
The Role of Hydrogen Sulfide in Respiratory Diseases

 , , , ,
, , , ,
Abstract
1. Introduction
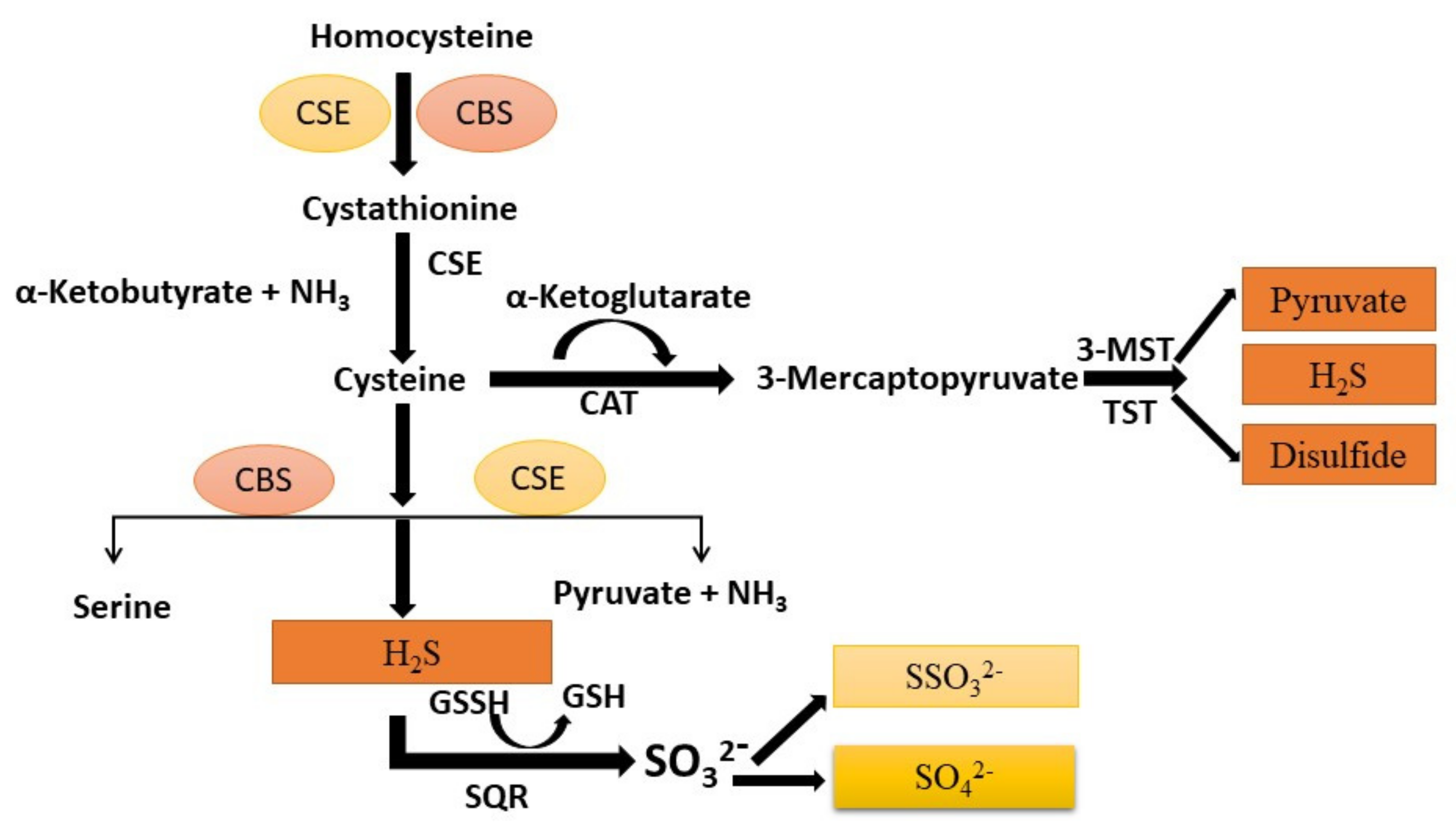
2. Regulation of H2S and H2S-Mediated Sulfhydration in the Lung
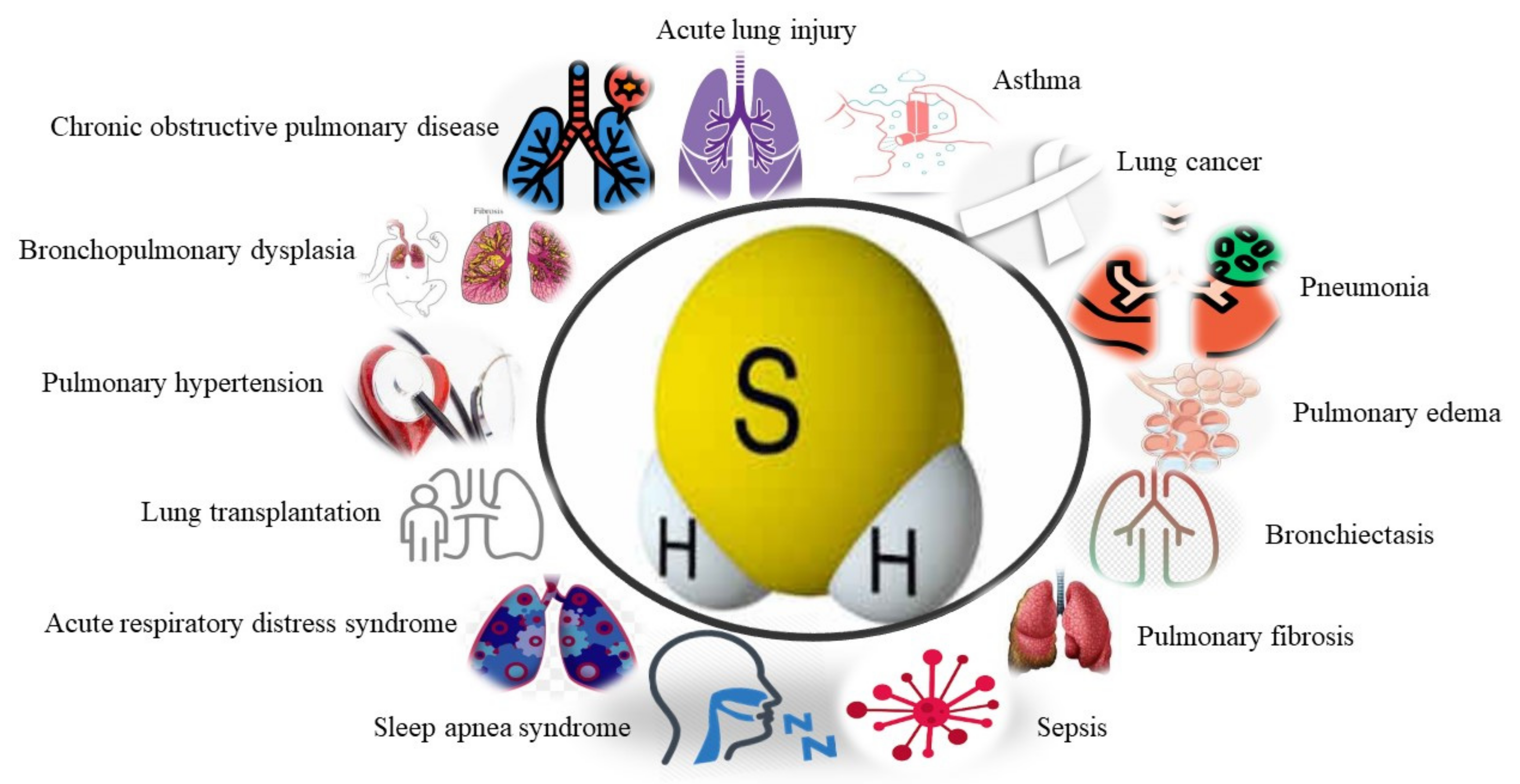
3. The Role of H2S in Respiratory Diseases
3.1. H2S and Chronic Obstructive Pulmonary Diseases
3.2. H2S and Acute Lung Injury (ALI)
3.2.1. Pancreatic Inflammatory Lung Injury
3.2.2. Inhalation-Induced Lung Injury
3.2.3. Ventilator-Induced Lung Injury (VILI)
3.3. H2S and Asthma
3.4. H2S and Lung Cancer
3.5. H2S and Pneumonia
3.6. H2S and Pulmonary Edema
3.7. H2S and Bronchiectasis
3.8. H2S and Pulmonary Fibrosis
3.9. H2S and Sepsis
3.10. H2S and Lung Transplantation
3.11. H2S and Pulmonary Hypertension
3.12. H2S and Sleep Apnea Syndrome (SAS)
3.13. H2S and Acute Respiratory Distress Syndrome
3.14. H2S and Bronchopulmonary Dysplasia (BPD)
4. H2S in the Physiopathology of Airways
5. H2S in Pulmonary Inflammation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Action | H2S | References |
|---|---|---|
| Vasodilation | ↑ | [153,154] |
| Stable asthma | ↓ | [35,36,38,52,68,102,174] |
| Bronchodilation | ↑ | [103] |
| Angiogenic activity | ↑ | [22] |
| Pro-inflammatory action | ↑ | [61] |
| Anti-inflammatory action | ↑ | [61] |
| Airway hyper-reactivity | ↑ | [63,64] |
| Asthma exacerbation | ↓ | [35,36,38,52,68,102,174] |
| Stable COPD | ↑ | [65] |
| COPD exacerbation | ↓ | [37] |
6. Clinical Trials of H2S Donors
7. Perceptions, Limitations and Prospects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| H2S | hydrogen sulfide |
| CBS | cystathionine beta-synthase |
| CSE | cystathionine γ-lyase |
| 3-MPST | 3-mercaptopyruavte sulfurtransferase |
| COPD | chronic obstructive pulmonary disease |
| SQR | sulfide-quinone reductase |
| TST | thiosulfate sulfurtransferase |
| CAT | cysteine aminotransferase |
| GSSH | glutathione |
| SMC | smooth muscle cells |
| ROS | reactive oxygen species |
| ALI | acute lung injury |
| NO | nitric oxide |
| ATF-30 | transcription factor-3 |
| OA | oleic acid |
| ECM | extracellular matrix |
| MLC | myosin light chain |
| PHT | pulmonary hypertension |
| SM | sulfur mustard |
| NSAID | non-steroidal anti-inflammatory drugs |
| MODS | multiple organ dysfunction syndrome |
| MDA | malondialdehyde |
| MPO | myeloperoxidase |
| ARDS | acute respiratory distress syndrome |
| LPVS | lung-protective ventilation strategies |
| VILI | ventilator-induced lung injury |
| FEV | forced expiratory volume |
| LTx | lung transplantation |
| PH | pulmonary hypertension |
| RSV | respiratory syncytial virus |
| IPF | idiopathic pulmonary fibrosis |
| CSAS | central sleep apnea syndrome |
| SAS | sleep apnea syndrome |
| hMPV | human metapneumovirus |
| CB | carotid body |
| CO | carbon monoxide |
| BPD | bronchopulmonary dysplasia |
| ATII | alveolar Type II |
| NaHS | sodium hydrosulfide |
| HO-1 | heme oxygenase-1 |
| STAT-3 | signal transducer and activator of transporter-1 |
| Nrf-2 | nuclear factor erythroid-2 related factor |
| NF-қβ | nuclear factor-kappa B |
| PI3K | phosphoinositide 3-kinase |
| ERK | extracellular signal-regulated kinase |
| AMPK | AMP-activated protein kinase |
| TNF-α | tumor necrosis factor-α |
| TGF-β1 | transforming growth factor beta 1 |
| SOD | superoxide dismutase |
| IL | interleukin |
| IKK | IқB kinase |
| Keap1 | Kelch-like-ECH-associated protein |
| ADT-OH | 5-(hydroxyphenyl)-3H-1:2-dithiole-3-thione |
| ATB-429 | 4-(5-sulfanylidenedithiol-3-y) phenyl 5-amino-2-hydroxybenzoate |
| STS | sodium thiosulfate |
References
- Scano, G.; Innocenti-Bruni, G.; Stendardi, L. Do obstructive and restrictive lung diseases share common underlying mechanisms of breathlessness? Respir. Med. 2010, 104, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Dasaraju, P.V.; Liu, C. Infections of the Respiratory System, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Bartlett, J.G. Management of Respiratory Tract Infections; Williams & Wilkins: Baltimore, MD, USA, 1997. [Google Scholar]
- Perez-Padilla, R.P.B.; Marks, G.; Wong, G.; Bateman, E.; Jarvis, D.; Prabhakaran, D.; Anand, S.; Gaziano, T.A.; Mbanya, J.-C.; Wu, Y.; et al. Chronic Lower Respiratory Tract Diseases. In Cardiovascular, Respiratory, and Related Disorders, 3rd ed.; World Bank: Washington, DC, USA, 2017; Volume 5, pp. 263–285. [Google Scholar]
- Zar, H.J.; Ferkol, T.W. The global burden of respiratory disease-Impact on child health. Pediatr. Pulmonol. 2014, 49, 430–434. [Google Scholar] [CrossRef]
- World Health Organization. WHO Strategy for Prevention and Control of Chronic Respiratory Diseases; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Abou-Hamdan, A.; Guedouari-Bounihi, H.; Lenoir, V.; Andriamihaja, M.; Blachier, F.; Bouillaud, F. Oxidation of H2S in Mammalian Cells and Mitochondria. Methods Enzymol. 2015, 554, 201–228. [Google Scholar] [CrossRef] [PubMed]
- Ramazzini, B. De morbis artificum Bernardini Ramazzini diatribe; University of Chicago Press: Chicago, IL, USA, 1940. [Google Scholar]
- Reiffenstein, R.J.; Hulbert, W.C.; Roth, S.H. Toxicology of hydrogen sulfide. Annu. Rev. Pharmacol. Toxicol. 1992, 32, 109–134. [Google Scholar] [CrossRef]
- Bhatia, M.; Wong, F.L.; Fu, D.; Lau, H.Y.; Moochhala, S.M.; Moore, P.K. Role of hydrogen sulfide in acute pancreatitis and associated lung injury. FASEB J. 2005, 19, 1–17. [Google Scholar] [CrossRef]
- Wu, D.; Luo, N.; Wang, L.; Zhao, Z.; Bu, H.; Xu, G.; Yan, Y.; Che, X.; Jiao, Z.; Zhao, T.; et al. Hydrogen sulfide ameliorates chronic renal failure in rats by inhibiting apoptosis and inflammation through ROS/MAPK and NF-κB signaling pathways. Sci. Rep. 2017, 7, 455. [Google Scholar] [CrossRef]
- Li, L.; Moore, P.K. Could hydrogen sulfide be the next blockbuster treatment for inflammatory disease? Expert Rev. Clin. Pharmacol. 2013, 6, 593–595. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fiorucci, S. Hydrogen sulfide: From physiology to pharmacology. Inflamm. Allergy Drug Targets 2011, 10, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, J.; Li, H.; Xue, M.; Ji, A.; Li, Y. Role of Hydrogen Sulfide in Ischemia-Reperfusion Injury. Oxidative Med. Cell. Longev. 2015, 2015, 1–16. [Google Scholar] [CrossRef]
