Peucedanum ostruthium Inhibits E-Selectin and VCAM-1 Expression in Endothelial Cells through Interference with NF-κB Signaling

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Herbal/Fungal Extracts
2.2. Cell Culture
2.3. Antibodies and Reagents
2.4. Cytotoxicity Assays
2.5. Real-Time PCR
2.6. Cell ELISA
2.7. Western Blotting
2.8. Transfection and Reporter Gene Assays
2.9. Immunofluorescence Microscopy
2.10. Ultra Performance Liquid Chromatography
2.11. Statistical Significance Calculations
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mayer, H.; Bilban, M.; Kurtev, V.; Gruber, F.; Wagner, O.; Binder, B.R.; De Martin, R. Deciphering regulatory patterns of inflammatory gene expression from interleukin-1-stimulated human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1192–1198. [Google Scholar] [CrossRef]
- De Martin, R.; Hoeth, M.; Hofer-Warbinek, R.; Schmid, J.A. The transcription factor NF-kappa B and the regulation of vascular cell function. Arterioscler. Thromb. Vasc. Biol. 2000, 20, e83–e88. [Google Scholar]
- Fiordelisi, A.; Iaccarino, G.; Morisco, C.; Coscioni, E.; Sorriento, D. NFkappaB is a key player in the crosstalk between inflammation and cardiovascular diseases. Int. J. Mol. Sci. 2019, 20, 1599. [Google Scholar] [CrossRef]
- Renner, F.; Schmitz, M.L. Autoregulatory feedback loops terminating the NF-kappaB response. Trends Biochem. Sci. 2009, 34, 128–135. [Google Scholar] [CrossRef]
- Winsauer, G.; de Martin, R. Resolution of inflammation: Intracellular feedback loops in the endothelium. Thromb. Haemost. 2007, 97. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Kanatsu, J.; Toh, M.; Naka, A.; Kondo, K.; Iida, K. The Dietary Isoflavone Daidzein Reduces Expression of Pro-Inflammatory Genes through PPARalpha/gamma and JNK Pathways in Adipocyte and Macrophage Co-Cultures. PLoS ONE 2016, 11, e0149676. [Google Scholar] [CrossRef]
- Schmitz, M.L.; Baeuerle, P.A. The p65 subunit is responsible for the strong transcription activating potential of NF-kappa B. EMBO J. 1991, 10, 3805–3817. [Google Scholar] [CrossRef]
- Zhong, L.; Simard, M.J.; Huot, J. Endothelial microRNAs regulating the NF-kappaB pathway and cell adhesion molecules during inflammation. FASEB J. 2018, 32, 4070–4084. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Gerlach, S.; Saukel, J.; Kubelka, W. Pflanzen in der österreichischen Volksmedizin. Die “VOLKSMED-Datenbank”. Sci. Pharm. 2006, 74, S36. [Google Scholar]
- Kratz, J.M.; Mair, C.E.; Oettl, S.K.; Saxena, P.; Scheel, O.; Schuster, D.; Hering, S.; Rollinger, J.M. hERG channel blocking ipecac alkaloids identified by combined in silico—In vitro screening. Planta Med. 2016, 82, 1009–1015. [Google Scholar] [CrossRef]
- Camp, D.; Davis, R.A.; Campitelli, M.; Ebdon, J.; Quinn, R.J. Drug-like properties: Guiding principles for the design of natural product libraries. J. Nat. Prod. 2012, 75, 72–81. [Google Scholar] [CrossRef]
- Tu, Y.; Jeffries, C.; Ruan, H.; Nelson, C.; Smithson, D.; Shelat, A.A.; Brown, K.M.; Li, X.C.; Hester, J.P.; Smillie, T.; et al. Automated high-throughput system to fractionate plant natural products for drug discovery. J. Nat. Prod. 2010. [Google Scholar] [CrossRef]
- Mair, C.E.; Grienke, U.; Wilhelm, A.; Urban, E.; Zehl, M.; Schmidtke, M.; Rollinger, J.M. Anti-influenza triterpene saponins from the bark of Burkea africana. J. Nat. Prod. 2018, 81, 515–523. [Google Scholar] [CrossRef]
- Grienke, U.; Mair, C.E.; Kirchmair, J.; Schmidtke, M.; Rollinger, J.M. Discovery of bioactive natural products for the treatment of acute respiratory infections—An integrated approach *. Planta Med. 2018, 84, 684–695. [Google Scholar] [CrossRef]
- Zwirchmayr, J.; Kirchweger, B.; Lehner, T.; Tahir, A.; Pretsch, D.; Rollinger, J.M. A robust and miniaturized screening platform to study natural products affecting metabolism and survival in Caenorhabditis elegans. Sci. Rep. 2020. [Google Scholar] [CrossRef]
- Grienke, U.; Zwirchmayr, J.; Peintner, U.; Urban, E.; Zehl, M.; Schmidtke, M.; Rollinger, J.M. Lanostane triterpenes from Gloeophyllum odoratum and their anti-influenza effects. Planta Med. 2019, 85, 195–202. [Google Scholar] [CrossRef]
- Zwirchmayr, J.; Grienke, U.; Hummelbrunner, S.; Seigner, J.; de Martin, R.; Dirsch, V.M.; Rollinger, J.M. A biochemometric approach for the identification of in vitro anti-inflammatory constituents in masterwort. Biomolecules 2020, 28, 679. [Google Scholar] [CrossRef]
- Hoeth, M.; Niederleithner, H.; Hofer-Warbinek, R.; Bilban, M.; Mayer, H.; Resch, U.; Lemberger, C.; Wagner, O.; Hofer, E.; Petzelbauer, P.; et al. The transcription factor SOX18 regulates the expression of matrix metalloproteinase 7 and guidance molecules in human endothelial cells. PLoS ONE 2012, 7, e30982. [Google Scholar] [CrossRef]
- Seigner, J.; Junker-Samek, M.; Plaza, A.; D’Urso, G.; Masullo, M.; Piacente, S.; Holper-Schichl, Y.M.; de Martin, R. A Symphytum officinale Root Extract Exerts Anti-inflammatory Properties by Affecting Two Distinct Steps of NF-kappaB Signaling. Front. Pharmacol. 2019, 10, 289. [Google Scholar] [CrossRef]
