
RT-QuIC Using C-Terminally Truncated α-Synuclein Forms Detects Differences in Seeding Propensity of Different Brain Regions from Synucleinopathies

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Expression and Purification of Recombinant αSyn Proteins
2.2. Circular Dichroism Spectra
2.3. Isolation of TBS (aqueous)-Soluble and Detergent-Soluble Fractions from Brain Tissue
2.4. RT-QuIC Assay
2.5. Proteinase K Digestion and Western Blotting
2.6. Transmission Electron Microscopy (TEM)
2.7. Statistical Analysis
3. Results
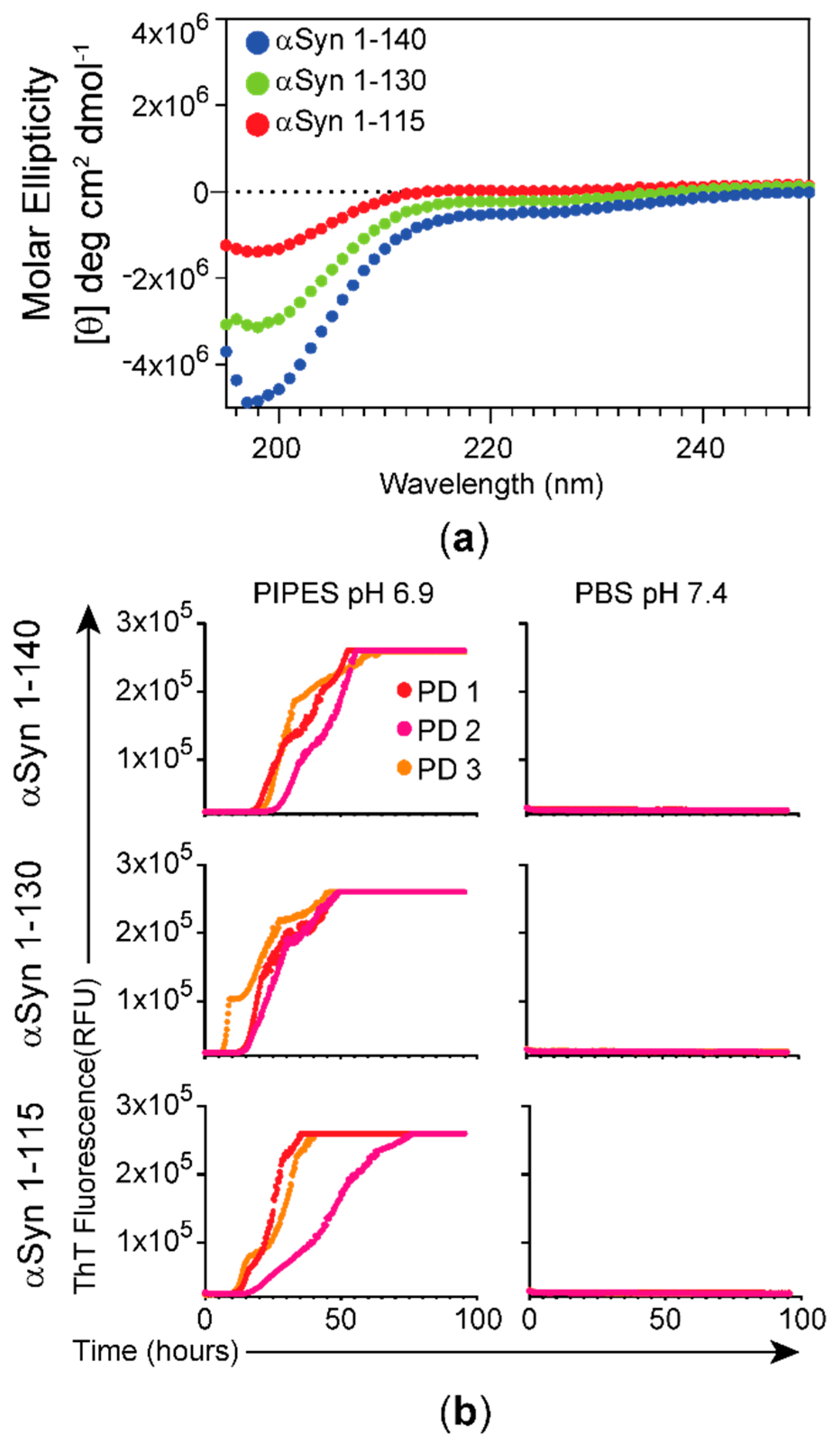
3.1. Characterization of αSyn Forms Used as Substrates and Effect of Buffer Composition on the RT-QuIC Assay
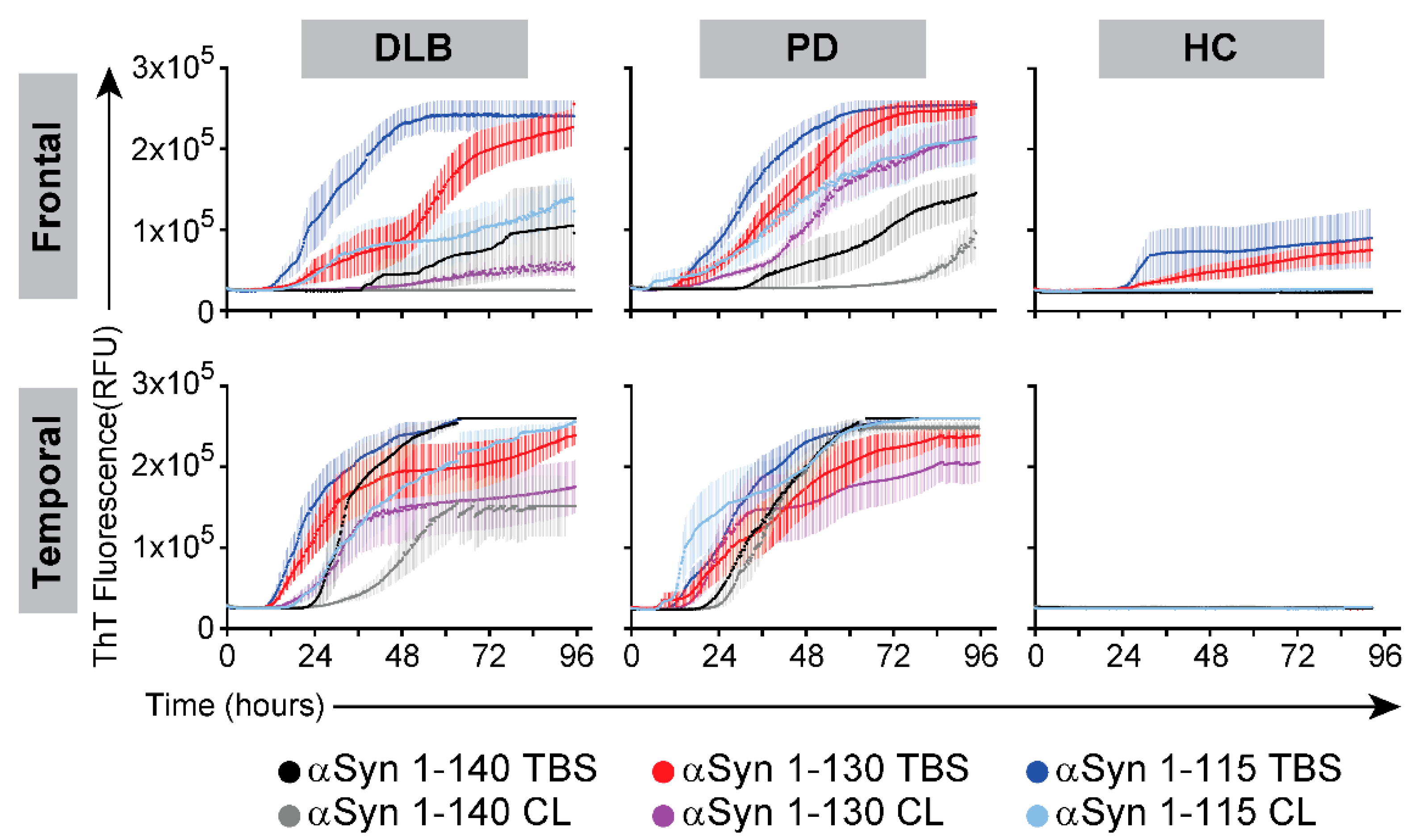
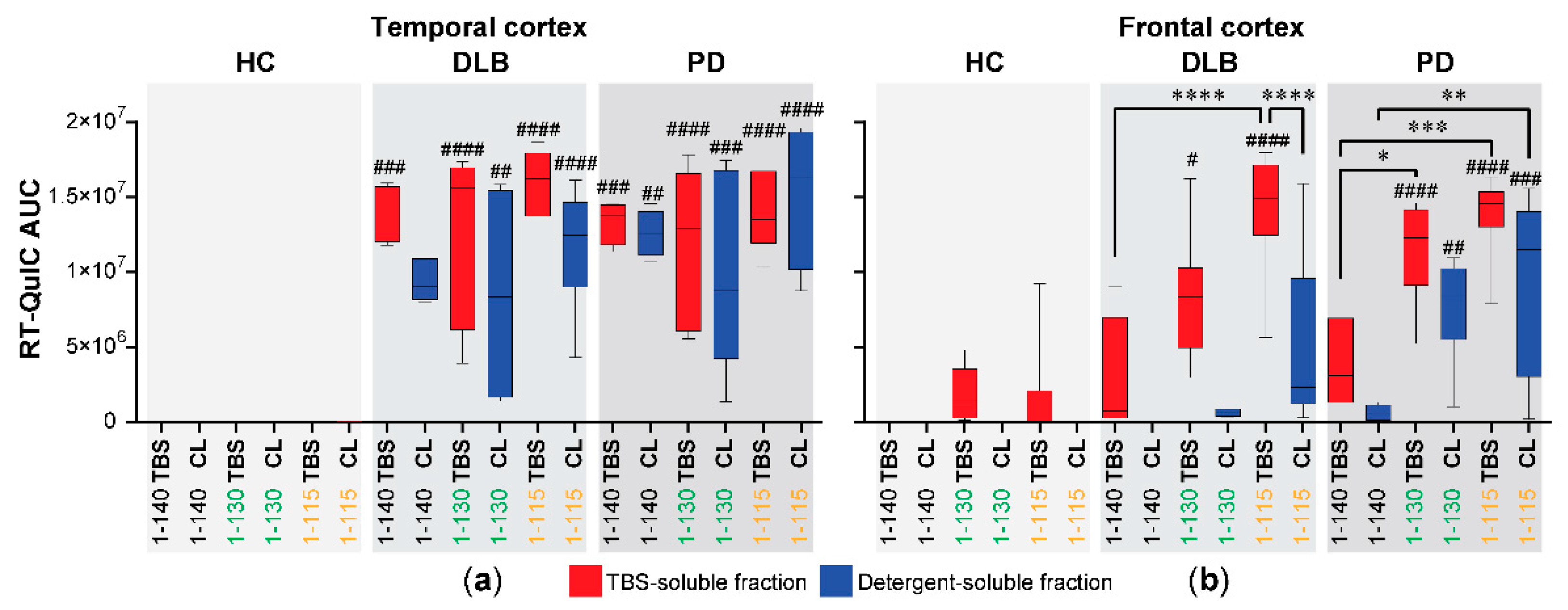
3.2. Seeding RT-QuIC Reactions with Brain Homogenates from PD and DLB Cases Using Different Forms of αSyn
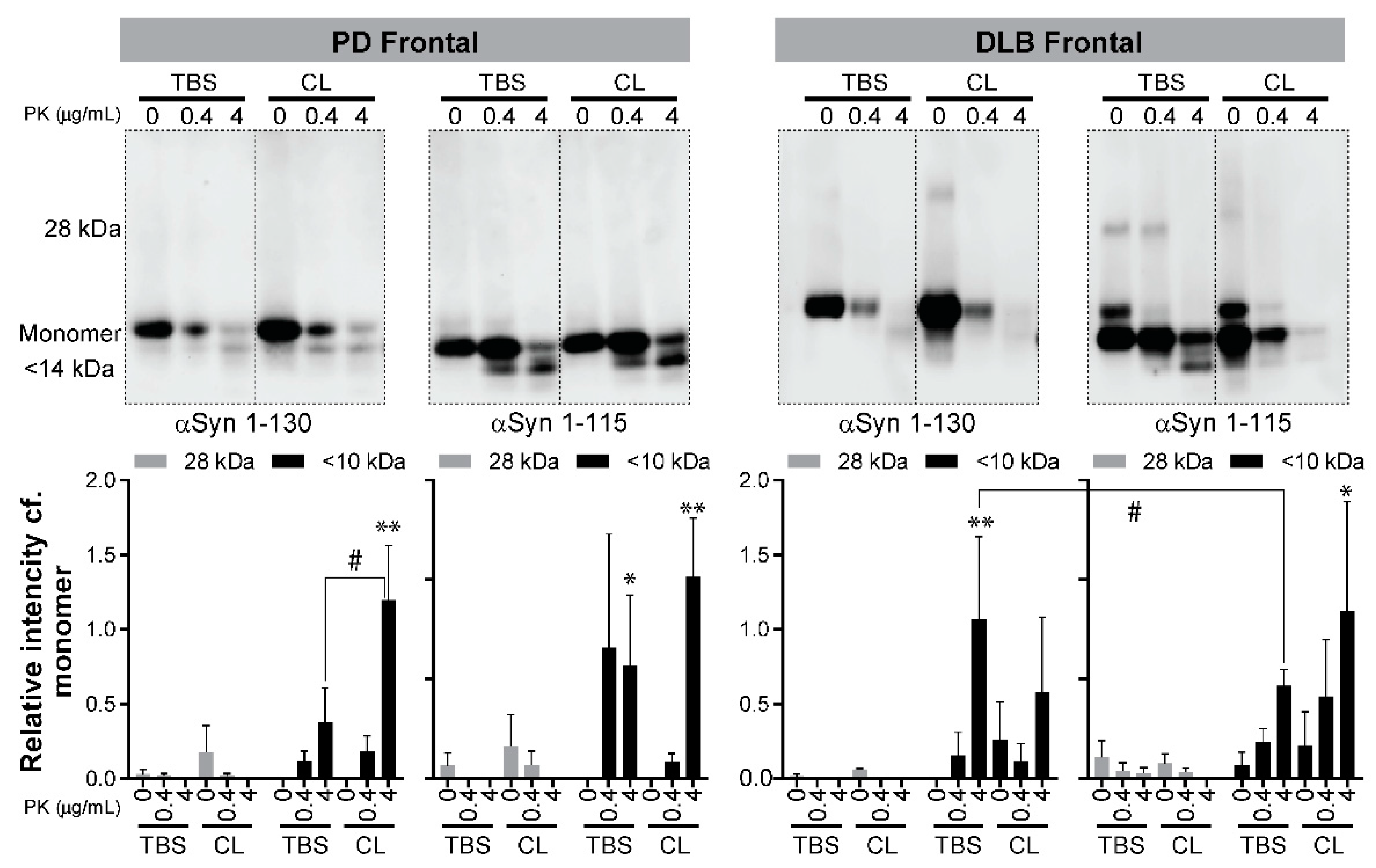
3.3. Detection of Proteinase K (PK)-Resistant Fragments of C-Terminal Truncated αSyn in the RT-QuIC End Products
3.4. Morphological Characterization of RT-QuIC End Products by TEM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCann, H.; Stevens, C.H.; Cartwright, H.; Halliday, G.M. alpha-Synucleinopathy phenotypes. Parkinsonism Relat. Disord. 2014, 20, S62–S67. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. alpha-Synuclein in filamentous inclusions of Lewy bodies from Parkinson’s disease and dementia with lewy bodies. Proc. Natl. Acad. Sci. USA 1998, 95, 6469–6473. [Google Scholar] [CrossRef]
