
CB2 Receptor Involvement in the Treatment of Substance Use Disorders

 ,
, 
 ,
, 

Abstract
:1. Introduction
The Endocannabinoid System: Role in SUD
2. Methods
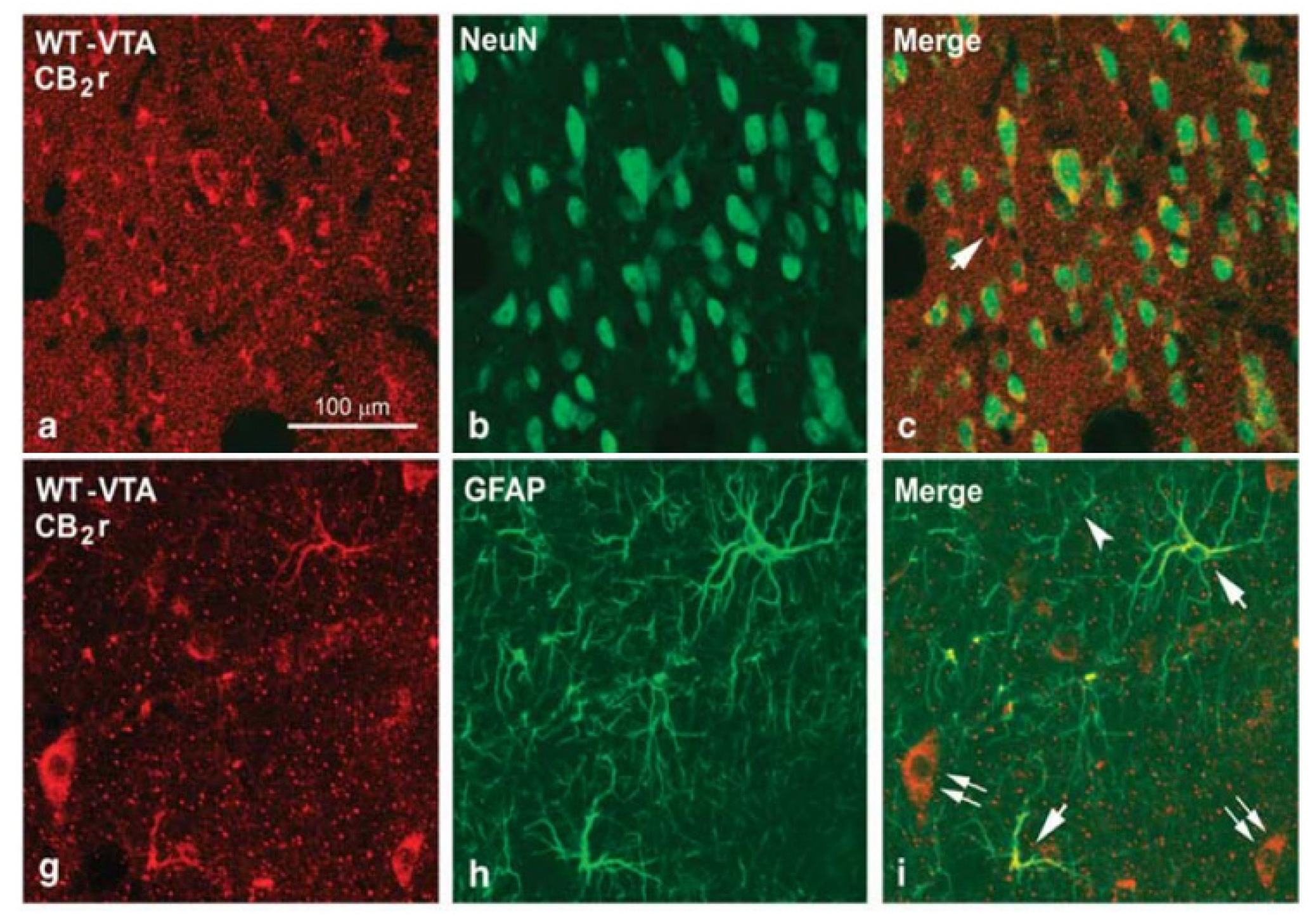
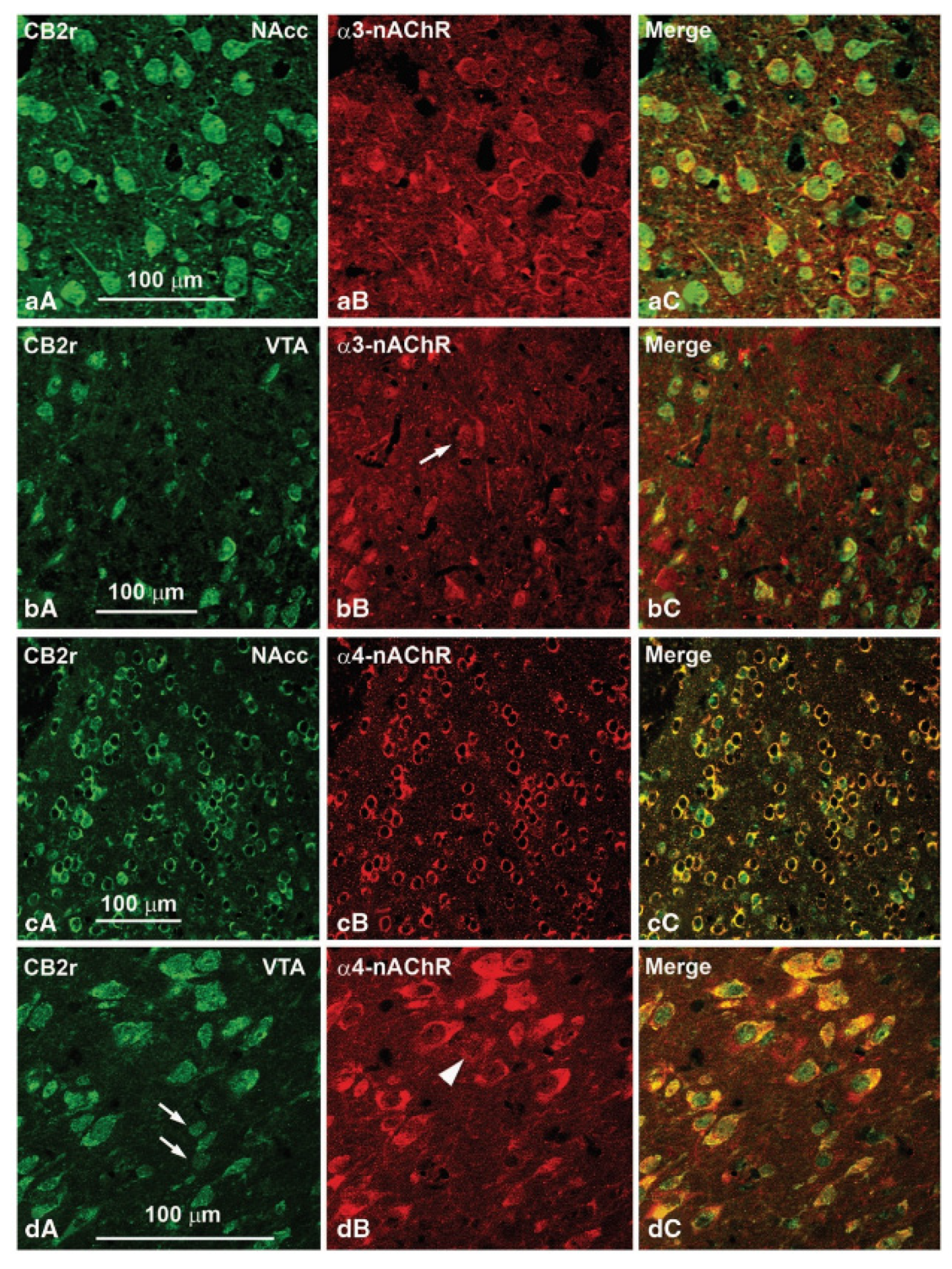
3. Expression, Distribution, and Functional Involvement of CB2r in the Reward System
4. Therapeutic Potential of CB2r in the Management of Substance Use Disorders
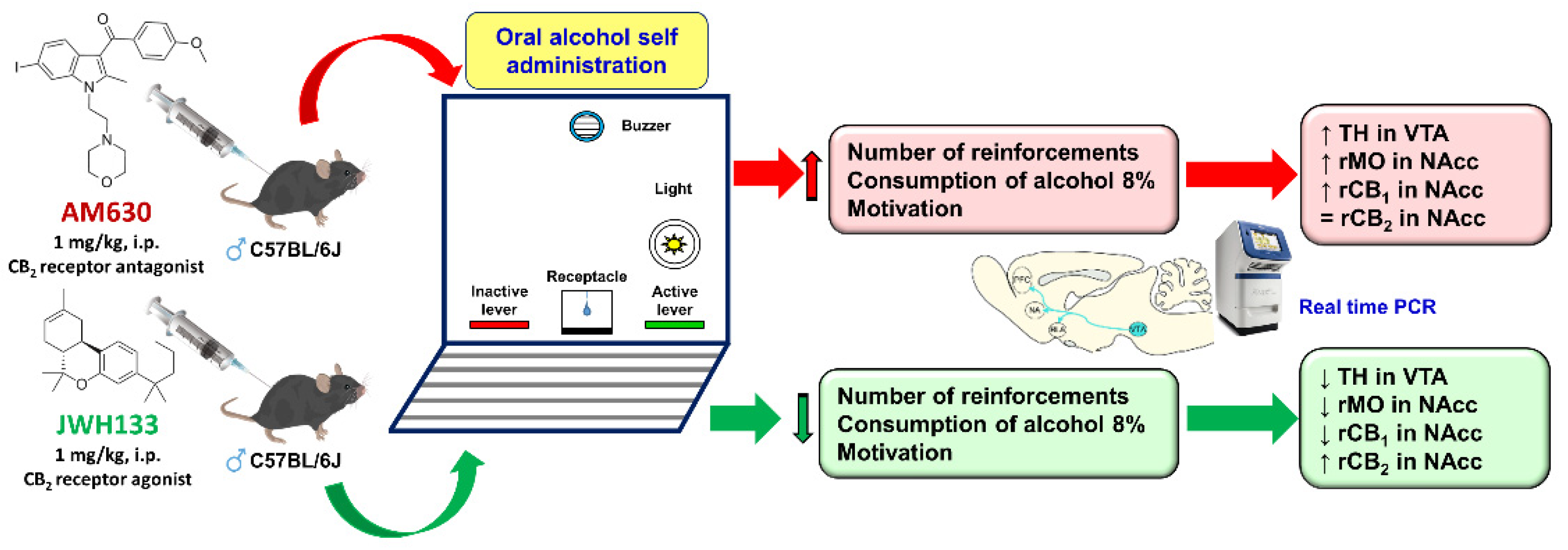
4.1. CB2r and Alcohol
4.2. CB2r and Cocaine
4.3. CB2r and Nicotine
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- APA. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; DSM-V; American Psychiatric Association (APA): Washington, DC, USA, 2014. [Google Scholar]
- United Nations Office on Drugs and Crime. World Drug Report; UNODC: Vienna, Austria, 2021. [Google Scholar]
- WHO. Global Status Report on Alcohol and Health; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Piomelli, D. The molecular logic of endocannabinoid signalling. Nat. Rev. Neurosci. 2003, 4, 873–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Katona, I.; Freund, T.F. Multiple Functions of Endocannabinoid Signaling in the Brain. Annu. Rev. Neurosci. 2012, 35, 529–558. [Google Scholar] [CrossRef] [Green Version]
- Mackie, K. Distribution of cannabinoid receptors in the central and peripheral nervous system. In Cannabinoids. Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 299–325. [Google Scholar] [CrossRef]
- De Fonseca, F.R.; Del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The Endocannabinoid System: Physiology and Pharmacology. Alcohol Alcohol. 2005, 40, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Tsou, K.; Brown, S.; Sañudo-Peña, M.; Mackie, K.; Walker, J. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 1998, 83, 393–411. [Google Scholar] [CrossRef]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Puente, N.; Elezgarai, I.; Ruehle, S.; Lutz, B.; Reguero, L.; Gerrikagoitia, I.; Marsicano, G.; Grandes, P. Anatomical characterization of the cannabinoid CB1receptor in cell-type-specific mutant mouse rescue models. J. Comp. Neurol. 2017, 525, 302–318. [Google Scholar] [CrossRef] [PubMed]
- Piazza, P.V.; Cota, D.; Marsicano, G. The CB1 Receptor as the Cornerstone of Exostasis. Neuron 2017, 93, 1252–1274. [Google Scholar] [CrossRef] [PubMed]
- Busquets-Garcia, A.; Bains, J.; Marsicano, G. CB1 Receptor Signaling in the Brain: Extracting Specificity from Ubiquity. Neuropsychopharmacology 2018, 43, 4–20. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of Central and Peripheral Cannabinoid Receptors in Human Immune Tissues and Leukocyte Subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Cabral, G.A.; Griffin-Thomas, L. Emerging role of the cannabinoid receptor CB2in immune regulation: Therapeutic prospects for neuroinflammation. Expert Rev. Mol. Med. 2009, 11, e3. [Google Scholar] [CrossRef] [Green Version]