- Seigner, J.; Basilio, J.; Resch, U.; de Martin, R. CD40L and TNF both activate the classical NF-κB pathway, which is not required for the CD40L induced alternative pathway in endothelial cells. Biochem. Biophys. Res. Commun. 2017, 495, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Grienke, U.; Zoll, M.; Peintner, U.; Rollinger, J.M. European medicinal polypores--a modern view on traditional uses. J. Ethnopharmacol. 2014, 154, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziem, S.H.; El-Nekeety, A.A.; Barakat, I.A.; Mohamed, M.I.; Abdel-Wahhab, M.A. Aquilegia vulgaris extract protects against the oxidative stress and the mutagenic effects of cadmium in Balb/c mice. Exp. Toxicol. Pathol. 2011, 63, 337–344. [Google Scholar] [CrossRef]
- Jodynis-Liebert, J.; Adamska, T.; Ewertowska, M.; Bylka, W.; Matławska, I. Aquilegia vulgaris extract attenuates carbon tetrachloride-induced liver fibrosis in rats. Exp. Toxicol. Pathol. 2009, 61, 443–451. [Google Scholar] [CrossRef]
- Hassan, A.M.; Mohamed, S.R.; El-Nekeety, A.A.; Hassan, N.S.; Abdel-Wahhab, M.A. Aquilegia vulgaris L. extract counteracts oxidative stress and cytotoxicity of fumonisin in rats. Toxicon 2010, 56, 8–18. [Google Scholar] [CrossRef]
- Adamska, T.; Mlynarczyk, W.; Jodynis-Liebert, J.; Bylka, W.; Matlawska, I. Hepatoprotective effect of the extract and isocytisoside from Aquilegia vulgaris. Phytother. Res. 2003, 17, 691–696. [Google Scholar] [CrossRef]
- Subapriya, R.; Nagini, S. Medicinal properties of Neem leaves: A review. Currrent Med. Chem. Anti Cancer Agents 2005, 5, 149–156. [Google Scholar] [CrossRef]
- Xiang, X.; Wu, L.; Mao, L.; Liu, Y. Antioxidative and antiapoptotic neuroprotective effects of Azadirachta indica in Parkinsoninduced functional damage. Mol. Med. Rep. 2018, 17, 7959–7965. [Google Scholar] [CrossRef]
- Patel, S.M.; Nagulapalli Venkata, K.C.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Hao, F.; Kumar, S.; Yadav, N.; Chandra, D. Neem components as potential agents for cancer prevention and treatment. Biochim. Biophys. Acta Rev. Cancer 2014, 1846, 247–257. [Google Scholar] [CrossRef]
- Joy Sinha, D.; Nandha, K.D.S.; Jaiswal, N.; Vasudeva, A.; Prabha Tyagi, S.; Pratap Singh, U. Antibacterial Effect of Azadirachta indica (Neem) or Curcuma longa (Turmeric) against Enterococcus faecalis Compared with That of 5% Sodium Hypochlorite or 2% Chlorhexidine in vitro. Bull. Tokyo Dent. Coll 2017, 58, 103–109. [Google Scholar] [CrossRef]
- Batista, F.L.A.; Lima, L.M.G.; Abrante, I.A.; de Araujo, J.I.F.; Batista, F.L.A.; Abrante, I.A.; Magalhaes, E.A.; de Lima, D.R.; Lima, M.; do Prado, B.S.; et al. Antinociceptive activity of ethanolic extract of Azadirachta indica A. Juss (Neem, Meliaceae) fruit through opioid, glutamatergic and acid-sensitive ion pathways in adult zebrafish (Danio rerio). Biomed. Pharmacother. 2018, 108, 408–416. [Google Scholar] [CrossRef]
- Kushwaha, P.; Khedgikar, V.; Haldar, S.; Gautam, J.; Mulani, F.A.; Thulasiram, H.V.; Trivedi, R. Azadirachta indica triterpenoids promote osteoblast differentiation and mineralization in vitro and in vivo. Bioorg. Med. Chem. Lett. 2016, 26, 3719–3724. [Google Scholar] [CrossRef]
- Mathisen, E.; Diallo, D.; Andersen, O.M.; Malterud, K.E. Antioxidants from the bark of Burkea africana, an African medicinal plant. Phytother. Res. 2002, 16, 148–153. [Google Scholar] [CrossRef]
- Peng, J.; Hu, T.; Li, J.; Du, J.; Zhu, K.; Cheng, B.; Li, K. Shepherd’s Purse Polyphenols Exert Its Anti-Inflammatory and Antioxidative Effects Associated with Suppressing MAPK and NF-kappaB Pathways and Heme Oxygenase-1 Activation. Oxid. Med. Cell. Longev. 2019, 2019, 7202695. [Google Scholar] [CrossRef]
- Cha, J.M.; Kim, D.H.; Lee, T.H.; Subedi, L.; Kim, S.Y.; Lee, K.R. Phytochemical constituents of Capsella bursa-pastoris and their anti-inflammatory activity. Nat. Prod. Sci. 2018, 24, 132–138. [Google Scholar] [CrossRef]
- Vogl, S.; Picker, P.; Mihaly-Bison, J.; Fakhrudin, N.; Atanasov, A.G.; Heiss, E.H.; Wawrosch, C.; Reznicek, G.; Dirsch, V.M.; Saukel, J.; et al. Ethnopharmacological in vitro studies on Austria’s folk medicine-an unexplored lore in vitro anti-inflammatory activities of 71 Austrian traditional herbal drugs. J. Ethnopharmacol. 2013, 149, 750–771. [Google Scholar] [CrossRef]
- Ghalandari, S.; Kariman, N.; Sheikhan, Z.; Mojab, F.; Mirzaei, M.; Shahrahmani, H. Effect of hydroalcoholic extract of Capsella bursa pastoris on early postpartum hemorrhage: A clinical trial study. J. Altern. Complement. Med. 2017, 23, 794–799. [Google Scholar] [CrossRef]
- Nadkarni, K.M. Indian Materia Medica: With Ayurvedic, Unani-Tibbi, Siddha, Allopathic, Homeopathic, Naturopathic & Home Remedies, Appendices & Indexes; Popular Prakashan Private Ltd.: Mumbai, India, 1996; Volume 1. [Google Scholar]
- Gilani, A.H.; Jabeen, Q.; Khan, A.; Shah, A.J. Gut modulatory, blood pressure lowering, diuretic and sedative activities of cardamom. J. Ethnopharmacol. 2008, 115, 463–472. [Google Scholar] [CrossRef]
- Souissi, M.; Azelmat, J.; Chaieb, K.; Grenier, D. Antibacterial and anti-inflammatory activities of cardamom (Elettaria cardamomum) extracts: Potential therapeutic benefits for periodontal infections. Anaerobe 2019, 61, 102089. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Carr, R.I. In vitro investigation of the potential immunomodulatory and anti-cancer activities of black pepper (Piper nigrum) and cardamom (Elettaria cardamomum). J. Med. Food 2010, 13, 371–381. [Google Scholar] [CrossRef]
- Das, I.; Acharya, A.; Berry, D.L.; Sen, S.; Williams, E.; Permaul, E.; Sengupta, A.; Bhattacharya, S.; Saha, T. Antioxidative effects of the spice cardamom against non-melanoma skin cancer by modulating nuclear factor erythroid-2-related factor 2 and NF-kappaB signalling pathways. Br. J. Nutr. 2012, 108, 984–997. [Google Scholar] [CrossRef]