- Halliday, G.M.; Song, Y.J.C.; Harding, A.J. Striatal beta-amyloid in dementia with Lewy bodies but not Parkinson’s disease. J. Neural Transm. 2011, 118, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Peelaerts, W.; Bousset, L.; Van der Perren, A.; Moskalyuk, A.; Pulizzi, R.; Giugliano, M.; Van den Haute, C.; Melki, R.; Baekelandt, V. alpha-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature 2015, 522, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Masliah, E. Neurodegeneration: Aggregates feel the strain. Nature 2015, 522, 296–297. [Google Scholar] [CrossRef]
- Campbell, B.C.V.; McLean, C.A.; Culvenor, J.G.; Gai, W.P.; Blumbergs, P.C.; Jäkälä, P.; Beyreuther, K.; Masters, C.L.; Li, Q.-X. The solubility of alpha-synuclein in multiple system atrophy differs from that of dementia with Lewy bodies and Parkinson’s disease. J. Neurochem. 2001, 76, 87–96. [Google Scholar] [CrossRef]
- Van der Perren, A.; Gelders, G.; Fenyi, A.; Bousset, L.; Brito, F.; Peelaerts, W.; Van den Haute, C.; Gentleman, S.; Melki, R.; Baekelandt, V. The structural differences between patient-derived alpha-synuclein strains dictate characteristics of Parkinson’s disease, multiple system atrophy and dementia with Lewy bodies. Acta Neuropathol. 2020, 139, 977–1000. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, Z.A.; Goodwin, M.S.; Riffe, C.J.; Dhillon, J.-K.S.; Xia, Y.; Gorion, K.-M.; Vijayaraghavan, N.; McFarland, K.N.; Golbe, L.I.; Yachnis, A.T.; et al. Unique alpha-synuclein pathology within the amygdala in Lewy body dementia: Implications for disease initiation and progression. Acta Neuropathol. Commun. 2019, 7, 142. [Google Scholar] [CrossRef]