- Benito, C.; Nunez, E.; Tolon, R.M.; Carrier, E.J.; Rabano, A.; Hillard, C.J.; Romero, J. Cannabinoid CB2 receptors and fatty acid amide hydrolase are selectively overexpressed in neuritic plaque-associated glia in Alzheimer’s disease brains. J. Neurosci. 2003, 23, 11136–11141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yiangou, Y.; Facer, P.; Durrenberger, P.; Chessell, I.P.; Naylor, A.; Bountra, C.; Banati, R.R.; Anand, P. COX-2, CB2 and P2X7-immunoreactivities are increased in activated microglial cells/macrophages of multiple sclerosis and amyotrophic lateral sclerosis spinal cord. BMC Neurol. 2006, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, M.; Sanchez, C.; Galve-Roperh, I. Control of the cell survival/death decision by cannabinoids. J. Mol. Med. 2000, 78, 613–625. [Google Scholar] [CrossRef]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Identification and Functional Characterization of Brainstem Cannabinoid CB 2 Receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.-P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.-R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S. Neuropsychobiological Evidence for the Functional Presence and Expression of Cannabinoid CB2 Receptors in the Brain. Neuropsychobiology 2006, 54, 231–246. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the Presence and Functional Expression of Cannabinoid CB2 Receptors in Brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.; Pérez-Ortiz, J.; Gutiérrez-Adán, A.; Manzanares, J. Depression-resistant endophenotype in mice overexpressing cannabinoid CB2 receptors. Br. J. Pharmacol. 2010, 160, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Gao, M.; Liu, Q.-R.; Bi, G.-H.; Li, X.; Yang, H.-J.; Gardner, E.L.; Wu, J.; Xi, Z.-X. Cannabinoid CB2receptors modulate midbrain dopamine neuronal activity and dopamine-related behavior in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E5007–E5015. [Google Scholar] [CrossRef] [Green Version]
- Cabral, G.A.; Raborn, E.S.; Griffin, L.; Dennis, J.; Marciano-Cabral, F. CB2 receptors in the brain: Role in central immune function. Br. J. Pharmacol. 2008, 153, 240–251. [Google Scholar] [CrossRef] [Green Version]
- García-Gutiérrez, M.S.; Navarrete, F.; Navarro, G.; Reyes-Resina, I.; Franco, R.; Lanciego, J.L.; Giner, S.; Manzanares, J. Alterations in Gene and Protein Expression of Cannabinoid CB2 and GPR55 Receptors in the Dorsolateral Prefrontal Cortex of Suicide Victims. Neurotherapeutics 2018, 15, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.-R.; Canseco-Alba, A.; Zhang, H.-Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Escosteguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; García-Gutiérrez, M.S.; Aracil-Fernández, A.; Lanciego, J.L.; Manzanares, J. Cannabinoid CB1 and CB2 Receptors, and Monoacylglycerol Lipase Gene Expression Alterations in the Basal Ganglia of Patients with Parkinson’s Disease. Neurotherapeutics 2018, 15, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid Receptors and Their Ligands: Beyond CB1and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [Green Version]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- Alger, B.E.; Kim, J. Supply and demand for endocannabinoids. Trends Neurosci. 2011, 34, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Manzanares, J.; Cabañero, D.; Puente, N.; García-Gutiérrez, M.S.; Grandes, P.; Maldonado, R. Role of the endocannabinoid system in drug addiction. Biochem. Pharmacol. 2018, 157, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Filip, M.; Gołda, A.; Zaniewska, M.; McCreary, A.C.; Nowak, E.; Kolasiewicz, W.; Przegaliński, E. Involvement of cannabinoid CB1 receptors in drug addiction: Effects of rimonabant on behavioral responses induced by cocaine. Pharmacol. Rep. 2007, 58, 806–819. [Google Scholar]