- Zhong, G.Z.; Chen, F.R.; Cheng, Y.Q.; Tang, C.S.; Du, J.B. The role of hydrogen sulfide generation in the pathogenesis of hypertension in rats induced by inhibition of nitric oxide synthase. J. Hypertens. 2003, 21, 1879–1885. [Google Scholar] [CrossRef]
- Szabo, C.; Coletta, C.; Chao, C.; Módis, K.; Szczesny, B.; Papapetropoulos, A.; Hellmich, M.R. Tumor-derived hydrogen sulfide, produced by cystathionine-β-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 12474–12479. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, A.L.; et al. Cystathionine Beta-Synthase (CBS) Contributes to Advanced Ovarian Cancer Progression and Drug Resistance. PLoS ONE 2013, 8, e79167. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Kawahara, B.; Gupta, D.; Tsai, R.; Khachatryan, M.; Roy-Chowdhuri, S.; Bose, S.; Yoon, A.; Faull, K.; Farias-Eisner, R.; et al. Role of cystathionine β-synthase in human breast Cancer. Free. Radic. Biol. Med. 2015, 86, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Panza, E.; De Cicco, P.; Armogida, C.; Scognamiglio, G.; Gigantino, V.; Botti, G.; Germano, D.; Napolitano, M.; Papapetropoulos, A.; Bucci, M. Role of the cystathionine γ lyase/hydrogen sulfide pathway in human melanoma progression. Pigment Cell Melanoma Res. 2015, 28, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Bian, J. Hydrogen sulfide gas has cell growth regulatory role. Eur. J. Pharmacol. 2011, 656, 5–9. [Google Scholar] [CrossRef]
- Coletta, C.; Papapetropoulos, A.; Erdelyi, K.; Olah, G.; Modis, K.; Panopoulos, P.; Asimakopoulou, A.; Gero, D.; Sharina, I.; Martin, E.; et al. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc. Natl. Acad. Sci. USA 2012, 109, 9161–9166. [Google Scholar] [CrossRef]
- Szabo, C. Gasotransmitters in cancer: From pathophysiology to experimental therapy. Nat. Rev. Drug Discov. 2016, 15, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Módis, K.; Coletta, C.; Asimakopoulou, A.; Szczesny, B.; Chao, C.; Papapetropoulos, A.; Hellmich, M.R.; Szabo, C. Effect of S-adenosyl-L-methionine (SAM), an allosteric activator of cystathionine-β-synthase (CBS) on colorectal cancer cell proliferation and bioenergetics in vitro. Nitric Oxide 2014, 41, 146–156. [Google Scholar] [CrossRef]
- Szabó, C.; Papapetropoulos, A. Hydrogen sulphide and angiogenesis: Mechanisms and applications. Br. J. Pharmacol. 2011, 164, 853–865. [Google Scholar] [CrossRef]
- Szczesny, B.; Módis, K.; Yanagi, K.; Coletta, C.; Le Trionnaire, S.; Perry, A.; Wood, M.E.; Whiteman, M.; Szabo, C. AP39, a novel mitochondria-targeted hydrogen sulfide donor, stimulates cellular bioenergetics, exerts cytoprotective effects and protects against the loss of mitochondrial DNA integrity in oxidatively stressed endothelial cells in vitro. Nitric Oxide 2014, 41, 120–130. [Google Scholar] [CrossRef]
- Wang, M.-J.; Cai, W.-J.; Li, N.; Ding, Y.-J.; Chen, Y.; Zhu, Y.-C. The Hydrogen Sulfide Donor NaHS Promotes Angiogenesis in a Rat Model of Hind Limb Ischemia. Antioxid. Redox Signal. 2010, 12, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Sun, X.; Wang, R. Hydrogen sulfide-induced apoptosis of human aorta smooth muscle cells via the activation of mitogen-activated protein kinases and caspase-3. Faseb J. 2004, 18, 1782–1784. [Google Scholar] [CrossRef]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.-J.; Wang, M.-J.; Moore, P.K.; Jin, H.-M.; Yao, T.; Zhu, Y.-C. The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Jia, Y.; Field, L.; A Ridnour, L.; Sparatore, A.; Del Soldato, P.; Sowers, A.L.; Yeh, G.C.; Moody, T.W.; A Wink, D.; et al. Modulation of angiogenesis by dithiolethione-modified NSAIDs and valproic acid. Br. J. Pharmacol. 2007, 151, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Papapetropoulos, A.; Pyriochou, A.; Altaany, Z.; Yang, G.; Marazioti, A.; Zhou, Z.; Jeschke, M.G.; Branski, L.K.; Herndon, D.N.; Wang, R.; et al. Hydrogen sulfide is an endogenous stimulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21972–21977. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Goto, Y.-I.; Kimura, H. Hydrogen Sulfide Increases Glutathione Production and Suppresses Oxidative Stress in Mitochondria. Antioxid. Redox Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zheng, N.; Ziqiang, S.; Cheng, H.; Sun, Z.; Gao, B.; Zhang, Y.; Pang, W.; Huangfu, C.; Ji, S.; et al. Exogenous hydrogen sulfide mitigates the fatty liver in obese mice through improving lipid metabolism and antioxidant potential. Med. Gas. Res. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.; Yu, W.; Chen, L.; Wang, Z.; Zhang, T. Expression of pulmonary arterial elastin in rats with hypoxic pulmonary hypertension using H2S. J. Recept. Signal. Transduct. 2020, 40, 383–387. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Wu, R.; Geng, B.; Qi, Y.-F.; Wang, P.-P.; Yao, W.-Z.; Tang, C.-S. Endogenous hydrogen sulfide reduces airway inflammation and remodeling in a rat model of asthma. Cytokine 2009, 45, 117–123. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Yao, W.-Z.; Geng, B.; Ding, Y.-L.; Lu, M.; Zhao, M.-W.; Tang, C.-S. Endogenous Hydrogen Sulfide in Patients With COPD. Chest 2005, 128, 3205–3211. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Chen, Y.; Yao, W. Correlation between levels of exhaled hydrogen sulfide and airway inflammatory phenotype in patients with chronic persistent asthma. Respirology 2014, 19, 1165–1169. [Google Scholar] [CrossRef]
- Han, W.; Dong, Z.; Dimitropoulou, C.; Su, Y. Hydrogen Sulfide Ameliorates Tobacco Smoke-Induced Oxidative Stress and Emphysema in Mice. Antioxid. Redox Signal. 2011, 15, 2121–2134. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Ding, Y.; Li, H.; Nie, L.; Yan, X.; Zhou, H.; Zheng, Y. KATP channels of parafacial respiratory group (pFRG) neurons are involved in H2S-mediated central inhibition of respiratory rhythm in medullary slices of neonatal rats. Brain Res. 2013, 1527, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Bilska-Wilkosz, A.; Górny, M. Sulfane sulfur—New findings on an old topic. Acta Biochim. Pol. 2019, 66, 533–544. [Google Scholar] [CrossRef]
- Ishigami, M.; Hiraki, K.; Umemura, K.; Ogasawara, Y.; Ishii, K.; Kimura, H. A Source of Hydrogen Sulfide and a Mechanism of Its Release in the Brain. Antioxid. Redox Signal. 2009, 11, 205–214. [Google Scholar] [CrossRef]