- Candelise, N.; Schmitz, M.; Llorens, F.; Villar-Piqué, A.; Cramm, M.; Thom, T.; da Silva Correia, S.M.; da Cunha, J.E.G.; Möbius, W.; Outeiro, T.F.; et al. Seeding variability of different alpha synuclein strains in synucleinopathies. Ann. Neurol. 2019, 85, 691–703. [Google Scholar] [CrossRef]
- Shahnawaz, M.; Mukherjee, A.; Pritzkow, S.; Mendez, N.; Rabadia, P.; Liu, X.; Hu, B.; Schmeichel, A.; Singer, W.; Wu, G.; et al. Discriminating alpha-synuclein strains in Parkinson’s disease and multiple system atrophy. Nature 2020, 578, 273–277. [Google Scholar] [CrossRef]
- Lewis, K.A.; Su, Y.; Jou, O.; Ritchie, C.; Foong, C.; Hynan, L.S.; White, C.L., III; Thomas, P.J.; Hatanpaa, K.J. Abnormal neurites containing C-terminally truncated alpha-synuclein are present in Alzheimer’s disease without conventional Lewy body pathology. Am. J. Pathol. 2010, 177, 3037–3050. [Google Scholar] [CrossRef]
- Rossi, M.; Candelise, N.; Baiardi, S.; Capellari, S.; Giannini, G.; Orru, C.D.; Antelmi, E.; Mammana, A.; Hughson, A.G.; Calandra-Buonaura, G.; et al. Ultrasensitive RT-QuIC assay with high sensitivity and specificity for Lewy body-associated synucleinopathies. Acta Neuropathol. 2020, 140, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Giasson, B.I.; Lewis, K.A.; Lee, V.M.; Demartino, G.N.; Thomas, P.J. A precipitating role for truncated alpha-synuclein and the proteasome in alpha-synuclein aggregation: Implications for pathogenesis of Parkinson disease. J. Biol. Chem. 2005, 280, 22670–22678. [Google Scholar] [CrossRef] [PubMed]