- Parolaro, D.; Vigano’, D.; Realini, N.; Rubino, T. Role of endocannabinoids in regulating drug dependence. Neuropsychiatr. Dis. Treat. 2008, 3, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Le Foll, B.; Forget, B.; Aubin, H.-J.; Goldberg, S.R. Blocking cannabinoid CB1 receptors for the treatment of nicotine dependence: Insights from pre-clinical and clinical studies. Addict. Biol. 2008, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, B.; Goldberg, S.R. Cannabinoid CB1 Receptor Antagonists as Promising New Medications for Drug Dependence. J. Pharmacol. Exp. Ther. 2005, 312, 875–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinen, H.; Hyytiä, P. Ethanol Self-Administration Is Regulated by CB1 Receptors in the Nucleus Accumbens and Ventral Tegmental Area in Alcohol-Preferring AA Rats. Alcohol. Clin. Exp. Res. 2008, 32, 1976–1983. [Google Scholar] [CrossRef]
- Lallemand, F.; De Witte, P. SR147778, a CB1 Cannabinoid Receptor Antagonist, Suppresses Ethanol Preference in Chronically Alcoholized Wistar Rats. Alcohol 2006, 39, 125–134. [Google Scholar] [CrossRef]
- Lallemand, F.; Soubrié, P.; De Witte, P. Effects of cb1 cannabinoid receptor blockade on ethanol preference after chronic alcohol administration combined with repeated re-exposures and withdrawals. Alcohol Alcohol. 2004, 39, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Lallemand, F.; Soubrié, P.H.; De Witte, P.H. Effects of CB1 cannabinoid receptor blockade on ethanol preference after chronic ethanol administration. Alcohol. Clin. Exp. Res. 2001, 25, 1317–1323. [Google Scholar] [CrossRef]
- Soyka, M.; Koller, G.; Schmidt, P.; Lesch, O.-M.; Leweke, M.; Fehr, C.; Gann, H.; Mann, K.F. Cannabinoid Receptor 1 Blocker Rimonabant (SR 141716) for Treatment of Alcohol Dependence: Results from a placebo-controlled, double-blind trial. J. Clin. Psychopharmacol. 2008, 28, 317–324. [Google Scholar] [CrossRef] [PubMed]
- George, D.T.; Herion, D.W.; Jones, C.L.; Phillips, M.J.; Hersh, J.; Hill, D.; Heilig, M.; Ramchandani, V.A.; Geyer, C.; Spero, D.E.; et al. Rimonabant (SR141716) has no effect on alcohol self-administration or endocrine measures in nontreatment-seeking heavy alcohol drinkers. Psychopharmacology 2009, 208, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Bonci, A. Getting to the core of addiction: Hooking CB2 receptor into drug abuse? Nat. Med. 2012, 18, 504–505. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wang, L.; Xie, X.-Q. Latest advances in novel cannabinoid CB2 ligands for drug abuse and their therapeutic potential. Future Med. Chem. 2012, 4, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Aracil-Fernández, A.; Trigo, J.M.; García-Gutiérrez, M.S.; Álvaro, A.O.; Ternianov, A.; Navarro, D.; Robledo, P.; Berbel, P.; Maldonado, R.; Manzanares, J. Decreased Cocaine Motor Sensitization and Self-Administration in Mice Overexpressing Cannabinoid CB2 Receptors. Neuropsychopharmacology 2012, 37, 1749–1763. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; Rodriguez-Arias, M.; Martin-García, E.; Navarro, D.; García-Gutiérrez, M.S.; Aguilar, M.A.; Aracil-Fernández, A.; Berbel, P.; Miñarro, J.; Maldonado, R.; et al. Role of CB2 Cannabinoid Receptors in the Rewarding, Reinforcing, and Physical Effects of Nicotine. Neuropsychopharmacology 2013, 38, 2515–2524. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Álvaro, A.; Ternianov, A.; Aracil-Fernández, A.; Navarrete, F.; García-Gutiérrez, M.S.; Manzanares, J. Role of cannabinoid CB2receptor in the reinforcing actions of ethanol. Addict. Biol. 2015, 20, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.-P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Brain Neuronal CB2 Cannabinoid Receptors in Drug Abuse and Depression: From Mice to Human Subjects. PLoS ONE 2008, 3, e1640. [Google Scholar] [CrossRef] [Green Version]
