
Nourin-Dependent miR-137 and miR-106b: Novel Early Inflammatory Diagnostic Biomarkers for Unstable Angina Patients



 , , , ,
, , , ,
Abstract
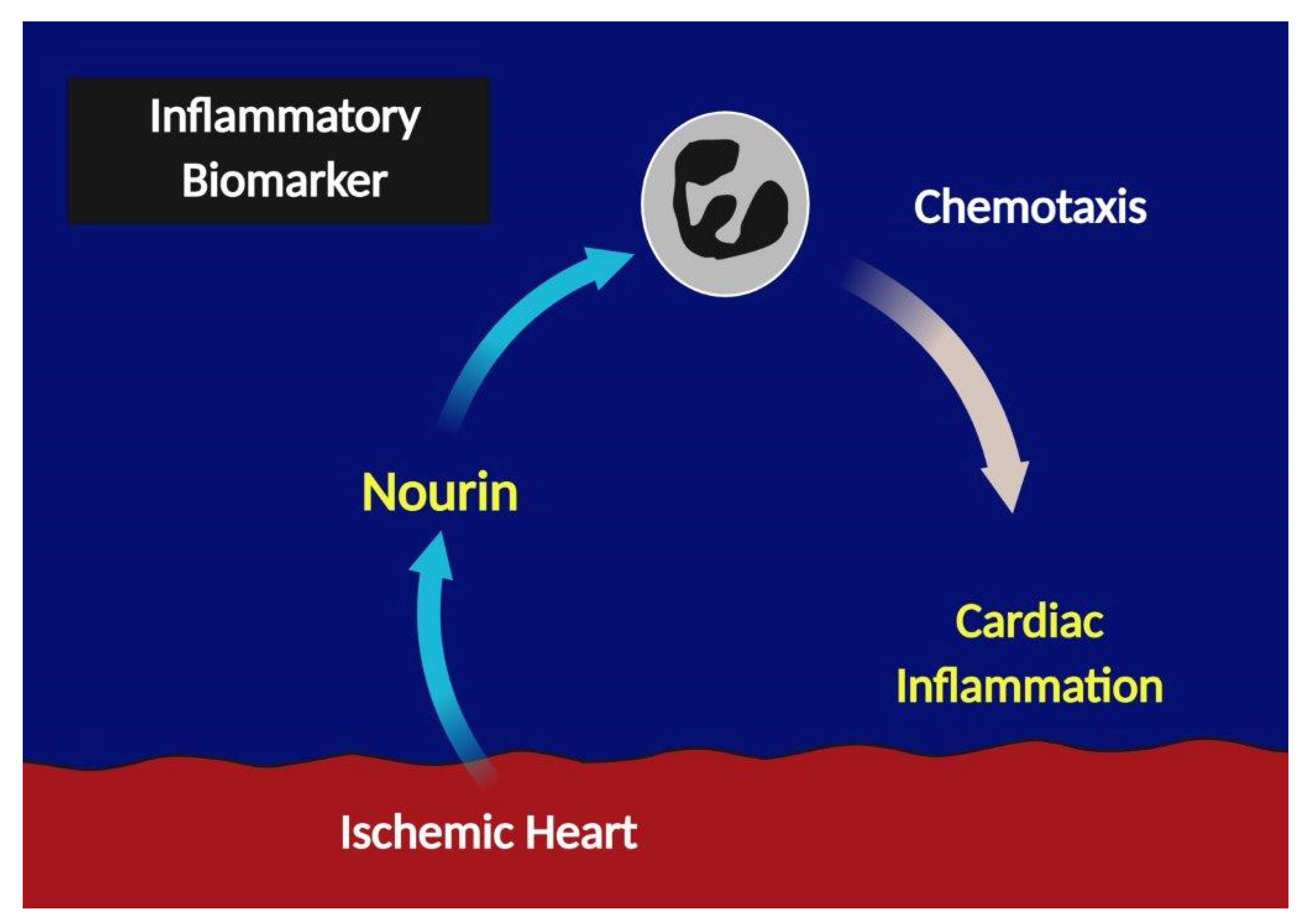
1. Introduction
2. Subjects and Methods
2.1. Bioinformatics Analysis
2.2. Study Design/Participant’s Selection
2.3. Total and miRNA Extraction and Purification
2.4. Gene Amplification Analysis by Quantitative Real Time PCR (qPCR)
2.5. Statistical Analysis
3. Results
3.1. Demographic Characteristics of Participants
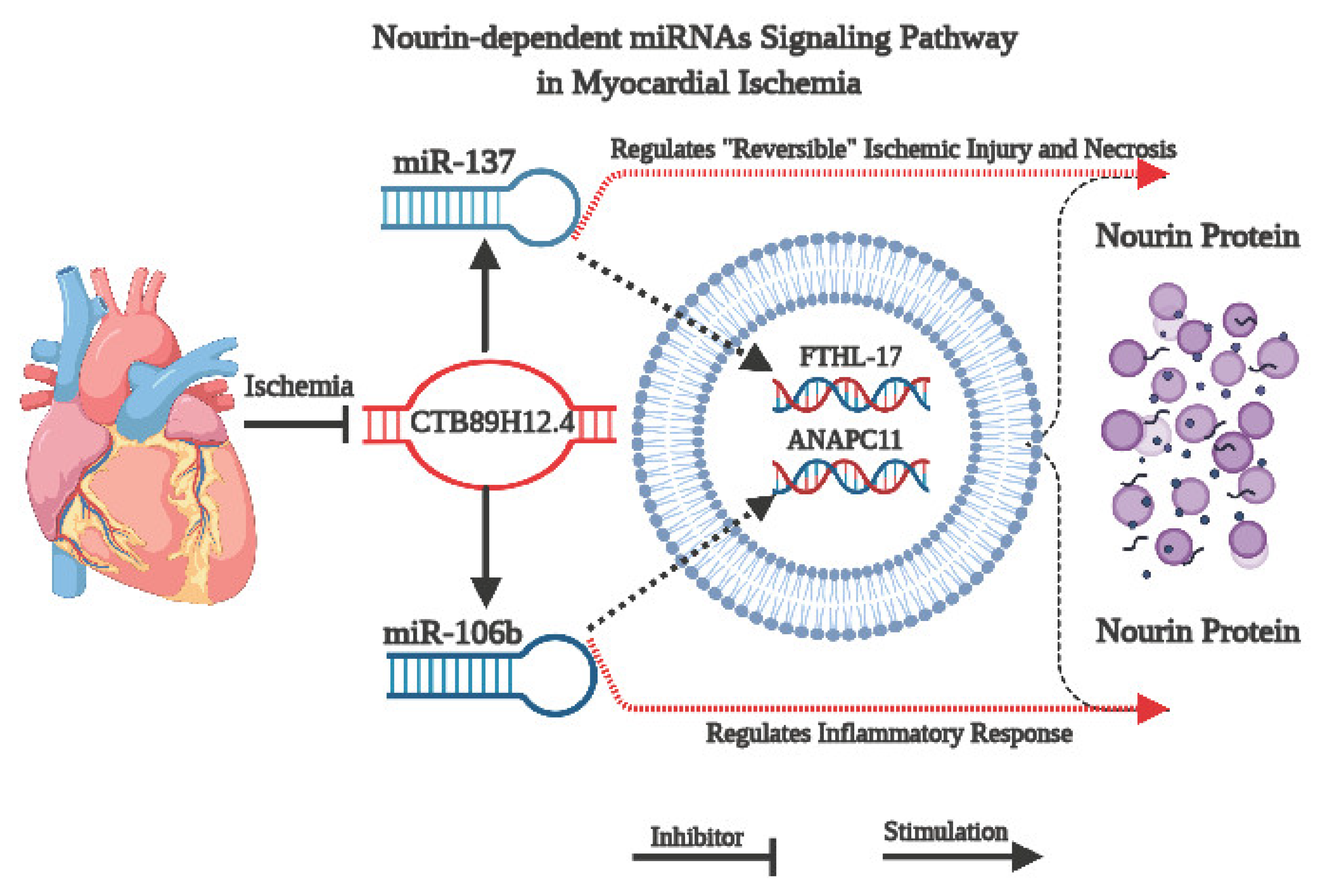
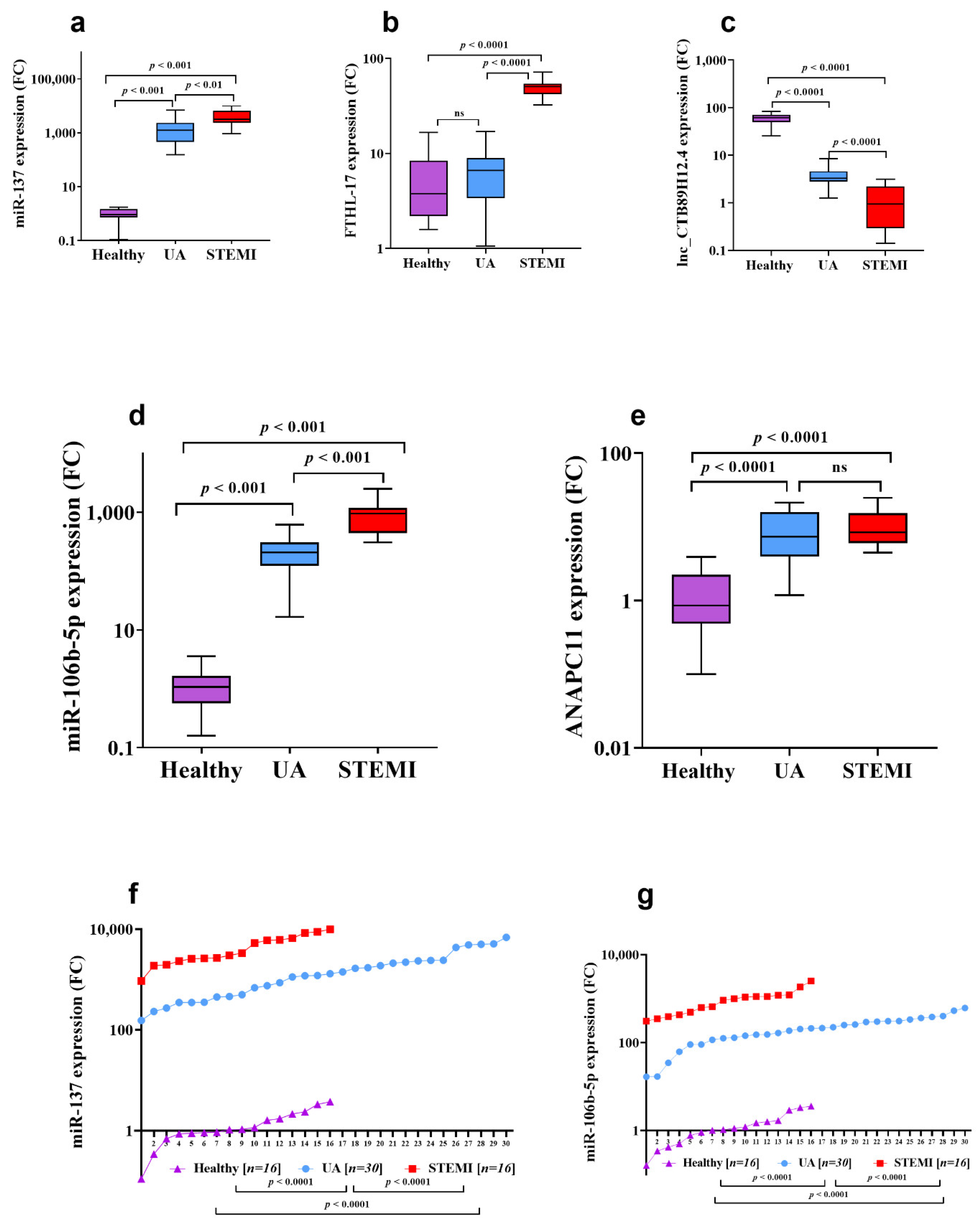
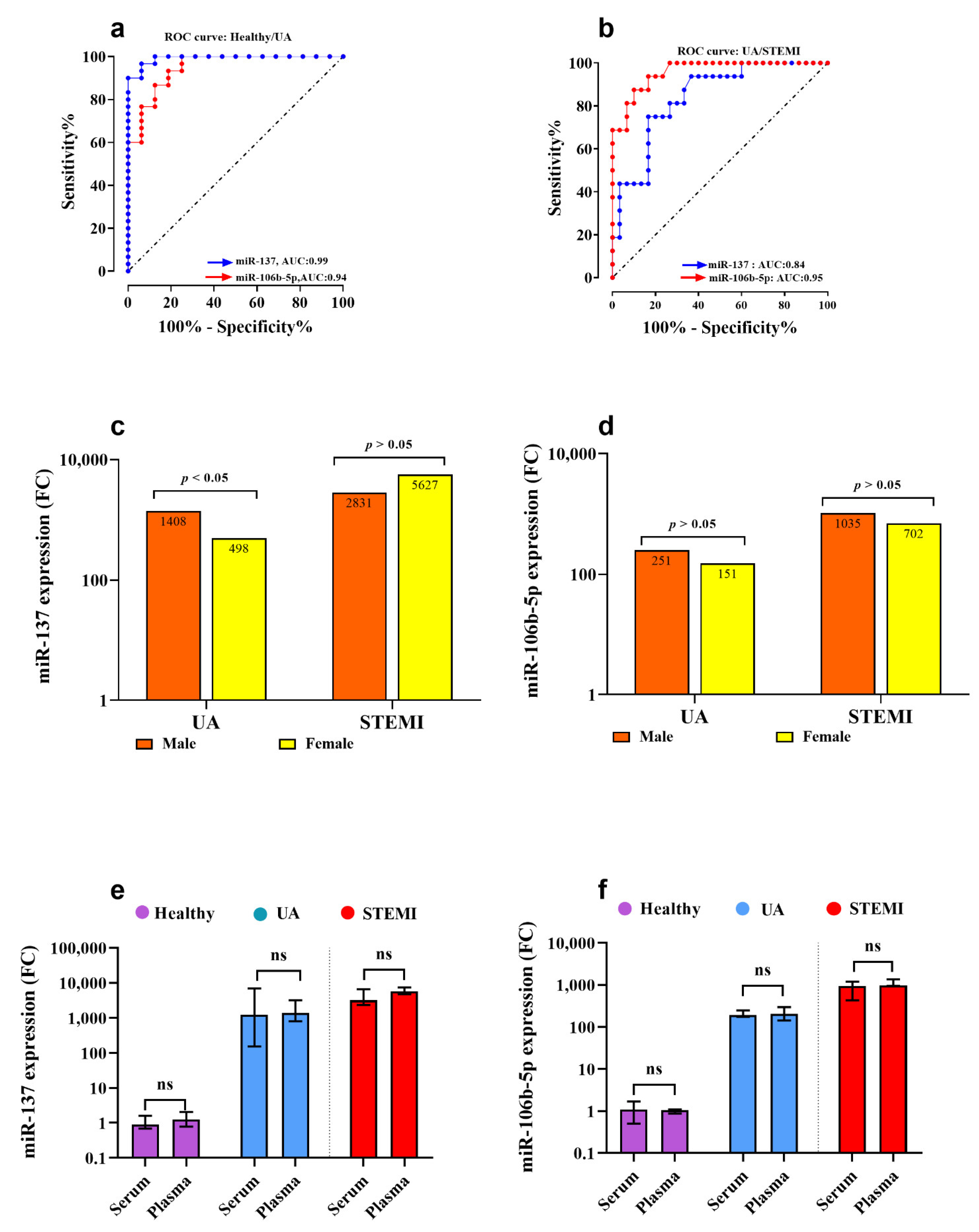
3.2. Assessment of CTB89H12.4/miR-137/FTHL-17 Network
3.3. Assessment of CTB89H12.4/miR-106b-5p/ANAPC11 Network
3.4. Correlation between lncRNA/miRNA/mRNA Genes in Acute Coronary Syndrome (ACS) Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| UA | unstable angina |
| STEMI | ST-elevation myocardial infarction |
| ACS | acute coronary syndrome |
| CAD | coronary artery disease |
| ED | emergency department |
| FTHL-17 | ferritin heavy chain like 17 |
| ANAPC11 | anaphase promoting complex subunit |
| CTB89H12.4 | lncRNA-CTB89H12.4 |
| PI3K | phosphatidylinositol-3 kinase |
| Akt | protein kinase B (PKB) |
| mTOR | mammalian target of rapamycin |
| Jak-STAT | janus kinase/signal transducer and activator of transcription |
| NGS | next-generation sequencing |
| TNF-α | tumor necrosis factor-α |
| IL-1β | interleukin 1β |
| IL-8 | interleukin 8 |
| LECAM-1 | leukocyte-endothelial cell adhesion molecule 1 |
| ICAM-1 | intercellular adhesion molecule 1 |
| ELAM-1 | endothelial-leukocyte adhesion molecule 1 |
| FPR | formyl peptide receptor |
| CRP | c-reactive protein |
| TLR | toll-like receptor |
| HIF-1α | hypoxia-inducible factor-1α |
| ROC | receiver operator characteristics |
| AUC | area under the curve |
| VECs | vascular endothelial cells |
| DEGs | different expressed genes |
| GO | gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PPI | protein–protein Interaction |
| ATP | adenosine triphosphate |
References