- Murray, I.V.; Giasson, B.I.; Quinn, S.M.; Koppaka, V.; Axelsen, P.H.; Ischiropoulos, H.; Trojanowski, J.Q.; Lee, V.M. Role of alpha-synuclein carboxy-terminus on fibril formation in vitro. Biochemistry 2003, 42, 8530–8540. [Google Scholar] [CrossRef] [PubMed]
- Periquet, M.; Fulga, T.; Myllykangas, L.; Schlossmacher, M.G.; Feany, B.M. Aggregated alpha-synuclein mediates dopaminergic neurotoxicity in vivo. J. Neurosci. 2007, 27, 3338–3346. [Google Scholar] [CrossRef] [PubMed]
- Michell, A.W.; Tofaris, G.K.; Gossage, H.; Tyers, P.; Spillantini, M.G.; Barker, R.A. The effect of truncated human alpha-synuclein (1-120) on dopaminergic cells in a transgenic mouse model of Parkinson’s disease. Cell Transpl. 2007, 16, 461–474. [Google Scholar] [CrossRef]
- Baba, M.; Nakajo, S.; Tu, P.H.; Tomita, T.; Nakaya, K.; Lee, V.M.; Trojanowski, J.Q.; Iwatsubo, T. Aggregation of alpha-synuclein in Lewy bodies of sporadic Parkinson’s disease and dementia with Lewy bodies. Am. J. Pathol. 1998, 152, 879–884. [Google Scholar]
- Sorrentino, Z.A.; Vijayaraghavan, N.; Gorion, K.M.; Riffe, C.J.; Strang, K.H.; Caldwell, J.; Giasson, B.I. Physiological C-terminal truncation of alpha-synuclein potentiates the prion-like formation of pathological inclusions. J. Biol. Chem. 2018, 293, 18914–18932. [Google Scholar] [CrossRef]
- Chakroun, T.; Evsyukov, V.; Nykanen, N.P.; Hollerhage, M.; Schmidt, A.; Kamp, F.; Ruf, V.C.; Wurst, W.; Rosler, T.W.; Hoglinger, G.U. Alpha-synuclein fragments trigger distinct aggregation pathways. Cell Death Dis. 2020, 11, 84. [Google Scholar] [CrossRef]
