
New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice

 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Bacteria
2.2. Minimum Inhibitory (MIC) and Minimum Bactericidal Concentration (MBC)
2.3. Macromolecular Synthesis
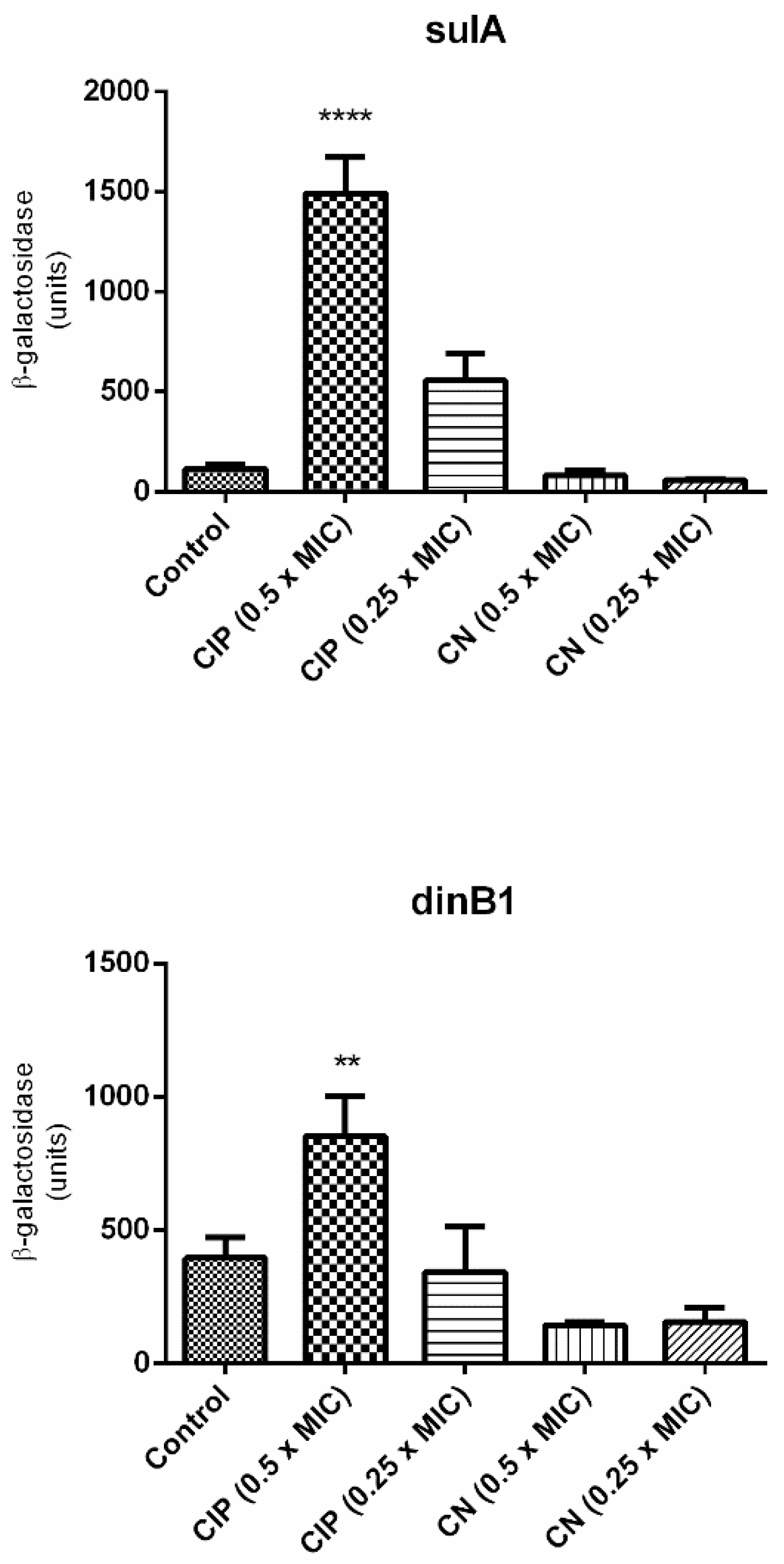
2.4. Evaluation of the Expression of Genes Associated with SOS Response
2.5. Adhesion Test with Human Epithelial Type 2 (Hep-2) Cells
2.6. Cytotoxicity Assay
2.7. Effect of Cinnamaldehyde on Tenebrio molitor Larvae
2.8. Mouse Colonization
2.9. Statistical Analysis
3. Results
3.1. Cinnamaldehyde Inhibits E. coli Growth
3.2. Cinnamaldehyde Interferes in the Macromolecular Synthesis in E. coli
3.3. Cinnamaldehyde Does Not Interfere with Cell Viability and Is Not Toxic against T. molitor Larvae
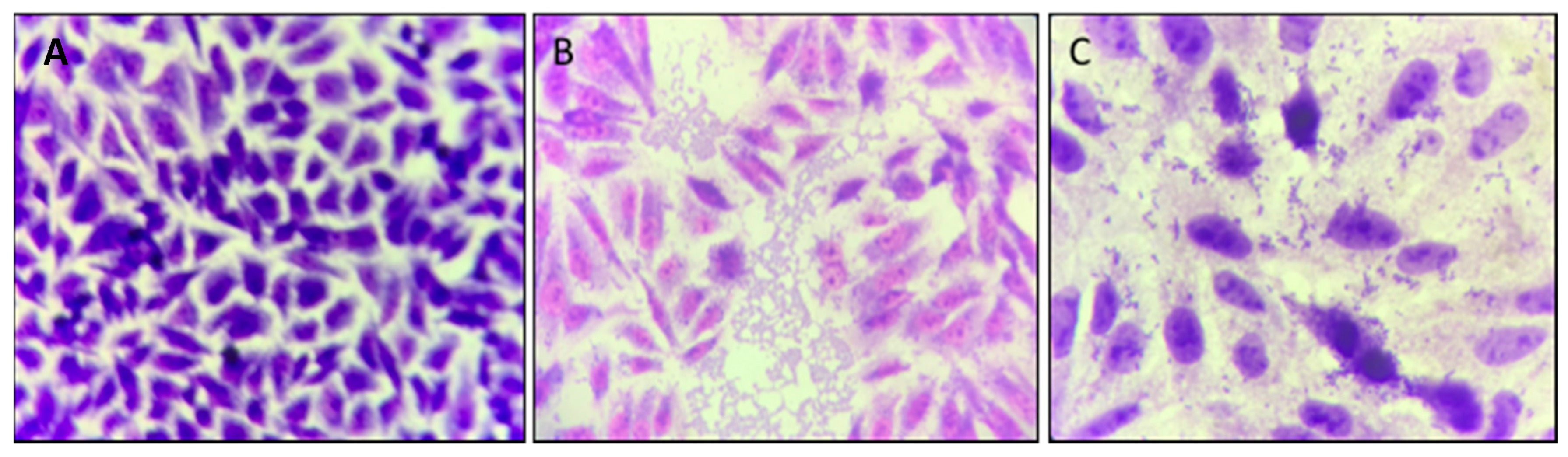
3.4. Cinnamaldehyde Promotes Reduction of EAEC 042 Aggregative Adhesion on HEp-2 Cells
3.5. Cinnamaldehyde Treatment Reduced the Intestinal Colonization of Mice by E. coli
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taghadosi, R.; Shakibaie, M.R.; Hosseini-Nave, H. Antibiotic resistance, ESBL genes, integrons, phylogenetic groups and MLVA profiles of Escherichia coli pathotypes isolated from patients with diarrhea and farm animals in south-east of Iran. Comp. Immunol. Microbiol. Infect. Dis. 2019, 63, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Tharwat, N.; El-Sherif, R.; Elnagdy, S.; Marzaban, R.; Amer, S. Virulent Escherichia coli strains among Egyptian patients with acute diarrhoea versus urinary tract infection, and their antibiotic susceptibility. Arab J. Gastroenterol. 2019, 20, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Venkatesh, V.; Kumar, R.; Kashyap, S.; Kumar, M.; Maurya, A.K.; Singh, M. Etiological agents of diarrhea in hospitalized pediatric patients with special emphasis on diarrheagenic Escherichia coli in North India. J. Lab. Phys. 2019, 11, 68. [Google Scholar] [CrossRef]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.S. Origins of the E. coli strain causing an outbreak of hemolytic-uremic syndrome in Germany. Nejm 2011, 365, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.H.; Olsen, K.E.; Struve, C.; Krogfelt, K.A.; Petersen, A.M. Epidemiology and clinical manifestations of enteroaggregative Escherichia coli. Clin. Microbiol. Rev. 2014, 27, 614–630. [Google Scholar] [CrossRef]
- Aijuka, M.; Santiago, A.E.; Girón, J.A.; Nataro, J.P.; Buys, E.M. Enteroaggregative Escherichia coli is the predominant diarrheagenic E. coli pathotype among irrigation water and food sources in South Africa. Int. J. Food Microbiol. 2018, 278, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. MAL-ED Network Investigators. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef]
