
A New Pentafluorothio-Substituted Curcuminoid with Superior Antitumor Activity

 ,
,  , , , ,
, , , ,  and
and 
Abstract
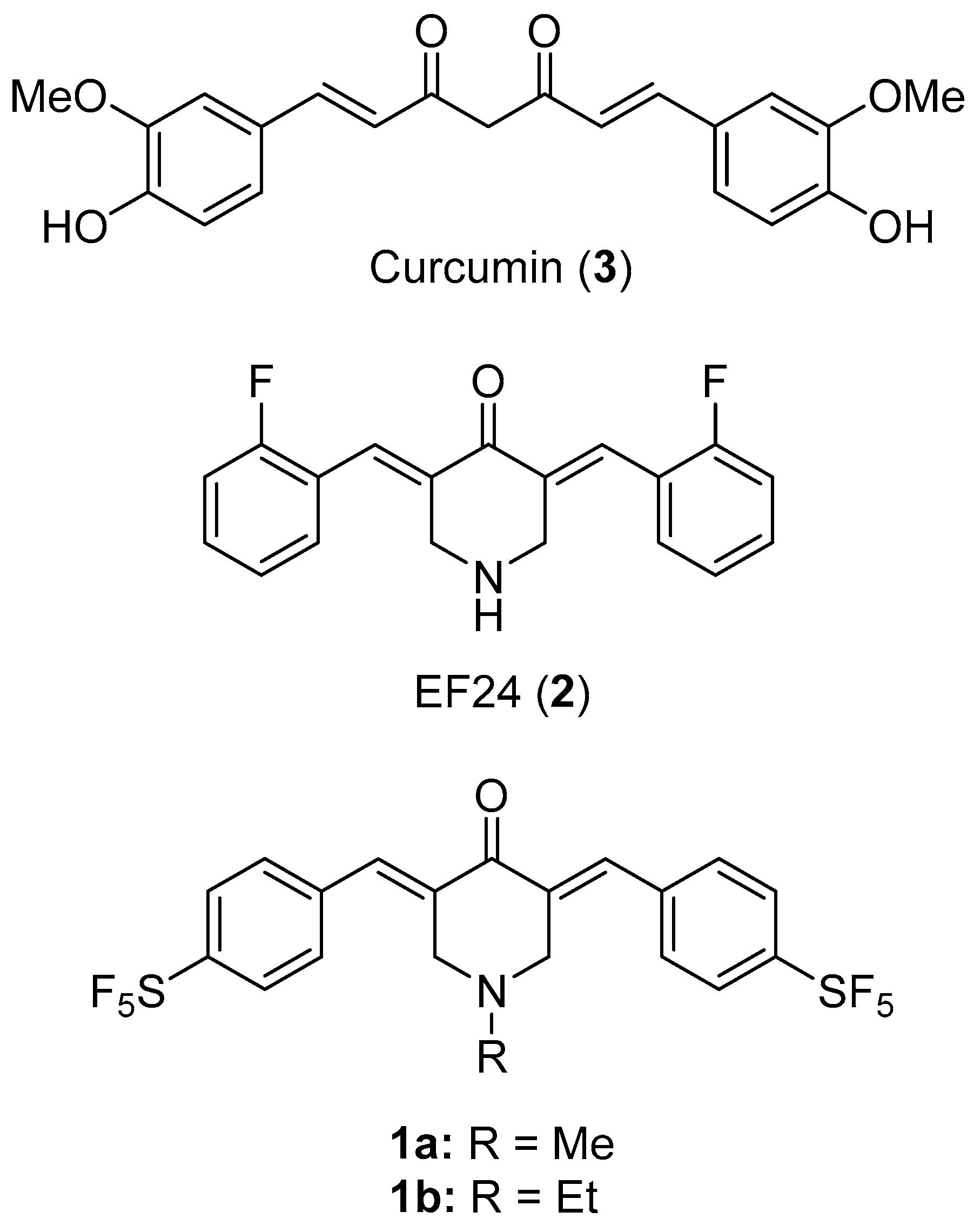
:1. Introduction
2. Materials and Methods
2.1. Chemistry
2.1.1. General
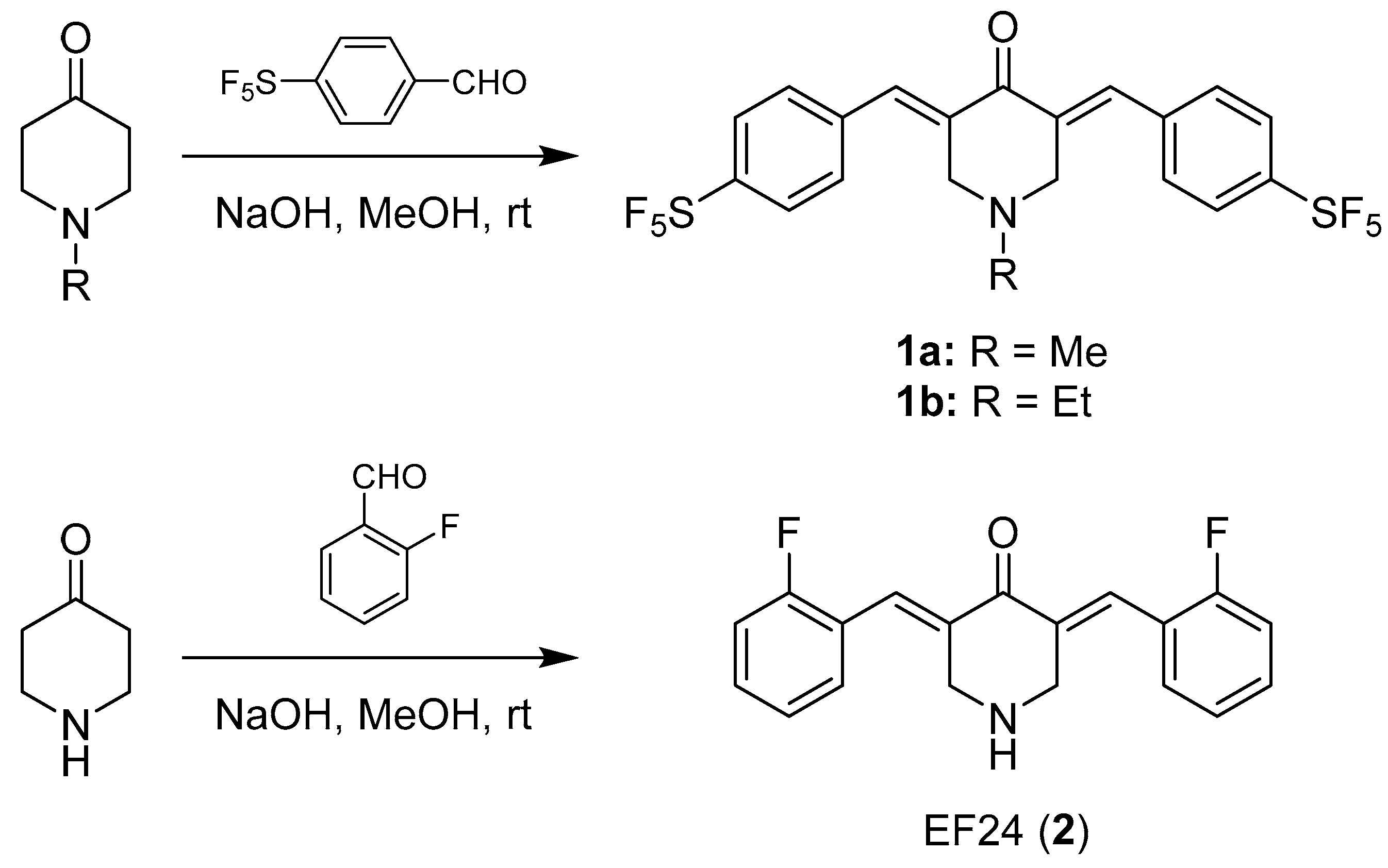
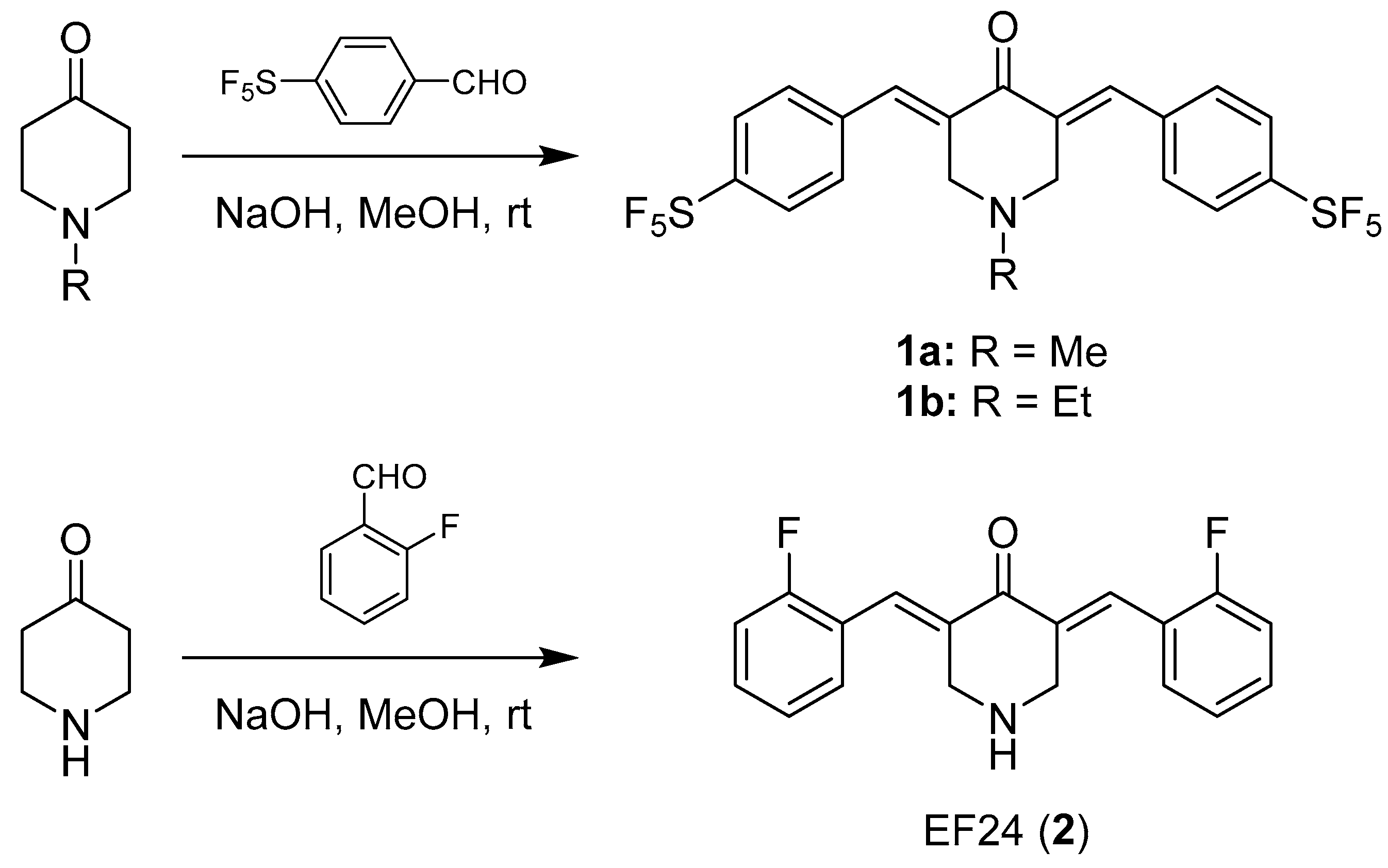
2.1.2. (E)-1-Methyl-3,5-bis(4-pentafluorothiobenzylidene)-4-piperidone (1a)
2.2. Biological Evaluation
2.2.1. Cell Culture Conditions
2.2.2. MTT Assay
2.2.3. Hexoseaminidase Enzyme Assay
2.2.4. Zebrafish Angiogenesis Assay
2.2.5. Caspase-3/7 Activity Assay
2.2.6. DCFH-DA Assay
2.2.7. Cell-Cycle Analysis
2.2.8. Immunofluorescence Staining of Actin Cytoskeleton
2.2.9. Limiting Dilution Assay
2.2.10. Measuring Apoptosis-Induction Using Flow Cytometry
3. Results
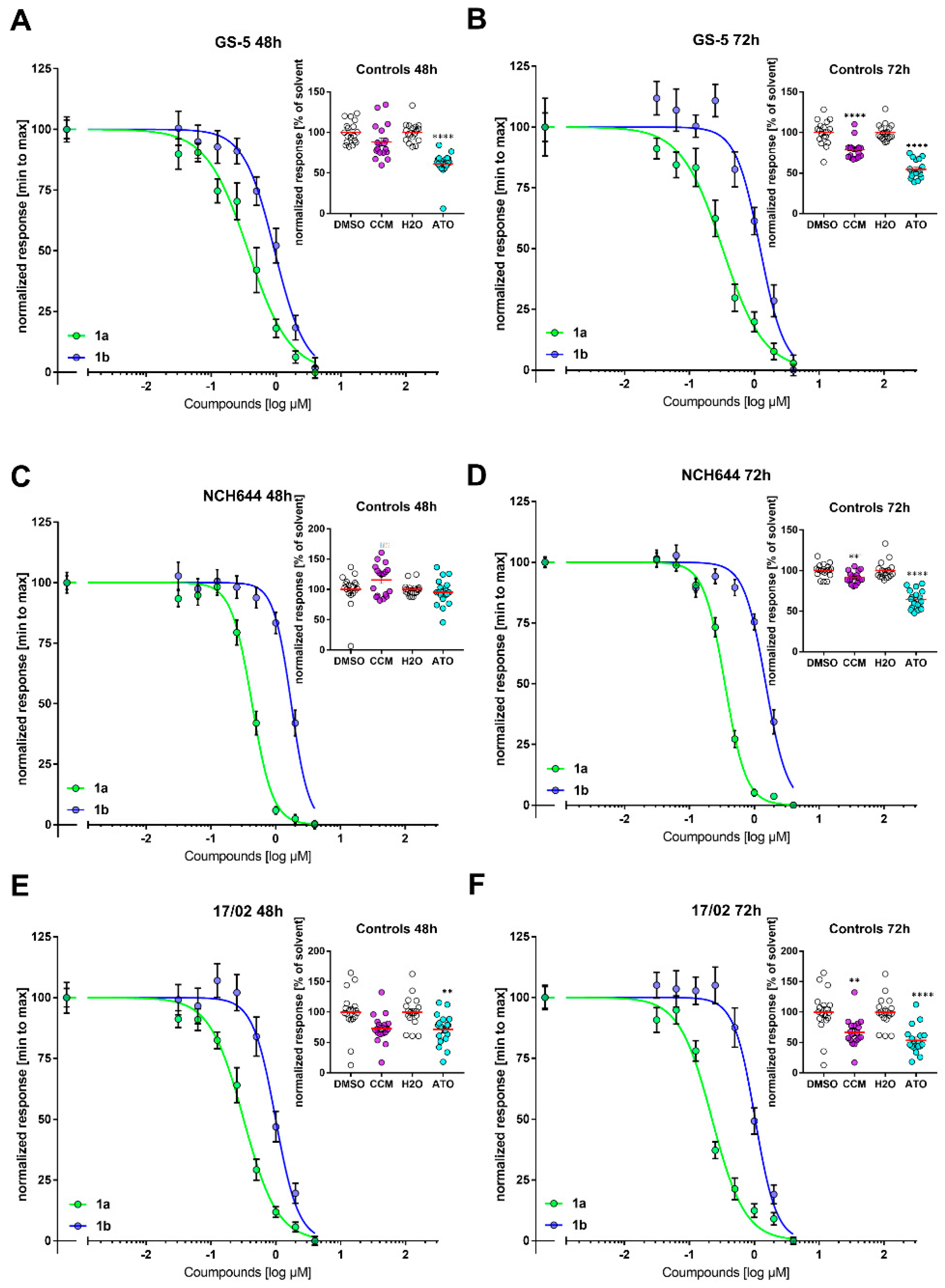
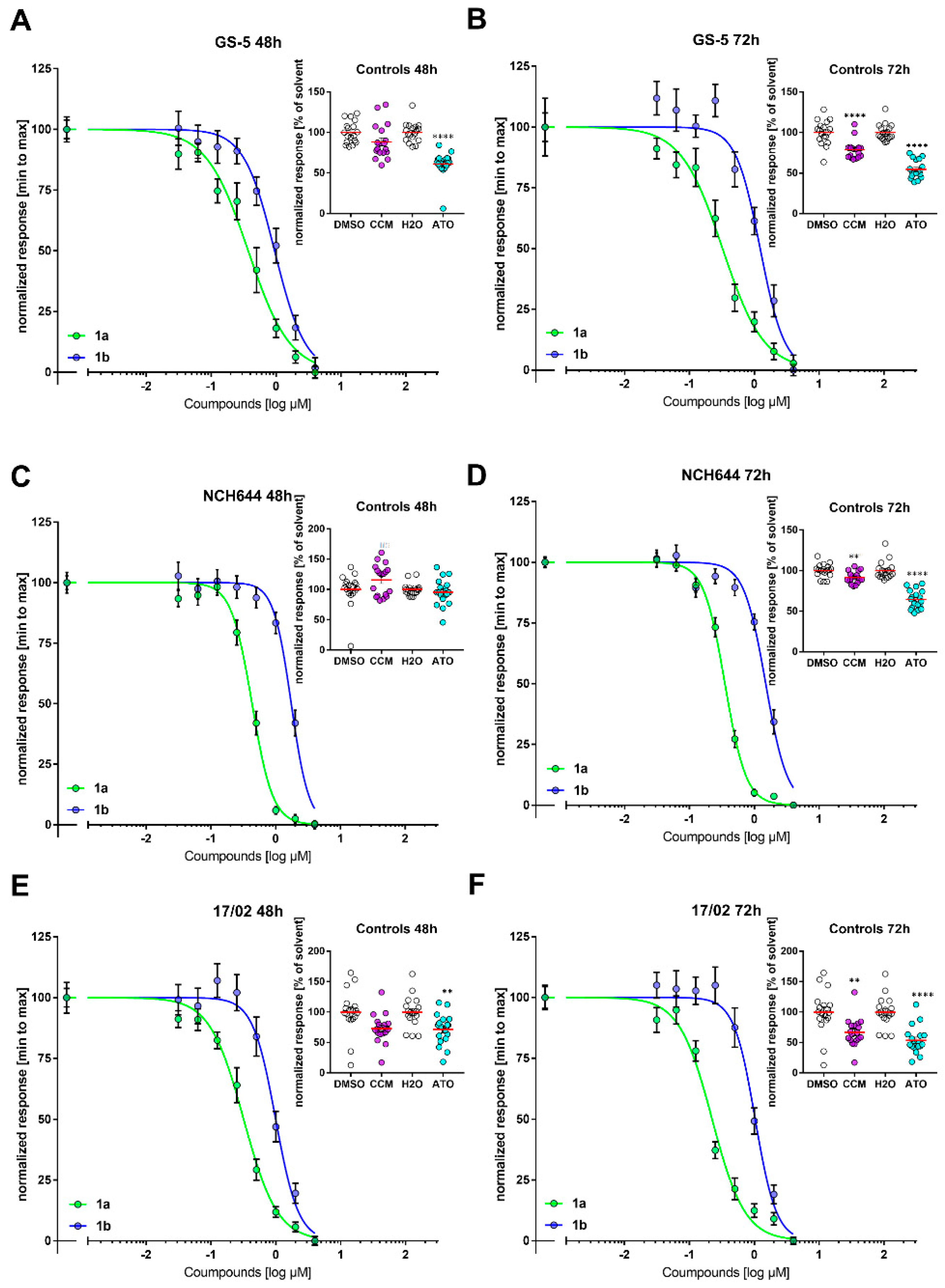
3.1. Antiproliferative Activity
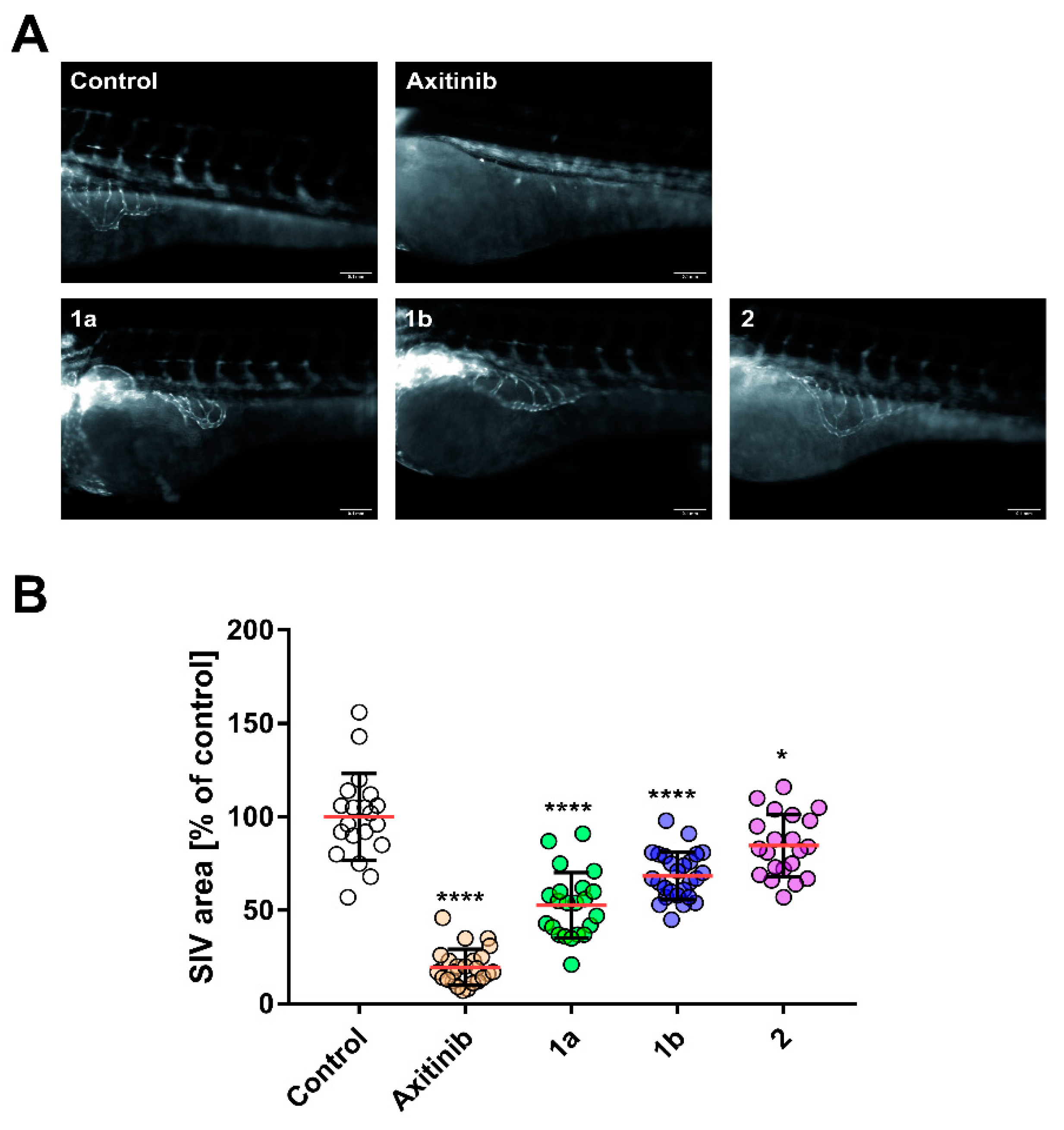
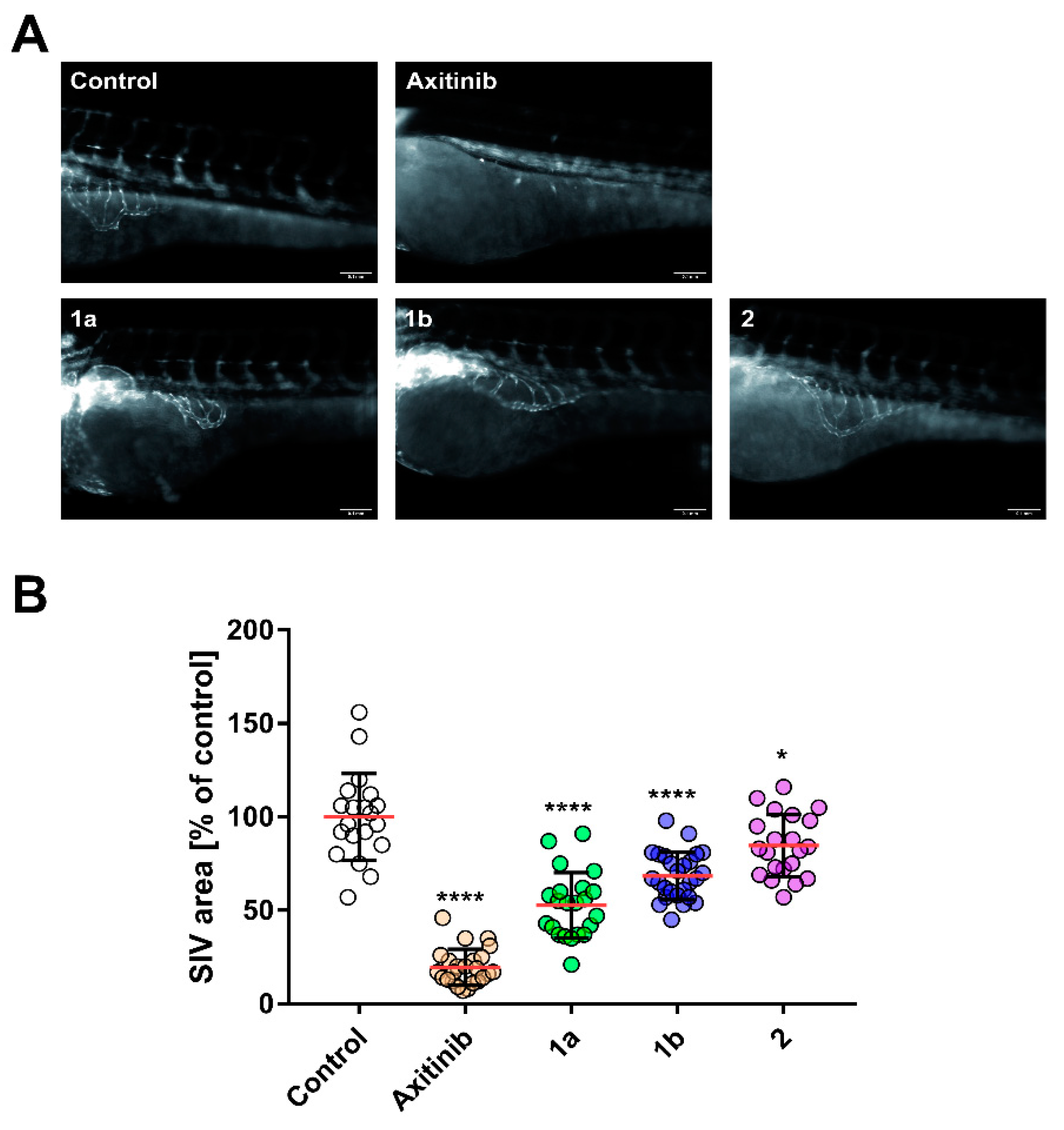
3.2. Anti-Angiogenic Effect
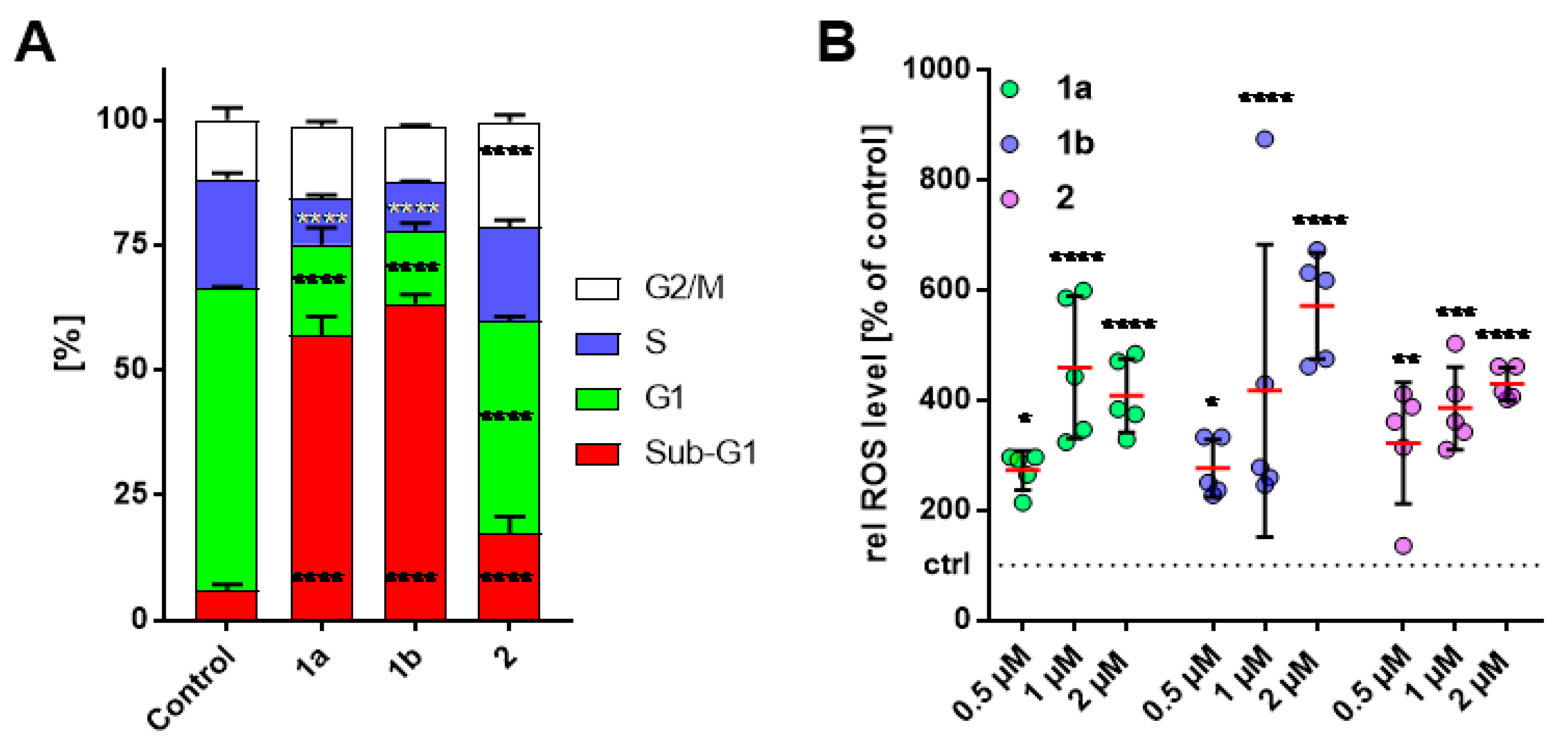
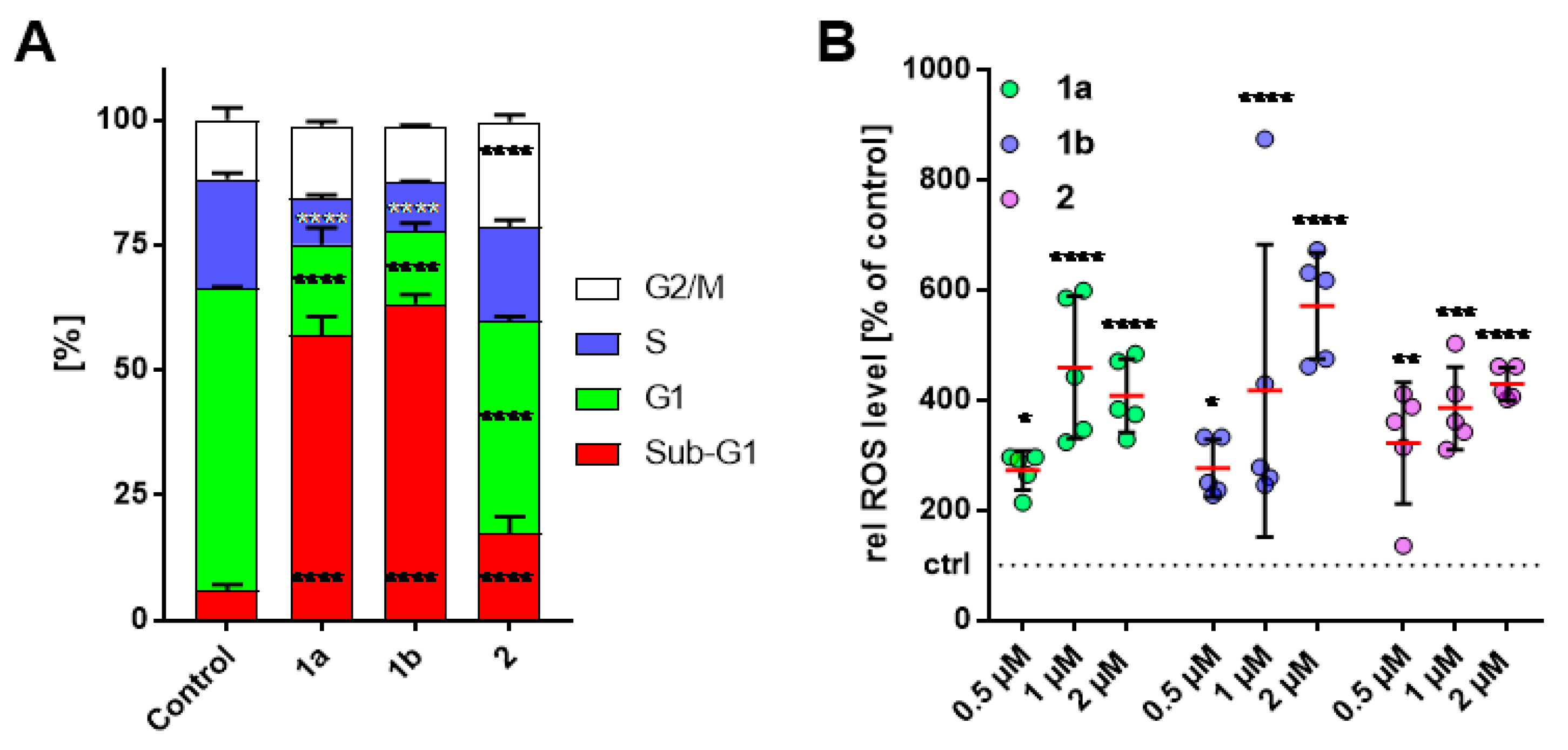
3.3. Alteration in Cell-Cycle Progression of HT-29 Cells
3.4. Increase of Intracellular ROS Levels in HT-29 Cells
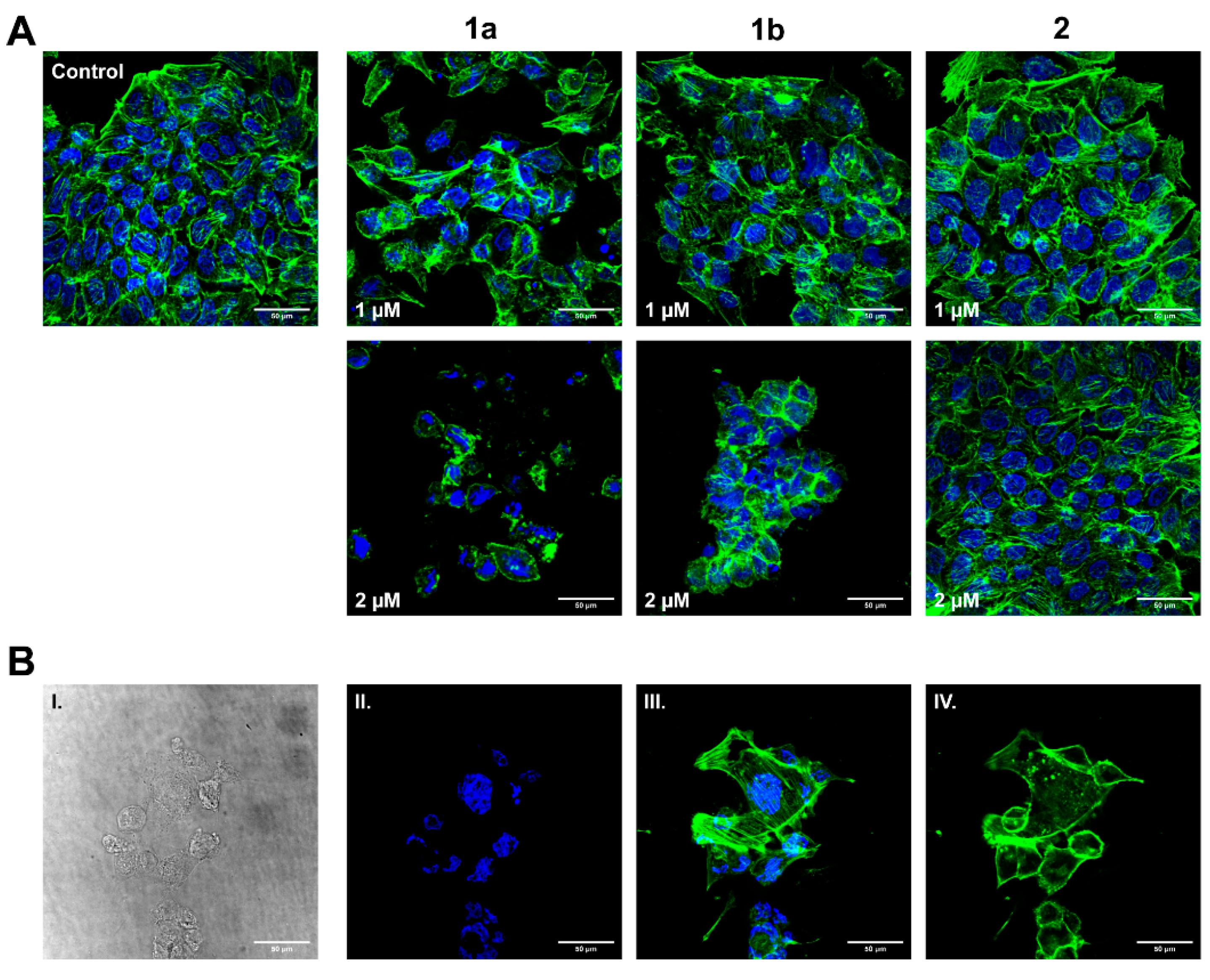
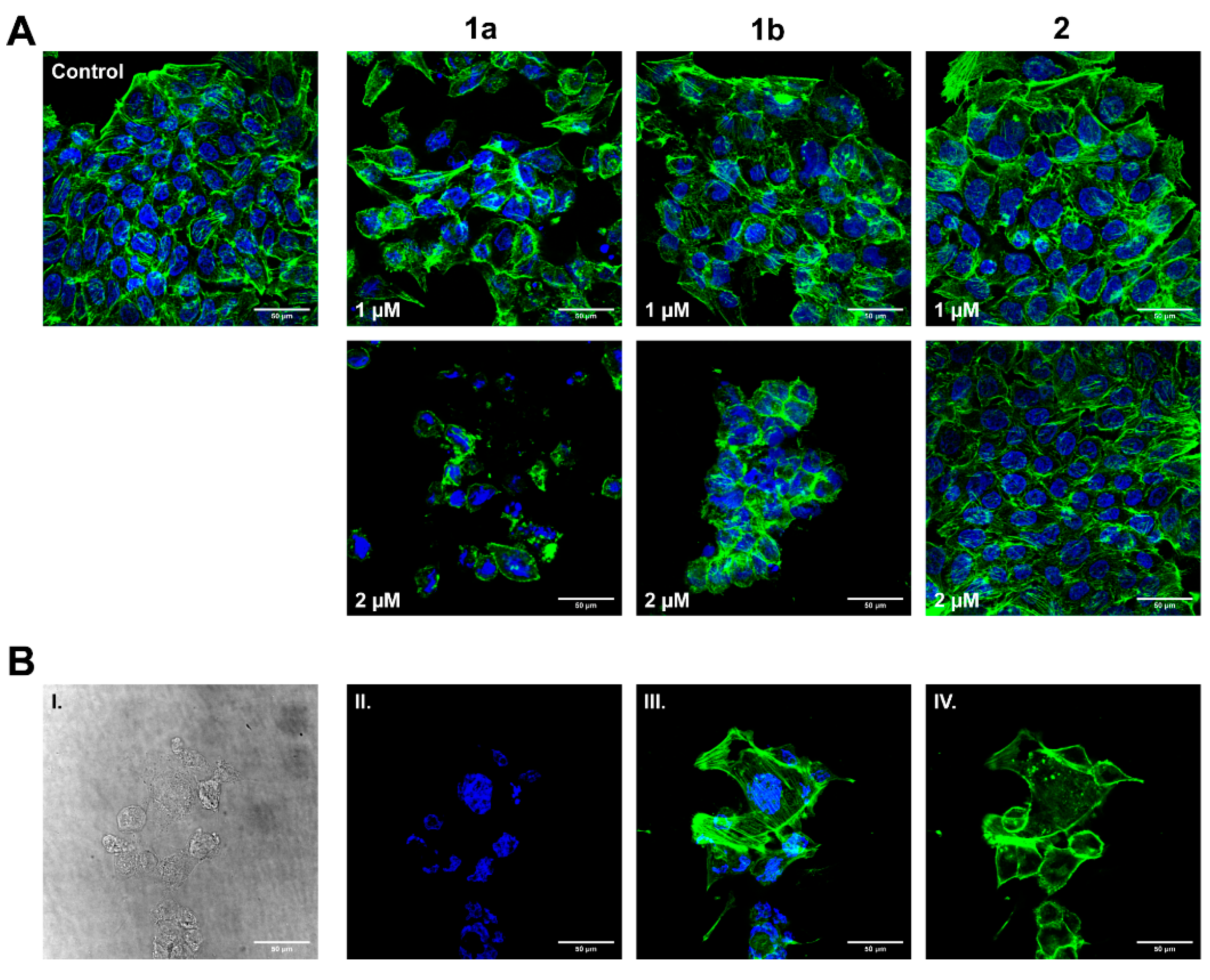
3.5. Influence on HT-29 Cell Morphology and Actin Cytoskeleton
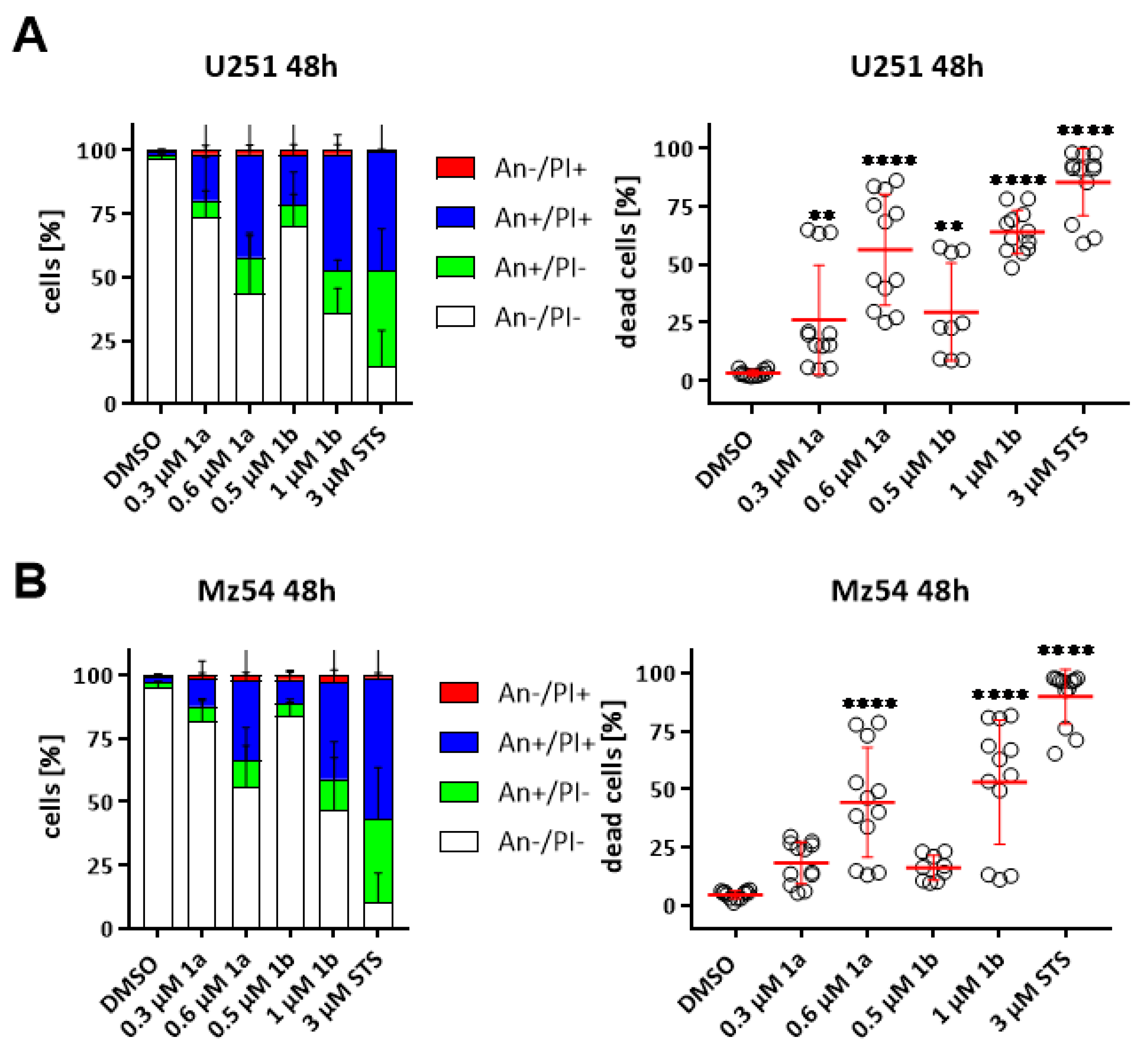
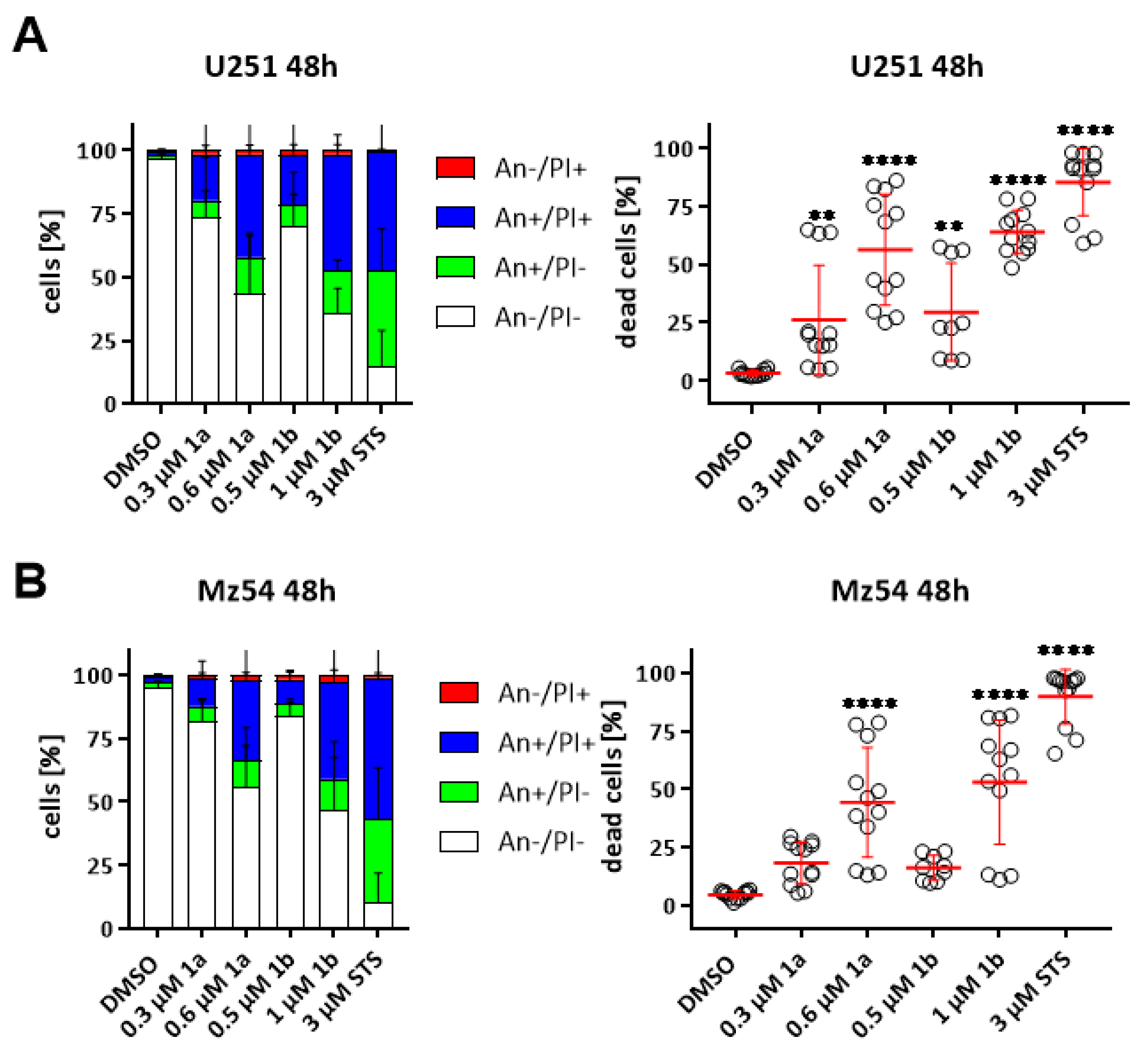
3.6. Induction of Cell Death in GBM Cell Lines
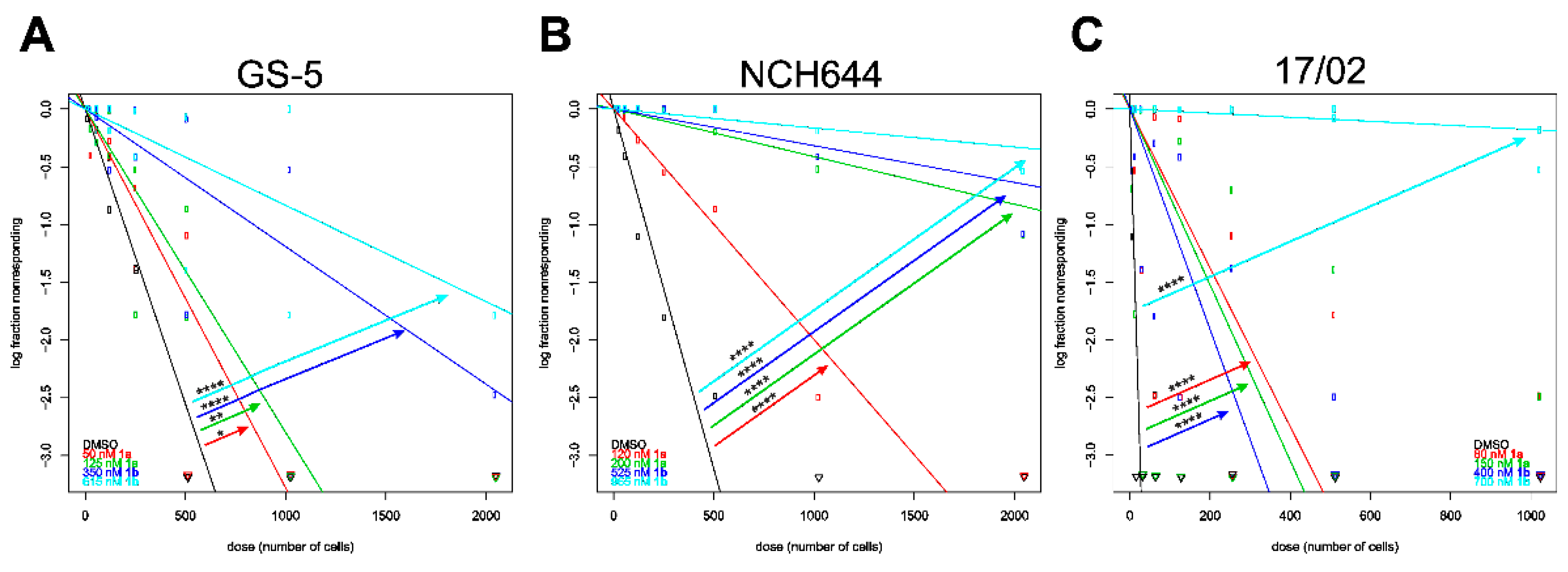
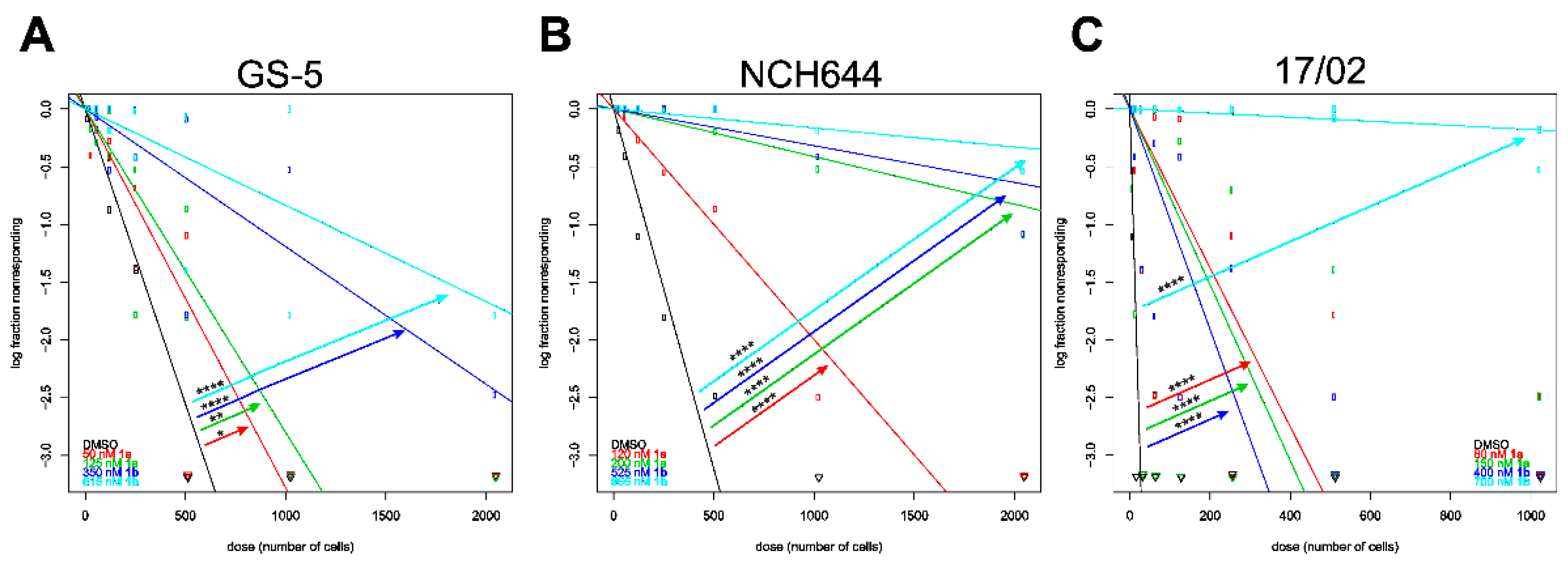
3.7. Stemness Decrease of GSCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, R.A.; McLelland, H.R.; Hill, K.A.; Ireson, C.R.; Euden, S.A.; Manson, M.M.; Pirmohamed, M.; Marnett, L.J.; Gescher, A.J.; Steward, W.P. Pharmacodynamic and pharmacokinetic study of oral Curcuma extract in patients with colorectal cancer. Clin. Cancer Res. 2001, 7, 1894–1900. [Google Scholar]
- Pan, M.-H.; Huang, T.M.; Lin, J.K. Biotransformation of curcumin through reduction and glucuronidation in mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar]