- Benito, C.; Tolón, R.M.; Pazos, M.R.; Nuñez, E.; Castillo, A.I.; Romero, J. Cannabinoid CB2 receptors in human brain inflammation. Br. J. Pharmacol. 2008, 153, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turcotte, C.; Blanchet, M.-R.; LaViolette, M.; Flamand, N. The CB2 receptor and its role as a regulator of inflammation. Cell. Mol. Life Sci. 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Ruiz, J.; Pazos, M.R.; García-Arencibia, M.; Sagredo, O.; Ramos, J.A. Role of CB2 receptors in neuroprotective effects of cannabinoids. Mol. Cell. Endocrinol. 2008, 286, S91–S96. [Google Scholar] [CrossRef] [Green Version]
- Ternianov, A.; Pérez-Ortiz, J.M.; Solesio, M.E.; García-Gutiérrez, M.S.; Álvaro, A.O.; Navarrete, F.; Leiva, C.; Galindo, M.F.; Manzanares, J. Overexpression of CB2 cannabinoid receptors results in neuroprotection against behavioral and neurochemical alterations induced by intracaudate administration of 6-hydroxydopamine. Neurobiol. Aging 2012, 33, 421.e1–421.e6. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; García-Bueno, B.; Zoppi, S.; Leza, J.C.; Manzanares, J. Chronic blockade of cannabinoid CB2 receptors induces anxiolytic-like actions associated with alterations in GABAA receptors. Br. J. Pharmacol. 2012, 165, 951–964. [Google Scholar] [CrossRef] [Green Version]
- García-Gutiérrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Doods, H.; Treede, R.-D.; Ceci, A. Depression-like behaviour in rats with mononeuropathy is reduced by the CB2-selective agonist GW405833. Pain 2009, 143, 206–212. [Google Scholar] [CrossRef]
- Liu, Q.-R.; Pan, C.-H.; Hishimoto, A.; Li, C.-Y.; Xi, Z.-X.; Llorente-Berzal, A.; Viveros, M.-P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.-X.; Peng, X.; Li, X.; Song, R.; Zhang, H.; Liu, Q.-R.; Yang, H.-J.; Bi, G.-H.; Li, J.; Gardner, E.L. Brain cannabinoid CB2 receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-Y.; Xi, Z.-X. Chapter 70—Cannabinoid CB2 receptor: A new target for treatment of cocaine addiction. In The Neuroscience of Cocaine Mechanisms and Treatment; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Ma, Z.; Gao, F.; Larsen, B.; Gao, M.; Luo, Z.; Chen, D.; Ma, X.; Qiu, S.; Zhou, Y.; Xie, J.; et al. Mechanisms of cannabinoid CB2 receptor-mediated reduction of dopamine neuronal excitability in mouse ventral tegmental area. EBioMedicine 2019, 42, 225–237. [Google Scholar] [CrossRef] [Green Version]
- García-Gutiérrez, M.S.; Ortega-Álvaro, A.; Busquets-García, A.; Pérez-Ortiz, J.M.; Caltana, L.; Ricatti, M.J.; Brusco, A.; Maldonado, R.; Manzanares, J. Synaptic plasticity alterations associated with memory impairment induced by deletion of CB2 cannabinoid receptors. Neuropharmacology 2013, 73, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Ratano, P.; Petrella, C.; Forti, F.; Passeri, P.P.; Morena, M.; Palmery, M.; Trezza, V.; Severini, C.; Campolongo, P. Pharmacological inhibition of 2-arachidonoilglycerol hydrolysis enhances memory consolidation in rats through CB2 receptor activation and mTOR signaling modulation. Neuropharmacology 2018, 138, 210–218. [Google Scholar] [CrossRef]