- Qiblawi, S.; Al-Hazimi, A.; Al-Mogbel, M.; Hossain, A.; Bagchi, D. Chemopreventive effects of cardamom (Elettaria cardamomum L.) on chemically induced skin carcinogenesis in Swiss albino mice. J. Med. Food 2012, 15, 576–580. [Google Scholar] [CrossRef]
- Mutlu-Ingok, A.; Karbancioglu-Guler, F. Cardamom, cumin, and dill weed essential oils: Chemical compositions, antimicrobial activities, and mechanisms of action against campylobacter spp. Molecules 2017, 22, 1191. [Google Scholar] [CrossRef]
- Kazemi, S.; Yaghooblou, F.; Siassi, F.; Rahimi Foroushani, A.; Ghavipour, M.; Koohdani, F.; Sotoudeh, G. Cardamom supplementation improves inflammatory and oxidative stress biomarkers in hyperlipidemic, overweight, and obese pre-diabetic women: A randomized double-blind clinical trial. J. Sci. Food Agric. 2017, 97, 5296–5301. [Google Scholar] [CrossRef]
- Stoss, M.; Michels, C.; Peter, E.; Beutke, R.; Gorter, R.W. Prospective cohort trial of Euphrasia single-dose eye drops in conjunctivitis. J. Altern. Complement. Med. 2000, 6, 499–508. [Google Scholar] [CrossRef]
- Liu, Y.; Hwang, E.; Ngo, H.T.T.; Perumalsamy, H.; Kim, Y.J.; Li, L.; Yi, T.-H. Protective effects of Euphrasia officinalis Extract against Ultraviolet B-Induced Photoaging in Normal Human Dermal Fibroblasts. Int. J. Mol. Sci. 2018, 19, 3327. [Google Scholar] [CrossRef]
- Paduch, R.; Woźniak, A.; Niedziela, P.; Rejdak, R. Assessment of eyebright (Euphrasia Officinalis L.) extract activity in relation to human corneal cells using in vitro tests. Balk. Med. J. 2014, 31, 29–36. [Google Scholar] [CrossRef]
- Bigagli, E.; Cinci, L.; D’Ambrosio, M.; Luceri, C. Pharmacological activities of an eye drop containing Matricaria chamomilla and Euphrasia officinalis extracts in UVB-induced oxidative stress and inflammation of human corneal cells. J. Photochem. Photobiol. B Biol. 2017, 173, 618–625. [Google Scholar] [CrossRef]
- Hao, L.; Sheng, Z.; Lu, J.; Tao, R.; Jia, S. Characterization and antioxidant activities of extracellular and intracellular polysaccharides from Fomitopsis pinicola. Carbohydr. Polym. 2016, 141, 54–59. [Google Scholar] [CrossRef]
- Wu, H.-T.; Lu, F.-H.; Su, Y.-C.; Ou, H.-Y.; Hung, H.-C.; Wu, J.-S.; Yang, Y.-C.; Chang, C.-J. In vivo and in vitro anti-tumor effects of fungal extracts. Molecules 2014, 19, 2546–2556. [Google Scholar] [CrossRef]
- Bhattarai, G.; Lee, Y.-H.; Lee, N.-H.; Lee, I.-K.; Yun, B.-S.; Hwang, P.-H.; Yi, H.-K. Fomitoside-K from Fomitopsis nigra Induces Apoptosis of Human Oral Squamous Cell Carcinomas (YD-10B) via Mitochondrial Signaling Pathway. Biol. Pharm. Bull. 2012, 35, 1711–1719. [Google Scholar] [CrossRef][Green Version]
- Choi, D.; Park, S.-S.; Ding, J.-L.; Cha, W.-S. Effects of Fomitopsis pinicola extracts on antioxidant and antitumor activities. Biotechnol. Bioprocess. Eng. 2007, 12, 516. [Google Scholar] [CrossRef]
- Hwang, C.H.; Jaki, B.U.; Klein, L.L.; Lankin, D.C.; McAlpine, J.B.; Napolitano, J.G.; Fryling, N.A.; Franzblau, S.G.; Cho, S.H.; Stamets, P.E.; et al. Chlorinated Coumarins from the Polypore Mushroom Fomitopsis officinalis and Their Activity against Mycobacterium tuberculosis. J. Nat. Prod. 2013, 76, 1916–1922. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Inoue, M.; Matsumoto, Y.; Sakakibara, C.; Miyataka, H.; Matsumoto, H.; Arihara, S. Lanostane Triterpenoids and Triterpene Glycosides from the Fruit Body of Fomitopsis pinicola and Their Inhibitory Activity against COX-1 and COX-2. J. Nat. Prod. 2005, 68, 69–73. [Google Scholar] [CrossRef]
- Tai, S.-H.; Kuo, P.-C.; Hung, C.-C.; Lin, Y.-H.; Hwang, T.-L.; Lam, S.H.; Kuo, D.-H.; Wu, J.-B.; Hung, H.-Y.; Wu, T.-S. Bioassay-guided purification of sesquiterpenoids from the fruiting bodies of Fomitopsis pinicola and their anti-inflammatory activity. RSC Adv. 2019, 9, 34184–34195. [Google Scholar] [CrossRef]
- Zhao, L.; Xiang, K.-L.; Liu, R.-X.; Xie, Z.-P.; Zhang, S.-M.; Dai, S.-J. Anti-inflammatory and anti-viral labdane diterpenoids from the fruits of Forsythia suspensa. Bioorg. Chem. 2020, 96, 103651. [Google Scholar] [CrossRef]
- Zhao, L.; Yan, X.; Shi, J.; Ren, F.; Liu, L.; Sun, S.; Shan, B. Ethanol extract of Forsythia suspensa root induces apoptosis of esophageal carcinoma cells via the mitochondrial apoptotic pathway. Mol. Med. Rep. 2015, 11, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-C.; Wei, B.-L.; Chiou, W.-F. Dual regulatory effect of plant extracts of Forsythia suspense on RANTES and MCP-1 secretion in influenza A virus-infected human bronchial epithelial cells. J. Ethnopharmacol. 2005, 102, 418–423. [Google Scholar] [CrossRef]
- Kim, M.-S.; Na, H.-J.; Han, S.-W.; Jin, J.-S.; Song, U.-Y.; Lee, E.-J.; Song, B.-K.; Hong, S.-H.; Kim, H.-M. Forsythia fructus Inhibits the Mast-Cell-Mediated Allergic Inflammatory Reactions. Inflammation 2003, 27, 129–135. [Google Scholar] [CrossRef]
