
Pib2 as an Emerging Master Regulator of Yeast TORC1


Abstract
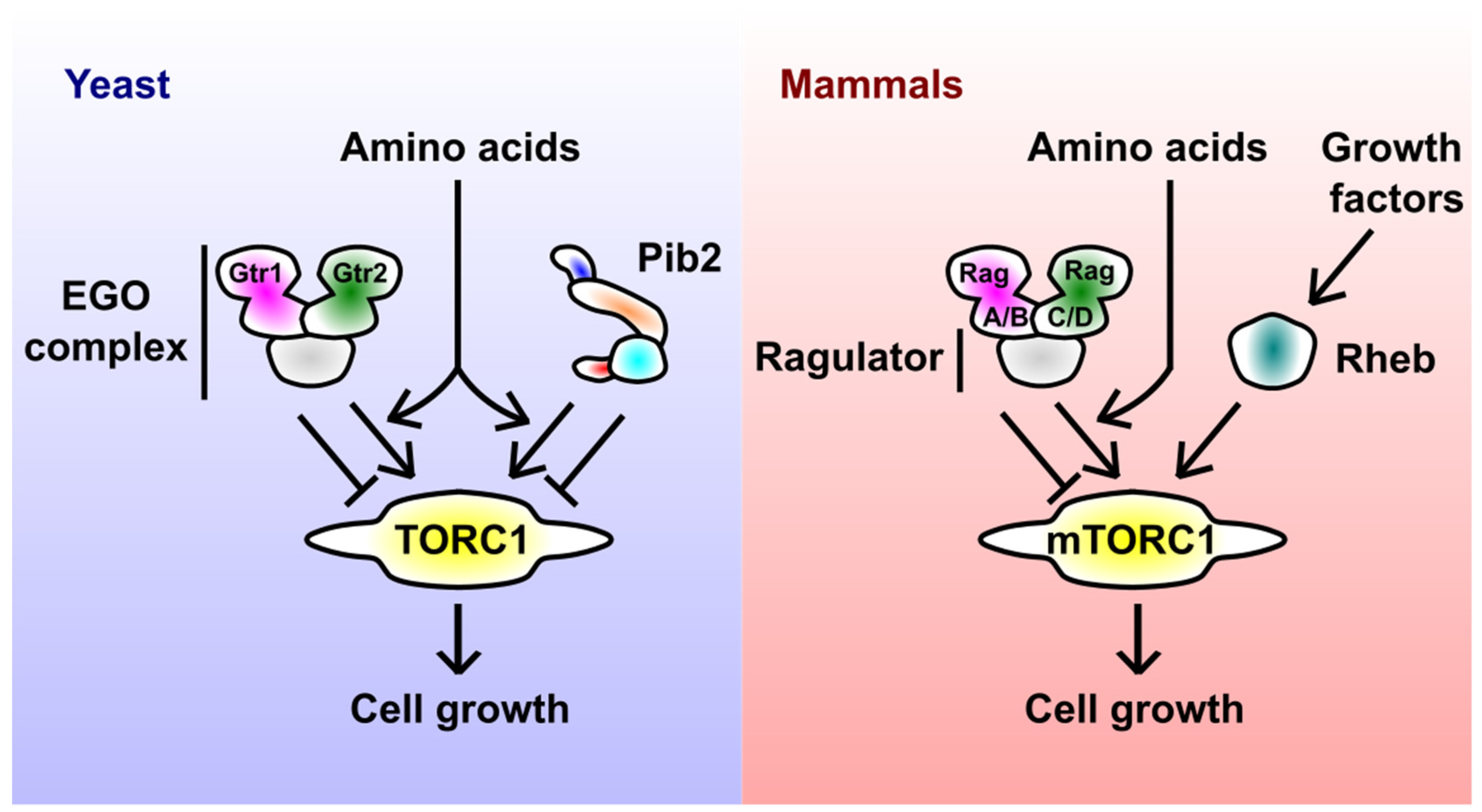
:1. Introduction: Major Players in TORC1 Regulation
2. Pib2 before 2015
3. Identification of Pib2 as a TORC1 Regulator
4. Pib2 in Amino Acid Response
4.1. Pib2 as a Glutamine Sensor
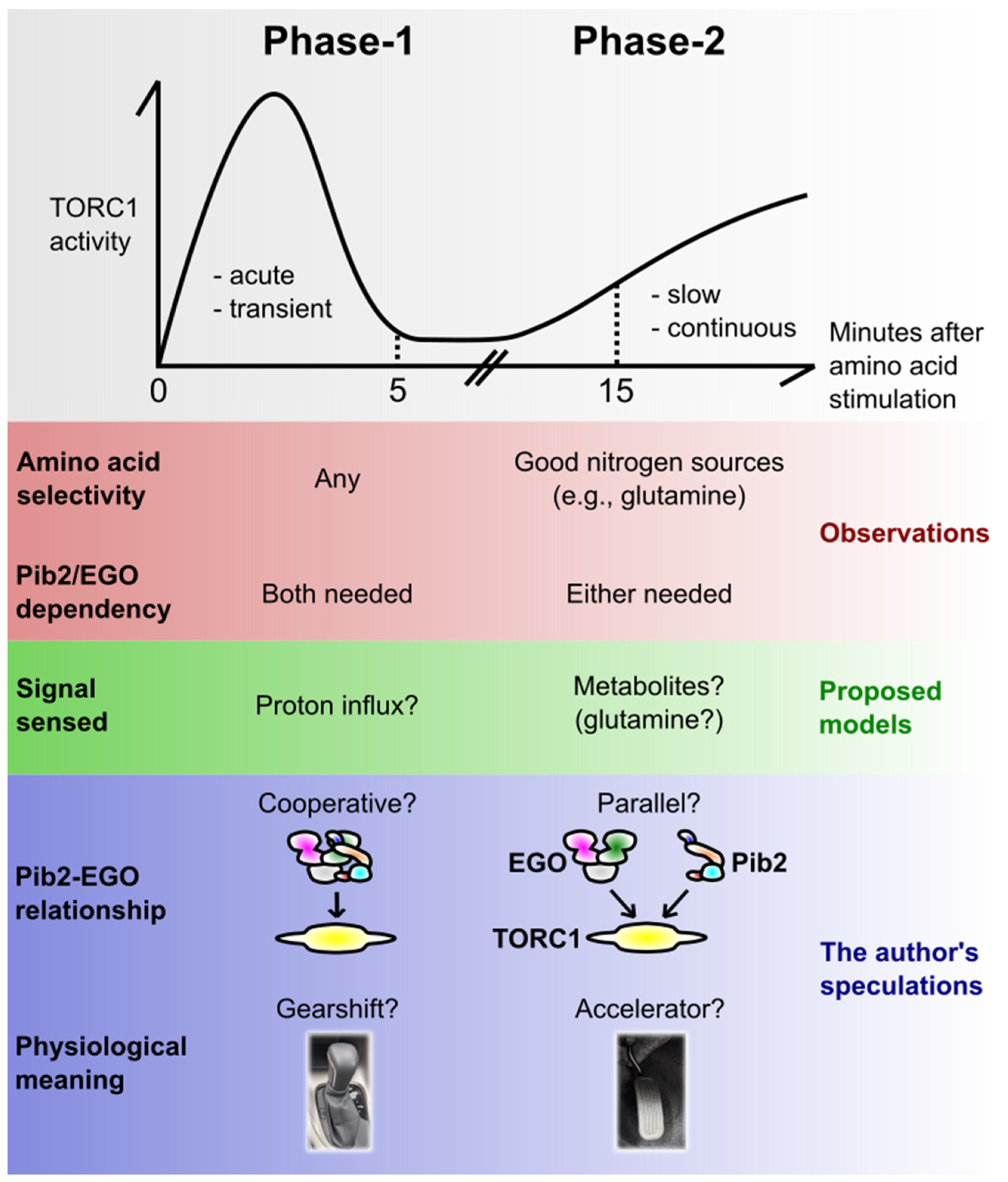
4.2. Dual-Phase Activation of TORC1
4.3. Pib2 and the EGO Complex in the Two Phases
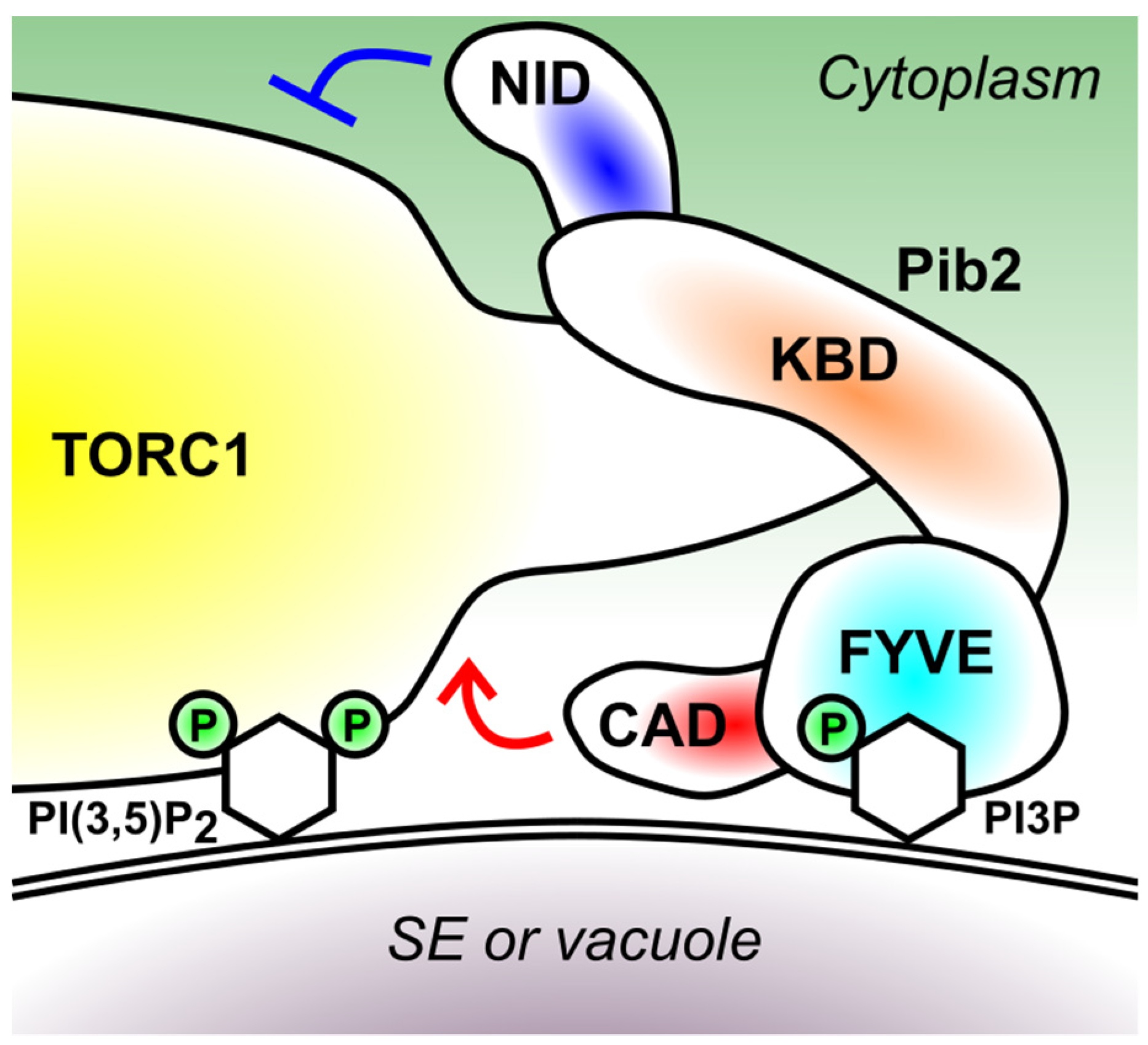
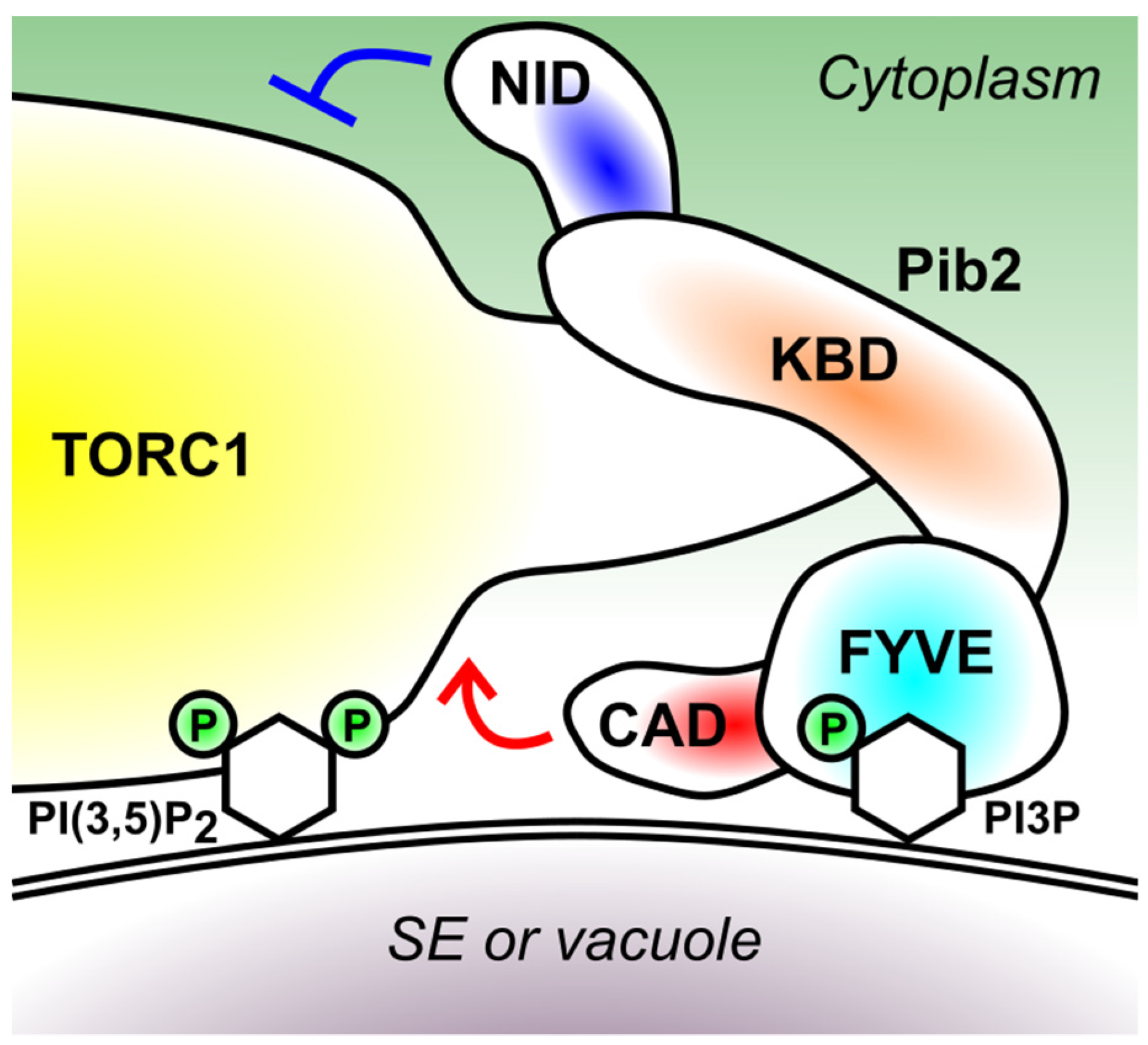
5. Domain Structure of Pib2
5.1. FYVE Domain (426–532 Amino Acid Residues)
5.2. Kog1-Binding Domain (KBD) (165–425 Residues)
5.3. C-Terminal TORC1-Activating Domain (CAD) (533–635 Residues)
5.4. N-Terminal Inhibitory Domain (NID) (1–164 Residues)
6. Regulation of Pib2
7. Pib2 in Other Organisms
8. Final Remarks: What’s Next?
Funding
Acknowledgments
Conflicts of Interest
References
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Shimobayashi, M.; Hall, M.N. Making new contacts: The mTOR network in metabolism and signalling crosstalk. Nat. Rev. Mol. Cell Biol. 2014, 15, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Tafur, L.; Kefauver, J.; Loewith, R. Structural insights into TOR signaling. Genes 2020, 11, 885. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Hall, M.N. Regulation of MTORC2 signaling. Genes 2020, 11, 1045. [Google Scholar] [CrossRef] [PubMed]
- Riggi, M.; Kusmider, B.; Loewith, R. The flipside of the TOR coin—TORC2 and plasma membrane homeostasis at a glance. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef]
- Dubouloz, F.; Deloche, O.; Wanke, V.; Cameroni, E.; De Virgilio, C. The TOR and EGO protein complexes orchestrate microautophagy in yeast. Mol. Cell 2005, 19, 15–26. [Google Scholar] [CrossRef]
- Sancak, Y.; Peterson, T.R.; Shaul, Y.D.; Lindquist, R.A.; Thoreen, C.C.; Bar-Peled, L.; Sabatini, D.M. The rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 2008, 320, 1496–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Goraksha-Hicks, P.; Li, L.; Neufeld, T.P.; Guan, K.L. Regulation of TORC1 by Rag GTPases in nutrient response. Nat. Cell Biol. 2008, 10, 935–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Loewith, R.; Jacinto, E.; Wullschleger, S.; Lorberg, A.; Crespo, J.L.; Bonenfant, D.; Oppliger, W.; Jenoe, P.; Hall, M.N. Two TOR complexes, only one of which is rapamycin sensitive, have distinct roles in cell growth control. Mol. Cell 2002, 10, 457–468. [Google Scholar] [CrossRef]