- Navarrete, F.; Pérez-Ortiz, J.M.; Manzanares, J. Cannabinoid CB2 receptor-mediated regulation of impulsive-like behaviour in DBA/2 mice. Br. J. Pharmacol. 2012, 165, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Arias, M.; Navarrete, F.; Blanco-Gandía, M.C.; Arenas, M.C.; Aguilar, M.A.; Bartoll-Andrés, A.; Valverde, O.; Miñarro, J.; Manzanares, J. Role of CB2 receptors in social and aggressive behavior in male mice. Psychopharmacology 2015, 232, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Marco, E.M.; García-Gutiérrez, M.S.; Bermúdez-Silva, F.-J.; Moreira, F.; Guimarães, F.; Manzanares, J.; Viveros, M.-P. Endocannabinoid system and psychiatry: In search of a neurobiological basis for detrimental and potential therapeutic effects. Front. Behav. Neurosci. 2011, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, H.; Iwasaki, S.; Teasenfitz, L.; Higuchi, S.; Horiuchi, Y.; Saito, T.; Arinami, T.; Onaivi, E.S. Involvement of cannabinoid CB2 receptor in alcohol preference in mice and alcoholism in humans. Pharmacogenom. J. 2006, 7, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, A.; Rivera, P.; Pavon, F.J.; Decara, J.; Suárez, J.; de Fonseca, F.R.; Parsons, L.H. Differential Effects of Single versus Repeated Alcohol Withdrawal on the Expression of Endocannabinoid System-Related Genes in the Rat Amygdala. Alcohol. Clin. Exp. Res. 2012, 36, 984–994. [Google Scholar] [CrossRef] [Green Version]
- Marín, L.S.; Pavon, F.J.; Decara, J.; Suarez, J.; Gavito, A.; Castilla-Ortega, E.; De Fonseca, F.R.; Serrano, A. Effects of Intermittent Alcohol Exposure on Emotion and Cognition: A Potential Role for the Endogenous Cannabinoid System and Neuroinflammation. Front. Behav. Neurosci. 2017, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gutierrez, M.S.; Aracil-Fernandez, A.; Navarrete, F.; Manzanares, J. Cannabinoid CB2 receptor gene expression alterations in the dorsolateral prefrontal cortex and nucleus accumbens of alcoholic patients. In Proceedings of the 3rd International Congress on Dual Pathology, Barcelona, Spain, 23–26 October 2013. [Google Scholar]
- Pradier, B.; Erxlebe, E.; Markert, A.; Rácz, I. Interaction of cannabinoid receptor 2 and social environment modulates chronic alcohol consumption. Behav. Brain Res. 2015, 287, 163–171. [Google Scholar] [CrossRef]
- Powers, M.S.; Breit, K.R.; Chester, J.A. Genetic Versus Pharmacological Assessment of the Role of Cannabinoid Type 2 Receptors in Alcohol Reward-Related Behaviors. Alcohol. Clin. Exp. Res. 2015, 39, 2438–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Mansouri, S.; Ojha, S.; Al Maamari, E.; Al Ameri, M.; Nurulain, S.M.; Bahi, A. The cannabinoid receptor 2 agonist, β-caryophyllene, reduced voluntary alcohol intake and attenuated ethanol-induced place preference and sensitivity in mice. Pharmacol. Biochem. Behav. 2014, 124, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; García-Gutiérrez, M.S.; Manzanares, J. Pharmacological regulation of cannabinoid CB2 receptor modulates the reinforcing and motivational actions of ethanol. Biochem. Pharmacol. 2018, 157, 227–234. [Google Scholar] [CrossRef]