- Thayyullathil, F.; Chathoth, S.; Hago, A.; Patel, M.; Galadari, S. Rapid reactive oxygen species (ROS) generation induced by curcumin leads to caspase-dependent and -independent apoptosis in L929 cells. Free. Radic. Biol. Med. 2008, 45, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, S.; Anjum, R.; Rangaraj, N.; Pardhasaradhi, B.; Khar, A. Curcumin mediated apoptosis in AK-5 tumor cells involves the production of reactive oxygen intermediates. FEBS Lett. 1999, 456, 311–314. [Google Scholar] [CrossRef]
- Zou, P.; Xia, Y.; Chen, W.; Chen, X.; Ying, S.; Feng, Z.; Chen, T.; Ye, Q.; Wang, Z.; Qiu, C.; et al. EF24 induces ROS-mediated apoptosis via targeting thioredoxin reductase 1 in gastric cancer cells. Oncotarget 2016, 7, 18050–18064. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.-L.; Liang, Y.-J.; Zheng, T.-S.; Song, R.-P.; Wang, J.-B.; Sun, B.-S.; Pan, S.-H.; Qu, L.-D.; Liu, J.-R.; Jiang, H.-C.; et al. EF24 inhibits tumor growth and metastasis via suppressing NF-kappaB dependent pathways in human cholangiocarcinoma. Sci. Rep. 2016, 6, 32167. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.R.; Hussain, A.; Xie, J.; Kosanke, S.; Awasthi, V. The salutary effects of diphenyldifluoroketone EF24 in liver of a rat hemorrhagic shock model. Scand. J. Trauma Resusc. Emerg. Med. 2015, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Feng, C.; Vinothkumar, R.; Chen, W.; Dai, X.; Chen, X.; Ye, Q.; Qiu, C.; Zhou, H.; Wang, Y.; et al. Curcumin analog EF24 induces apoptosis via ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Cancer Chemother. Pharmacol. 2016, 78, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.M.; Buhrow, S.A.; Gilbert, J.A.; Jia, L.; Shoji, M.; Snyder, J.P.; Ames, M.M. Mouse pharmacokinetics and metabolism of the curcumin analog, 4-piperidinone,3,5-bis[(2-fluorophenyl)methylene]-acetate(3E,5E) (EF-24; NSC 716993). Cancer Chemother. Pharmacol. 2014, 73, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, F.; Gold, M.; Begemann, G.; Andronache, I.; Biersack, B.; Schobert, R. Fluoro and pentafluorothio analogs of the antitumoral curcuminoid EF24 with superior antiangiogenic and vascular-disruptive effects. Bioorganic. Med. Chem. 2017, 25, 4894–4903. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, F.; Subramaniam, D.; Anant, S.; Padhye, S.; Begemann, G.; Schobert, R.; Biersack, B. Halogenated Bis(methoxybenzylidene)-4-piperidone curcuminoids with improved anticancer activity. ChemMedChem 2018, 13, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, S.; Zanda, M. Synthetic chemistry and biological activity of pentafluorosulphanyl (SF5) organic molecules. J. Fluor. Chem. 2012, 143, 57–93. [Google Scholar] [CrossRef] [Green Version]
- Lagisetty, P.; Powell, D.R.; Awasthi, V. Synthesis and structural determination of 3,5-bis(2-fluorobenzylidene)-4-piperidone analogs of curcumin. J. Mol. Struct. 2009, 936, 23–28. [Google Scholar] [CrossRef]
- Jansen, B.; Inoue, S.A.; Wadl, H.; Eichler, H.-G.; Wolff, K.; Van Elsas, A.; Schrier, P.I.; Pehamberger, H. N-ras oncogene expression changes the growth characteristics of human melanoma in two independent SCID-hu mouse models. Int. J. Cancer 1996, 67, 821–825. [Google Scholar] [CrossRef]
- Pontén, J.; MacIntyre, E.H. Long term culture of normal and neoplastic human glia. Acta Pathol. Microbiol. Scand. 2009, 74, 465–486. [Google Scholar] [CrossRef] [PubMed]
- Hetschko, H.; Voss, V.; Seifert, V.; Prehn, J.H.M.; Kögel, D. Upregulation of DR5 by proteasome inhibitors potently sensitizes glioma cells to TRAIL-induced apoptosis. FEBS J. 2008, 275, 1925–1936. [Google Scholar] [CrossRef]
- Günther, H.S.; O Schmidt, N.; Phillips, H.S.; Kemming, D.; Kharbanda, S.; Soriano, R.; Modrusan, Z.; Meissner, H.; Westphal, M.; Lamszus, K. Glioblastoma-derived stem cell-enriched cultures form distinct subgroups according to molecular and phenotypic criteria. Oncogene 2007, 27, 2897–2909. [Google Scholar] [CrossRef]
- Campos, B.; Wan, F.; Farhadi, M.; Ernst, A.; Zeppernick, F.; Tagscherer, K.E.; Ahmadi, R.; Lohr, J.; Dictus, C.; Gdynia, G.; et al. Differentiation therapy exerts antitumor effects on stem-like glioma cells. Clin. Cancer Res. 2010, 16, 2715–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, B.; Wehle, A.; Hehlgans, S.; Bonn, F.; Dikic, I.; Rödel, F.; Seifert, V.; Kögel, D. Arsenic Trioxide and (−)-gossypol synergistically target glioma stem-like cells via inhibition of hedgehog and notch signaling. Cancers 2019, 11, 350. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Landegren, U. Measurement of cell numbers by means of the endogenous enzyme hexosaminidase. Applications to detection of lymphokines and cell surface antigens. J. Immunol. Methods 1984, 67, 379–388. [Google Scholar] [CrossRef]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- White, R.M.; Sessa, A.; Burke, C.; Bowman, T.; LeBlanc, J.; Ceol, C.; Bourque, C.; Dovey, M.; Goessling, W.; Burns, C.E.; et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell 2008, 2, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Smyth, G.K. ELDA: Extreme limiting dilution analysis for comparing depleted and enriched populations in stem cell and other assays. J. Immunol. Methods 2009, 347, 70–78. [Google Scholar] [CrossRef]
- Cen, L.; Hutzen, B.; Ball, S.; DeAngelis, S.; Chen, C.-L.; Fuchs, J.R.; Li, C.; Li, P.-K.; Lin, J. New structural analogues of curcumin exhibit potent growth suppressive activity in human colorectal carcinoma cells. BMC Cancer 2009, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Friedman, L.; Lin, L.; Ball, S.; Bekaii-Saab, T.; Fuchs, J.; Li, P.-K.; Li, C.; Lin, J. Curcumin analogues exhibit enhanced growth suppressive activity in human pancreatic cancer cells. Anti-Cancer Drugs 2009, 20, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, D.; May, R.; Sureban, S.; Lee, K.B.; George, R.; Kuppusamy, P.; Ramanujam, R.P.; Hideg, K.; Dieckgraefe, B.K.; Houchen, C.W.; et al. Diphenyl difluoroketone: A curcumin derivative with potent in vivo anticancer activity. Cancer Res. 2008, 68, 1962–1969. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Sidell, N.; Mancini, A.; Huang, R.-P.; Wang, S.; Horowitz, I.R.; Liotta, D.C.; Taylor, R.N.; Wieser, F. Multiple anticancer activities of EF24, a novel curcumin analog, on human ovarian carcinoma cells. Reprod. Sci. 2010, 17, 931–940. [Google Scholar] [CrossRef]
- Serbedzija, G.N.; Flynn, E.; Willett, C.E. Zebrafish angiogenesis: A new model for drug screening. Angiogenesis 1999, 3, 353–359. [Google Scholar] [CrossRef]
- Draut, H.; Rehm, T.; Begemann, G.; Schobert, R. Antiangiogenic and toxic effects of genistein, usnic acid, and their copper complexes in zebrafish embryos at different developmental stages. Chem. Biodivers. 2017, 14, e1600302. [Google Scholar] [CrossRef]