- Kim, A.; Cunningham, K.W. A LAPF/phafin1-like protein regulates TORC1 and lysosomal membrane permeabilization in response to endoplasmic reticulum membrane stress. Mol. Biol. Cell 2015, 26, 4631–4645. [Google Scholar] [CrossRef]
- Hatakeyama, R.; De Virgilio, C. Unsolved mysteries of Rag GTPase signaling in yeast. Small GTPases 2016, 7, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Condon, K.J.; Sabatini, D.M. Nutrient regulation of mTORC1 at a glance. J. Cell Sci. 2019, 132, jcs222570. [Google Scholar] [CrossRef]
- Panchaud, N.; Péli-Gulli, M.P.; De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Sci. Signal. 2013, 6, ra42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchaud, N.; Péli-Gulli, M.P.; De Virgilio, C. SEACing the GAP that nEGOCiates TORC1 activation: Evolutionary conservation of Rag GTPase regulation. Cell Cycle 2013, 12, 2948–2952. [Google Scholar] [CrossRef] [Green Version]
- Bar-Peled, L.; Chantranupong, L.; Cherniack, A.D.; Chen, W.W.; Ottina, K.A.; Grabiner, B.C.; Spear, E.D.; Carter, S.L.; Meyerson, M.; Sabatini, D.M. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science 2013, 340, 1100–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péli-Gulli, M.P.; Sardu, A.; Panchaud, N.; Raucci, S.; De Virgilio, C. Amino Acids Stimulate TORC1 through Lst4-Lst7, a GTPase-Activating Protein Complex for the Rag Family GTPase Gtr2. Cell Rep. 2015, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsun, Z.Y.; Bar-Peled, L.; Chantranupong, L.; Zoncu, R.; Wang, T.; Kim, C.; Spooner, E.; Sabatini, D.M. The folliculin tumor suppressor is a GAP for the RagC/D GTPases that signal amino acid levels to mTORC1. Mol. Cell 2013, 52, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, R.; Sardu, A.; Panchaud, N.; De Virgilio, C. The architecture of the Rag GTPase signaling network. Biomolecules 2017, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Chantranupong, L.; Wolfson, R.L.; Sabatini, D.M. Nutrient-sensing mechanisms across evolution. Cell 2015, 161, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Sancak, Y.; Bar-Peled, L.; Zoncu, R.; Markhard, A.L.; Nada, S.; Sabatini, D.M. Ragulator-rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 2010, 141, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Nakashima, A.; Guo, L.; Tamanoi, F. Specific activation of mTORC1 by Rheb G-protein in vitro involves enhanced recruitment of its substrate protein. J. Biol. Chem. 2009, 284, 12783–12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, Y.; Thoreen, C.C.; Peterson, T.R.; Lindquist, R.A.; Kang, S.A.; Spooner, E.; Carr, S.A.; Sabatini, D.M. PRAS40 is an insulin-regulated inhibitor of the mTORC1 protein Kinase. Mol. Cell 2007, 25, 903–915. [Google Scholar] [CrossRef]
- Yang, H.; Jiang, X.; Li, B.; Yang, H.J.; Miller, M.; Yang, A.; Dhar, A.; Pavletich, N.P. Mechanisms of mTORC1 activation by RHEB and inhibition by PRAS40. Nature 2017, 552, 368–373. [Google Scholar] [CrossRef]
- Aspuria, P.J.; Tamanoi, F. The Rheb family of GTP-binding proteins. Cell. Signal. 2004, 16, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Kira, S.; Tabata, K.; Shirahama-Noda, K.; Nozoe, A.; Yoshimori, T.; Noda, T. Reciprocal conversion of Gtr1 and Gtr2 nucleotidebinding states by Npr2-Npr3 inactivates TORC1 and induces autophagy. Autophagy 2014, 10, 1565–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demetriades, C.; Doumpas, N.; Teleman, A.A. Regulation of TORC1 in response to amino acid starvation via lysosomal recruitment of TSC2. Cell 2014, 156, 786–799. [Google Scholar] [CrossRef] [Green Version]
- Hughes Hallett, J.E.; Luo, X.; Capaldi, A.P. Snf1/AMPK promotes the formation of Kog1/raptor-bodies to increase the activation threshold of TORC1 in budding yeast. Elife 2015, 4, e09181. [Google Scholar] [CrossRef]
- Yuan, W.; Guo, S.; Gao, J.; Zhong, M.; Yan, G.; Wu, W.; Chao, Y.; Jiang, Y. General control nonderepressible 2 (GCN2) kinase inhibits target of rapamycin complex 1 in response to amino acid starvation in saccharomyces cerevisiae. J. Biol. Chem. 2017, 292, 2660–2669. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, G.; Zhang, Y.; Dayhoff-Brannigan, M.; Diny, N.L.; Zhao, M.; He, G.; Sing, C.N.; Metz, K.A.; Stolp, Z.D.; et al. Whi2 is a conserved negative regulator of TORC1 in response to low amino acids. PLoS Genet. 