
Apolipoprotein Mimetic Peptides: An Emerging Therapy against Diabetic Inflammation and Dyslipidemia

Abstract
1. Lipoprotein Abnormalities in Patients with Diabetes
2. Modification of Apolipoprotein (apo) A-I in Type 2 Diabetes Patients and Consequences
3. Apolipoprotein Mimetic Peptides in the Treatment of Diabetes
4. Effect of ApoE Mimetic Peptides on Plasma Cholesterol and Diabetes in Mouse Models of Diabetes
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luscher, T.F.; Creager, M.A.; Beckman, J.A. Diabetes and Vascular Disease. Circulation 2003, 108, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Krauss, R.M. Lipids and Lipoproteins in Type 2 Diabetes. Diabetes Care 2004, 27, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.T.; Superko, H.R.; Haskell, W.L.; Alderman, E.L.; Blanche, P.J.; Hall, L.G.; Krauss, R.M. Smallest LDL Particles are Most Strongly Related to Coronary Disease Progression in Men. Arteriosclr. Throm. Vasc. Biol. 2003, 23, 314–321. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pyrala, K.; Pederson, T.R.; Kjekshus, J.; Faergeman, O.; Olson, A.G. Cholessterol Lowering with Simvastatin Improves Prognosis of Diabetic Patients with Coronary Heart Disease: A Subgroup Ananysis of Scandinavian Simvastatin Survival Study (4S). Diabetes Care 1997, 20, 614–620. [Google Scholar] [CrossRef]
- Navab, M.; Reddy, S.T.; Van Lenten, B.J.; Fogelman, A.M. HDL and Cardiovascular Disease: Atherogenic and Atheroprotective Mechanisms. Nat. Rev. Cardiol. 2011, 8, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Couret, D.; Tran-Dinh, A.; Duranteau, J.; Montravers, P.; Schwendeman, A.; Meilhac, O. High-Density Lipoproteins during Sepsis: From Bench to Bedside. Crit. Care 2020, 24, 1–11. [Google Scholar] [CrossRef]
- Castelli, W.P.; Garrison, R.J.; Wilson, P.W.; Abbot, R.D.; Kolousdian, S.; Kannel, W.B. Incedence of Coronary Heart Disease and Lipoprotein Cholesterol levels: The Framingham Study. JAMA 1986, 256, 2835–2838. [Google Scholar] [CrossRef]
- Briel, M.; Ferreira-Gonzalez, I.; You, J.J.; Karanicolas, P.J.; Akl, E.A.; Wu, P.; Blechacz, B.; Bassler, D.; Wei, X.; Sharman, A.; et al. Association between Change in High Density Lipoprotein Cholesterol and Cardiovascular Disease Morbidity and Mortality: Systematic Review and Meta-Regression Analysis. BMJ. 2009, 338, 1–8. [Google Scholar] [CrossRef]
- Barter, P.J.; Rye, K.A. HDL Cholesterol Concentration or HDL Function: Which Matters? Eur. Heart J. 2017, 38, 2487–2489. [Google Scholar] [CrossRef]
- Drew, B.G.; Rye, K.A.; Duffy, S.J.; Barter, P.; Kingwell, B.A. The Emerging Role of HDL in Glucose Metabolism. Nat. Rev. Endocrinol. 2012, 8, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Haase, C.L.; Tybjærg-Hansen, A.; Nordestgaard, B.G.; Frikke-Schmidt, R. HDL Cholesterol and Risk of Type 2 Diabetes: A Mendelian Randomization Study. Diabetes 2015, 64, 3328–3333. [Google Scholar] [CrossRef]
- Brunham, L.R.; Kruit, J.K.; Pape, T.D.; Timmins, J.M.; Reuwer, A.Q.; Vasanji, Z.; Marsh, B.J.; Rodrigues, B.; Johnson, J.D.; Parks, J.S.; et al. Beta-Cell ABCA1 Influences Insulin Secretion, Glucose Homeostasis and Response to Thiazolidinedione Treatment. Nat. Med. 2007, 3, 340–347. [Google Scholar] [CrossRef]