- Toohey, J.I. Sulfur signaling: Is the agent sulfide or sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Toohey, J.I. The conversion of H2S to sulfane sulfur. Nat. Rev. Mol. Cell Biol. 2012, 13, 803. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen sulfide, an enhancer of vascular nitric oxide signaling: Mechanisms and implications. Am. J. Physiol. Physiol. 2017, 312, C3–C15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yu, B.; De La Cruz, L.K.; Choudhury, M.R.; Anifowose, A.; Wang, B. Toward Hydrogen Sulfide Based Therapeutics: Critical Drug Delivery and Developability Issues. Med. Res. Rev. 2018, 38, 57–100. [Google Scholar] [CrossRef]
- Gojon, G.; Morales, G.A. SG1002 and Catenated Divalent Organic Sulfur Compounds as Promising Hydrogen Sulfide Prodrugs. Antioxid. Redox Signal. 2020, 33, 1010–1045. [Google Scholar] [CrossRef] [PubMed]
- Olhemus, D.J.; Li, Z.; Pattillo, C.B.; Gojon, G., Sr.; Gojon, G., Jr.; Giordano, T.; Krum, H. AA Novel Hydrogen Sulfide Prodrug, SG1002, Promotes Hydrogen Sulfide and Nitric Oxide Bioavailability in Heart Failure Patients. Cardiovasc Ther. 2015, 33, 216–226. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Ma, L.; Ji, R.; Qu, Y.; Xin, Y.; Lv, G. Chemopreventive activity of sulforaphane. Drug Des. Dev. Ther. 2018, 12, 2905–2913. [Google Scholar] [CrossRef] [PubMed]
- Szabó, C. Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 2007, 6, 917–935. [Google Scholar] [CrossRef]
- Madden, J.A.; Ahlf, S.B.; Dantuma, M.W.; Olson, K.R.; Roerig, D.L. Precursors and inhibitors of hydrogen sulfide synthesis affect acute hypoxic pulmonary vasoconstriction in the intact lung. J. Appl. Physiol. 2012, 112, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, G.; Wondimu, T.; Ross, B.; Wang, R. Hydrogen sulfide and asthma. Exp. Physiol. 2011, 96, 847–852. [Google Scholar] [CrossRef]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S Signals Through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef]
- Ju, Y.; Untereiner, A.; Wu, L.; Yang, G. H2S-induced S-sulfhydration of pyruvate carboxylase contributes to gluconeogenesis in liver cells. Biochim. Biophys. Acta 2015, 1850, 2293–2303. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, R.A.; Buist, A.S.; Calverley, P.M.; Jenkins, C.R.; Hurd, S.S. Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease: NHLBI/WHO Global Initiative for Chronic Obstructive Lung Disease (GOLD) Workshop summary. Am. J. Respir. Crit. Care. Med. 2001, 163, 1256–1276. [Google Scholar] [CrossRef]
- Wang, L.; Meng, J.; Wang, C.; Yang, C.; Wang, Y.; Li, Y.; Li, Y. Hydrogen sulfide alleviates cigarette smoke-induced COPD through inhibition of the TGF-β1/smad pathway. Exp. Biol. Med. 2020, 245, 190–200. [Google Scholar] [CrossRef]
- Viegas, J.; Esteves, A.F.; Cardoso, E.M.; Arosa, F.A.; Vitale, M.; Taborda-Barata, L. Biological Effects of Thermal Water-Associated Hydrogen Sulfide on Human Airways and Associated Immune Cells: Implications for Respiratory Diseases. Front. Public Heal. 2019, 7, 128. [Google Scholar] [CrossRef]
- Whiteman, M.; Winyard, P.G. Hydrogen sulfide and inflammation: The good, the bad, the ugly and the promising. Expert Rev. Clin. Pharmacol. 2011, 4, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhao, B.; Wang, C.; Wang, H.; Liu, Z.; Li, W.; Jin, H.; Tang, C.; Du, J. Regulatory Effects of Hydrogen Sulfide on IL-6, IL-8 and IL-10 Levels in the Plasma and Pulmonary Tissue of Rats with Acute Lung Injury. Exp. Biol. Med. 2008, 233, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Grommes, J.; Soehnlein, O. Contribution of Neutrophils to Acute Lung Injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Esechie, A.; Kiss, L.; Olah, G.; Horvath, E.; Hawkins, H.; Szabo, C.; Traber, D. Protective effect of hydrogen sulfide in a murine model of combined burn and smoke inhalation-induced acute lung injury. Clin. Sci. 2008, 115, 91–97. [Google Scholar] [CrossRef]
- Rose, P.; Moore, P.K.; Zhu, Y.Z. H2S biosynthesis and catabolism: New insights from molecular studies. Cell. Mol. Life Sci. 2017, 74, 1391–1412. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Wang, P.-P.; Wang, X.-M.; He, Y.-J.; Yao, W.-Z.; Qi, Y.-F.; Tang, C.-S. Involvement of endogenous hydrogen sulfide in cigarette smoke-induced changes in airway responsiveness and inflammation of rat lung. Cytokine 2011, 53, 334–341. [Google Scholar] [CrossRef]
- Roviezzo, F.; Bertolino, A.; Sorrentino, R.; Terlizzi, M.; Matteis, M.; Calderone, V.; Mattera, V.; Martelli, A.; Spaziano, G.; Pinto, A.; et al. Hydrogen sulfide inhalation ameliorates allergen induced airway hypereactivity by modulating mast cell activation. Pharmacol. Res. 2015, 100, 85–92. [Google Scholar] [CrossRef]
- Saito, J.; Mackay, A.J.; Rossios, C.; Gibeon, D.; Macedo, P.; Sinharay, R.; Bhavsar, P.K.; A Wedzicha, J.; Chung, K.F. Sputum-to-serum hydrogen sulfide ratio in COPD. Thorax 2014, 69, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.M.; Tildy, B.; Papi, A.; Casolari, P.; Caramori, G.; Rempel, K.L.; Halayko, A.J.; Adcock, I.; Chung, K.F. The anti-proliferative and anti-inflammatory response of COPD airway smooth muscle cells to hydrogen sulfide. Respir. Res. 2018, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, P.A.; Whiteman, M.; Winyard, P.G.; Caramori, G.; Gordon, F.; Ford, P.A.; Barnes, P.J.; Adcock, I.M.; Chung, K.F. Impact of theophylline/corticosteroid combination therapy on sputum hydrogen sulfide levels in patients with COPD. Eur. Respir. J. 2014, 43, 1504–1506. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Chen, Y.; Yao, W.; Rosner, E.; Mastropietro, C.W. Exhaled Hydrogen Sulfide Predicts Airway Inflammation Phenotype in COPD. Respir. Care 2014, 60, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, S.; Yao, W. Imbalance of Endogenous Hydrogen Sulfide and Homocysteine Pathway in Chronic Obstructive Pulmonary Disease Combined with Cardiovascular Disease. In Proceedings of the American Thoracic Society 2012 International Conference, San Francisco, CA, USA, 18–23 May 2012; American Thoracic Society: New York, NY, USA, 2012; p. 3985. [Google Scholar]
- Fitzgerald, R.; DeSantiago, B.; Lee, D.Y.; Yang, G.; Kim, J.Y.; Foster, D.B.; Chan-Li, Y.; Horton, M.R.; Panettieri, R.A.; Wang, R.; et al. H2S relaxes isolated human airway smooth muscle cells via the sarcolemmal KATP channel. Biochem. Biophys. Res. Commun. 2014, 446, 393–398. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Bernard, G.R. Acute lung injury and the acute respiratory distress syndrome: A clinical review. Lancet 2007, 369, 1553–1564. [Google Scholar] [CrossRef]