- Martín-Sánchez, A.; Warnault, V.; Montagud-Romero, S.; Pastor, A.; Mondragón, N.; De La Torre, R.; Valverde, O. Alcohol-induced conditioned place preference is modulated by CB2 cannabinoid receptors and modifies levels of endocannabinoids in the mesocorticolimbic system. Pharmacol. Biochem. Behav. 2019, 183, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Canseco-Alba, A.; Schanz, N.; Ishiguro, H.; Liu, Q.-R.; Onaivi, E.S. Behavioral Evaluation of Seeking and Preference of Alcohol in Mice Subjected to Stress. Bio-Protocol 2018, 8, e3061. [Google Scholar] [CrossRef] [PubMed]
- Trebicka, J.; Racz, I.; Siegmund, S.V.; Cara, E.; Granzow, M.; Schierwagen, R.; Klein, S.; Wojtalla, A.; Hennenberg, M.; Huss, S.; et al. Role of cannabinoid receptors in alcoholic hepatic injury: Steatosis and fibrogenesis are increased in CB2 receptor-deficient mice and decreased in CB1 receptor knockouts. Liver Int. 2011, 31, 860–870. [Google Scholar] [CrossRef]
- Denaës, T.; Lodder, J.; Chobert, M.-N.; Ruiz, I.; Pawlotsky, J.-M.; Lotersztajn, S.; Teixeira-Clerc, F. The Cannabinoid Receptor 2 Protects Against Alcoholic Liver Disease via a Macrophage Autophagy-Dependent Pathway. Sci. Rep. 2016, 6, 28806. [Google Scholar] [CrossRef] [Green Version]
- Rivera, P.; Blanco, E.; Bindila, L.; Alen, F.; Vargas, A.; Rubio, L.; Pavon, F.J.; Serrano, A.; Lutz, B.; De Fonseca, F.R.; et al. Pharmacological activation of CB2 receptors counteracts the deleterious effect of ethanol on cell proliferation in the main neurogenic zones of the adult rat brain. Front. Cell. Neurosci. 2015, 9, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczyk, P.; Miszkiel, J.; McCreary, A.C.; Filip, M.; Papp, M.; Przegaliński, E. The effects of cannabinoid CB1, CB2 and vanilloid TRPV1 receptor antagonists on cocaine addictive behavior in rats. Brain Res. 2012, 1444, 45–54. [Google Scholar] [CrossRef]
- Ignatowska-Jankowska, B.M.; Muldoon, P.P.; Lichtman, A.H.; Damaj, M.I. The cannabinoid CB2 receptor is necessary for nicotine-conditioned place preference, but not other behavioral effects of nicotine in mice. Psychopharmacology 2013, 229, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Bi, G.-H.; Li, X.; Li, J.; Qu, H.; Zhang, S.-J.; Li, C.-Y.; Onaivi, E.S.; Gardner, E.L.; Xi, Z.-X.; et al. Species Differences in Cannabinoid Receptor 2 and Receptor Responses to Cocaine Self-Administration in Mice and Rats. Neuropsychopharmacology 2015, 40, 1037–1051. [Google Scholar] [CrossRef] [Green Version]
- García-Cabrerizo, R.; García-Fuster, M.J. Opposite regulation of cannabinoid CB1 and CB2 receptors in the prefrontal cortex of rats treated with cocaine during adolescence. Neurosci. Lett. 2016, 615, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Bystrowska, B.; Frankowska, M.; Smaga, I.; Pomierny-Chamiolo, L.; Filip, M. Effects of Cocaine Self-Administration and Its Extinction on the Rat Brain Cannabinoid CB1 and CB2 Receptors. Neurotox. Res. 2018, 34, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Canseco-Alba, A.; Schanz, N.; Sanabria, B.; Zhao, J.; Lin, Z.; Liu, Q.-R.; Onaivi, E.S. Behavioral effects of psychostimulants in mutant mice with cell-type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behav. Brain Res. 2019, 360, 286–297. [Google Scholar] [CrossRef]