- Drew, B.G.; Duffy, S.J.; Formosa, M.F.; Natoli, A.K.; Henstridge, D.C.; Penfold, S.A.; Thomas, W.G.; Mukhamedova, N.; de Courten, B.; Forbes, J.M.; et al. High-Density Lipoprotein Modulates Glucose Metabolism in Patients with Type 2 Diabetes Mellitus. Circulation 2009, 119, 2103–2111. [Google Scholar] [CrossRef]
- Rader, D.J.; Hovingh, G.K. HDL and Cardiovascular Disease. Lancet 2014, 384, 618–625. [Google Scholar] [CrossRef]
- Luscher., F.; Landmesser, U.; von Eckerdstein, A.; Fogelman, A.M. High Density Lipoprotein: Vascular Protective Effects, Dysfunction and Potential as Therapeutic Target. Circulation 2014, 114, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Ansell, B.J.; Navab, M.; Hama, S.; Kamaranpour, N.; Fonarrow, G.; Hough, G.; Rahmani, S.; Mottahedeh, R.; Dave, R.; Reddy, S.T.; et al. Inflammatory/Anti-Inflammatory Properties of High Density Lipoprotein Distinguish Patients from Control Subjects Better than HDL Cholesterol Levels and are Favorably Affected by Simvastatin Treatment. Circulation 2003, 108, 2751–2756. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.A.K. Dysfunctional HDL in Diabetes Mellitus and its Role in the Pathogenesis of Cardiovascular Disease. Mol. Cell Biochem. 2018, 440, 167–187. [Google Scholar] [CrossRef] [PubMed]
- Orchared, T.J. Dyslipidemia and Diabetes. Endocrinnol. Metabol. Clin. N. Am. 1991, 19, 19361–19380. [Google Scholar]
- Nobercourt, E.; Davis, M.J.; Brown, B.E.; Curtiss, L.K.; Bonnet, D.; Charlton, E.; Januszewski, A.S.; Jenkins, A.J.; Barter, P.J.; Rye, K.-A. The Impact of Glycation on Apolipoprotein A-I Structure and its Ability to Activate Lecithin: Cholesterol Acyltransferase. Diabetologia 2017, 50, 643–653. [Google Scholar] [CrossRef]
- Tan, K.C.B. Reverse Cholesterol Transport in Type2 Diabetes Mellitus. Diabetes Obes. Metab. 2009, 11, 534–543. [Google Scholar] [CrossRef]
- Apro, J.; Tietge, J.F.; Kikkers, A.; Parini, P.; Angelin, B.; Rudling, M. Impaired Cholesterol Efflux Capacity of High-Density Lipoprotein Isolated from Interstitial Fluid in Type 2 Diabetes Mellitus—Brief Report. Athero. Thromb. Vasc. Biol. 2016, 36, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Zannis, V.I.; Chroni, A.; Krieger, M. Role of apoA-I, ABCA1, LCAT, and SR-BI in the Biogenesis of HDL. J. Mol. Med. 2006, 84, 276–294. [Google Scholar] [CrossRef]
- Pu, L.J.; Lu, L.; Zhang, R.Y.; Du, R.; Shen, Y.; Zhang, Q.; Yang, Z.K.; Chen, Q.; Shen, W.F. Glycation of Apoprotein A-I is Associated with Coronary Artery Plaque Progression in Type 2 Diabetic Patients. Diabetes Care 2013, 36, 1312–1320. [Google Scholar]
- Han, R.; Lai, R.; Ding, Q.; Wang, Z.; Luo, X.; Zhang, Y.; Cui, G.; He, J.; Liu, W.; Chen, Y. Apolipoprotein A-I Stimulates AMP-Activated Protein Kinase and Improves Glucose Metabolism. Diabetologia 2007, 50, 1960–1968. [Google Scholar] [CrossRef]
- Liu, D.; Ji, L.; Zhao, M.; Wang, Y.; Guo, Y.; Li, L.; Zhang, D.; Xu, L.; Pan, B.; Su, J.; et al. Lysine Glycation of Apolipoprotein A-I Impairs Its Anti-Inflammatory Properties in Type 2 Diabetes Mellitus. J. Mol. Cell Biol. 2018, 122, 47–57. [Google Scholar]
- Han, A.; Murphy, A.J.; Couglhan, M.T.; Thomas, M.C.; Forbes, J.M.; O’Brian, R.; Cooper, M.E.; Chin-Dusting, J.P.F.; Sviridov, V. Advanced glycation of alolipoprotein A-I impairs its anti-atherogenic properties. Diabitologia 2007, 50, 1770–1779. [Google Scholar]