- Zhong, W.; Wu, Y.; Xie, X.; Zhou, X.; Wei, M.; Soromou, L.-W.; Ci, X.; Wang, D. Phillyrin attenuates LPS-induced pulmonary inflammation via suppression of MAPK and NF-κB activation in acute lung injury mice. Fitoterapia 2013, 90, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Liu, F.; Zhang, C.; Wang, K.; Jia, X.; Wang, X.; Chen, M.; Li, P.; Su, H.; Wang, Y.; et al. Anti-melanoma activity of Forsythiae Fructus aqueous extract in mice involves regulation of glycerophospholipid metabolisms by UPLC/Q-TOF MS-based metabolomics study. Sci. Rep. 2016, 6, 39415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Piao, X.; Pan, L.; Zeng, Z.; Li, Q.; Xu, X.; Wang, H. Forsythia suspensa extract attenuates lipopolysaccharide-induced inflammatory liver injury in rats via promoting antioxidant defense mechanisms. Anim. Sci. J. 2017, 88, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Kahlos, K. Gloeophyllum odoratum (Brown Rot Fungus); In vitro culture, growth, and production of volatiles, sterols and triterpenes. In Biotechnology in Agriculture and Forestry 37: Medicinal and Aromativ Plants IX; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1996; Volume 1, pp. 144–164. [Google Scholar]
- Tel-Cayan, G. Phenolic profiles, antioxidant, and anticholinesterase activities of three Gloeophyllum species with chemometric approach. J. Food Biochem. 2019, 43, e12790. [Google Scholar] [CrossRef] [PubMed]
- Grienke, U.; Kaserer, T.; Kirchweger, B.; Lambrinidis, G.; Kandel, R.T.; Foster, P.A.; Schuster, D.; Mikros, E.; Rollinger, J.M. Steroid sulfatase inhibiting lanostane triterpenes - Structure activity relationship and in silico insights. Bioorg. Chem. 2020, 95, 103495. [Google Scholar] [CrossRef]
- Francesca, C.; Vittorio, L.; Marko, A.; Paolo, M.; Marina, Z.; Andrej, P.; Bojan, D. Triterpenes from Gloeophyllum odoratum as Potential Leads Towards Potent Thrombin Inhibitors. Lett. Drug Des. Discov. 2010, 7, 521–527. [Google Scholar] [CrossRef]
- Zanoli, P.; Zavatti, M. Pharmacognostic and pharmacological profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef]
- Štulíková, K.; Karabín, M.; Nešpor, J.; Dostálek, P. Therapeutic Perspectives of 8-Prenylnaringenin, a potent phytoestrogen from hops. Molecules 2018, 23, 660. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Fumagalli, M.; Santagostini, L.; Forino, M.; Piazza, S.; Colombo, E.; Taglialatela-Scafati, O.; Fico, G.; Dell’Agli, M. A bio-guided assessment of the anti-inflammatory activity of hop extracts (Humulus lupulus L. cv. Cascade) in human gastric epithelial cells. J. Funct. Foods 2019, 57, 95–102. [Google Scholar] [CrossRef]
- Schink, A.; Neumann, J.; Leifke, A.L.; Ziegler, K.; Froehlich-Nowoisky, J.; Cremer, C.; Thines, E.; Weber, B.; Poeschl, U.; Schuppan, D.; et al. Screening of herbal extracts for TLR2- and TLR4-dependent anti-inflammatory effects. PLoS ONE 2018, 13, e0203907/1–e0203907/27. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, C.; Li, X.; Zhao, Y.; Jing, J.; Ma, Y.; Li, J.; Wei, X.; Li, N. Dietary functional flavonoids as natural hepatoprotective agents against acute liver injury from hop (Humulus lupulus L.). J. Funct. Foods 2018, 45, 471–479. [Google Scholar] [CrossRef]
- Xia, T.; Lin, L.; Zhang, Q.; Jiang, Y.; Li, C.; Liu, X.; Qin, L.; Xin, H. Humulus lupulus L. Extract prevents ovariectomy-induced osteoporosis in mice and regulates activities of osteoblasts and osteoclasts. Chin. J. Integr. Med. 2019. [Google Scholar] [CrossRef]
- Kyrou, I.; Christou, A.; Panagiotakos, D.; Stefanaki, C.; Skenderi, K.; Katsana, K.; Tsigos, C. Effects of a hops (Humulus lupulus L.) dry extract supplement on self-reported depression, anxiety and stress levels in apparently healthy young adults: A randomized, placebo-controlled, double-blind, crossover pilot study. Hormones 2017, 16, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Aghamiri, V.; Mirghafourvand, M.; Mohammad-Alizadeh-Charandabi, S.; Nazemiyeh, H. The effect of Hop (Humulus lupulus L.) on early menopausal symptoms and hot flashes: A randomized placebo-controlled trial. Complement. Ther. Clin. Pract. 2016, 23, 130–135. [Google Scholar] [CrossRef]
- Llorach, R.; Martínez-Sánchez, A.; Tomás-Barberán, F.A.; Gil, M.I.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Azzini, E.; Lazzé, M.C.; Raguzzini, A.; Pizzala, R.; Maiani, G.; Palomba, L.; Maiani, G. Antioxidants in Italian Head Lettuce (Lactuca sativa var. capitata L.) Grown in Organic and Conventional Systems under Greenhouse Conditions. J. Food Biochem. 2014, 38, 56–61. [Google Scholar] [CrossRef]
- Moreno-Escamilla, J.O.; Alvarez-Parrilla, E.; de la Rosa, L.A.; Núñez-Gastélum, J.A.; González-Aguilar, G.A.; Rodrigo-García, J. Effect of Different Elicitors and Preharvest Day Application on the Content of Phytochemicals and Antioxidant Activity of Butterhead Lettuce (Lactuca sativa var. capitata) Produced under Hydroponic Conditions. J. Agric. Food Chem. 2017, 65, 5244–5254. [Google Scholar] [CrossRef] [PubMed]
- Culenova, M.; Sychrova, A.; Hassan, S.T.S.; Berchova-Bimova, K.; Svobodova, P.; Helclova, A.; Michnova, H.; Hosek, J.; Vasilev, H.; Suchy, P.; et al. Multiple In vitro biological effects of phenolic compounds from Morus alba root bark. J. Ethnopharmacol. 2020, 248, 112296. [Google Scholar] [CrossRef]
- Chung, K.O.; Kim, B.Y.; Lee, M.H.; Kim, Y.R.; Chung, H.Y.; Park, J.H.; Moon, J.O. In-vitro and in-vivo anti-inflammatory effect of oxyresveratrol from Morus alba L. J. Pharm. Pharmacol. 2003, 55, 1695–1700. [Google Scholar] [CrossRef]
- Lim, H.J.; Jin, H.-G.; Woo, E.-R.; Lee, S.K.; Kim, H.P. The root barks of Morus alba and the flavonoid constituents inhibit airway inflammation. J. Ethnopharmacol. 2013, 149, 169–175. [Google Scholar] [CrossRef]
- Vogl, S.; Zehl, M.; Picker, P.; Urban, E.; Wawrosch, C.; Reznicek, G.; Saukel, J.; Kopp, B. Identification and quantification of coumarins in Peucedanum ostruthium (L.) Koch by HPLC-DAD and HPLC-DAD-MS. J. Agric. Food Chem. 2011, 59, 4371–4377. [Google Scholar] [CrossRef] [PubMed]