- Mackay, A.; Al-Haddad, M. Acute lung injury and acute respiratory distress syndrome. Contin. Educ. Anaesth. Crit. Care Pain 2009, 9, 152–156. [Google Scholar] [CrossRef]
- Ge, X.; Sun, J.; Fei, A.; Gao, C.; Pan, S.; Wu, Z. Hydrogen sulfide treatment alleviated ventilator-induced lung injury through regulation of autophagy and endoplasmic reticulum stress. Int. J. Biol. Sci. 2019, 15, 2872–2884. [Google Scholar] [CrossRef]
- Spassov, S.G.; Donus, R.; Ihle, P.M.; Engelstaedter, H.; Hoetzel, A.; Faller, S. Hydrogen sulfide prevents formation of reactive oxygen species through PI3K/Akt signaling and limits ventilator-induced lung injury. Oxid. Med. Cell. Longev. 2017, 2017, 3715037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-X.; Liu, S.-J.; Tang, X.-L.; Duan, G.-L.; Ni, X.; Zhu, X.-Y.; Liu, Y.-J.; Wang, C.-N. H2S Attenuates LPS-Induced Acute Lung Injury by Reducing Oxidative/Nitrative Stress and Inflammation. Cell. Physiol. Biochem. 2016, 40, 1603–1612. [Google Scholar] [CrossRef]
- Faller, S.; Zimmermann, K.K.; Strosing, K.M.; Engelstaedter, H.; Buerkle, H.; Schmidt, R.; Spassov, S.G.; Hoetzel, A. Inhaled hydrogen sulfide protects against lipopolysaccharide-induced acute lung injury in mice. Med. Gas. Res. 2012, 2, 26. [Google Scholar] [CrossRef]
- Zimmermann, K.K.; Spassov, S.G.; Strosing, K.M.; Ihle, P.M.; Engelstaedter, H.; Hoetzel, A.; Faller, S. Hydrogen Sulfide Exerts Anti-oxidative and Anti-inflammatory Effects in Acute Lung Injury. Inflammation 2017, 41, 249–259. [Google Scholar] [CrossRef]
- Han, Z.-H.; Jiang, Y.; Duan, Y.-Y.; Wang, X.-Y.; Huang, Y.; Fang, T.-Z. Protective effects of hydrogen sulfide inhalation on oxidative stress in rats with cotton smoke inhalation-induced lung injury. Exp. Ther. Med. 2015, 10, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.Y.C.; Chen, W.; Li, X.L.; Wang, Y.W.; Xie, X.H. H₂S protecting against lung injury following limb ischemia-reperfusion by alleviating inflammation and water transport abnormality in rats. Biomed. Environ. Sci. 2014, 27, 410–418. [Google Scholar] [PubMed]
- Sakaguchi, M.; Marutani, E.; Shin, H.-s.; Chen, W.; Hanaoka, K.; Xian, M.; Ichinose, F. Sodium thiosulfate attenuates acute lung injury in mice. Anesthesiology 2014, 121, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Spassov, S.; Pfeifer, D.; Strosing, K.; Ryter, S.; Hummel, M.; Faller, S.; Hoetzel, A. Genetic Targets of Hydrogen Sulfide in Ventilator-Induced Lung Injury—A Microarray Study. PLoS ONE 2014, 9, e102401. [Google Scholar] [CrossRef]
- Geng, P.; Ma, T.; Xing, J.; Jiang, L.; Sun, H.; Zhu, B.; Zhang, H.; Xiao, H.; Wang, J.; Zhang, J. Dexamethasone ameliorates H2S-induced acute lung injury by increasing claudin-5 expression via the PI3K pathway. Hum. Exp. Toxicol. 2017, 37, 626–635. [Google Scholar] [CrossRef]
- Ning, J.; Mo, L.; Zhao, H.; Lu, K.; Lai, X.; Luo, X.; Zhao, H.; Ma, D. Sodium Hydrosulphide Alleviates Remote Lung Injury Following Limb Traumatic Injury in Rats. PLoS ONE 2013, 8, e59100. [Google Scholar] [CrossRef] [PubMed]
- Vadivel, A.; Alphonse, R.S.; Ionescu, L.; Machado, D.S.; O’Reilly, M.; Eaton, F.; Haromy, A.; Michelakis, E.D.; Thébaud, B. Exogenous Hydrogen Sulfide (H2S) Protects Alveolar Growth in Experimental O2-Induced Neonatal Lung Injury. PLoS ONE 2014, 9, e90965. [Google Scholar] [CrossRef]
- Liu, Z.-W.; Wang, H.-Y.; Guan, L.; Zhao, B. Regulatory effects of hydrogen sulfide on alveolar epithelial cell endoplasmic reticulum stress in rats with acute lung injury. World J. Emerg. Med. 2015, 6, 67–73. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Su, C.; Chen, J.; Zhu, B.; Zhang, H.; Xiao, H.; Zhang, J. Dexamethasone Ameliorates H2S-Induced Acute Lung Injury by Alleviating Matrix Metalloproteinase-2 and -9 Expression. PLoS ONE 2014, 9, e94701. [Google Scholar] [CrossRef] [PubMed]
- Aslami, H.; Heinen, A.; Roelofs, J.J.T.H.; Zuurbier, C.J.; Schultz, M.J.; Juffermans, N.P. Suspended animation inducer hydrogen sulfide is protective in an in vivo model of ventilator-induced lung injury. Intensiv. Care Med. 2010, 36, 1946–1952. [Google Scholar] [CrossRef] [PubMed]
- Shields, C.J.; Winter, D.C.; Redmond, H.P. Lung injury in acute pancreatitis: Mechanisms, prevention, and therapy. Curr. Opin. Crit. Care 2002, 8, 158–163. [Google Scholar] [CrossRef]
- Bhatia, M.; Sidhapuriwala, J.N.; Sparatore, A.; Moore, P.K. Treatment with H2S-releasing diclofenac protects mice against acute pancreatitis-associated lung injury. Shock 2008, 29, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Tamizhselvi, R.; Moore, P.K.; Bhatia, M. Inhibition of Hydrogen Sulfide Synthesis Attenuates Chemokine Production and Protects Mice against Acute Pancreatitis and Associated Lung Injury. Pancreas 2008, 36, e24–e31. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Pei, Z.; Feng, Y.; Zhao, J.; Chen, Y.; Shi, W.; Xu, Q.; Lin, F.; Sun, M.; Xiao, K. Neglected role of hydrogen sulfide in sulfur mustard poisoning: Keap1 S-sulfhydration and subsequent Nrf2 pathway activation. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Zhi, L.; Zhang, H.; Ng, S.-W.; Moore, P.K. Role of substance P in hydrogen sulfide-induced pulmonary inflammation in mice. Am. J. Physiol. Cell. Mol. Physiol. 2006, 291, L896–L904. [Google Scholar] [CrossRef]
- Sidhapuriwala, J.N.; Ng, S.W.; Bhatia, M. Effects of hydrogen sulfide on inflammation in caerulein-induced acute pancreatitis. J. Inflamm. 2009, 6, 35. [Google Scholar] [CrossRef]
- Esechie, A.; Enkhbaatar, P.; Traber, D.L.; Jonkam, C.; Lange, M.; Hamahata, A.; Djukom, C.; Whorton, E.B.; Hawkins, H.K.; Traber, L.D. Beneficial effect of a hydrogen sulphide donor (sodium sulphide) in an ovine model of burn-and smoke-induced acute lung injury. Br. J. Pharmacol. 2009, 158, 1442–1453. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.J.; Ng, M.Y.; Lee, Z.-W.; Dai, W.; Hagen, T.; Moore, P.K.; Huang, D.; Deng, L.-W.; Tan, C.-H. Hydrogen sulfide donors in research and drug development. Med. Chem. Comm. 2014, 5, 557–570. [Google Scholar] [CrossRef]
- Ahmad, A.; Szabo, C. Both the H 2 S biosynthesis inhibitor aminooxyacetic acid and the mitochondrially targeted H 2 S donor AP39 exert protective effects in a mouse model of burn injury. Pharmacol. Res. 2016, 113, 348–355. [Google Scholar] [CrossRef]