- Bashtovyy, D.; Jones, M.K.; Anantharamaiah, G.M.; Segrest, J.P. Sequence Conservation of Apolipoprotein A-I Affords Novel Insights into HDL Structure-Function. J. Lipid. Res. 2011, 52, 435–450. [Google Scholar] [CrossRef]
- Palgunachari, M.N.; Mishra, V.K.; Lund-Katz, S.; Phillips, M.C.; Adeyeye, S.O.; Alluri, S.; Anantharamaiah, G.M.; Segrest, J.P. Only the Two End Helixes of Eight Tandem Amphipathic Helical Domains of Human Apo A-I have Significant Lipid Affinity: Implications for HDL Assembly. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 328–338. [Google Scholar] [CrossRef]
- He, Y.; Song, H.D.; Anantharamaiah, G.M.; Palgunachari, M.N.; Bornfeldt, K.E.; Segrest, J.P.; Heinecke, J.W. Apolipoprotein A1 Forms 5/5 and 5/4 Antiparallel Dimers in Human High-density Lipoprotein. Mol. Cell Proteomics 2019, 18, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Kothari, V.; Bornfeldt, K.E. A New Treatment Strategy for Diabetic Dyslipidemia? Diabetes 2020, 69, 2061–2063. [Google Scholar] [CrossRef] [PubMed]
- Anantharamaiah, G.M.; Jones, J.L.; Brouillette, C.G.; Schmidt, C.F.; Chung, B.H.; Hughes, T.A.; Bhown, A.S.; Segrest, J.P. Studies of Synthetic Peptide Analogs of the Amphipathic Helix. Structure of Complexes with Dimyristoyl Phosphatidylcholine. J. Biol. Chem. 1985, 260, 10248–10255. [Google Scholar] [CrossRef]
- Segrest, J.P.; Garber, D.W.; Brouillette, C.G.; Harvey, S.S.; Anantharamaiah, G.M. The Amphipathic Helix in Exchangeable Apolipoproteins: A Multifunctional and Structural Motif in Plasma Apolipoproteins. Adv. Protein. Chem. 1994, 45, 303–369. [Google Scholar]
- Garber, D.W.; Datta, G.; Chaddha, M.; Palgunachari, M.N.; Hama, S.Y.; Navab, M.; Fogelman, A.M.; Segrest, J.P.; Anantharamaiah, G.M. A New Synthetic Class A Amphipathic Peptide Analogue Protects Mice from Diet-Induced Atherosclerosis. J. Lipid. Res. 2001, 42, 545–552. [Google Scholar] [CrossRef]
- Peterson, S.J.; Drummond, G.; Kim, D.H.; Li, M.; Kruger, A.L.; Ikehara, S.; Abraham, N.G. L-4F Treatment Reduces Adiposity, Increases Adiponectin Levels, and Improves Insulin Sensitivity in Obese Mice. J. Lipid. Res. 2008, 49, 1658–1669. [Google Scholar] [CrossRef]
- Peterson, S.J.; Kim, D.H.; Li, M.; Positano, V.; Vanella, L.; Rodella, L.F.; Piccolomini, F.; Puri, N.; Gastaldelli, A.; Kusmic, C.; et al. The L-4F Mimetic Peptide Prevents Insulin Resistance through Increased Levels of HO-1, pAMPK, and pAKT in Obese Mice. J. Lipid. Res. 2009, 50, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Kruger, A.L.; Peterson, S.; Turkseven, S.; Kaminski, P.M.; Zhang, F.F.; Quan, S.; Wolin, M.S.; Abraham, N.G. D-4F Induces Heme Oxygenase-1 and Extracellular Superoxide Dismutase, Decreases Endothelial Cell Sloughing, and Improves Vascular Reactivity in Rat Model of Diabetes. Circulation 2005, 111, 3126–3134. [Google Scholar] [CrossRef]
- Benson, M.; Peterson, S.J.; Mehta, P.; Abraham, N.G. ApoA-I Mimetic Peptides and Diabetes. In Apolipoprotein Mimetics in the Management of Human Disease; Anantharamaiah, G.M., Goldberg, D., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 43–54. [Google Scholar]