- Palmioli, A.; Bertuzzi, S.; De Luigi, A.; Colombo, L.; La Ferla, B.; Salmona, M.; De Noni, I.; Airoldi, C. bioNMR-based identification of natural anti-Abeta compounds in Peucedanum ostruthium. Bioorg. Chem. 2019, 83, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Hiermann, A.; Schantl, D. Antiphlogistic and antipyretic activity of Peucedanum ostruthium. Planta Med. 1998, 64, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Joa, H.; Vogl, S.; Atanasov, A.G.; Zehl, M.; Nakel, T.; Fakhrudin, N.; Heiss, E.H.; Picker, P.; Urban, E.; Wawrosch, C.; et al. Identification of ostruthin from Peucedanum ostruthium rhizomes as an inhibitor of vascular smooth muscle cell proliferation. J. Nat. Prod. 2011, 74, 1513–1516. [Google Scholar] [CrossRef]
- Padmakumari, K.P.; Sasidharan, I.; Sreekumar, M.M. Composition and antioxidant activity of essential oil of pimento (Pimenta dioica (L) Merr.) from Jamaica. Nat. Prod. Res. 2011, 25, 152–160. [Google Scholar] [CrossRef]
- Doyle, B.J.; Lawal, T.O.; Locklear, T.D.; Hernandez, L.; Perez, A.L.; Patel, U.; Patel, S.; Mahady, G.B. Isolation and identification of three new chromones from the leaves of Pimenta dioica with cytotoxic, oestrogenic and anti-oestrogenic effects. Pharm. Biol. 2018, 56, 235–244. [Google Scholar] [CrossRef]
- Pereira, A.S.P.; Banegas-Luna, A.J.; Pena-Garcia, J.; Perez-Sanchez, H.; Apostolides, Z. Evaluation of the anti-diabetic activity of some common herbs and spices: Providing new insights with inverse virtual screening. Molecules 2019, 24, 4030. [Google Scholar] [CrossRef]
- Ladurner, A.; Zehl, M.; Grienke, U.; Hofstadler, C.; Faur, N.; Pereira, F.C.; Berry, D.; Dirsch, V.M.; Rollinger, J.M. Allspice and clove as source of triterpene acids activating the G protein-coupled bile acid receptor TGR5. Front. Pharmacol. 2017, 8, 468/1–468/13. [Google Scholar] [CrossRef]
- Kamo, T.; Asanoma, M.; Shibata, H.; Hirota, M. Anti-inflammatory Lanostane-Type Triterpene Acids from Piptoporus betulinus. J. Nat. Prod. 2003, 66, 1104–1106. [Google Scholar] [CrossRef]
- Tohtahon, Z.; Xue, J.; Han, J.; Liu, Y.; Hua, H.; Yuan, T. Cytotoxic lanostane triterpenoids from the fruiting bodies of Piptoporus betulinus. Phytochemistry 2017, 143, 98–103. [Google Scholar] [CrossRef]
- Khalilov, Q.; Li, L.; Liu, Y.; Tohtahon, Z.; Chen, X.; Aisa, H.A.; Yuan, T. Piptolinic acids F-J, five new lanostane-type triterpenoids from Piptoporus betulinus. Nat. Prod. Res. 2019, 33, 3044–3051. [Google Scholar] [CrossRef] [PubMed]
- Jaric, S.; Kostic, O.; Mataruga, Z.; Pavlovic, D.; Pavlovic, M.; Mitrovic, M.; Pavlovic, P. Traditional wound-healing plants used in the Balkan region (Southeast Europe). J. Ethnopharmacol. 2018, 211, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Kovac, I.; Durkac, J.; Holly, M.; Jakubcova, K.; Perzelova, V.; Mucaji, P.; Svajdlenka, E.; Sabol, F.; Legath, J.; Belak, J.; et al. Plantago lanceolata L. water extract induces transition of fibroblasts into myofibroblasts and increases tensile strength of healing skin wounds. J. Pharm. Pharmacol. 2015, 67, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, H. Falsification of herba Potentillae aureae L. Pharm. Acta Helv. 1938, 13, 67–70. [Google Scholar]
- Schappi, B. Growth dynamics and population development in an alpine grassland under elevated CO2. Oecologia 1996, 106, 93–99. [Google Scholar] [CrossRef]
- Kołodziejek, J.; Patykowski, J.; Wala, M. Dormancy, germination, and sensitivity to salinity stress in five species of Potentilla (Rosaceae). Botany 2019, 97, 452–462. [Google Scholar] [CrossRef]
- Tomczyk, M.; Latté, K.P. Potentilla—A review of its phytochemical and pharmacological profile. J. Ethnopharmacol. 2009, 122, 184–204. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M.F. Medicinal Plants Traditionally Used for Treatment of Obesity and Diabetes Mellitus—Screening for Pancreatic Lipase and α-Amylase Inhibition. Phytother. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef]
- Hřibová, P.; Khazneh, E.; Žemlička, M.; Švajdlenka, E.; Ghoneim, M.M.; Elokely, K.M.; Ross, S.A. Antiurease activity of plants growing in the Czech Republic. Nat. Prod. Res. 2014, 28, 868–873. [Google Scholar] [CrossRef]
- Kwon, H.J.; Hong, Y.K.; Kim, K.H.; Han, C.H.; Cho, S.H.; Choi, J.S.; Kim, B.-W. Methanolic extract of Pterocarpus santalinus induces apoptosis in HeLa cells. J. Ethnopharmacol. 2006, 105, 229–234. [Google Scholar] [CrossRef]
- Wu, S.-F.; Hwang, T.-L.; Chen, S.-L.; Wu, C.-C.; Ohkoshi, E.; Lee, K.-H.; Chang, F.-R.; Wu, Y.-C. Bioactive components from the heartwood of Pterocarpus santalinus. Bioorg. Med. Chem. Lett. 2011, 21, 5630–5632. [Google Scholar] [CrossRef] [PubMed]
- Challa, C.S.; Lokesh, T.; Nayakanti, D.; Varadacharyulu, N.C. Phytochemical and anti-microbial study of Pterocarpus santalinus Linn Heartwood. Res. J. Life Sci. Bioinf. Pharm. Chem. Sci. 2019, 5, 451–464. [Google Scholar] [CrossRef]
- Dhande, P.P.; Gupta, A.O.; Jain, S.; Dawane, J.S. Anti-inflammatory and analgesic activities of topical formulations of Pterocarpus santalinus powder in rat model of chronic inflammation. J. Clin. Diagn. Res. 2017, 11, Ff01–Ff04. [Google Scholar] [CrossRef] [PubMed]