- Faller, S.; Ryter, S.W.; Choi, A.M.K.; Loop, T.; Schmidt, R.; Hoetzel, A. Inhaled Hydrogen Sulfide Protects against Ventilator-induced Lung Injury. Anesthesiology 2010, 113, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.C.; Vaporidi, K.; Bloch, K.D.; Ichinose, F.; Zapol, W.M. Protective and detrimental effects of sodium sulfide and hydrogen sulfide in murine ventilator-induced lung injury. Anesthesiology 2011, 115, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Asher, I.; Pearce, N. Global burden of asthma among children. Int. J. Tuberc. Lung Dis. 2014, 18, 1269–1278. [Google Scholar] [CrossRef]
- Derwall, M.; Francis, R.C.; Kida, K.; Bougaki, M.; Crimi, E.; Adrie, C.; Zapol, W.M.; Ichinose, F. Administration of hydrogen sulfide via extracorporeal membrane lung ventilation in sheep with partial cardiopulmonary bypass perfusion: A proof of concept study on metabolic and vasomotor effects. Crit. Care 2011, 15, R51. [Google Scholar] [CrossRef]
- Perry, M.M.; Hui, C.; Whiteman, M.; Wood, M.E.; Adcock, I.; Kirkham, P.; Chung, K.F. Hydrogen Sulfide Inhibits Proliferation and Release of IL-8 from Human Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2011, 45, 746–752. [Google Scholar] [CrossRef]
- Tian, M.; Wang, Y.; Lu, Y.-Q.; Yan, M.; Jiang, Y.-H.; Zhao, D.-Y. Correlation between serum H2S and pulmonary function in children with bronchial asthma. Mol. Med. Rep. 2012, 6, 335–338. [Google Scholar] [CrossRef]
- Zhang, P.; Li, F.; Wiegman, C.H.; Zhang, M.; Hong, Y.; Gong, J.; Chang, Y.; Zhang, J.; Adcock, I.; Chung, K.F.; et al. Inhibitory Effect of Hydrogen Sulfide on Ozone-Induced Airway Inflammation, Oxidative Stress, and Bronchial Hyperresponsiveness. Am. J. Respir. Cell Mol. Biol. 2015, 52, 129–137. [Google Scholar] [CrossRef]
- Ma, Y.; Yan, Z.; Deng, X.; Guo, J.; Hu, J.; Yu, Y.; Jiao, F. Anticancer effect of exogenous hydrogen sulfide in cisplatin-resistant A549/DDP cells. Oncol. Rep. 2018, 39, 2969–2977. [Google Scholar] [CrossRef]
- Bhatia, M. Role of Hydrogen Sulfide in the Pathology of Inflammation. Scientifica 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Fiorucci, S.; Orlandi, S.; Mencarelli, A.; Caliendo, G.; Santagada, V.; Distrutti, E.; Santucci, L.; Cirino, G.; Wallace, J.L. Enhanced activity of a hydrogen sulphide-releasing derivative of mesalamine (ATB-429) in a mouse model of colitis. Br. J. Pharmacol. 2007, 150, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-Q.; Gao, C.; Niu, W.; Li, Y.; Wang, Y.-X.; Gao, C.-J.; Ding, Q.; Yao, L.-N.; Chai, W.; Li, Z.-C. Sodium hydrosulfide alleviates lung inflammation and cell apoptosis following resuscitated hemorrhagic shock in rats. Acta Pharmacol. Sin. 2013, 34, 1515–1525. [Google Scholar] [CrossRef]
- Bhatia, M. H2S and Inflammation: An Overview. Handb. Exp. Pharmacol. 2015, 230, 165–180. [Google Scholar] [CrossRef]
- Guo, F.; Yu, T.; Hong, J.; Fang, J. Emerging Roles of Hydrogen Sulfide in Inflammatory and Neoplastic Colonic Diseases. Front. Physiol. 2016, 7, 156. [Google Scholar] [CrossRef]
- Saglam, K.; Alhan, E.; Turkyilmaz, S.; Vanizor, B.K.; Ercin, C. The anti-inflammatory effect of hydrogen sulphide on acute necrotizing pancreatitis in rats. Turk. J. Surg. 2017, 33, 158–163. [Google Scholar] [CrossRef]
- Campos, D.; Ravagnani, F.G.; Gurgueira, S.A.; Vercesi, A.E.; Teixeira, S.A.; Costa, S.K.; Muscará, M.N.; Ferreira, H.H. Increased glutathione levels contribute to the beneficial effects of hydrogen sulfide and inducible nitric oxide inhibition in allergic lung inflammation. Int. Immunopharmacol. 2016, 39, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Aslami, H.; Pulskens, W.P.; Kuipers, M.T.; Bos, A.P.; Van Kuilenburg, A.B.P.; Wanders, R.J.A.; Roelofsen, J.; Roelofs, J.J.T.H.; Kerindongo, R.P.; Beurskens, C.J.P.; et al. Hydrogen Sulfide Donor NaHS Reduces Organ Injury in a Rat Model of Pneumococcal Pneumosepsis, Associated with Improved Bio-Energetic Status. PLoS ONE 2013, 8, e63497. [Google Scholar] [CrossRef] [PubMed]
- Großhennig, S.; Ischebeck, T.; Gibhardt, J.; Busse, J.; Feussner, I.; Stülke, J. Hydrogen sulfide is a novel potential virulence factor of M ycoplasma pneumoniae: Characterization of the unusual cysteine desulfurase/desulfhydrase HapE. Mol. Microbiol. 2016, 100, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Oliver, E.; Ma, Y.; Escaffre, O.; Ivanciuc, T.; Komaravelli, N.; Kelley, J.P.; Coletta, C.; Szabo, C.; Rockx, B.; Garofalo, R.P.; et al. Role of Hydrogen Sulfide in Paramyxovirus Infections. J. Virol. 2015, 89, 5557–5568. [Google Scholar] [CrossRef]
- Cowley, E.S.; Kopf, S.H.; Lariviere, A.; Ziebis, W.; Newman, D.K. Pediatric Cystic Fibrosis Sputum Can Be Chemically Dynamic, Anoxic, and Extremely Reduced Due to Hydrogen Sulfide Formation. mBio 2015, 6, e00767-15. [Google Scholar] [CrossRef] [PubMed]
- Gaddam, R.R.; Fraser, R.; Badiei, A.; Chambers, S.; Cogger, V.C.; Le Couteur, D.G.; Ishii, I.; Bhatia, M. Cystathionine-Gamma-Lyase Gene Deletion Protects Mice against Inflammation and Liver Sieve Injury following Polymicrobial Sepsis. PLoS ONE 2016, 11, e0160521. [Google Scholar] [CrossRef]
- Zhang, H.; Zhi, L.; Moochhala, S.; Moore, P.K.; Bhatia, M. Hydrogen sulfide acts as an inflammatory mediator in cecal ligation and puncture-induced sepsis in mice by upregulating the production of cytokines and chemokines via NF-κB. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, L960–L971. [Google Scholar] [CrossRef] [PubMed]
- El-Chemaly, S.; Levine, S.J.; Moss, J. Lymphatics in Lung Disease. Ann. N. Y. Acad. Sci. 2008, 1131, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Reed, H.O.; Wang, L.; Sonett, J.; Chen, M.; Yang, J.; Li, L.; Aradi, P.; Jakus, Z.; D’Armiento, J.M.; Hancock, W.W.; et al. Lymphatic impairment leads to pulmonary tertiary lymphoid organ formation and alveolar damage. J. Clin. Investig. 2019, 129, 2514–2526. [Google Scholar] [CrossRef] [PubMed]
- Erb, A.; Althaus, M. Actions of Hydrogen Sulfide on Sodium Transport Processes across Native Distal Lung Epithelia (Xenopus laevis). PLoS ONE 2014, 9, e100971. [Google Scholar] [CrossRef] [PubMed]
- Althaus, M.; Clauss, W.G.; Fronius, M. Amiloride-Sensitive Sodium Channels and Pulmonary Edema. Pulm. Med. 2010, 2011, 1–8. [Google Scholar] [CrossRef]