- Kaski, J.C.; Crea, F.; Gersh, B.J.; Camic, P.G.i. Reappraisal of Ischemic Heart Disease: Fundamental Role of Coronary Microvascular Dysfunction in the Pathogenesis of Angina Pectoris. Circulation 2018, 138, 1463–1480. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.-P.; Thiele, H.; Barbato, E.; Barthélémy, O.; Bauersachs, J.; Bhatt, D.L.; Dendale, P.; Dorobantu, M.; Edvardsen, T.; Folliguet, T.; et al. 2020 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevationThe Task Force for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2020, 1–79. [Google Scholar] [CrossRef]
- Jakimov, T.; Mrdović, I.; Filipović, B.; Zdravković, M.; Djoković, A.; Hinić, S.; Milić, N.; Filipović, B. Comparison of RISK-PCI, GRACE, TIMI risk scores for prediction of major adverse cardiac events in patients with acute coronary syndrome. Croat. Med. J. 2017, 58, 406–415. [Google Scholar] [CrossRef]
- Yan, A.T.; Yan, R.T.; Tan, M.; Casanova, A.; Labinaz, M.; Sridhar, K.; Fitchett, D.H.; Langer, A.; Goodman, S.G. Risk scores for risk stratification in acute coronary syndromes: Useful but simpler is not necessarily better. Eur. Heart J. 2007, 28, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Bing, R.; Dweck, M.R.; van Beek, E.J.R.; Mills, N.L.; Williams, M.C.; Villines, T.C.; Newby, D.E.; Adamson, P.D. Exercise Electrocardiography and Computed Tomography Coronary Angiography for Patients With Suspected Stable Angina Pectoris: A Post Hoc Analysis of the Randomized SCOT-HEART Trial. JAMA Cardiol. 2020, 5, 920–928. [Google Scholar] [CrossRef] [PubMed]
- BreweR, L.C.; Svatikova, A.; Mulvagh, S.L. The Challenges of Prevention, Diagnosis and Treatment of Ischemic Heart Disease in Women. Cardiovasc. Drugs Ther. 2015, 29, 355–368. [Google Scholar] [CrossRef]
- Parsonage, W.A.; Cullen, L.; Younger, J.F. The approach to patients with possible cardiac chest pain. Med. J. Aust. 2013, 199, 30–34. [Google Scholar] [CrossRef][Green Version]
- Aydin, S.; Ugur, K.; Aydin, S.; Sahin, I.; Yardim, M. Biomarkers in acute myocardial infarction: Current perspectives. Vasc. Health Risk Manag. 2019, 15, 1–10. [Google Scholar] [CrossRef]
- Cavarretta, E.; Frati, G. MicroRNAs in Coronary Heart Disease: Ready to Enter the Clinical Arena? Biomed Res. Int. 2016, 2016, 2150763. [Google Scholar] [CrossRef]