- Bouzari, S.; Dashti, A.; Jafari, A.; Oloomi, M. Immune response against adhesins of enteroaggregative Escherichia coli immunized by three different vaccination strategies (DNA/DNA, Protein/Protein, and DNA/Protein) in mice. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 215–225. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance; World Health Organization (WHO): Geneva, Switzerland, 2014. [Google Scholar]
- Wang, Y.; Zhang, Y.; Shi, Y.Q.; Pan, X.H.; Lu, Y.H.; Cao, P. Antibacterial effects of cinnamon (Cinnamomum zeylanicum) bark essential oil on Porphyromonas gingivalis. Microb. Pathog. 2018, 116, 26–32. [Google Scholar] [CrossRef]
- Narayanan, A.; Muyyarikkandy, M.S.; Mooyottu, S.; Venkitanarayanan, K.; Amalaradjou, M. Oral supplementation of trans-cinnamaldehyde reduces uropathogenic Escherichia coli colonization in a mouse model. Lett. Appl. microbiol. 2017, 64, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Yang, H.; Tang, P.; Liu, J.; Wang, W.; Li, H. Interactions of cinnamaldehyde and its metabolite cinnamic acid with human serum albumin and interference of other food additives. Food Chem. 2018, 243, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiari, S.; Jafari, S.; Taheri, J.B.; Kashi, T.S.J.; Namazi, Z.; Iman, M.; Poorberafeyi, M. The Effects of Cinnamaldehyde (Cinnamon Derivatives) and Nystatin on Candida Albicans and Candida Glabrata. Open Access Maced. J. Med. Sci. 2019, 7, 1067. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.N.; Khan, S.; Iqbal, J.; Khan, R.; Khan, A.U. Enhanced killing and antibiofilm activity of encapsulated cinnamaldehyde against Candida albicans. Front Microbiol. 2017, 8, 1641. [Google Scholar] [CrossRef] [PubMed]
- He, T.F.; Wang, L.H.; Niu, D.B.; Wen, Q.H.; Zeng, X.A. Cinnamaldehyde inhibit Escherichia coli associated with membrane disruption and oxidative damage. Arch. Microbiol. 2018, 201, 1–8. [Google Scholar] [CrossRef]
- Rogiers, G.; Kebede, B.T.; Van Loey, A.; Michiels, C.W. Membrane fatty acid composition as a determinant of Listeria monocytogenes sensitivity to trans-cinnamaldehyde. Res. Microbiol. 2017, 168, 536–546. [Google Scholar] [CrossRef]
- Topa, S.H.; Subramoni, S.; Palombo, E.A.; Kingshott, P.; Rice, S.A.; Blackall, L.L. Cinnamaldehyde disrupts biofilm formation and swarming motility of Pseudomonas aeruginosa. Microbiology 2018, 164, 1087–1097. [Google Scholar] [CrossRef]
- Ferro, T.A.; Araújo, J.M.; Pinto, B.L.S.; Santos, J.S.; Souza, E.B.; Silva, B.L.; Monteiro-Neto, V. Cinnamaldehyde Inhibits Staphylococcus aureus Virulence Factors and Protects against Infection in a Galleria mellonella Model. Front. Microbiol. 2016, 7, 2052. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Lin, I.F.; Chen, C.J.; Chang, S.T. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food Chem. Toxicol. 2015, 46, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.J.; Sousa, F.I.; Pereira, D.M.; Ferro, T.A.; Pereira, I.C.; Silva, B.L.; Nascimento, J.L. Cinnamaldehyde modulates LPS-induced systemic inflammatory response syndrome through TRPA1-dependent and independent mechanisms. Int. Immunopharmacol. 2016, 34, 60–70. [Google Scholar] [CrossRef]