- Egli, M.; Duplain, H.; Lepori, M.; Cook, S.; Nicod, P.; Hummler, E.; Sartori, C.; Scherrer, U. Defective respiratory amiloride-sensitive sodium transport predisposes to pulmonary oedema and delays its resolution in mice. J. Physiol. 2004, 560, 857–865. [Google Scholar] [CrossRef]
- Kubo, S.; Doe, I.; Kurokawa, Y.; Kawabata, A. Hydrogen sulfide causes relaxation in mouse bronchial smooth muscle. J. Pharmacol. Sci. 2007, 104, 392–396. [Google Scholar] [CrossRef]
- Gharib-Naseri, M.K.; Saberi, S.; Mard, S.A.; Latifi, S.M. Bronchodilatory Effect of Hydrogen Sulfide in Rat. Iran. J. Basic Med. Sci 2012, 15, 907–915. [Google Scholar]
- Zhang, Y.-L.; Chen, P.-X.; Guan, W.-J.; Guo, H.-M.; Qiu, Z.-E.; Xu, J.-W.; Luo, Y.-L.; Lan, C.-F.; Xu, J.-B.; Hao, Y.; et al. Increased intracellular Cl− concentration promotes ongoing inflammation in airway epithelium. Mucosal Immunol. 2018, 11, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; An, G.; Chen, J. Inhibitory effects of hydrogen sulphide on pulmonary fibrosis in smoking rats via attenuation of oxidative stress and inflammation. J. Cell. Mol. Med. 2014, 18, 1098–1103. [Google Scholar] [CrossRef]
- Song, K.; Li, Q.; Yin, X.-Y.; Lu, Y.; Liu, C.-F.; Hu, L.-F. Hydrogen Sulfide: A Therapeutic Candidate for Fibrotic Disease? Oxidative Med. Cell. Longev. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Zhou, X.; Cao, Y.; Ao, G.; Hu, L.; Liu, H.; Wu, J.; Wang, X.; Jin, M.; Zheng, S.; Zhen, X. CaMKKβ-dependent activation of AMP-activated protein kinase is critical to suppressive effects of hydrogen sulfide on neuroinflammation. Antioxid. Redox Signal. 2014, 21, 1741–1758. [Google Scholar] [CrossRef]
- Wang, Z.; Yin, X.; Gao, L.; Feng, S.; Song, K.; Li, L.; Lu, Y.; Shen, H. The protective effect of hydrogen sulfide on systemic sclerosis associated skin and lung fibrosis in mice model. SpringerPlus 2016, 5, 1084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pan, C.; Zhou, F.; Yuan, Z.; Wang, H.; Cui, W.; Zhang, G. Hydrogen Sulfide as a Potential Therapeutic Target in Fibrosis. Oxidative Med. Cell. Longev. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, M.C.; Scherag, A.; Adhikari, N.K.J.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K. Assessment of Global Incidence and Mortality of Hospital-treated Sepsis. Current Estimates and Limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef]
- Ang, S.-F.; Moochhala, S.M.; MacAry, P.A.; Bhatia, M. Hydrogen sulfide and neurogenic inflammation in polymicrobial sepsis: Involvement of substance P and ERK-NF-κB signaling. PLoS ONE 2011, 6, e24535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hegde, A.; Ng, S.W.; Adhikari, S.; Moochhala, S.M.; Bhatia, M. Hydrogen sulfide up-regulates substance P in polymicrobial sepsis-associated lung injury. J. Immunol. 2007, 179, 4153–4160. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Yang, W.; Zhang, H.; Song, Z.; Liu, T.; Lv, X. Hydrogen Sulfide Ameliorates Lung Ischemia-Reperfusion Injury Through SIRT1 Signaling Pathway in Type 2 Diabetic Rats. Front. Physiol. 2020, 11, 596. [Google Scholar] [CrossRef] [PubMed]
- George, T.J.; Arnaoutakis, G.J.; Beaty, C.A.; Jandu, S.K.; Santhanam, L.; Berkowitz, D.E.; Shah, A.S. Inhaled hydrogen sulfide improves graft function in an experimental model of lung transplantation. J. Surg. Res. 2012, 178, 593–600. [Google Scholar] [CrossRef][Green Version]
- Wu, J.-X.; Wei, J.; You, X.; Chen, X.; Zhu, H.; Zhu, X.; Liu, Y.; Xu, M. Inhibition of hydrogen sulfide generation contributes to lung injury after experimental orthotopic lung transplantation. J. Surg. Res. 2013, 182, e25–e33. [Google Scholar] [CrossRef] [PubMed]
- Wanamaker, B.; Cascino, T.; McLaughlin, V.; Oral, H.; Latchamsetty, R.; Siontis, K.C. Atrial Arrhythmias in Pulmonary Hypertension: Pathogenesis, Prognosis and Management. Arrhythmia Electrophysiol. Rev. 2018, 7, 43–48. [Google Scholar] [CrossRef]
- Xiaohui, L.; Junbao, D.; Lin, S.; Jian, L.; Xiuying, T.; Jianguang, Q.; Bing, W.; Hongfang, J.; Chaoshu, T. Down-Regulation of Endogenous Hydrogen Sulfide Pathway in Pulmonary Hypertension and Pulmonary Vascular Structural Remodeling Induced by High Pulmonary Blood Flow in Rats. Circ. J. 2005, 69, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Pan, W.; Wang, C.; Dong, H.; Xing, L.; Hou, J.; Fang, S.; Li, H.; Yang, F.; Yu, B. H2S attenuates endoplasmic reticulum stress in hypoxia-induced pulmonary artery hypertension. Biosci. Rep. 2019, 39, BSR20190304. [Google Scholar] [CrossRef]
- Wei, H.-L.; Zhang, C.-Y.; Jin, H.-F.; Tang, C.-S.; Du, J.-B. Hydrogen sulfide regulates lung tissue-oxidized glutathione and total antioxidant capacity in hypoxic pulmonary hypertensive rats. Acta Pharmacol. Sin. 2008, 29, 670–676. [Google Scholar] [CrossRef]
- Peng, Y.-J.; Zhang, X.; Gridina, A.; Chupikova, I.; McCormick, D.L.; Thomas, R.J.; Scammell, T.E.; Kim, G.; Vasavda, C.; Nanduri, J.; et al. Complementary roles of gasotransmitters CO and H2S in sleep apnea. Proc. Natl. Acad. Sci. USA 2017, 114, 1413–1418. [Google Scholar] [CrossRef]
- Yuan, G.; Peng, Y.-J.; Khan, S.A.; Nanduri, J.; Singh, A.; Vasavda, C.; Semenza, G.L.; Kumar, G.K.; Snyder, S.H.; Prabhakar, N.R. H2S production by reactive oxygen species in the carotid body triggers hypertension in a rodent model of sleep apnea. Sci. Signal. 2016, 9, ra80. [Google Scholar] [CrossRef] [PubMed]
- Claudet, I.; Marcoux, M.-O.; Karsenty, C.; Rittié, J.-L.; Honorat, R.; Lelong-Tissier, M. Intoxication accidentelle grave à l’hydrogène sulfuré: Un cas pédiatrique de survie. Ann. Françaises D’anesthésie Et De Réanimation 2012, 31, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Muzaffar, S.; Jeremy, J.Y.; Sparatore, A.; Del Soldato, P.; Angelini, G.D.; Shukla, N. H2 S-donating sildenafil (ACS6) inhibits superoxide formation and gp91phox expression in arterial endothelial cells: Role of protein kinases A and G. Br. J. Pharmacol. 2008, 155, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, X. The effects of gasotransmitters on bronchopulmonary dysplasia. Eur. J. Pharmacol. 2020, 873, 172983. [Google Scholar] [CrossRef] [PubMed]
- Madurga, A.; Mižíková, I.; Ruiz-Camp, J.; Vadász, I.; Herold, S.; Mayer, K.; Fehrenbach, H.; Seeger, W.; Morty, R.E. Systemic hydrogen sulfide administration partially restores normal alveolarization in an experimental animal model of bronchopulmonary dysplasia. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L684–L697. [Google Scholar] [CrossRef]