- Elgebaly, S.A.; Poston, R.; Todd, R.; Helmy, T.; Almaghraby, A.M.; Elbayoumi, T.; Kreutzer, D.L. Cyclocreatine protects against ischemic injury and enhances cardiac recovery during early reperfusion. Expert Rev. Cardiovasc. Ther. 2019, 17, 683–697. [Google Scholar] [CrossRef]
- Elgebaly, S.A.; Hashmi, F.H.; Houser, S.L.; Allam, M.E.; Doyle, K. Cardiac-derived neutrophil chemotactic factors: Detection in coronary sinus effluents of patients undergoing myocardial revascularization. J. Thorac. Cardiovasc. Surg. 1992, 103, 952–959. [Google Scholar] [CrossRef]
- Elgebaly, S.A.; Masetti, P.; Allam, M.; Forouhar, F. Cardiac derived neutrophil chemotactic factors; preliminary biochemical characterization. J. Mol. Cell. Cardiol. 1989, 21, 585–593. [Google Scholar] [CrossRef]
- Elgebaly, S.A.; van Buren, C.; Todd, R.; Poston, R.; Rabie, M.A.; Mohamed, A.F.; Ahmed, L.A.; El Sayed, N.S. Cyclocreatine Phosphate: A Novel Mechanism for Preventing Development of Heart Failure. Circulation 2020, 142 (Suppl. 3), A13311. [Google Scholar]
- Elgebaly, S.A.; Christenson, R.H.; Kandil, H.; El-Khazragy, N.; Rashed, L.; Yacoub, B.; Sharafieh, R.; Klue, U.; Kreutzer, D.L. Nourin-dependent Mirna-137: A Novel Early Diagnostic Biomarker for Unstable Angina Patients. Circulation 2020, 142 (Suppl. 3), A13051. [Google Scholar] [CrossRef]
- Elgebaly, S.A.; Christenson, R.H.; Kandil, H.; Elkhazragy, N.; Rashed, L.; Yacou, B.; Sharafieh, R.; Klueh, U.; Kreutzer, D.L. Nourin-dependent Mirna-106b: A Novel Early Inflammatory Diagnostic Biomarker for Cardiac Injury. Circulation 2020, 142 (Suppl. 3), A13103. [Google Scholar] [CrossRef]
- Ozeren, A.; Aydin, M.; Tokac, M.; Demircan, N.; Unalacak, M.; Gurel, A.; Yazici, M. Levels of serum IL-1beta, IL-2, IL-8 and tumor necrosis factor-alpha in patients with unstable angina pectoris. Mediat. Inflamm. 2003, 12, 361–365. [Google Scholar] [CrossRef]
- Elgebaly, S.; Christenson, R.; Schiffmann, E.; Yi, Q.; Kreutzer, D. Early identification of cardiac ischemia patients in the emergency department. Catheter. Cardiovasc. Interv. 2013, 81 (Suppl. 1), S2–S3. [Google Scholar]
- Cordeddu, L.; Pilbrow, A.P.; Cameron, V.A.; Troughton, R.W.; Richards, M.A.; Foo, R.S. Circulating miRNAs as Biomarkers in Acute Coronary Syndrome. Circ. Res. 2012, 111, A290. [Google Scholar] [CrossRef]