- Vaikath, N.N.; Erskine, D.; Morris, C.M.; Majbour, N.K.; Vekrellis, K.; Li, J.Y.; El-Agnaf, O.M.A. Heterogeneity in alpha-synuclein subtypes and their expression in cortical brain tissue lysates from Lewy body diseases and Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2019, 45, 597–608. [Google Scholar] [CrossRef]
- Fairfoul, G.; McGuire, L.I.; Pal, S.; Ironside, J.W.; Neumann, J.; Christie, S.; Joachim, C.; Esiri, M.; Evetts, S.G.; Rolinski, M.; et al. Alpha-synuclein RT-QuIC in the CSF of patients with alpha-synucleinopathies. Ann. Clin. Transl. Neurol. 2016, 3, 812–818. [Google Scholar] [CrossRef]
- Koss, D.J.; Dubini, M.; Buchanan, H.; Hull, C.; Platt, B. Distinctive temporal profiles of detergent-soluble and -insoluble tau and Abeta species in human Alzheimer’s disease. Brain Res. 2018, 1699, 121–134. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.M.G.; Elia, A.E.; Portaleone, S.M.; Cazzaniga, F.A.; Rossi, M.; Bistaffa, E.; De Cecco, E.; Narkiewicz, J.; Salzano, G.; Carletta, O.; et al. Efficient RT-QuIC seeding activity for alpha-synuclein in olfactory mucosa samples of patients with Parkinson’s disease and multiple system atrophy. Transl. Neurodegener. 2019, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Manne, S.; Kondru, N.; Jin, H.; Anantharam, V.; Huang, X.; Kanthasamy, A.; Kanthasamy, A.G. alpha-Synuclein real-time quaking-induced conversion in the submandibular glands of Parkinson’s disease patients. Mov. Disord. 2020, 35, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Candelise, N.; Schmitz, M.; Thune, K.; Cramm, M.; Rabano, A.; Zafar, S.; Stoops, E.; Vanderstichele, H.; Villar-Pique, A.; Llorens, F.; et al. Effect of the micro-environment on alpha-synuclein conversion and implication in seeded conversion assays. Transl. Neurodegener. 