- Jensen, L.; Cao, Z.; Nakamura, M.; Yang, Y.; Bräutigam, L.; Andersson, P.; Zhang, Y.; Wahlberg, E.; Länne, T.; Hosaka, K.; et al. Opposing effects of circadian clock genes Bmal1 and Period2 in regulation of VEGF-dependent angiogenesis in developing zebrafish. Cell Rep. 2012, 2, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta Bioenerg. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader11Mention of a trade name, proprietary product, or specific equipment does not constitute a guarantee by the United States Department of Agriculture and does not imply its approval to the exclusion of other products that may be suitable. Free. Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.T.; Wang, Y.; Chiu, J.-F. Reactive oxygen species: Current knowledge and applications in cancer research and therapeutic. J. Cell. Biochem. 2008, 104, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.C.; Stone, N.L.; Pittman, R.N. Extranuclear Apoptosis. J. Cell Biol. 1999, 146, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tixeira, R.; Caruso, S.; Paone, S.; Baxter, A.A.; Atkin-Smith, G.K.; Hulett, M.D.; Poon, I.K.H. Defining the morphologic features and products of cell disassembly during apoptosis. Apoptosis 2017, 22, 475–477. [Google Scholar] [CrossRef]
- Coleman, M.; Olson, M. Rho GTPase signalling pathways in the morphological changes associated with apoptosis. Cell Death Differ. 2002, 9, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Sordillo, P.P.; Helson, L. Curcumin and cancer stem cells: Curcumin has asymmetrical effects on cancer and normal stem cells. Anticancer. Res. 2015, 35, 599–614. [Google Scholar]
- A Sordillo, L.; Sordillo, P.P.; Helson, L. Curcumin for the Treatment of Glioblastoma. Anticancer. Res. 2015, 35, 6373–6378. [Google Scholar]
- Adams, B.K.; Ferstl, E.M.; Davis, M.C.; Herold, M.; Kurtkaya, S.; Camalier, R.F.; Hollingshead, M.G.; Kaur, G.; Sausville, E.A.; Rickles, F.R.; et al. Synthesis and biological evaluation of novel curcumin analogs as anti-cancer and anti-angiogenesis agents. Bioorganic. Med. Chem. 2004, 12, 3871–3883. [Google Scholar] [CrossRef]