- Elbaz, M.; Faccini, J.; Laperche, C.; Grousset, E.; Roncalli, J.; Ruidavets, J.B.; Vindis, C. Identification of a miRNA Based-Signature Associatetd with Acute Coronary Syndrome: Evidence from the FLORINF Study. J. Clin. Med. 2020, 9, 1674. [Google Scholar] [CrossRef]
- Gacoń, J.; Kabłak-Ziembicka, A.; Stępień, E.; Enguita, F.J.; Karch, I.; Derlaga, B.; Żmudka, K.; Przewłocki, T. Decision-making microRNAs (miR-124,-133a/b,-34a and-134) in patients with occluded target vessel in acute coronary syndrome. Kardiol. Pol. 2016, 74, 280–288. [Google Scholar]
- Wang, A.; Kwee, L.C.; Grass, E.; Neely, M.L.; Gregory, S.G.; Fox, K.A.A.; Armstrong, P.W.; White, H.D.; Ohman, E.M.; Roe, M.T.; et al. Whole blood sequencing reveals circulating microRNA associations with high-risk traits in non-ST-segment elevation acute coronary syndrome. Atherosclerosis 2017, 261, 19–25. [Google Scholar] [CrossRef]
- Zhang, M.; Ge, D.J.; Su, Z.; Qi, B. miR-137 alleviates focal cerebral ischemic injury in rats by regulating JAK1/STAT1 signaling pathway. Hum. Exp. Toxicol. 2020, 39, 816–827. [Google Scholar] [CrossRef]
- Suzuki, T.; Toba, K.; Kato, K.; Ozawa, T.; Higasimura, M.; Kitajima, T.; Oda, H.; Tsuchida, K.; Tomosugi, N.; Saitoh, H.; et al. Serum ferritin levels adversely affect cardiac function in patients with ST-elevation myocardial infarction who underwent successful percutaneous coronary intervention. Int. J. Cardiol. 2013, 167, 286–288. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Z.; Liu, J.; Wang, J.; Qu, C. MicroRNA-137 regulates hypoxia-induced retinal ganglion cell apoptosis through Notch1. Int. J. Mol. Med. 2018, 41, 1774–1782. [Google Scholar] [CrossRef]
- Gorabi, A.M.; Kiaie, N.; Sathyapalan, T.; Al-Rasadi, K.; Jamialahmadi, T.; Sahebkar, A. The Role of MicroRNAs in Regulating Cytokines and Growth Factors in Coronary Artery Disease: The Ins and Outs. J. Immunol. Res. 2020, 2020, 5193036. [Google Scholar] [CrossRef]
- Bostjancic EGlavac, D. miRNome in myocardial infarction: Future directions and perspective. World J. Cardiol. 2014, 6, 939–958. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, D.; Xie, P.; Ren, G.; Sun, G.; Zeng, X.; Sun, X. MiR-106b and MiR-15b modulate apoptosis and angiogenesis in myocardial infarction. Cell. Physiol. Biochem. 2012, 29, 851–862. [Google Scholar] [CrossRef]