- Calderone, V.; Martelli, A.; Testai, L.; Citi, V.; Breschi, M.C. Using hydrogen sulfide to design and develop drugs. Expert Opin. Drug Discov. 2015, 11, 163–175. [Google Scholar] [CrossRef]
- Citi, V.; Piragine, E.; Testai, L.; Breschi, M.C.; Calderone, V.; Martelli, A. The Role of Hydrogen Sulfide and H2S-donors in Myocardial Protection against Ischemia/Reperfusion Injury. Curr. Med. Chem. 2018, 25, 4380–4401. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, P.; Yang, G.; Cao, Q.; Wang, R. The Inhibitory Role of Hydrogen Sulfide in Airway Hyperresponsiveness and Inflammation in a Mouse Model of Asthma. Am. J. Pathol. 2013, 182, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Heer, J.; Garle, M.; Alexander, S.; Roberts, R. Hydrogen sulphide-induced relaxation of porcine peripheral bronchioles. Br. J. Pharmacol. 2013, 168, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Castro-Piedras, I.; Perez-Zoghbi, J.F. Hydrogen sulphide inhibits Ca2+release through InsP3receptors and relaxes airway smooth muscle. J. Physiol. 2013, 591, 5999–6015. [Google Scholar] [CrossRef]
- Huang, J.; Luo, Y.-L.; Hao, Y.; Zhang, Y.-L.; Chen, P.-X.; Xu, J.-W.; Chen, M.; Luo, Y.-F.; Zhong, N.-S.; Xu, J.; et al. Cellular mechanism underlying hydrogen sulfide induced mouse tracheal smooth muscle relaxation: Role of BKCa. Eur. J. Pharmacol. 2014, 741, 55–63. [Google Scholar] [CrossRef]
- Hosoki, R.; Matsuki, N.; Kimura, H. The Possible Role of Hydrogen Sulfide as an Endogenous Smooth Muscle Relaxant in Synergy with Nitric Oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Mainali, P.; Tang, C.-S.; Shi, L.; Zhang, C.-Y.; Yan, H.; Liu, X.-Q.; Du, J.-B. Effects of nitric oxide and hydrogen sulfide on the relaxation of pulmonary arteries in rats. Chin. Med. J. 2008, 121, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Costantino, M.; Lampa, E.; Nappi, G. Effectiveness of sulphur spa therapy with politzer in the treatment of rhinogenic deafness. Acta Otorhinolaryngol. Ital. 2006, 26, 7–13. [Google Scholar]
- Pouokam, E.; Althaus, M. Epithelial Electrolyte Transport Physiology and the Gasotransmitter Hydrogen Sulfide. Oxidative Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Faller, S.; Hausler, F.; Goeft, A.; Von Itter, M.-N.A.; Gyllenram, V.; Hoetzel, A.; Spassov, S.G. Hydrogen sulfide limits neutrophil transmigration, inflammation, and oxidative burst in lipopolysaccharide-induced acute lung injury. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, X.; Chen, S.; Chen, S.; Yu, W.; Liu, X.; Yang, G.; Tao, Y.; Tang, X.; Bu, D.; et al. Endogenous hydrogen sulfide sulfhydrates IKKβ at cysteine 179 to control pulmonary artery endothelial cell inflammation. Clin. Sci. 2019, 133, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, R.C.O.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. Faseb J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Brancaleone, V.; Mitidieri, E.; Flower, R.J.; Cirino, G.; Perretti, M. Annexin A1 Mediates Hydrogen Sulfide Properties in the Control of Inflammation. J. Pharmacol. Exp. Ther. 2014, 351, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Pugliesi, I.; Barresi, E.; Nesi, G.; Rapposelli, S.; Taliani, S.; Da Settimo, F.; et al. Arylthioamides as H2S Donors: L-Cysteine-Activated Releasing Properties and Vascular effects in vitro and in vivo. ACS Med. Chem. Lett. 2013, 4, 904–908. [Google Scholar] [CrossRef]
- Liu, Y.; Li, A.; Feng, X.; Sun, X.; Zhu, X.; Zhao, Z. Pharmacological Investigation of the Anti-Inflammation and Anti-Oxidation Activities of Diallyl Disulfide in a Rat Emphysema Model Induced by Cigarette Smoke Extract. Nutrients 2018, 10, 79. [Google Scholar] [CrossRef]
- Lucarini, E.; Micheli, L.; Trallori, E.; Citi, V.; Martelli, A.; Testai, L.; De Nicola, G.R.; Iori, R.; Calderone, V.; Ghelardini, C.; et al. Effect of glucoraphanin and sulforaphane against chemotherapy-induced neuropathic pain: Kv7 potassium channels modulation by H2 S release in vivo. Phytother. Res. 2018, 32, 2226–2234. [Google Scholar] [CrossRef]
- Citi, V.; Corvino, A.; Fiorino, F.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Brogi, S.; Flori, L.; Gorica, E.; et al. Structure-activity relationships study of isothiocyanates for H2S releasing properties: 3-Pyridyl-isothiocyanate as a new promising cardioprotective agent. J. Adv. Res. 2021, 27, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Martelli, A.; Gorica, E.; Brogi, S.; Testai, L.; Calderone, V. Role of hydrogen sulfide in endothelial dysfunction: Pathophysiology and therapeutic approaches. J. Adv. Res. 2021, 27, 99–113. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Testai, L.; Marino, A.; Breschi, M.C.; Calderone, V. Hydrogen Sulfide Releasing Capacity of Natural Isothiocyanates: Is It a Reliable Explanation for the Multiple Biological Effects of Brassicaceae? Planta Med. 2014, 80, 610–613. [Google Scholar] [CrossRef]
- Citi, V.; Piragine, E.; Pagnotta, E.; Ugolini, L.; Mannelli, L.D.C.; Testai, L.; Ghelardini, C.; Lazzeri, L.; Calderone, V.; Martelli, A. Anticancer properties of erucin, an H2S-releasing isothiocyanate, on human pancreatic adenocarcinoma cells (AsPC-1). Phytother. Res. 2019, 33, 845–855. [Google Scholar] [CrossRef]
- Martelli, A.; Citi, V.; Testai, L.; Brogi, S.; Calderone, V. Organic Isothiocyanates as Hydrogen Sulfide Donors. Antioxid. Redox Signal. 2020, 32, 110–144. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Piragine, E.; Citi, V.; Testai, L.; Pagnotta, E.; Ugolini, L.; Lazzeri, L.; Mannelli, L.D.C.; Manzo, O.L.; Bucci, M.; et al. Erucin exhibits vasorelaxing effects and antihypertensive activity by H2S-releasing properties. Br. J. Pharmacol. 2020, 177, 824–835. [Google Scholar] [CrossRef]
- Sestito, S.; Pruccoli, L.; Runfola, M.; Citi, V.; Martelli, A.; Saccomanni, G.; Calderone, V.; Tarozzi, A.; Rapposelli, S. Design and synthesis of H2S-donor hybrids: A new treatment for Alzheimer’s disease? Eur. J. Med. Chem. 2019, 184, 111745. [Google Scholar] [CrossRef]
- Takayama, K.; Hirose, A.; Suda, I.; Miyazaki, A.; Oguchi, M.; Onotogi, M.; Fotopoulos, G. Comparison of the Anti-Inflammatory and Analgesic Effects in Rats of Diclofenac-Sodium, Felbinac and Indomethacin Patches. Int. J. Biomed. Sci. 2011, 7, 222–229. [Google Scholar]