2018, 14, e1007592. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.S.; Soto Diaz, S.; Van Vooren, P.; Godard, P.; Marini, A.M.; Boeckstaens, M. Pib2-dependent feedback control of the TORC1 Signaling network by the Npr1 Kinase. iScience 2019, 20, 415–433. [Google Scholar] [CrossRef] [Green Version]
- Magasanik, B.; Kaiser, C.A. Nitrogen regulation in Saccharomyces cerevisiae. Gene 2002, 290, 1–18. [Google Scholar] [CrossRef]
- Boeckstaens, M.; Llinares, E.; Van Vooren, P.; Marini, A.M. The TORC1 effector kinase Npr1 fine tunes the inherent activity of the Mep2 ammonium transport protein. Nat. Commun. 2014, 5, 3101. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.H.; Hatakeyama, R.; Kimmig, P.; Arter, M.; Peter, M.; Matos, J.; De Virgilio, C.; Kornmann, B.T. Functional mapping of yeast genomes by saturated transposition. Elife 2017, 6, e2357. [Google Scholar] [CrossRef]
- Tanigawa, M.; Maeda, T. An in vitro TORC1 kinase assay that recapitulates the Gtr-independent glutamine-responsive TORC1 activation mechanism on yeast vacuoles. Mol. Cell. Biol. 2017, 37, e00075-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanigawa, M.; Yamamoto, K.; Nagatoishi, S.; Noda, N.N.; Tsumoto, K.; Maeda, T. A glutamine sensor that directly activates TORC1. Commun. Biol. 2021, 4, 1–11. [Google Scholar] [CrossRef]
- Ukai, H.; Araki, Y.; Kira, S.; Oikawa, Y.; May, A.I.; Noda, T. Gtr/Ego-independent TORC1 activation is achieved through a glutamine-sensitive interaction with Pib2 on the vacuolar membrane. PLoS Genet. 2018, 14, e1007334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlakhanova, N.V.; Mihalevic, M.J.; Bernstein, K.A.; Ford, M.G.J. Pib2 and the EGO complex are both required for activation of TORC1. J. Cell Sci. 2017, 130, 3878–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, A.; Wallace, R.L.; Wellington, R.; Luo, X.; Capaldi, A.P. Multilayered regulation of TORC1-body formation in budding yeast. Mol. Biol. Cell 2019, 30, 400–410. [Google Scholar] [CrossRef]