2020, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Koob, A.O.; Shaked, G.M.; Bender, A.; Bisquertt, A.; Rockenstein, E.; Masliah, E. Neurogranin binds alpha-synuclein in the human superior temporal cortex and interaction is decreased in Parkinson’s disease. Brain. Res. 2014, 1591, 102–110. [Google Scholar] [CrossRef]
- Kellie, J.F.; Higgs, R.E.; Ryder, J.W.; Major, A.; Beach, T.G.; Adler, C.H.; Merchant, K.; Knierman, M.D. Quantitative measurement of intact alpha-synuclein proteoforms from post-mortem control and Parkinson’s disease brain tissue by intact protein mass spectrometry. Sci. Rep. 2014, 4, 5797. [Google Scholar] [CrossRef]
- Dufty, B.M.; Warner, L.R.; Hou, S.T.; Jiang, S.X.; Gomez-Isla, T.; Leenhouts, K.M.; Oxford, J.T.; Feany, M.B.; Masliah, E.; Rohn, T.T. Calpain-cleavage of alpha-synuclein: Connecting proteolytic processing to disease-linked aggregation. Am. J. Pathol. 2007, 170, 1725–1738. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a | TBS-Soluble Fraction | Detergent-Soluble Fraction | ||||||
| Temporal | Sens. | Spec. | p * | Cutoff | Sens. | Spec. | p * | Cutoff |
| (RT-QuIC AUC) | (RT-QuIC AUC) | |||||||
| αSyn 1-140 | ||||||||
| Cases vs. HC | 100% | 100% | 0.006 | >5,686,027 | 100% | 100% | 0.007 | >4,010,801 |
| PD vs. HC | 100% | 100% | 0.02 | >5,686,027 | 100% | 100% | 0.02 | >5,343,731 |
| DLB vs. HC | 100% | 100% | 0.02 | >5,889,846 | 100% | 100% | 0.02 | >4,010,801 |
| PD vs. DLB | 75% | 50% | 0.563 | <14,474,992 | 100% | 75% | 0.043 | >10,077,548 |
| αSyn 1-130 | ||||||||
| Cases vs. HC | 100% | 100% | 0.003 | >1,957,147 | 94% | 100% | 0.003 | >1,367,850 |