- Baotic, I.; Ge, Z.D.; Sedlic, F.; Coon, A.; Weihrauch, D.; Warltier, D.C.; Kersten, J.R. Apolipoprotein A-1 Mimetic D-4F Enhances Isoflurane-Induced eNOS Signaling and Cardioprotection during Acute Hyperglycemia. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, 219–227. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Morgantini, C.; Imaizumi, S.; Grijalva, V.; Navab, M.; Fogelman, A.M.; Reddy, S.T. Apolipoprotein A-I Mimetic Peptides Prevent Atherosclerosis Development and Reduce Plaque Inflammation in a Murine Model of Diabetes. Diabetes 2010, 59, 3223–3228. [Google Scholar] [CrossRef]
- Cao, J.; Puri, N.; Sodhi, K.; Bellner, L.; Abraham, N.G.; Kappas, A. Apo A1 Mimetic Rescues the Diabetic Phenotype of HO-2 Knockout Mice via an Increase in HO-1 Adiponectin and LKBI Signaling Pathway. Int. J. Hypertens. 2012, 2012, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Mahley, R.W.; Ji, Z.-S. Remnant Lipoprotein Metabolism: Key Pathways Involving Cell-Surface Heparin Sulfate Proteoglycans and Apolipoprotein E. J. Lipid. Res. 1999, 40, 1–16. [Google Scholar] [CrossRef]
- Gonzales, J.C.; Gordts, P.I.; Foley, E.M.; Esko, J.D. Apolipoprotein E and AV Mediate Lipoprotein Clearance by Hepatic Proteoglycans. J. Clin. Invest. 2013, 135, 2742–2751. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J. Apolipoprotein E and Atherosclerosis:Beyond Lipid Effect. Arterio. Thromb. Vasc. Biol. 2005, 25, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Weisgraber, K.H. Apolipoprotein E: Structure-Function Relationship. Adv. Protein. Chem. 1994, 45, 249–302. [Google Scholar] [PubMed]
- Datta, G.; Chaddha, M.; Garber, D.W.; Chung, B.H.; Tyler, E.M.; Dashti, N.; Bradley, W.A.; Gianturco, S.H.; Anantharamaiah, G.M. The Receptor Binding Domain of apoE, Linked to a Model Class A Amphipathic Helix., Enhances Internalization and Degradation of LDL by Fibroblasts. Biochemistry 2000, 39, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Handattu, S.P.; Nayyar, G.; Garber, D.W.; Palgunachari, M.N.; Monroe, C.E.; Keenam, T.D.; Mishra, M.K.; Datta, G.; Anantharamaiah, G.M. Two Apolipoprotein E Mimetic Peptides with Similar Cholesterol Reducing Properties Exhibit Different Atheroprotective Effects in apoE Null Mice. Atherosclerosis 2013, 227, 58–68. [Google Scholar] [CrossRef]
- Gupta, H.; White, C.R.; Handattu, S.P.; Garber, D.W.; Datta, G.; Chaddha, M.; Dai, L.; Gianturco, S.H.; Bradley, W.A.; Anantharamaiah, G.M. Apolipoprotein E Mimetic Peptide Dramatically Lowers Plasma Cholesterol and Restores Endothelial Function in Watanabe Heritable Hyperlipidemic Rabbits. Circulation 2005, 111, 3112–3118. [Google Scholar] [CrossRef][Green Version]
- Nayyar, G.; Garber, D.W.; Palgunachari, M.N.; Moore, C.E.; Keenam, C.E.; Handattu, S.P.; Mishra, V.K.; Anantharamaiah, G.M. Apolipoprotein E Mimetic Peptide is More Effective than Apolipoprotein A-I Mimetic in Reducing Lesion Formation in Older apoE Null Mice. Atherosclerosis 2012, 224, 326–331. [Google Scholar] [CrossRef]
- White, C.R.; Garber, D.W.; Anantharamaiah, G.M. Anti-Inflammatory and Cholesterol Reducing Properties of Apolipoprotein Mimetics: A Review. J. Lipid. Res. 2014, 55, 2007–2021. [Google Scholar] [CrossRef]
- White, R.C.; Anantharamaiah, G.M. Unpublished Results.