- Adams, B.K.; Cai, J.; Armstrong, J.; Herold, M.; Lu, Y.J.; Sun, A.; Snyder, J.P.; Liotta, D.C.; Jones, D.P.; Shoji, M. EF24, a novel synthetic curcumin analog, induces apoptosis in cancer cells via a redox-dependent mechanism. Anti-Cancer Drugs 2005, 16, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Kasinski, A.L.; Du, Y.; Thomas, S.L.; Zhao, J.; Sun, S.-Y.; Khuri, F.R.; Wang, C.-Y.; Shoji, M.; Sun, A.; Snyder, J.P.; et al. Inhibition of IκB kinase-nuclear factor-κB signaling pathway by 3,5-Bis(2-flurobenzylidene)piperidin-4-one (EF24), a novel monoketone analog of curcumin. Mol. Pharmacol. 2008, 74, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.L.; Zhong, D.; Zhou, W.; Malik, S.; Liotta, D.; Snyder, J.P.; Hamel, E.; Giannakakou, P. EF24, a novel curcumin analog, disrupts the microtubule cytoskeleton and inhibits HIF-1. Cell Cycle 2008, 7, 2409–2417. [Google Scholar] [CrossRef] [Green Version]
- Moreira, J.; Saraiva, L.; Pinto, M.; Cidade, H. Diarylpentanoids with antitumor activity: A critical review of structure-activity relationship studies. Eur. J. Med. Chem. 2020, 192, 112177. [Google Scholar] [CrossRef]
- Weissenberger, J.; Priester, M.; Bernreuther, C.; Rakel, S.; Glatzel, M.; Seifert, V.; Kögel, D. Dietary curcumin attenuates glioma growth in a syngeneic mouse model by inhibition of the JAK1,2/STAT3 signaling pathway. Clin. Cancer Res. 2010, 16, 5781–5795. [Google Scholar] [CrossRef] [Green Version]
- Senft, C.; Polacin, M.; Priester, M.; Seifert, V.; Kögel, D.; Weissenberger, J. The nontoxic natural compound Curcumin exerts anti-proliferative, anti-migratory, and anti-invasive properties against malignant gliomas. BMC Cancer 2010, 10, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bimonte, S.; Barbieri, A.; Palma, G.; Luciano, A.; Rea, D.; Arra, C. Curcumin inhibits tumor growth and angiogenesis in an orthotopic mouse model of human pancreatic cancer. BioMed Res. Int. 2013, 2013, 810423. [Google Scholar] [CrossRef]
- Carroll, C.E.; Ellersieck, M.R.; Hyder, S.M. Curcumin inhibits MPA-induced secretion of VEGF from T47-D human breast cancer cells. Menopause 2008, 15, 570–574. [Google Scholar] [CrossRef]