- Bonjardim, L.R.; Silva, A.M.; Oliveira, M.G.; Guimaraes, A.G.; Antoniolli, A.R.; Santana, M.F.; Serafini, M.R.; Santos, R.C.; Araujo, A.A.; Estevam, C.S.; et al. Sida cordifolia leaf extract reduces the orofacial nociceptive response in mice. Phytother. Res. 2011, 25, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Lakshmi, P.K.; Sahi, C.; Pawar, R.S. Sida cordifolia accelerates wound healing process delayed by dexamethasone in rats: Effect on ROS and probable mechanism of action. J. Ethnopharmacol. 2019, 235, 279–292. [Google Scholar] [CrossRef]
- Rejitha, S.; Prathibha, P.; Indira, M. Amelioration of alcohol-induced hepatotoxicity by the administration of ethanolic extract of Sida cordifolia Linn. Br. J. Nutr. 2012, 108, 1256–1263. [Google Scholar] [CrossRef]
- Martins, C.A.F.; Campos, M.L.; Irioda, A.C.; Stremel, D.P.; Trindade, A.; Pontarolo, R. Anti-Inflammatory effect of Malva sylvestris, Sida cordifolia, and Pelargonium graveolens is related to inhibition of prostanoid production. Molecules 2017, 22, 1883. [Google Scholar] [CrossRef]
- Fateh, A.H.; Mohamed, Z.; Chik, Z.; Alsalahi, A.; Md Zain, S.R.; Alshawsh, M.A. Mutagenicity and genotoxicity effects of Verbena officinalis leaves extract in Sprague-Dawley Rats. J. Ethnopharmacol. 2019, 235, 88–99. [Google Scholar] [CrossRef]
- Shu, J.; Chou, G.; Wang, Z. Two new iridoids from Verbena officinalis L. Molecules 2014, 19, 10473–10479. [Google Scholar] [CrossRef]
- Casanova, E.; García-Mina, J.M.; Calvo, M.I. Antioxidant and antifungal activity of Verbena officinalis L. Leaves. Plant. Foods Hum. Nutr. 2008, 63, 93–97. [Google Scholar] [CrossRef]
- Rehecho, S.; Hidalgo, O.; García-Iñiguez de Cirano, M.; Navarro, I.; Astiasarán, I.; Ansorena, D.; Cavero, R.Y.; Calvo, M.I. Chemical composition, mineral content and antioxidant activity of Verbena officinalis L. LWT Food Sci. Technol. 2011, 44, 875–882. [Google Scholar] [CrossRef]
- Calvo, M.I. Anti-inflammatory and analgesic activity of the topical preparation of Verbena officinalis L. J. Ethnopharmacol. 2006, 107, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.W.; Khan, A.U.; Ahmed, T. Anticonvulsant, Anxiolytic, and Sedative Activities of Verbena officinalis. Front. Pharmacol. 2016, 7, 499. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, A.; Kazemi, F.; Mehrzadi, S.; Dehpour, A.R.; Mehr, S.E.; Rezayat, S.M. Anticonvulsant effects of aerial parts of Verbena officinalis extract in mice: Involvement of benzodiazepine and opioid receptors. J. Evid. Based Complement. Altern. Med. 2017, 22, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Deepak, M.; Handa, S.S. Antiinflammatory activity and chemical composition of extracts of Verbena officinalis. Phytother. Res. 2000, 14, 463–465. [Google Scholar] [CrossRef]
- Grawish, M.E.; Anees, M.M.; Elsabaa, H.M.; Abdel-Raziq, M.S.; Zedan, W. Short-term effects of Verbena officinalis Linn decoction on patients suffering from chronic generalized gingivitis: Double-blind randomized controlled multicenter clinical trial. Quintessence Int. 2016, 47, 491–498. [Google Scholar] [CrossRef]
- Mechtcheriakova, D.; Schabbauer, G.; Lucerna, M.; Clauss, M.; De Martin, R.; Binder, B.R.; Hofer, E. Specificity, diversity, and convergence in VEGF and TNF-α signaling events leading to tissue factor upregulation via EGR-1 in endothelial cells. FASEB J. 2001, 15. [Google Scholar] [CrossRef]
- Hoffmann, A.; Levchenko, A.; Scott, M.L.; Baltimore, D. The IkappaB-NF-kappaB signaling module: Temporal control and selective gene activation. Science 2002, 298, 1241–1245. [Google Scholar] [CrossRef]
- Delhase, M.; Hayakawa, M.; Chen, Y.; Karin, M. Positive and negative regulation of IkappaB kinase activity through IKKbeta subunit phosphorylation. Science 1999, 284, 309–313. [Google Scholar] [CrossRef]
- Sarkhail, P. Traditional uses, phytochemistry and pharmacological properties of the genus Peucedanum: A review. J. Ethnopharmacol. 2014, 156, 235–270. [Google Scholar] [CrossRef]
- Chun, J.M.; Lee, A.R.; Kim, H.S.; Lee, A.Y.; Gu, G.J.; Moon, B.C.; Kwon, B.I. Peucedanum japonicum extract attenuates allergic airway inflammation by inhibiting Th2 cell activation and production of pro-inflammatory mediators. J. Ethnopharmacol. 2018, 211, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Y.; Wang, J.S.; Wu, F.H.; Li, J.; Kong, L.Y. The effects of (+/−)-Praeruptorin A on airway inflammation, remodeling and transforming growth factor-beta1/Smad signaling pathway in a murine model of allergic asthma. Int. Immunopharmacol. 2012, 14, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Y.; Wu, F.H.; Wang, J.S.; Li, J.; Kong, L.Y. Attenuation of airway hyperreactivity and T helper cell type 2 responses by coumarins from Peucedanum praeruptorum Dunn in a murine model of allergic airway inflammation. J. Ethnopharmacol. 2012, 141, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.J.; Ci, W.; Wang, G.F.; Zhang, J.Y.; Wu, S.Y.; Xu, W.; Jin, H.; Zhu, Z.G.; Zhang, J.J.; Pang, J.X.; et al. Praeruptorin A inhibits lipopolysaccharide-induced inflammatory response in murine macrophages through inhibition of NF-kappaB pathway activation. Phytother. Res. 2011, 25, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Sun, Z.; Lu, Y.; Li, Y.Z.; Xu, H.R.; Zeng, C.Q. Osthole inhibits proliferation and induces apoptosis in BV-2 microglia cells in kainic acid-induced epilepsy via modulating PI3K/AKt/mTOR signalling way. Pharm. Biol. 2019, 57, 238–244. [Google Scholar] [CrossRef]