- Sartorius, T.; Peter, A.; Schulz, N.; Drescher, A.; Bergheim, I.; Machann, J.; Schick, F.; Siegel-Axel, D.; Schürmann, A.; Weigert, C.; et al. Cinnamon extract improves insulin sensitivity in the brain and lowers liver fat in mouse models of obesity. PLoS ONE 2014, 9, e92358. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.K.; Hwang, J.S.; So, J.S.; Lee, C.G.; Sahoo, A.; Ryu, J.H.; Jeon, W.K.; Ko, B.S.; Im, C.R.; Lee, S.H.; et al. Cinnamon extract induces tumor cell death through inhibition of NFkB and AP1. BMC Cancer 2010, 10, 392. [Google Scholar] [CrossRef]
- Schoene, N.W.; Kelly, M.A.; Polansky, M.M.; Anderson, R.A. Water-soluble polymeric polyphenols from cinnamon inhibit proliferation and alter cell cycle distribution patterns of hematologic tumor cell lines. Cancer Lett. 2005, 230, 134–140. [Google Scholar] [CrossRef]
- Nataro, J.P.; Baldini, M.M.; Kaper, J.B.; Black, R.E.; Bravo, N.; Levine, M.M. Detection of an adherence factor of enteropathogenic Escherichia coli with a DNA probe. J. Infect. Dis. 1985, 152, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Boyer, H.W.; Roulland-Dussoix, D. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J Molbiol 1969, 41, 459–472. [Google Scholar] [CrossRef]
- Frimodt-Møller, J.; Charbon, G.; Krogfelt, K.A.; Løbner-Olesen, A. Control regions for chromosome replication are conserved with respect to sequence and location among Escherichia coli strains. Front. Microbiol. 2015, 6, 1011. [Google Scholar] [CrossRef] [PubMed]
- Frimodt-Møller, J.; Charbon, G.; Krogfelt, K.A.; Løbner-Olesen, A. DNA replication control is linked to genomic positioning of control regions in Escherichia coli. PLoS Genet. 2016, 12, e1006286. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.J.; Walker, G.C. DNA-damaging agents stimulate gene expression at specific loci in Escherichia coli. Proc. Natl. Acad. Sci. USA 1980, 77, 2819–2823. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972; pp. 352–355. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Seventh Informational Supplement; CLSI Document M100-S27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Scaletsky, I.C.; Silva, M.L.; Trabulsi, L.R. Distinctive patterns of adherence of enteropathogenic Escherichia coli to HeLa cells. Infect. Immun. 1984, 45, 534–536. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Silva, T.F.; Filho, J.R.N.C.; Fonsêca, M.M.L.B.; Santos, N.M.; Silva, A.C.B.; Zagmignan, A.; Abreu, A.G.; Silva, A.P.S.; Lima, V.L.M.; Silva, N.H.; et al. Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury. Pharmaceuticals 2020, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Harrington, S.M.; Sheikh, J.; Henderson, I.R.; Ruiz-Perez, F.; Cohen, P.S.; Nataro, J.P. The Pic protease of enteroaggregative Escherichia coli promotes intestinal colonization and growth in the presence of mucin. Infect. Immun. 2009, 77, 2465–2473. [Google Scholar] [CrossRef]