- Lu, Z.J.; Ren, Y.Q.; Wang, G.P.; Song, Q.; Li, M.; Jiang, S.S.; Ning, T.; Guan, Y.S.; Yang, J.L.; Luo, F. Biological behaviors and proteomics analysis of hybrid cell line EAhy926 and its parent cell line A549. J. Exp. Clin. Cancer Res. 2009, 28, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebradshaw, A.; Ewickremesekera, A.; Tan, S.T.; Epeng, L.; Davis, P.F.; Itinteang, T. Cancer Stem Cell Hierarchy in Glioblastoma Multiforme. Front. Surg. 2016, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Atashzar, M.R.; Baharlou, R.; Karami, J.; Abdollahi, H.; Rezaei, R.; Pourramezan, F.; Moghaddam, S.H.Z. Cancer stem cells: A review from origin to therapeutic implications. J. Cell. Physiol. 2020, 235, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Zendehdel, E.; Abdollahi, E.; Momtazi-Borojeni, A.A.; Korani, M.; Alavizadeh, S.H.; Sahebkar, A. The molecular mechanisms of curcumin’s inhibitory effects on cancer stem cells. J. Cell. Biochem. 2019, 120, 4739–4747. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | Tumor Type | Reference |
|---|---|---|
| 518A2 | Melanoma | [14] |
| Panc-1 | Pancreatic ductular adenocarcinoma | ACC-783 |
| KB-V1Vbl | Cervix carcinoma, Vinblastine resistant | ACC-149 |
| MCF-7Topo | Breast Carcinoma, Topotecan resistant | ACC-115 |
| HT-29 | Colon Carcinoma | ACC-299 |
| HCT-116 | Colon Carcinoma | ACC-581 |
| DLD-1 | Colon Carcinoma | ACC-278 |
| EA.hy926 | Endothelial hybrid | CRL-2922 |
| SW-480 | Colon Carcinoma | CCL-228 |
| U251 | Glioblastoma | ECACC 09063001 |
| Mz54 | Glioblastoma | CVCL_M406 |
| GS-5 | Glioblastoma, stem-like cells | [17] |
| NCH644 | Glioblastoma, stem-like cells | [18] |
| 17/02 | Glioblastoma, stem-like cells | [19] |
| Compounds | 1a | 1b [b] | EF24 (2) [b] | Curcumin (3) |
|---|---|---|---|---|
| 518A2 | 0.17 ± 0.02 | 0.99 ± 0.14 | 1.8 ± 0.2 | - |
| HT-29 | 0.20 ± 0.01 | 0.29 ± 0.02 | 1.6 ± 0.1 | 13.3 [c] |
| HCT-116 | 0.11 ± 0.01 | 0.24 ± 0.02 | 1.5 ± 0.2 | 10.9 [c] |
| DLD-1 | 0.16 ± 0.01 | 0.33 ± 0.01 | 1.3 ± 0.1 | - |
| KB-V1Vbl | 0.15 ± 0.01 | 0.64 ± 0.05 | 1.1 ± 0.1 | - |
| MCF-7Topo | 0.22 ± 0.02 | 0.40 ± 0.01 | 2.2 ± 0.2 | - |
| Panc-1 | 0.19 ± 0.01 | 1.1 ± 0.1 | 1.5 ± 0.2 | 20.4 [c] |
| EA.hy926 | 0.31 ± 0.03 | 0.54 ± 0.03 | 1.4 ± 0.1 | |
| U251 | 0.22 ± 0.02 | 0.79 ± 0.03 | - | - |