- Shin, M.E.; Ogburn, K.D.; Varban, O.A.; Gilbert, P.M.; Burd, C.G. FYVE domain targets Pib1p ubiquitin ligase to endosome and vacuolar membranes. J. Biol. Chem. 2001, 276, 41388–41393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, C.G.; Emr, S.D. Phosphatidylinositol (3)-phosphate signaling mediated by specific binding to RING FYVE domains. Mol. Cell 1998, 2, 157–162. [Google Scholar] [CrossRef]
- Lemmon, M.A. Phosphoinositide recognition domains. Traffic 2003, 4, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lo, W.T.; Haucke, V. Phosphoinositide switches in endocytosis and in the endolysosomal system. Curr. Opin. Cell Biol. 2019, 59, 50–57. [Google Scholar] [CrossRef]
- Sturgill, T.W.; Cohen, A.; Diefenbacher, M.; Trautwein, M.; Martin, D.E.; Hall, M.N. TOR1 and TOR2 have distinct locations in live cells. Eukaryot. Cell 2008, 7, 1819–1830. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, R.; Péli-Gulli, M.P.; Hu, Z.; Jaquenoud, M.; Garcia Osuna, G.M.; Sardu, A.; Dengjel, J.; De Virgilio, C. Spatially distinct pools of TORC1 balance protein Homeostasis. Mol. Cell 2019, 73, 325–338.e8. [Google Scholar] [CrossRef] [Green Version]
- Macgurn, J.A.; Hsu, P.-C.; Smolka, M.B.; Emr, S.D. TORC1 regulates endocytosis via Npr1-mediated phosphoinhibition of a ubiquitin ligase adaptor. Cell 2011, 147, 1104–1117. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Li, N.; Chen, T.; Han, Y.; Li, C.; Wang, Y.; He, W.; Zhang, L.; Wan, T.; Cao, X. The lysosome-associated apoptosis-inducing protein containing the pleckstrin homology (PH) and FYVE domains (LAPF), representative of a novel family of PH and FYVE domain-containing proteins, induces caspase-independent apoptosis via the lysosomal-mitocho. J. Biol. Chem. 2005, 280, 40985–40995. [Google Scholar] [CrossRef] [Green Version]
- Tarassov, K.; Messier, V.; Landry, C.R.; Radinovic, S.; Serna Molina, M.M.; Shames, I.; Malitskaya, Y.; Vogel, J.; Bussey, H.; Michnick, S.W. An in vivo map of the yeast protein interactome. Science 2008, 320, 1465–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfson, R.L.; Sabatini, D.M. The dawn of the age of amino acid sensors for the mTORC1 pathway. Cell Metab. 