- Abreu, A.G.; Abe, C.M.; Nunes, K.O.; Moraes, C.T.; Chavez-Duenas, L.; Navarro-Garcia, F.; Elias, W.P. The serine protease Pic as a virulence factor of atypical enteropathogenic Escherichia coli. Gut Microbes. 2016, 7, 115–125. [Google Scholar] [CrossRef]
- Goodman, M.F.; Woodgate, R. Translesion DNA polymerases. Cold Spring Harb. Perspect. Biol. 2013, 5, a010363. [Google Scholar] [CrossRef]
- Ordonez, H.; Uson, M.L.; Shuman, S. Characterization of three mycobacterial DinB (DNA polymerase IV) paralogs highlights DinB2 as naturally adept at ribonucleotide incorporation. Nucleic Acids Res. 2014, 42, 11056–11070. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.E.; Prakash, S.; Prakash, L. The human DINB1 gene encodes the DNA polymerase Polθ. Proc. Natl. Acad. Sci. USA 2000, 97, 3838–3843. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.D.; Smith, B.T.; Godoy, V.G.; Walker, G.C. The SOS response: Recent insights into umuDC-dependent mutagenesis and DNA damage tolerance. Annu. Rev. Genet. 2000, 34. [Google Scholar] [CrossRef]
- Nazir, A.; Harinarayanan, R. Inactivation of cell division protein FtsZ by SulA makes Lon indispensable for the viability of a ppGpp0 strain of Escherichia coli. J. Bacteriol. 2016, 198, 688–700. [Google Scholar] [CrossRef]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sheng, J.; Huang, G.; Ma, R.; Yin, F.; Song, D.; Ma, S. Design, synthesis and antibacterial activity of cinnamaldehyde derivatives as inhibitors of the bacterial cell division protein FtsZ. Eur. J. Med. Chem. 2015, 97, 32–41. [Google Scholar] [CrossRef]
- Yuan, W.; Yuk, H.G. Effect of sublethal thymol, carvacrol and trans-cinnamaldehyde adaptation on virulence properties of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2019, 85, e00271-19. [Google Scholar] [CrossRef] [PubMed]
- Malheiro, J.F.; Maillard, J.Y.; Borges, F.; Simões, M. Evaluation of cinnamaldehyde and cinnamic acid derivatives in microbial growth control. Int. Biodeter. Biodegrad. 2019, 141, 71–78. [Google Scholar] [CrossRef]
- Firmino, D.F.; Cavalcante, T.T.; Gomes, G.A.; Firmino, N.; Rosa, L.D.; de Carvalho, M.G.; Catunda, F.E., Jr. Antibacterial anAntibiofilm Activities of Cinnamomum Sp. Essential Oil and Cinnamaldehyde: Antimicrobial Activities. Sci. World J. 2018, 2018, 7405736. [Google Scholar] [CrossRef]
- Field, D.; Baghou, I.; Rea, M.; Gardiner, G.; Ross, R.; Hill, C. Nisin in Combination with Cinnamaldehyde and EDTA to Control Growth of Escherichia coli Strains of Swine Origin. Antibiotics 2017, 6, 35. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference |
|---|---|---|