- Li, J.; Fan, G. Identification of key genes and pathways in obstructive coronary artery disease by integrated analysis. J. Xiangya Med. 2020, 5, 14. [Google Scholar] [CrossRef]
- Park, K.C.; Gaze, D.C.; Collinson, P.O.; Marber, M.S. Cardiac troponins: From myocardial infarction to chronic disease. Cardiovasc. Res. 2017, 113, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.T.; Sörensen, N.A.; Ojeda, F.; Renné, T.; Schnabel, R.B.; Zeller, T.; Karakas, M.; Blankenberg, S.; Westermann, D. Early diagnosis of acute myocardial infarction using high-sensitivity troponin I. PLoS ONE 2017, 12, e0174288. [Google Scholar] [CrossRef]
- Pleister, A.; Selemon, H.; Elton, S.M.; Elton, T.S. Circulating miRNAs: Novel biomarkers of acute coronary syndrome? Biomark. Med. 2013, 7, 287–305. [Google Scholar] [CrossRef]
- Barraclough, J.Y.; Joglekar, M.V.; Januszewski, A.S.; Martínez, G.; Celermajer, D.S.; Keech, A.C.; Hardikar, A.A.; Patel, S. A MicroRNA Signature in Acute Coronary Syndrome Patients and Modulation by Colchicine. J. Cardiovasc. Pharmacol. Ther. 2020, 25, 444–455. [Google Scholar] [CrossRef]
- Oerlemans, M.I.F.J.; Mosterd, A.; Dekker, M.S.; de Very, E.A.; van Mil, A.; Pasterkamp, G.; Doevendans, P.A.; Hoes, A.W.; Sluijter, J.P.G. Early assessment of acute coronary syndromes in the emergency department: The potential diagnostic value of circulating microRNAs. EMBO Mol. Med. 2012, 4, 1176–1185. [Google Scholar] [CrossRef]
- Zhao, T.; Qiu, Z.; Gao, Y. MiR-137-3p exacerbates the ischemia-reperfusion injured cardiomyocyte apoptosis by targeting KLF15. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1013–1024. [Google Scholar] [CrossRef]
- Wang, J.; Xu, R.; Wu, J.; Li, Z. MicroRNA-137 Negatively Regulates H2O2-Induced Cardiomyocyte Apoptosis Through CDC42. Med. Sci. Monit. 2015, 21, 3498–3504. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, J.; Li, J.; Wei, T.; Li, J. Down-Regulation of MicroRNA-137 Improves High Glucose-Induced Oxidative Stress Injury in Human Umbilical Vein Endothelial Cells by Up-Regulation of AMPKalpha1. Cell. Physiol. Biochem. 2016, 39, 847–859. [Google Scholar] [CrossRef]