| PD vs. HC | 100% | 100% | 0.007 | >2,775,490 | 100% | 100% | 0.007 | >700,526 |
| DLB vs. HC | 100% | 100% | 0.007 | >1,957,147 | 88% | 100% | 0.007 | >1,404,507 |
| PD vs. DLB | 75% | 50% | 0.834 | <16,198,128 | 88% | 50% | 0.401 | >3,158,315 |
| αSyn 1-115 | ||||||||
| Cases vs. HC | 100% | 100% | 0.003 | >5,198,217 | 94% | 100% | 0.003 | >6,530,860 |
| PD vs. HC | 100% | 100% | 0.007 | >5,198,217 | 100% | 100% | 0.007 | >4,395,742 |
| DLB vs. HC | 100% | 100% | 0.007 | >6,335,727 | 100% | 100% | 0.007 | >2,164,070 |
| PD vs. DLB | 63% | 75% | 0.142 | <14,687,906 | 63% | 63% | 0.207 | <13,688,774 |
| b | TBS-soluble fraction | Detergent-soluble fraction | ||||||
| Frontal | Sens. | Spec. | p * | Cutoff | Sens. | Spec. | p * | Cutoff |
| (RT-QuIC AUC) | (RT-QuIC AUC) | |||||||
| αSyn 1-140 | ||||||||
| Cases vs. HC | 100% | 100% | 0.006 | <35,870 | 100% | 100% | 0.006 | >64.66 |
| PD vs. HC | 100% | 100% | 0.02 | >515,271 | 100% | 100% | 0.02 | >72.59 |
| DLB vs. HC | 100% | 100% | 0.02 | >35,870 | 100% | 100% | 0.006 | >64.66 |
| PD vs. DLB | 75% | 100% | 0.25 | <893,180 | 100% | 75% | 0.15 | <200,163 |
| αSyn 1-130 | ||||||||
| Cases vs. HC | 100% | 100% | 0.001 | >5,142,617 | 100% | 50% | 0.024 | 0.7891 |
| PD vs. HC | 100% | 100% | 0.001 | >5,142,617 | 88% | 88% | 0.003 | 0.9375 |
| DLB vs. HC | 88% | 88% | 0.005 | >4,666,272 | 88% | 50% | 0.345 | >318,073 |
| PD vs. DLB | 75% | 88% | 0.093 | >10,642,878 | 88% | 100% | 0.001 | <957,841 |
| αSyn 1-115 | ||||||||
| Cases vs. HC | 88% | 75% | 0.006 | >11,421,461 | 87% | 88% | 0.001 | >673,667 |