| E. coli 042 | Standard strain for studies with enteroaggregative E. coli (EAEC), isolated from an outbreak of diarrhea in Peru. | [24] |
| E. coli HB101 | Non-pathogenic strain, used as negative control of adhesion assays. | [25] |
| E. coli MG1655 | Non-pathogenic wild type strain used as control in SOS and radioisotope assays. | [26] [27] |
| E. coli ALO4696 | MG1655; sulA::lacZ. | This work |
| E. coli ALO562 | GW1010 dinB1::Mud(Ap, lac) | [28] |
| E. coli BW25113 (ALO 4628) | Wild-type strain from Keio Collection | [29] |
| E. coli JW3682 (ALO 3542) | BW25113; ΔyidD::kan, KanR | [29] |
| E. coli JW3820 (ALO 3544) | BW25113; Δfre::kan, KanR | [29] |
| E. coli JW2513 (ALO 3545) | BW25113; ΔiscU::kan, KanR | [29] |
| E. coli JW3879-1 (ALO 4554) | BW25113; ΔsodA::kan, KanR | [29] |
| E. coli JW1648-1 (ALO 4555) | BW25113; ΔsodB::kan, KanR | [29] |
| Precursors | Marked Precursor | Function | Time of Incorporation | Antibiotic Control |
|---|---|---|---|---|
| Thymidine | H3-Thymidine | DNA Replication | 4 min | Nalidixic Acid |
| Uridine | H3-Uridine | RNA Synthesis | 2 min | Rifampicin |
| Glucosamine | H3-Glucosamine | Cell wall Synthesis | 20 min | Ampicillin |
| Arginine | H3-Arginine | Protein Synthesis | 4 min | Chloramphenicol |
| E. coli Strain | MIC (µg/mL) | MBC (µg/mL) |
|---|---|---|
| 042 | 780 | 1560 |
| HB101 | 780 | 1560 |
| 4628 | 1560 | 1560 |
| 3542 | 780 | 3120 |
| 3544 | 780 | 780 |
| 3545 | 780 | 3120 |
| 4554 | 3120 | 3120 |
| 4555 | 3120 | 3120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, W.A.; Pereira, C.D.S.; Assunção, R.G.; da Silva, I.S.C.; Rego, F.S.; Alves, L.S.R.; Santos, J.S.; Nogueira, F.J.R.; Zagmignan, A.; Thomsen, T.T.; et al. New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice. Biomolecules 2021, 11, 302. https://doi.org/10.3390/biom11020302
Pereira WA, Pereira CDS, Assunção RG, da Silva ISC, Rego FS, Alves LSR, Santos JS, Nogueira FJR, Zagmignan A, Thomsen TT, et al. New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice. Biomolecules. 2021; 11(2):302. https://doi.org/10.3390/biom11020302
Chicago/Turabian StylePereira, Wellison A., Carlos Drielson S. Pereira, Raíssa G. Assunção, Iandeyara Savanna C. da Silva, Fabrícia S. Rego, Leylane S. R. Alves, Juliana S. Santos, Francisco Jonathas R. Nogueira, Adrielle Zagmignan, Thomas T. Thomsen, and et al. 2021. "New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice" Biomolecules 11, no. 2: 302. https://doi.org/10.3390/biom11020302
APA StylePereira, W. A., Pereira, C. D. S., Assunção, R. G., da Silva, I. S. C., Rego, F. S., Alves, L. S. R., Santos, J. S., Nogueira, F. J. R., Zagmignan, A., Thomsen, T. T., Løbner-Olesen, A., Krogfelt, K. A., da Silva, L. C. N., & Abreu, A. G. (2021). New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice. Biomolecules, 11(2), 302. https://doi.org/10.3390/biom11020302