Iwona Sidorkiewicz, Anna Trochimiuk, Sławomir Ławicki, Adam Jacek Krętowski, and Monika Zbucka-Krętowska. 2021. "Novel Approaches to an Integrated Route for Trisomy 21 Evaluation" Biomolecules 11, no. 9: 1328. https://doi.org/10.3390/biom11091328
APA StyleBuczyńska, A., Sidorkiewicz, I., Trochimiuk, A., Ławicki, S., Krętowski, A. J., & Zbucka-Krętowska, M. (2021). Novel Approaches to an Integrated Route for Trisomy 21 Evaluation. Biomolecules, 11(9), 1328. https://doi.org/10.3390/biom11091328