- Yang, Y.; Li, F.; Saha, M.N.; Abdi, J.; Qiu, L.; Chang, H. miR-137 and miR-197 Induce Apoptosis and Suppress Tumorigenicity by Targeting MCL-1 in Multiple Myeloma. Clin. Cancer Res. 2015, 21, 2399–2411. [Google Scholar] [CrossRef]
- Zhang, B.; Ma, L.; Wei, J.; Hu, J.; Zhao, Z.; Wang, Y.; Chen, Y.; Zhao, F. miR-137 Suppresses the Phosphorylation of AKT and Improves the Dexamethasone Sensitivity in Multiple Myeloma Cells Via Targeting MITF. Curr. Cancer Drug Targets 2016, 16, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Hao, N.-B.; He, Y.-F.; Li, X.-Q.; Wang, K.; Wang, R.-L. The role of miRNA and lncRNA in gastric cancer. Oncotarget 2017, 8, 81572–81582. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, R.; Zhou, X.; Guo, R.; Yin, J.; Li, Y.; Ma, G. miR-137: A Novel Therapeutic Target for Human Glioma. Mol. Ther. Nucleic Acids 2020, 21, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Song, B. Notch 1 signalling inhibits cardiomyocyte apoptosis in ischaemic postconditioning. Heart Lung Circ. 2014, 23, 152–158. [Google Scholar] [CrossRef]
- Guarnieri, A.L.; Towers, C.G.; Drasin, D.J.; Oliphant, M.U.J.; Andrysik, Z.; Hotz, T.J.; Vartuli, R.L.; Linklater, E.S.; Pandey, A.; Khana, S.L.; et al. The miR-106b-25 cluster mediates breast tumor initiation through activation of NOTCH1 via direct repression of NEDD4L. Oncogene 2018, 37, 3879–3893. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.; Lang, F.; Liu, Y.-X.; Yang, C.-Q.; Gao, H.-J. In situ hybridization analysis of the expression of miR-106b in colonic cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 786–792. [Google Scholar] [PubMed]
- Zhang, J.; Li, S.F.; Chen, H.; Song, J.X. Role of miR-106b-5p in the regulation of gene profiles in endothelial cells. Beijing Da Xue Xue Bao Yi Xue Ban 2019, 51, 221–227. [Google Scholar] [PubMed]
- Kim, J.; Yoon, H.; Ramírez, C.M.; Lee, S.M.; Hoe, H.S.; Fernández-Hernando, C.; Kim, J. MiR-106b impairs cholesterol efflux and increases Aβ levels by repressing ABCA1 expression. Exp. Neurol. 2012, 235, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.H.; Mitchell, D.A.; McGowan, S.D.; Evanoff, R.; Griswold, M.D. Two miRNA clusters, Mir-17-92 (Mirc1) and Mir-106b-25 (Mirc3), are involved in the regulation of spermatogonial differentiation in mice. Biol. Reprod. 2012, 86, 72. [Google Scholar] [CrossRef]
