DNA Phosphorothioate Modifications Are Widely Distributed in the Human Microbiome

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Multiple Sequence Alignments
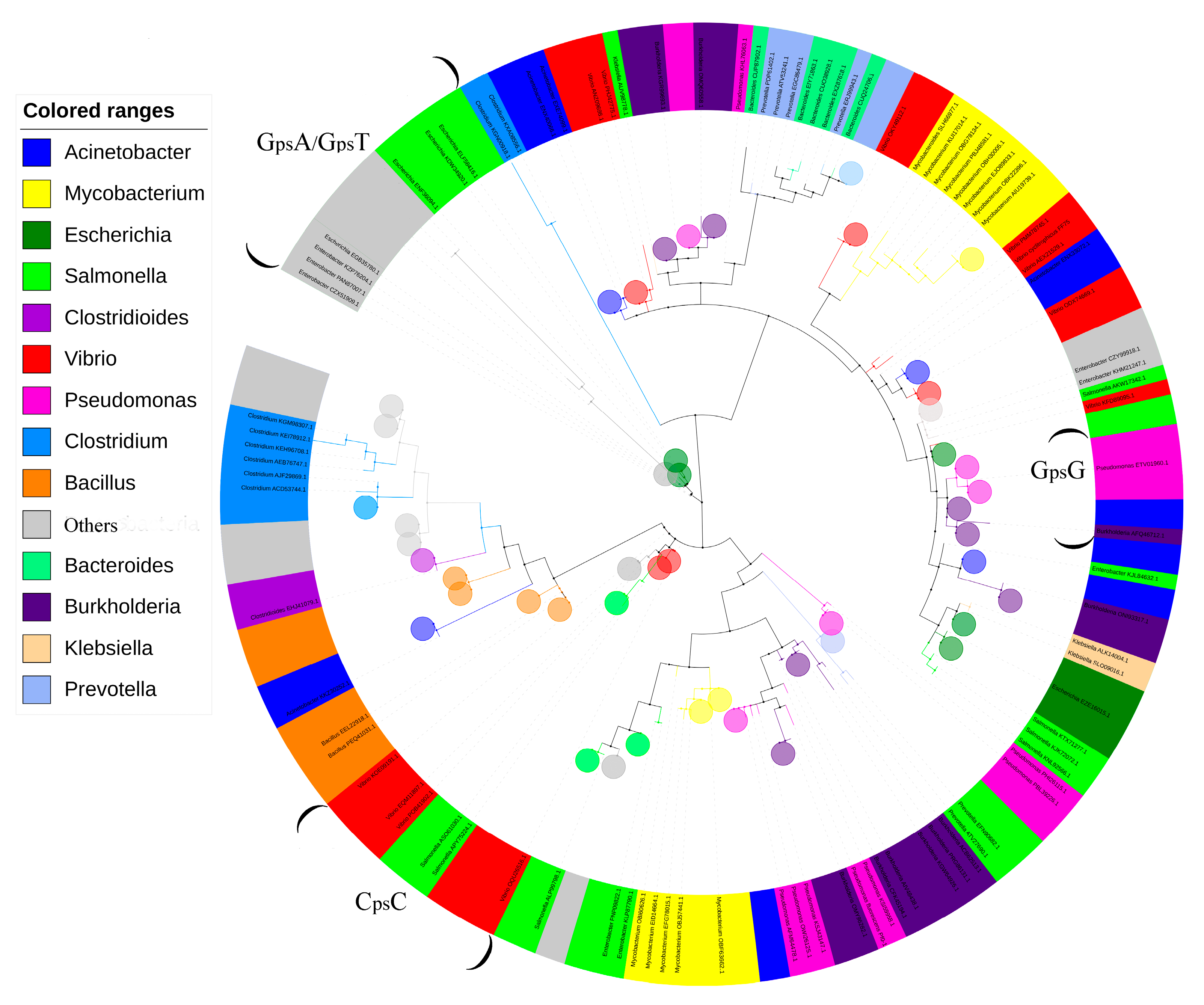
2.2. Phylogenic Analysis
2.3. Gene Abundance Calculation
2.4. Fecal DNA Preparation
2.5. Detection of PT-Linked Dinucleotides
3. Results