- Zaniewska, M.; McCreary, A.C.; Przegaliński, E.; Filip, M. Evaluation of the role of nicotinic acetylcholine receptor subtypes and cannabinoid system in the discriminative stimulus effects of nicotine in rats. Eur. J. Pharmacol. 2006, 540, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Gamaleddin, I.; Wertheim, C.; Zhu, A.Z.; Coen, K.M.; Vemuri, K.; Makryannis, A.; Goldberg, S.R.; Le Foll, B. Cannabinoid receptor stimulation increases motivation for nicotine and nicotine seeking. Addict. Biol. 2012, 17, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Gamaleddin, I.; Zvonok, A.; Makriyannis, A.; Goldberg, S.R.; Le Foll, B. Effects of a Selective Cannabinoid CB2 Agonist and Antagonist on Intravenous Nicotine Self Administration and Reinstatement of Nicotine Seeking. PLoS ONE 2012, 7, e29900. [Google Scholar] [CrossRef] [Green Version]
- Gado, F.; Di Cesare Mannelli, L.; Lucarini, E.; Bertini, S.; Cappelli, E.; Digiacomo, M.; Stevenson, L.A.; Macchia, M.; Tuccinardi, T.; Ghelardini, C.; et al. Identification of the First Synthetic Allosteric Modulator of the CB2 Receptors and Evidence of Its Efficacy for Neuropathic Pain Relief. J. Med. Chem. 2018, 62, 276–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| CB2r & Alcohol-ANIMAL STUDIES | ||||
|---|---|---|---|---|
| Genetic Manipulation CB2r | ||||
| Genetic manipulation | Specie | Experimental paradigm | Results | References |
| CB2r deletion, knock-out (CB2-/- and CNR2 KO) | Swiss ICR and C57BL/6J mice | CPP | ↑ Conditioned place preference | [50] |
| VC | ↑ Voluntary ethanol consumption and preference | |||
| OESA | ↑ Motivation to drink ethanol | |||
| CB2r deletion, knock-out (CNR2 KO) | C57BL/6J mice | FD IFD | No differences | [72] |
| VC | ↑ Alcohol drinking in group-housing conditions | |||
| CB2r deletion, knock-out (CNR2 KO) | C57BL/6J mice | CPP | ↑ Conditioned place preference | [73] |
| CB2r deletion, in dopaminergic neurons, knock out condicional (DAT-Cnr2-/-) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [77] |
| VC | ||||
| Pharmacological manipulation of CB2r | ||||
| Doses | Specie | Experimental paradigm | Results | References |
| β-caryophyllene, CB2r agonist (25, 50 and 100 mg/kg) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [74] |
| VC | ↓ Voluntary ethanol consumption and preference | |||
| JWH133, CB2r agonist (10 y 20 mg/kg) | HS/Ibg mice | CPP | No differences | [73] |
| VC | No differences | |||
| JWH133, CB2r agonist (5 mg/kg) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [28] |
| JWH133, CB2r agonist (1 mg/kg) | C57BL/6J mice | OESA | ↓ Motivation to drink ethanol | [75] |
| CB2r & Alcohol-Human Studies | ||||
| Diagnosis | Population | Variable | Results | References |
| SUD | Japanese | Q63R (CB2r) polymorphism association-SUD | ↑ Incidence | [51,68] |
| SUD | Caucasian | Relative gene expression (CB2r) | ↓ DLPFCx y NAcc | [71] |
| CB2r & Cocaine-Animals Studies | ||||
|---|---|---|---|---|
| Genetic Manipulation CB2r | ||||
| Genetic manipulation | Specie | Experimental paradigm | Results | References |
| Overexpression of the CB2r (CB2xP) | Swiss ICR mice | SM | ↓ Cocaine-induced motor sensitization | [48] |
| CPP | ↓ Conditioned place preference | |||
| ISA | ↓ Motivation for cocaine consumption | |||
| CB2r deletion in dopaminergic neurons, conditional knock-out (DAT-Cnr2-/-) | C57BL/6J mice | MA | ↑ Cocaine-induced hyperactivity | [86] |
| MS | ↑ Cocaine-induced motor sensitization (10 mg/kg) | |||
| CPP | ↓ Cocaine-induced motor sensitization (20 mg/kg) ↑ Conditioned place preference | |||
| Pharmacological manipulation of CB2r | ||||
| Doses | Specie | Experimental paradigm | Results | References |