- Anantharamaiah, G.M.; Garber, D.W.; Goldberg, D.; Morrel, E.; Datta, G.; Palgunachari, M.N.; Register, T.C.; Appt, S.E.; White, C.R. Novel Fatty Acyl apoE Mimetic Peptides have Increased Potency to Reduce Plasma Cholesterol in Mice and Macaques. J. Lipid. Res. 2018, 59, 2075–2083. [Google Scholar] [CrossRef]
- Giordano-Mooga, S.; Datta, G.; Wolkowicz, P.; Garber, D.W.; Palgunachari, M.N.; White, C.R.; Anantharamaiah, G.M. The Apolipoprotein E Mimetic Peptide AEM-2 Attenuates Mitochondrial Injury and Apoptosis in Human THP-1 Macrophages. Curr. Top. Pept. Protein. Res. 2018, 19, 15–25. [Google Scholar]



{kind=link}
{kind=link}
| Peptide | Animal Model | Biological Changes | References |
|---|---|---|---|
| 4F | Obese mice | Adiposity ↓ | [34] |
| Bone marrow adipogenesis ↓ | [34] | ||
| Insulin sensitivity ↑ | [34,35] | ||
| Glucose tolerance ↑ | [35,37] | ||
| IL-b, IL-6, superoxide ↓ | [37] | ||
| Hepatic lipid content ↓ | [35,37] | ||
| Adipocytes of small size ↑ | [35,37] | ||
| D-4F | Sprague–Dawley Rats with STZ | HO-1 ↑ SOD ↑ | [36] |
| Endothelial sloughing ↓ | [36] | ||
| 4F | HO-2 knockout mice | HO-1 ↑ Adeponectin ↑ | [40] |