3.1. PT Genes Are Widely Distributed in the Human Microbiome
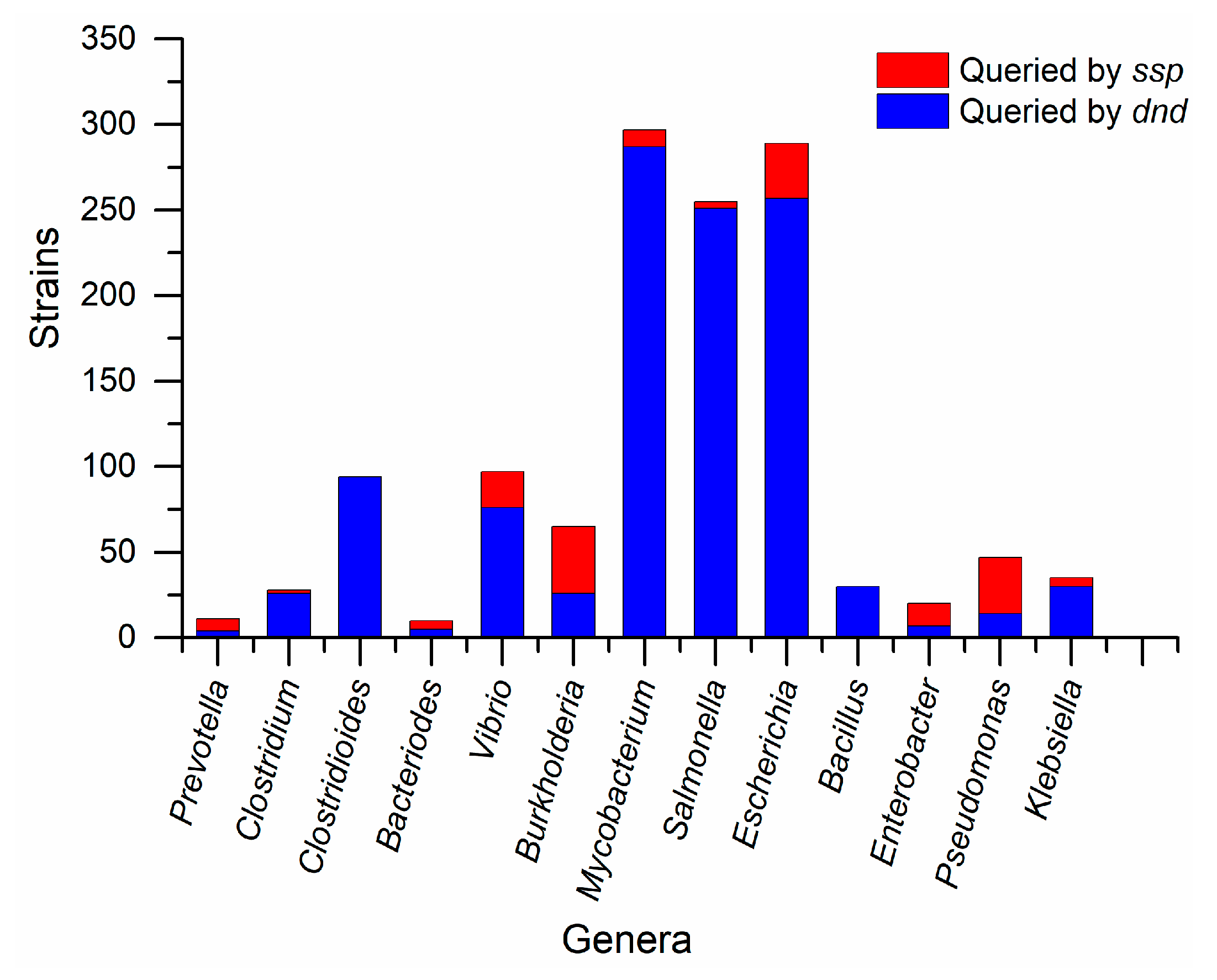
3.2. Species Difference of PT-Related Genes
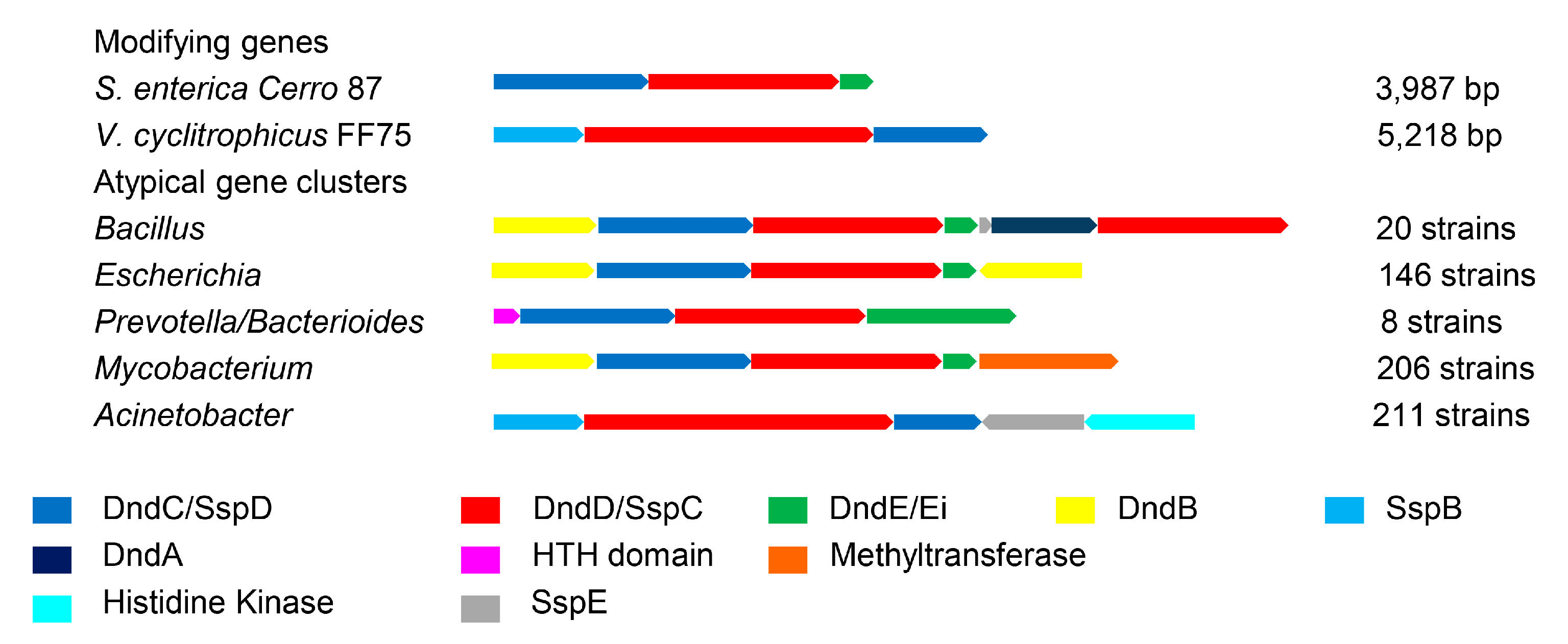