- Novák, J.; Olejníčková, V.; Tkáčová, N.; Santulli, G. Mechanistic Role of MicroRNAs in Coupling Lipid Metabolism and Atherosclerosis. Adv. Exp. Med. Biol. 2015, 887, 79–100. [Google Scholar]
- Pan, J.; Li, K.; Huang, W.; Zhang, X. MiR-137 inhibited cell proliferation and migration of vascular smooth muscle cells via targeting IGFBP-5 and modulating the mTOR/STAT3 signaling. PLoS ONE 2017, 12, e0186245. [Google Scholar] [CrossRef]
- Maimaiti, A.; Maimaiti, A.; Yang, Y.; Ma, Y. MiR-106b exhibits an anti-angiogenic function by inhibiting STAT3 expression in endothelial cells. Lipids Health Dis. 2016, 15, 51. [Google Scholar] [CrossRef]
- Eid, R.; Boucher, E.; Gharib, N.; Khoury, C.; Arab, N.T.T.; Murray, A.; Young, P.G.; Mandato, C.A.; Greenwood, M.T. Identification of human ferritin, heavy polypeptide 1 (FTH1) and yeast RGI1 (YER067W) as pro-survival sequences that counteract the effects of Bax and copper in Saccharomyces cerevisiae. Exp. Cell Res. 2016, 342, 52–61. [Google Scholar] [CrossRef]
- Zhong, S.; Xu, Y.; Yu, C.; Zhang, X.; Li, L.; Ge, H.; Ren, G.; Wang, Y.; Ma, J.; Zheng, Y.; et al. Anaphase-promoting complex/cyclosome regulates RdDM activity by degrading DMS3 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 3899–3908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhuang, J.; Liu, L.; He, Z.; Liu, C.; Ma, X.; Li, J.; Ding, X.; Sun, C. Integrative transcriptome data mining for identification of core lncRNAs in breast cancer. PeerJ 2019, 7, e7821. [Google Scholar] [CrossRef] [PubMed]
- Saddic, L.A.; Sigurdsson, M.I.; Chang, T.-W.; Mazaika, E.; Heydarpour, M.; Shernan, S.K.; Seidman, C.E.; Seidman, J.G.; Aranki, S.F.; Body, S.C.; et al. The Long Noncoding RNA Landscape of the Ischemic Human Left Ventricle. Circ. Cardiovasc. Genet. 2017, 10, e001534. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | ACS (n = 46) | Healthy (n = 16) | p-Value | |
|---|---|---|---|---|
| UA (n = 30) | STEMI (n = 16) | |||
| Age (years) mean ± SD | 60.1 ± 8.1 | 54.4 ± 12.7 | 32.9 ± 9.9 | 0.001 |
| Sex: Males: n (%) | 19 (63.3) | 12 (75) | 16 (100) | 0.014 |
| Risk factors | ||||