- Hadacek, F.; Muller, C.; Werner, A.; Greger, H.; Proksch, P. Analysis, isolation and insecticidal activity of linear furanocoumarins and other coumarin derivatives from Peucedanum (Apiaceae: Apioideae). J. Chem. Ecol. 1994, 20, 2035–2054. [Google Scholar] [CrossRef]
- Zimecki, M.; Artym, J.; Cisowski, W.; Mazol, I.; Włodarczyk, M.; Gleńsk, M. Immunomodulatory and anti-inflammatory activity of selected osthole derivatives. Z. Fur Naturforsch. Sect. C J. Biosci. 2009, 64, 361–368. [Google Scholar] [CrossRef]
- Grienke, U.; Foster, P.A.; Zwirchmayr, J.; Tahir, A.; Rollinger, J.M.; Mikros, E. (1)H NMR-MS-based heterocovariance as a drug discovery tool for fishing bioactive compounds out of a complex mixture of structural analogues. Sci. Rep. 2019, 9, 11113. [Google Scholar] [CrossRef]
- Werner, S.L.; Kearns, J.D.; Zadorozhnaya, V.; Lynch, C.; O’Dea, E.; Boldin, M.P.; Ma, A.; Baltimore, D.; Hoffmann, A. Encoding NF-kappaB temporal control in response to TNF: Distinct roles for the negative regulators IkappaBalpha and A20. Genes Dev. 2008, 22, 2093–2101. [Google Scholar] [CrossRef]







{kind=link}
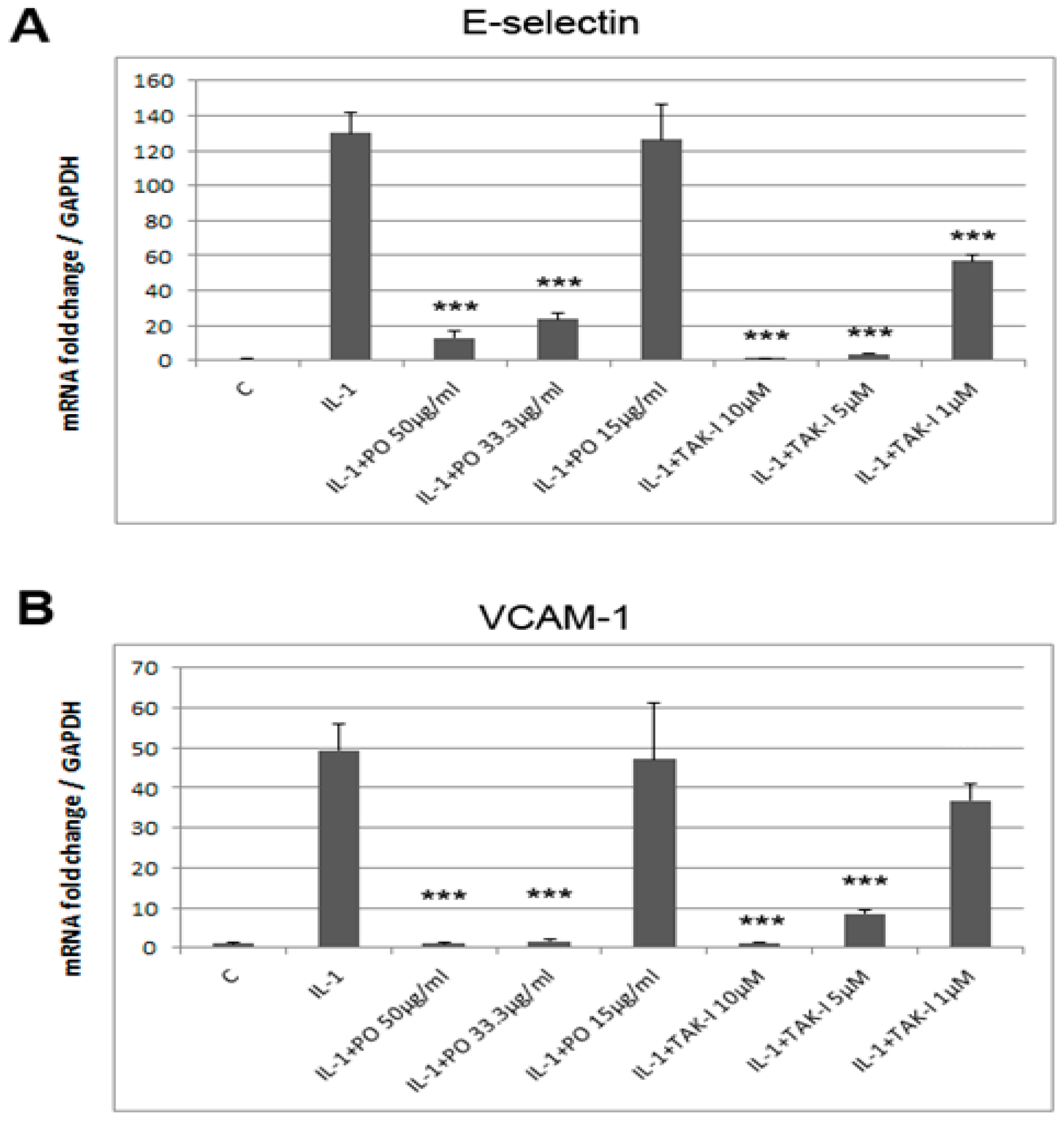{kind=link}
{kind=link}
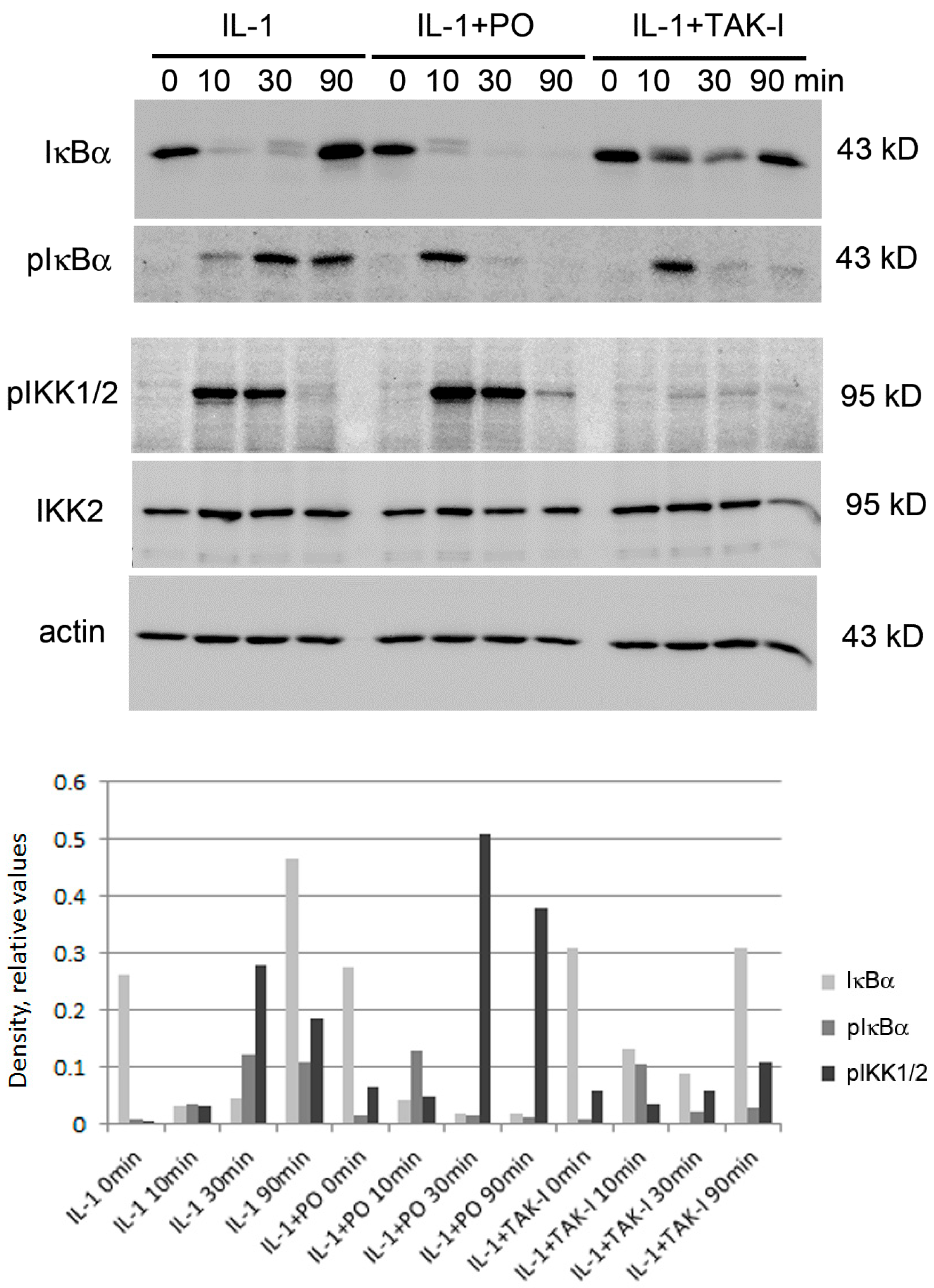{kind=link}
{kind=link}
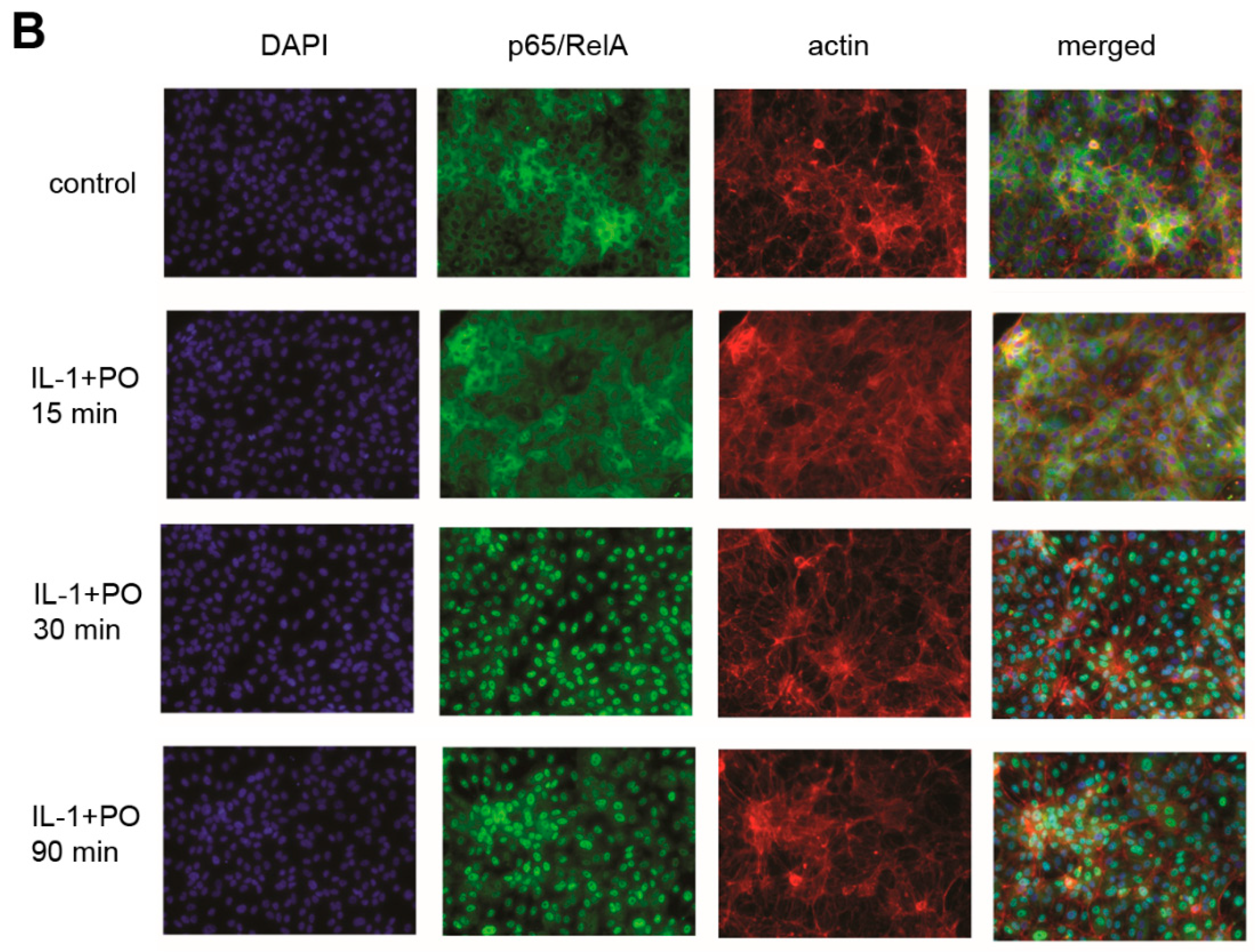{kind=link}
| Species | Family | Organ | Provider/Collection Site | Voucher Specimen/Charge Number | Used Extract |
|---|---|---|---|---|---|
| Aquilegia vulgaris L. | Ranunculaceae | herb | Padma AG, Wetzikon, Switzerland | Ch.Nr.: 21290300 | LLE |
| Azadirachta indica A.Juss. | Meliaceae | fruits | Padma AG, Wetzikon, Switzerland | JR-20150615-A1 Ch.Nr.: 2021108301 | LLE |
| Burkea africana Hook. | Leguminosae | bark/heart-wood | Northwest of Zeerust, South Africa | Voucher no 5858 [14] | Ethanol (EtOH) extract |
| Capsella bursa-pastoris (L.) Medik. | Brassicaceae | herb | Kottas Pharma GmbH, Vienna, Austria | Ch.Nr.: 1223296 | MeOH |
| Elettaria cardamomum (L.) Maton | Zingiberaceae | fruits | Padma AG, Wetzikon, Switzerland | Ch.Nr.: W17201998 | LLE w/o TD |
| Euphrasia officinalis L. | Orobanchaceae | herb | Kottas Pharma GmbH, Vienna, Austria | JR-20090625-A1 Ch.Nr.: KLA90309 | LLE |
| Fomitopsis pinicola (Sw.) P. Karst. (strain 10) | Fomitopsidaceae | fruit body | Viggartal, Ellbögen, Austria (grown on dead spruce trunk); Ursula Peintner | FompinE0010 | LLE w/o TD |
| Forsythia suspensa (Thunb.) Vahl | Oleaceae | fruits | Plantasia GmbH, Oberndorf bei Salzburg, Austria | Ch.Nr.: 020410 | LLE |
| Gloeophyllum odoratum (Wulfen) Imazeki (strain 54) | Gloeophyllaceae | fruit body | Oberperfuss, Austria (grown on spruce) Ursula Peintner | JR-20140310-A1 GloodoE0054 | EtOH extract [17] |
| Humulus lupulus L. | Cannabaceae | flower | Kottas Pharma GmbH, Vienna, Austria | Ch.Nr.: W12203440 | CH2Cl2 |
| Lactuca sativa L. | Compositae | leaves | Padma AG, Wetzikon, Switzerland | Ch.Nr.: 21400300 | LLE |
| Morus alba L. | Moraceae | root bark | Plantasia GmbH, Oberndorf bei Salzburg, Austria | Ch.Nr.: 710797 | LLE |
| Peucedanum ostruthium (L.) W.D.J.Koch | Apiaceae | roots/rhizom | Birgitzköpfl, Axamer Lizum, Austria | JR-20120814-A1 (small scale extract) | LLE |
| Peucedanum ostruthium (L.) W.D.J.Koch | Apiaceae | roots/rhizom | Kottas Pharma GmbH, Vienna, Austria | JR-20180119-A2 Ch.Nr.: P17301770 (large scale extract) | LLE w/o TD [18] |
| Pimenta dioica (L.) Merr. | Myrtaceae | fruits | Padma AG, Wetzikon, Switzerland | JR-20150615-A4 Ch.Nr.: 21362100 | LLE |
| Piptoporus betulinus (Bull.) P. Karst. (strain 39) | Fomitopsida-ceae | fruit body | Vahrn bei Brixen, Italy, grown on birch, Ursula Peintner | PipbetE0039 | LLE w/o TD |
| Plantago lanceolate L. | Plantaginaceae | herb | Padma AG, Wetzikon, Switzerland | Ch.Nr.: 21327101 | LLE |
| Potentilla aurea L. | Rosaceae | herb | Padma AG, Wetzikon, Switzerland | JR-20150615-A3 Ch.Nr.: 21161301 | LLE |
| Pterocarpus santalinus L.f. | Leguminosae | wood | Padma AG, Wetzikorn, Switzerland | JR-20190315–A1 Ch.Nr.: P16301836 | LLE |
| Sida cordifolia L. | Malvaceae | herb | Padma AG, Wetzikon, Switzerland | JR-20150615-A2 Ch.Nr.: 20981300 | LLE |
| Verbena officinalis L. | Verbenaceae | herb | Ehnbach-klamm, Zirl, Austria | JR-20120801-A12 | LLE |
| Herbal/Fungal Material | Traditional Use | In Vitro/In Vivo Studies | Clinical Studies |
|---|---|---|---|
| Herbs of Aquilegia vulgaris (European columbine) | Southern Europe, Asia and Africa; chronic skin inflammation, liver and bile duct disorders, jaundice [23,24,25,26]. | Hepatoprotective [24,26] and antioxidant effects [23,25]. | |
| Fruits of Azadirachta indica (Neem fruits) | Medicinal systems (Ayurveda [27,28], Unani and Siddha [29]); as astringents and antihelmintic agent [30]. | Antioxidant and neuroprotective effecs [27]; anti-bacterial [31], anti-nociceptive [32] and osteogenic activities [33]. | |
| Roots and bark of Burkea africana (Seringa tree) | Sub-Saharan Africa; Pain (e.g., headache and migraine), inflammation and as wound-healing agents [34]. | Antioxidant [34] and anti-viral activites [14,15]. | |
| Herbs of Capsella bursa-pastoris (Shepherd’s purse) | Traditional Chinese [35] and Korean folk medicine; treatment of hypertension and edema [36]. | Anti-inflammatory [35,36,37] and anti-viral activities [15]. | Reduced postpartum hemorrhage bleeding in women [38]. |
| Fruits of Elettaria cardamomum (Queen of spices) | Ayurveda, Siddha and Unani [39]; Treatment of asthma, teeth infections, cataracts, digestive and kidney disorders [40]. | Anti-inflammatory [41,42,43], anti-tumor [42,44], anti-microbial [41,45] activities;Immunomodulatory effects [42]. | Improved parameters of inflammation and oxidative stress in pre-diabetic women [46]. |