| Mz54 | 0.27 ± 0.02 | 0.83 ± 0.02 | - | - |
| HCT-116 | SW-480 | |||||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| 1a | 0.16 ± 0.02 | 0.073 ± 0.01 | 0.074 ± 0.01 | 0.56 ± 0,1 | 0.11 ± 0.02 | 0.1 ± 0.01 |
| 1b | 0.34 ± 0.04 | 0.14 ± 0.02 | 0.15 ± 0.01 | 0.75 ± 0.14 | 0.22 ± 0.04 | 0.17 ± 0.03 |
| 2 | 0.47 ± 0.07 | 0.26 ± 0.07 | 0.29 ± 0.03 | 1.1 ± 0.3 | 0.37 ± 0.07 | 0.24 ± 0.03 |
| Compound | 1a | 1b | ||
|---|---|---|---|---|
| IC50 after Incubation | 48 h | 72 h | 48 h | 72 h |
| GS-5 | 0.37 ± 0.04 | 0.32 ± 0.05 | 0.95 ± 0.04 | 1.3 ± 0.04 |
| NCH644 | 0.43 ± 0.02 | 0.35 ± 0.01 | 1.7 ± 0.02 | 1.5 ± 0.02 |
| 17/02 | 0.32 ± 0.03 | 0.22 ± 0.03 | 0.99 ± 0.04 | 1.0 ± 0.03 |
| Compound | DMSO | 1a | 1b | ||
|---|---|---|---|---|---|
| Stem-Cell Frequency (1/X) | IC25 | IC50 | IC25 | IC50 | |
| GS-5 | 196 | 305 | 357 | 839 | 1199 |
| NCH644 | 8 | 145 | 131 | 104 | 5626 |
| 17/02 | 216 | 1680 | 4288 | 5169 | 11,319 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linder, B.; Köhler, L.H.F.; Reisbeck, L.; Menger, D.; Subramaniam, D.; Herold-Mende, C.; Anant, S.; Schobert, R.; Biersack, B.; Kögel, D. A New Pentafluorothio-Substituted Curcuminoid with Superior Antitumor Activity. Biomolecules 2021, 11, 947. https://doi.org/10.3390/biom11070947
Linder B, Köhler LHF, Reisbeck L, Menger D, Subramaniam D, Herold-Mende C, Anant S, Schobert R, Biersack B, Kögel D. A New Pentafluorothio-Substituted Curcuminoid with Superior Antitumor Activity. Biomolecules. 2021; 11(7):947. https://doi.org/10.3390/biom11070947
Chicago/Turabian StyleLinder, Benedikt, Leonhard H. F. Köhler, Lisa Reisbeck, Dominic Menger, Dharmalingam Subramaniam, Christel Herold-Mende, Shrikant Anant, Rainer Schobert, Bernhard Biersack, and Donat Kögel. 2021. "A New Pentafluorothio-Substituted Curcuminoid with Superior Antitumor Activity" Biomolecules 11, no. 7: 947. https://doi.org/10.3390/biom11070947
APA StyleLinder, B., Köhler, L. H. F., Reisbeck, L., Menger, D., Subramaniam, D., Herold-Mende, C., Anant, S., Schobert, R., Biersack, B., & Kögel, D. (2021). A New Pentafluorothio-Substituted Curcuminoid with Superior Antitumor Activity. Biomolecules, 11(7), 947. https://doi.org/10.3390/biom11070947