2017, 26, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ou, Y.; Luo, R.; Wang, J.; Wang, D.; Guan, J.; Li, Y.; Xia, P.; Chen, P.R.; Liu, Y. SAR1B senses leucine levels to regulate mTORC1 signalling. Nature 2021, 596, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Stracka, D.; Jozefczuk, S.; Rudroff, F.; Sauer, U.; Hall, M.N. Nitrogen source activates TOR (Target of Rapamycin) complex 1 via glutamine and independently of Gtr/Rag proteins. J. Biol. Chem. 2014, 289, 25010–25020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliba, E.; Evangelinos, M.; Gournas, C.; Corrillon, F.; Georis, I.; André, B. The yeast H+ -ATPase Pma1 promotes Rag/Gtr-dependent TORC1 activation in response to H+-coupled nutrient uptake. Elife 2018, 7, e31981. [Google Scholar] [CrossRef] [PubMed]
- Saliba, E.; Primo, C.; Guarini, N.; André, B. A plant plasma-membrane H+-ATPase promotes yeast TORC1 activation via its carboxy-terminal tail. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Hatakeyama, R.; De Virgilio, C. A spatially and functionally distinct pool of TORC1 defines signaling endosomes in yeast. Autophagy 2019, 15, 915–916. [Google Scholar] [CrossRef]
- Chen, Z.; Malia, P.C.; Hatakeyama, R.; Nicastro, R.; Hu, Z.; Péli-Gulli, M.-P.; Gao, J.; Nishimura, T.; Eskes, E.; Stefan, C.J.; et al. TORC1 determines Fab1 lipid kinase function at signaling endosomes and vacuoles. Curr. Biol. 2020, 31, 297–309. [Google Scholar] [CrossRef]
- Mccartney, A.J.; Zhang, Y.; Weisman, L.S. Phosphatidylinositol 3,5-bisphosphate: Low abundance, high significance. BioEssays 2014, 36, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; Mao, K.; Jin, Y.; Tevzadze, G.; Kauffman, E.J.; Park, S.; Bridges, D.; Loewith, R.; Saltiel, A.R.; Klionsky, D.J.; et al. Roles for PI (3,5) P2in nutrient sensing through TORC1. Mol. Biol. Cell 2014, 25, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.G.; Cullen, P.J. Coincidence detection in phosphoinositide signaling. Trends Cell Biol. 2005, 15, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Prouteau, M.; Desfosses, A.; Sieben, C.; Bourgoint, C.; Mozaffari, N.L.; Demurtas, D.; Mitra, A.K.; Guichard, P.; Manley, S.; Loewith, R. TORC1 organized in inhibited domains (TOROIDs) regulate TORC1 activity. Nature 2017, 550, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Rogala, K.B.; Gu, X.; Kedir, J.F.; Abu-Remaileh, M.; Bianchi1, L.F.; Bottino1, A.M.S.; Dueholm1, R.; Niehaus1, A.; Overwijn1, D.; Priso Fils1, A.C.; et al. Structural basis for the docking of mTORC1 on the lysosomal surface. Science 2019, 366, 468–475. [Google Scholar] [CrossRef]