| Herbs of Euphrasia officinalis (Eyebright herb) | Anthroposophical medicine [47]; conjunctivitis, ophthalmia and ocular allergies [48] | Anti-inflammatory activity [37,49]; reduction of UVB-induced cell death and increased wound-healing ability [50]. | Effective against allergic conjunctivitis and conjunctivitis due to external irritants or other causes [47]. |
| Fruit bodies of Fomitopsis pinicola (Red banded polypore) | Traditional Asian [51] and European medicine; Nausea, headache and liver problems [22]. | Anti-tumor [52,53], anti-oxidant [54], anti-bacterial [55] and anti-inflammatory activities [56,57]. | |
| Fruits of Forsythia suspensa (Lian Qiao) | Traditional Chinese Medicine (TCM) as heart-clearing and detoxifying agent [58,59,60]. | Anti-inflammatory [59,61,62], anti-tumour [59,63], antioxidant [64] and anti-viral activities [15,58,60]. | |
| Fruit bodies of Gloeophyllum odoratum (Anise mazegill) | Central Europe, Asia and North America [17,65] | Anti-influenza virus activity [17]; antioxidant [66] and anti-cancer activities [67]; Thrombin inhibition [68]. | |
| Female inflorescences of Humulus lupulus (Hops) | Management of sleeping disorders, as sedative, and bitter stomachic [69,70]. | Anti-inflammatory [71,72], antioxidant activities; hepatoprotective [73] and osteoprotective effects [74]. | Effective against mild depression, anxiety and stress [75]; reduction of early postmenopausal symptoms (hot flashes, sexual dysfunction, anxiety and depression) [76]. |
| Leaves of Lactuca sativa (Butterhead lettuce) | Popular vegetable; Perception as “healthy” food [77]. | Antioxidant properties [78,79]. | |
| Root bark of Morus alba (Sang-Bai-Pi) | TCM; Throat infections, asthma, fever and inflammation [80]. | Antioxidant, anti-cancer, anti-inflammatory [80,81,82] and anti-viral activities [15]. | |
| Roots and rhizome of Peucedanum ostruthium (Masterwort) | Austrian and Italian folk medicine; gastro-intestinal, cardiovascular and respiratory diseases [37,83,84,85]. | Anti-inflammatory activity [37,85]; inhibition of vascular smooth muscle cell proliferation [86]; anti-amyloidogenic activity [84]. | |
| Fruits of Pimenta dioica (Allspice) | Jamaican [87] and Costa Rican medicine; menopausal symptoms, dysmenorrhea and dyspepsia [88]. | Anti-oxidant [87], anti-diabetic effects [89] and metabolic disorders via upregulation of TGR5 [90]; oestrogenic effects, inhibitory effect on breast and gastric cancer cells [88]. | |
| Fruit bodies of Piptoporus betulinus (Birch polypore) | Central European folk medicine [22]; fatigue and immune-enhancing properties [91] | Anti-microbial [92], anti-inflammatory [91] and anti-cancer activities [93]. | |
| Leaves of Plantago lanceolata (Ribwort plantain) | Slovakian and Southeast European folk medicine; gastric ulcers, respiratory infections and wound healing [94,95]. | Anti-inflammatory activities [36]; improved skin wound healing [95]. | |
| Herbs of Potentilla aurea (Goldfinger herb) | Southern and Central Europe [96,97,98], homeopathic medications; diarrhea, diabetes mellitus and inflammations [99]; | Pancreatic lipase and α-amylase inhibition [100]; anti-urease activity [101] | |
| Heartwood of Pterocarpus santalinus (Red sanders) | India and Korea; Inflammation, mental aberrations, cancer and ulcer [102,103] | Antioxidant properties; anti-microbial [104] and anti-inflammatory activities [103,105]. | |
| Herbs of Sida cordifolia (Indian mallow) | Indian, Chinese, African and Brazilian medicine; skin diseases [106,107], inflammation of oral mucosa, nasal congestion, asthmatic bronchitis and rheumatism [108]. | Anti-nociceptive [106], anti-inflammatory [109], antioxidant activity and improved wound healing [107]; Hepatoprotective effects [108]. | |
| Herbs of Verbena officinalis (Vervain) | European [110] and Chinese folk medicine; rheumatism and bronchitis [111]. | Anti-oxidant [112,113,114], anxiolytic, sedative, anti-convulsant effects [115,116]; anti-inflammatory activities [37,117]. | Ameliorative effect of chronic generalized gingivitis [118]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lammel, C.; Zwirchmayr, J.; Seigner, J.; Rollinger, J.M.; de Martin, R. Peucedanum ostruthium Inhibits E-Selectin and VCAM-1 Expression in Endothelial Cells through Interference with NF-κB Signaling. Biomolecules 2020, 10, 1215. https://doi.org/10.3390/biom10091215
Lammel C, Zwirchmayr J, Seigner J, Rollinger JM, de Martin R. Peucedanum ostruthium Inhibits E-Selectin and VCAM-1 Expression in Endothelial Cells through Interference with NF-κB Signaling. Biomolecules. 2020; 10(9):1215. https://doi.org/10.3390/biom10091215
Chicago/Turabian StyleLammel, Christoph, Julia Zwirchmayr, Jaqueline Seigner, Judith M. Rollinger, and Rainer de Martin. 2020. "Peucedanum ostruthium Inhibits E-Selectin and VCAM-1 Expression in Endothelial Cells through Interference with NF-κB Signaling" Biomolecules 10, no. 9: 1215. https://doi.org/10.3390/biom10091215
APA StyleLammel, C., Zwirchmayr, J., Seigner, J., Rollinger, J. M., & de Martin, R. (2020). Peucedanum ostruthium Inhibits E-Selectin and VCAM-1 Expression in Endothelial Cells through Interference with NF-κB Signaling. Biomolecules, 10(9), 1215. https://doi.org/10.3390/biom10091215