| BMI (kg/m2) mean ± SD | 29.3 ± 7.1 | 31.4 ± 4.8 | 26.8 ± 4.7 8 (50) | 0.145 |
| Smoking: n (%) | 16 (53.3) | 10 (52.5) | 0.09 | |
| Diabetes Mellitus: n (%) | 16 (53.3) | 6 (37.5) | 0.001 | |
| Hypertension: n (%) | 19 (63.3) | 7 (43) | 0.001 | |
| Dyslipidemia: n (%) | 12 (40) | 5 (31.3) | 0.001 | |
| Genes | Median Expression Level (log10) | Statistics FC (p-Value) | |||
|---|---|---|---|---|---|
| Healthy | UA | STEMI | UA/Healthy | STEMI/UA | |
| miR-137 (log10) | 0.9 | 1244 | 3163 | 1382 (<0.0001) | 2.5 (<0.001) |
| FTHL-17 (log10) | 3.76 | 6.6 | 50.4 | 1.7 (<0.0001) | 7.5 (<0.0001) |
| miR-106b-5p (log10) | 1.08 | 207 | 953 | 192 (<0.001) | 4.6 (<0.001) |
| ANAPC11 (log10) | 0.86 | 7.36 | 8.39 | 8.5 (<0.0001) | 1.1 (>0.05) |
| CTB89H12.4 (log10) | 65.3 | 3.27 | 0.94 | 20 (<0.0001) | 3.4 (<0.001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elgebaly, S.A.; Christenson, R.H.; Kandil, H.; El-Khazragy, N.; Rashed, L.; Yacoub, B.; Eldeeb, H.; Ali, M.; Sharafieh, R.; Klueh, U.; et al. Nourin-Dependent miR-137 and miR-106b: Novel Early Inflammatory Diagnostic Biomarkers for Unstable Angina Patients. Biomolecules 2021, 11, 368. https://doi.org/10.3390/biom11030368
Elgebaly SA, Christenson RH, Kandil H, El-Khazragy N, Rashed L, Yacoub B, Eldeeb H, Ali M, Sharafieh R, Klueh U, et al. Nourin-Dependent miR-137 and miR-106b: Novel Early Inflammatory Diagnostic Biomarkers for Unstable Angina Patients. Biomolecules. 2021; 11(3):368. https://doi.org/10.3390/biom11030368
Chicago/Turabian StyleElgebaly, Salwa A., Robert H. Christenson, Hossam Kandil, Nashwa El-Khazragy, Laila Rashed, Beshoy Yacoub, Heba Eldeeb, Mahmoud Ali, Roshanak Sharafieh, Ulrike Klueh, and et al. 2021. "Nourin-Dependent miR-137 and miR-106b: Novel Early Inflammatory Diagnostic Biomarkers for Unstable Angina Patients" Biomolecules 11, no. 3: 368. https://doi.org/10.3390/biom11030368
APA StyleElgebaly, S. A., Christenson, R. H., Kandil, H., El-Khazragy, N., Rashed, L., Yacoub, B., Eldeeb, H., Ali, M., Sharafieh, R., Klueh, U., & Kreutzer, D. L. (2021). Nourin-Dependent miR-137 and miR-106b: Novel Early Inflammatory Diagnostic Biomarkers for Unstable Angina Patients. Biomolecules, 11(3), 368. https://doi.org/10.3390/biom11030368