| PD vs. HC | 75% | 88% | 0.021 | >13,701,264 | 88% | 88% | 0.002 | >1,205,104 |
| DLB vs. HC | 88% | 75% | 0.016 | >11,421,461 | 88% | 88% | 0.009 | >673,667 |
| PD vs. DLB | 88% | 50% | 0.528 | <15,596,073 | 75% | 75% | 0.248 | <4,442,097 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poggiolini, I.; Erskine, D.; Vaikath, N.N.; Ponraj, J.; Mansour, S.; Morris, C.M.; El-Agnaf, O.M.A. RT-QuIC Using C-Terminally Truncated α-Synuclein Forms Detects Differences in Seeding Propensity of Different Brain Regions from Synucleinopathies. Biomolecules 2021, 11, 820. https://doi.org/10.3390/biom11060820
Poggiolini I, Erskine D, Vaikath NN, Ponraj J, Mansour S, Morris CM, El-Agnaf OMA. RT-QuIC Using C-Terminally Truncated α-Synuclein Forms Detects Differences in Seeding Propensity of Different Brain Regions from Synucleinopathies. Biomolecules. 2021; 11(6):820. https://doi.org/10.3390/biom11060820
Chicago/Turabian StylePoggiolini, Ilaria, Daniel Erskine, Nishant N. Vaikath, Janarthanan Ponraj, Said Mansour, Christopher M. Morris, and Omar M. A. El-Agnaf. 2021. "RT-QuIC Using C-Terminally Truncated α-Synuclein Forms Detects Differences in Seeding Propensity of Different Brain Regions from Synucleinopathies" Biomolecules 11, no. 6: 820. https://doi.org/10.3390/biom11060820
APA StylePoggiolini, I., Erskine, D., Vaikath, N. N., Ponraj, J., Mansour, S., Morris, C. M., & El-Agnaf, O. M. A. (2021). RT-QuIC Using C-Terminally Truncated α-Synuclein Forms Detects Differences in Seeding Propensity of Different Brain Regions from Synucleinopathies. Biomolecules, 11(6), 820. https://doi.org/10.3390/biom11060820