| JWH133, CB2r antagonist (10 and 20 mg/kg) | C57BL/6J mice | ISA | ↓ Motivation for cocaine consumption | [60] |
| MA | ↓ Cocaine-induced locomotor effect | |||
| GW405833, CB2r agonist (3 and 10 mg/kg) | C57BL/6J mice | ISA | ↓ Motivation for cocaine consumption | [81] |
| SR144528, CB2r antagonist (0.1, 0.3 and 1 mg/kg) | Wistar rats | ISA | No differences in motivation for cocaine consumption ↓ Cocaine-triggered relapse (not conditioned stimulus-triggered relapse) | |
| O-1966, CB2r antagonist (1, 3, 5, 10 and 20 mg/kg) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [82] |
| JWH133, CB2r agonist (10 and 20 mg/kg) | C57BL/6J mice | ISA | ↓ Motivation for cocaine consumption | [83] |
| Long-Evans rats | ISA | ↑ Motivation for cocaine consumption | ||
| JWH133, CB2r agonist (3 mg/kg) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [86] |
| CB2r & Nicotine-Animal studies | ||||
|---|---|---|---|---|
| Genetic manipulation CB2r | ||||
| Genetic manipulation | Specie | Experimental paradigm | Results | References |
| CB2r deletion, knock-out (CB2-/-) | Swiss ICR mice | CPP | ↓ Conditioned place preference | [49] |
| ISA | ↓ Motivation for nicotine consumption | |||
| WS | ↓ Withdrawal signs | |||
| CB2r deletion, knock-out (CB2-/-) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [82] |
| CB2r deletion in dopaminergic neurons, conditional knock-out (DAT-Cnr2-/-) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [86] |
| Pharmacological manipulation of CB2r | ||||
| Doses | Specie | Experimental paradigm | Results | References |
| SR144528, CB2r agonist (3 mg/kg) | Wistar Rats | DT | No differences | [87] |
| WIN55,212-2, CB1r/CB2r agonist (1 mg/kg) + Rimonabant, CB1r agonist (1 mg/kg) or AM630, CB2r antagonist (5 mg/kg) | Long-Evans rats | ISA | ↑ WIN55,212-2-induced nicotine relapse, blocked by rimonabant but not by AM630 | [88] |
| AM1241, CB2r agonist (1–10 mg/kg) | Long-Evans rats | ISA | No differences | [89] |
| AM630, CB2r agonist (1.25–5 mg/kg) | Long-Evans rats | ISA | No differences | |
| AM630, CB2r antagonist (1 mg/kg) | Swiss ICR mice | CPP | ↓ Conditioned place preference | [49] |
| ISA | ↓ Motivation for nicotine consumption | |||
| WA | ↓ Withdrawal signs | |||
| SR144528, CB2r antagonist (3 mg/kg) O-1966, CB2r agonist (1–20 mg/kg) | C57BL/6J mice | CPP | ↓ Conditioned place preference | [82] |
| C57BL/6J mice | CPP | ↑ Conditioned place preference | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarrete, F.; García-Gutiérrez, M.S.; Gasparyan, A.; Navarro, D.; Manzanares, J. CB2 Receptor Involvement in the Treatment of Substance Use Disorders. Biomolecules 2021, 11, 1556. https://doi.org/10.3390/biom11111556
Navarrete F, García-Gutiérrez MS, Gasparyan A, Navarro D, Manzanares J. CB2 Receptor Involvement in the Treatment of Substance Use Disorders. Biomolecules. 2021; 11(11):1556. https://doi.org/10.3390/biom11111556
Chicago/Turabian StyleNavarrete, Francisco, María S. García-Gutiérrez, Ani Gasparyan, Daniela Navarro, and Jorge Manzanares. 2021. "CB2 Receptor Involvement in the Treatment of Substance Use Disorders" Biomolecules 11, no. 11: 1556. https://doi.org/10.3390/biom11111556
APA StyleNavarrete, F., García-Gutiérrez, M. S., Gasparyan, A., Navarro, D., & Manzanares, J. (2021). CB2 Receptor Involvement in the Treatment of Substance Use Disorders. Biomolecules, 11(11), 1556. https://doi.org/10.3390/biom11111556