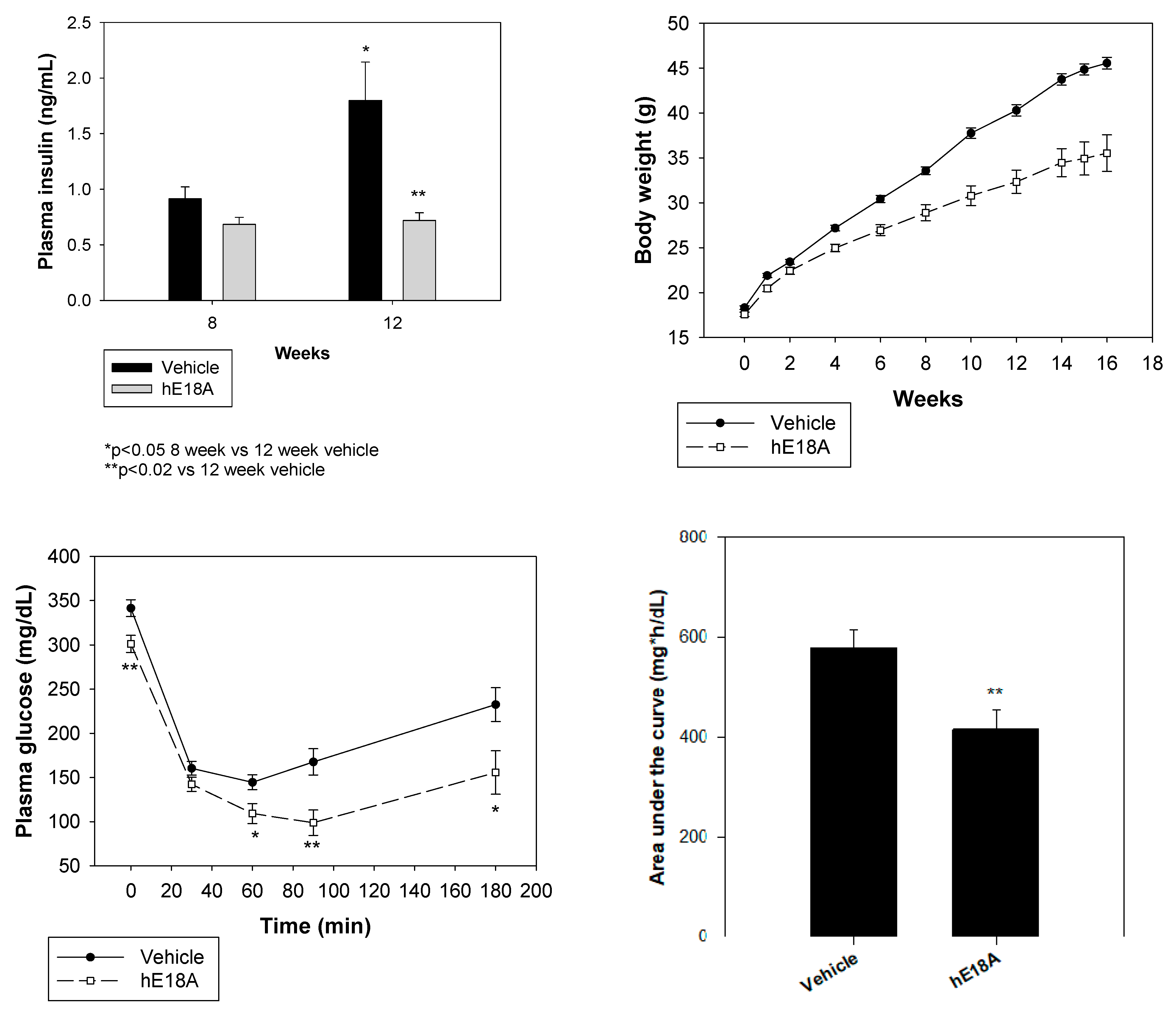
| Ac-hE18A-NH2 | C57B6/fat administered | Plasma insulin ↑ | [50] |
| Insulin sensitivity ↑ | |||
| Plasma glucose ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolkowicz, P.; White, C.R.; Anantharamaiah, G.M. Apolipoprotein Mimetic Peptides: An Emerging Therapy against Diabetic Inflammation and Dyslipidemia. Biomolecules 2021, 11, 627. https://doi.org/10.3390/biom11050627
Wolkowicz P, White CR, Anantharamaiah GM. Apolipoprotein Mimetic Peptides: An Emerging Therapy against Diabetic Inflammation and Dyslipidemia. Biomolecules. 2021; 11(5):627. https://doi.org/10.3390/biom11050627
Chicago/Turabian StyleWolkowicz, Paul, C. Roger White, and G. M. Anantharamaiah. 2021. "Apolipoprotein Mimetic Peptides: An Emerging Therapy against Diabetic Inflammation and Dyslipidemia" Biomolecules 11, no. 5: 627. https://doi.org/10.3390/biom11050627
APA StyleWolkowicz, P., White, C. R., & Anantharamaiah, G. M. (2021). Apolipoprotein Mimetic Peptides: An Emerging Therapy against Diabetic Inflammation and Dyslipidemia. Biomolecules, 11(5), 627. https://doi.org/10.3390/biom11050627