- Eltschinger, S.; Loewith, R. TOR Complexes and the maintenance of cellular homeostasis. Trends Cell Biol. 2016, 26, 148–159. [Google Scholar] [CrossRef]
- Lin, W.-J.; Yang, C.-Y.; Li, L.-L.; Yi, Y.-H.; Chen, K.-W.; Lin, Y.-C.; Liu, C.-C.; Lin, C.-H. Lysosomal targeting of phafin1 mediated by Rab7 induces autophagosome formation. Biochem. Biophys. Res. Commun. 2012, 417, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Noda, T.; Ohsumi, Y. Tor, a phosphatidylinositol kinase homologue, controls autophagy in yeast. J. Biol. Chem. 1998, 273, 3963–3966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauffer, B.; Powers, T. Target of rapamycin signaling mediates vacuolar fragmentation. Curr. Genet. 2017, 63, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.; Yang, Q.; Wang, H.; Melick, C.H.; Navlani, R.; Frank, A.R.; Jewell, J.L. Glutamine and asparagine activate mTORC1 independently of Rag GTPases. J. Biol. Chem. 2020, 295, 2890–2899. [Google Scholar] [CrossRef] [Green Version]
- Urano, J.; Comiso, M.J.; Guo, L.; Aspuria, P.J.; Deniskin, R.; Tabancay, A.P.; Kato-Stankiewicz, J.; Tamanoi, F. Identification of novel single amino acid changes that result in hyperactivation of the unique GTPase, Rheb, in fission yeast. Mol. Microbiol. 2005, 58, 1074–1086. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.M.; Canal, M.V.; Pereyra, C.M.; Welchen, E.; Martínez-Noël, G.M.A.; Estevez, J.M. The tip of the iceberg: Emerging roles of TORC1, and its regulatory functions in plant cells. J. Exp. Bot. 2021, 72, 4085–4101. [Google Scholar] [CrossRef]
- Cao, P.; Kim, S.-J.; Xing, A.; Schenck, C.A.; Liu, L.; Jiang, N.; Wang, J.; Last, R.L.; Brandizzi, F. Homeostasis of branched-chain amino acids is critical for the activity of TOR signaling in Arabidopsis. Elife 2019, 8, 1–24. [Google Scholar] [CrossRef]
- Ingargiola, C.; Duarte, G.T.; Robaglia, C.; Leprince, A.S.; Meyer, C. The plant target of rapamycin: A conductor of nutrition and metabolism in photosynthetic organisms. Genes 2020, 11, 1285. [Google Scholar] [CrossRef]