- Jia, G.; Yu, S.; Sun, W.; Yang, J.; Wang, Y.; Qi, Y.; Chen, Y. Hydrogen Sulfide Attenuates Particulate Matter-Induced Emphysema and Airway Inflammation Through Nrf2-Dependent Manner. Front. Pharmacol. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- McCook, O.; Radermacher, P.; Volani, C.; Asfar, P.; Ignatius, A.; Kemmler, J.; Möller, P.; Szabó, C.; Whiteman, M.; Wood, M.E.; et al. H2S during circulatory shock: Some unresolved questions. Nitric Oxide 2014, 41, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Pálinkás, Z.; Nagy, A.; Budai, B.; Tóth, I.; Vasas, A. Chemical aspects of hydrogen sulfide measurements in physiological samples. Biochim. Biophys. Acta 2014, 1840, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Du, X.; Wang, D.; Hao, F. Myocardial and lung injuries induced by hydrogen sulfide and the effectiveness of oxygen therapy in rats. Clin. Toxicol. 2011, 49, 161–166. [Google Scholar] [CrossRef]
- Olson, K.R.; DeLeon, E.R.; Gao, Y.; Hurley, K.; Sadauskas, V.; Batz, C.; Stoy, G.F. Thiosulfate: A readily accessible source of hydrogen sulfide in oxygen sensing. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R592–R603. [Google Scholar] [CrossRef]
- Szabo, C.; Papapetropoulos, A. International Union of Basic and Clinical Pharmacology. CII: Pharmacological Modulation of H2S Levels: H2S Donors and H2S Biosynthesis Inhibitors. Pharmacol. Rev. 2017, 69, 497–564. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yang, C.-T.; Meng, F.-H.; Pacheco, A.; Chen, L.; Xian, M. Ammonium tetrathiomolybdate as a water-soluble and slow-release hydrogen sulfide donor. Bioorganic Med. Chem. Lett. 2016, 26, 1585–1588. [Google Scholar] [CrossRef] [PubMed]
- Dyson, A.; Dal-Pizzol, F.; Sabbatini, G.; Lach, A.B.; Galfo, F.; Cardoso, J.D.S.; Mendonça, B.P.; Hargreaves, I.; Pinto, B.B.; Bromage, D.I.; et al. Ammonium tetrathiomolybdate following ischemia/reperfusion injury: Chemistry, pharmacology, and impact of a new class of sulfide donor in preclinical injury models. PLoS Med. 2017, 14, e1002310. [Google Scholar] [CrossRef]
- Donnarumma, E.; Ali, M.J.; Rushing, A.M.; Scarborough, A.L.; Bradley, J.M.; Organ, C.L.; Islam, K.N.; Polhemus, D.J.; Evangelista, S.; Cirino, G.; et al. Zofenopril Protects Against Myocardial Ischemia–Reperfusion Injury by Increasing Nitric Oxide and Hydrogen Sulfide Bioavailability. J. Am. Hear. Assoc. 2016, 5, 003531. [Google Scholar] [CrossRef]
- Neuwelt, E.A.; Gilmer-Knight, K.; Lacy, C.; Nicholson, H.S.; Kraemer, D.F.; Doolittle, N.D.; Hornig, G.W.; Muldoon, L.L. Toxicity profile of delayed high dose sodium thiosulfate in children treated with carboplatin in conjunction with blood-brain-barrier disruption. Pediatr. Blood Cancer 2006, 47, 174–182. [Google Scholar]
- Marutani, E.; Yamada, M.; Ida, T.; Tokuda, K.; Ikeda, K.; Kai, S.; Shirozu, K.; Hayashida, K.; Kosugi, S.; Hanaoka, K.; et al. Thiosulfate Mediates Cytoprotective Effects of Hydrogen Sulfide Against Neuronal Ischemia. J. Am. Heart Assoc. 2015, 4, e002125. [Google Scholar] [CrossRef]
- Wallace, J.L.; Caliendo, G.; Santagada, V.; Cirino, G. Markedly reduced toxicity of a hydrogen sulphide-releasing derivative of naproxen (ATB-346). Br. J. Pharmacol. 2010, 159, 1236–1246. [Google Scholar] [CrossRef]
- Ekundi-Valentim, E.; Mesquita, F.P.; Santos, K.T.; Paula, M.A.V.D.; Florenzano, J.; I Zanoni, C.; Rodrigues, L.; De Nucci, G.; A Teixeira, S.; Ferreira, H.H.; et al. A comparative study on the anti-inflammatory effects of single oral doses of naproxen and its hydrogen sulfide (H2S)-releasing derivative ATB-346 in rats with carrageenan-induced synovitis. Med. Gas. Res. 2013, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Nagy, P.; Feener, T.D.; Allain, T.; Ditrói, T.; Vaughan, D.J.; Muscara, M.N.; De Nucci, G.; Buret, A.G. A proof-of-concept, Phase 2 clinical trial of the gastrointestinal safety of a hydrogen sulfide-releasing anti-inflammatory drug. Br. J. Pharmacol. 2020, 177, 769–777. [Google Scholar] [CrossRef]
- Kashfi, K.; Chattopadhyay, M.; Kodela, R. NOSH-sulindac (AVT-18A) is a novel nitric oxide- and hydrogen sulfide-releasing hybrid that is gastrointestinal safe and has potent anti-inflammatory, analgesic, antipyretic, anti-platelet, and anti-cancer properties. Redox Biol. 2015, 6, 287–296. [Google Scholar] [CrossRef]
- Kodela, R.; Chattopadhyay, M.; Velázquez-Martínez, C.A.; Kashfi, K. NOSH-aspirin (NBS-1120), a novel nitric oxide- and hydrogen sulfide-releasing hybrid has enhanced chemo-preventive properties compared to aspirin, is gastrointestinal safe with all the classic therapeutic indications. Biochem. Pharmacol. 2015, 98, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Rose, P.; Tan, C.-H.; Parkinson, D.B.; Moore, P.K. The Effect of Hydrogen Sulfide Donors on Lipopolysaccharide-Induced Formation of Inflammatory Mediators in Macrophages. Antioxid. Redox Signal. 2010, 12, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Salto-Tellez, M.; Tan, C.-H.; Whiteman, M.; Moore, P.K. GYY4137, a novel hydrogen sulfide-releasing molecule, protects against endotoxic shock in the rat. Free Radic. Biol. Med. 2009, 47, 103–113. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khattak, S.; Zhang, Q.-Q.; Sarfraz, M.; Muhammad, P.; Ngowi, E.E.; Khan, N.H.; Rauf, S.; Wang, Y.-Z.; Qi, H.-W.; Wang, D.; et al. The Role of Hydrogen Sulfide in Respiratory Diseases. Biomolecules 2021, 11, 682. https://doi.org/10.3390/biom11050682
Khattak S, Zhang Q-Q, Sarfraz M, Muhammad P, Ngowi EE, Khan NH, Rauf S, Wang Y-Z, Qi H-W, Wang D, et al. The Role of Hydrogen Sulfide in Respiratory Diseases. Biomolecules. 2021; 11(5):682. https://doi.org/10.3390/biom11050682
Chicago/Turabian StyleKhattak, Saadullah, Qian-Qian Zhang, Muhammad Sarfraz, Pir Muhammad, Ebenezeri Erasto Ngowi, Nazeer Hussain Khan, Saqib Rauf, Yi-Zhen Wang, Hui-Wen Qi, Di Wang, and et al. 2021. "The Role of Hydrogen Sulfide in Respiratory Diseases" Biomolecules 11, no. 5: 682. https://doi.org/10.3390/biom11050682
APA StyleKhattak, S., Zhang, Q.-Q., Sarfraz, M., Muhammad, P., Ngowi, E. E., Khan, N. H., Rauf, S., Wang, Y.-Z., Qi, H.-W., Wang, D., Afzal, A., Ji, X.-Y., & Wu, D.-D. (2021). The Role of Hydrogen Sulfide in Respiratory Diseases. Biomolecules, 11(5), 682. https://doi.org/10.3390/biom11050682