- Tatebe, H.; Shiozaki, K. Evolutionary conservation of the components in the TOR signaling pathways. Biomolecules 2017, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Observation | Supported Model | Yeast Strain Background | Culture Media/ TORC1 Stimuli | TORC1 Readout | References |
|---|---|---|---|---|---|
| pib2∆-EGO∆ synthetic lethality | Parallel | BY4741/4742 (auxotroph) | YPD | Viability (rescued by hyperactive TORC1) | [12] |
| Pib2-EGO redundancy in the basal TORC1 activity | Parallel | BY4741 (auxotroph) | SD with casamino acid | Sch9 phosphorylation (C-terminus band-shift) | [38] |
| Partial defects in TORC1 activation (>10 min) in pib2∆ and EGO∆ | Parallel | Σ1278b (prototroph) [32]; S288C (prototroph) [36] | Minimal proline medium +glutamine or ammonium stimulation [32]; SD-N +glutamine stimulation [36] | Rps6 phosphorylation (phospho-specific antibody) or Npr1 phosphorylation (band-shift) [32]; Sch9 phosphorylation (phospho-specific antibody) [36] | [32,36] |
| Distinct Pib2-TORC1 and EGO-TORC1 supercomplexes (co-immunoprecipitation) | Parallel | BY4741 (auxotroph) | YPD | N/A | [38] |
| Pib2-EGO interaction (yeast two-hybrid) | Cooperative | N/A | N/A | N/A | [12,49] |
| Strict requirement of both Pib2 and EGO for acute and transient TORC1 activation (<5 min) | Cooperative | BY4741/4742 (prototroph) [35]; S288C (prototroph) [36]; W303A (auxotroph) [39] | Minimal proline medium +glutamine stimulation [35]; SD-N +glutamine or arginine stimulation [36]; SD-N +glutamine or leucine stimulation [39] | Sch9 phosphorylation (phospho-specific antibody) [35,36]; Rps6 phosphorylation (phospho-specific antibody [39] | [35,36,39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatakeyama, R. Pib2 as an Emerging Master Regulator of Yeast TORC1. Biomolecules 2021, 11, 1489. https://doi.org/10.3390/biom11101489
Hatakeyama R. Pib2 as an Emerging Master Regulator of Yeast TORC1. Biomolecules. 2021; 11(10):1489. https://doi.org/10.3390/biom11101489
Chicago/Turabian StyleHatakeyama, Riko. 2021. "Pib2 as an Emerging Master Regulator of Yeast TORC1" Biomolecules 11, no. 10: 1489. https://doi.org/10.3390/biom11101489
APA StyleHatakeyama, R. (2021). Pib2 as an Emerging Master Regulator of Yeast TORC1. Biomolecules, 11(10), 1489. https://doi.org/10.3390/biom11101489