3.3. Atypical PT Gene Clusters of Different Genera
3.4. Detection of PT-Linked Dinucleotides in Human Fecal DNA
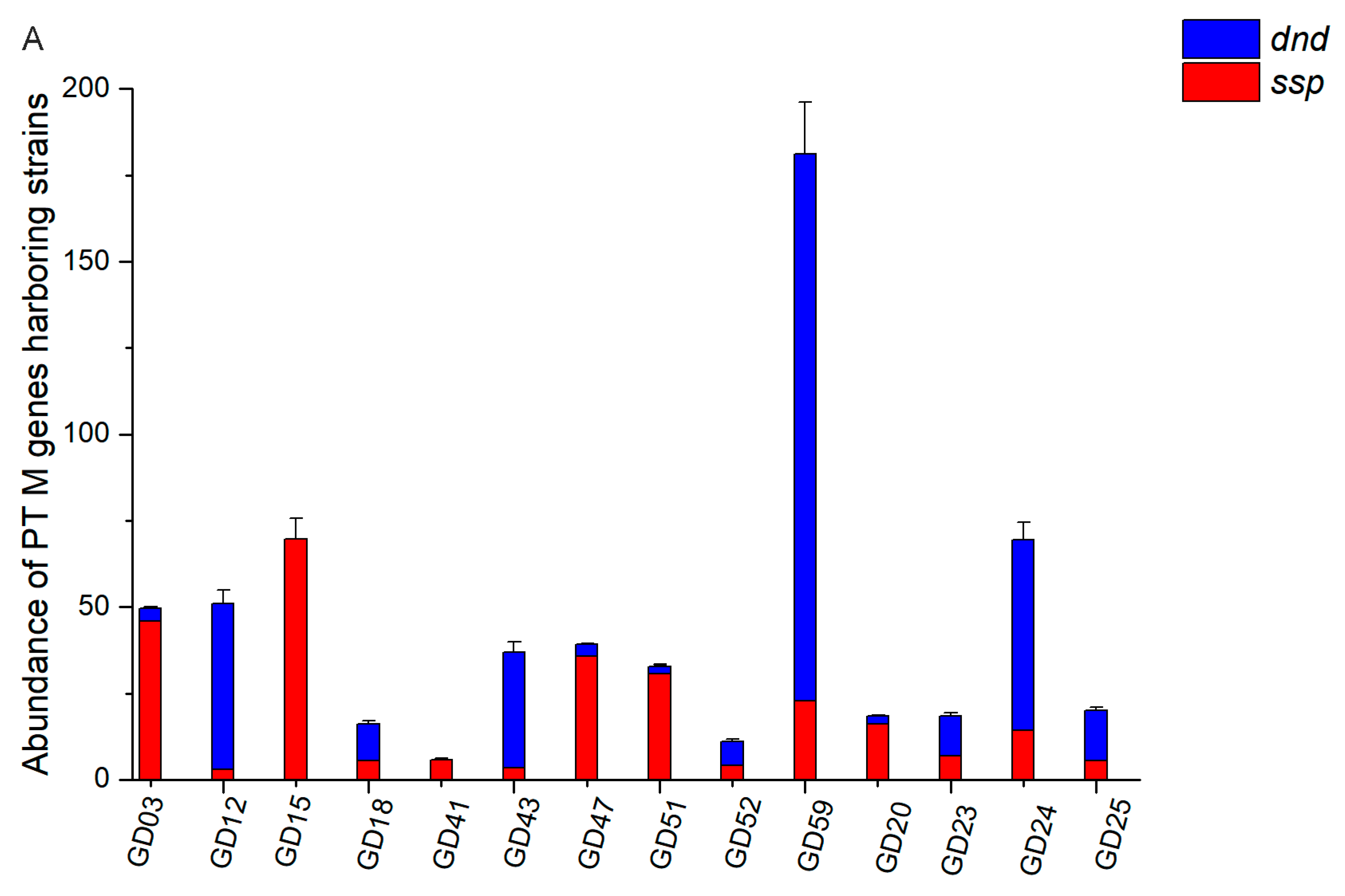
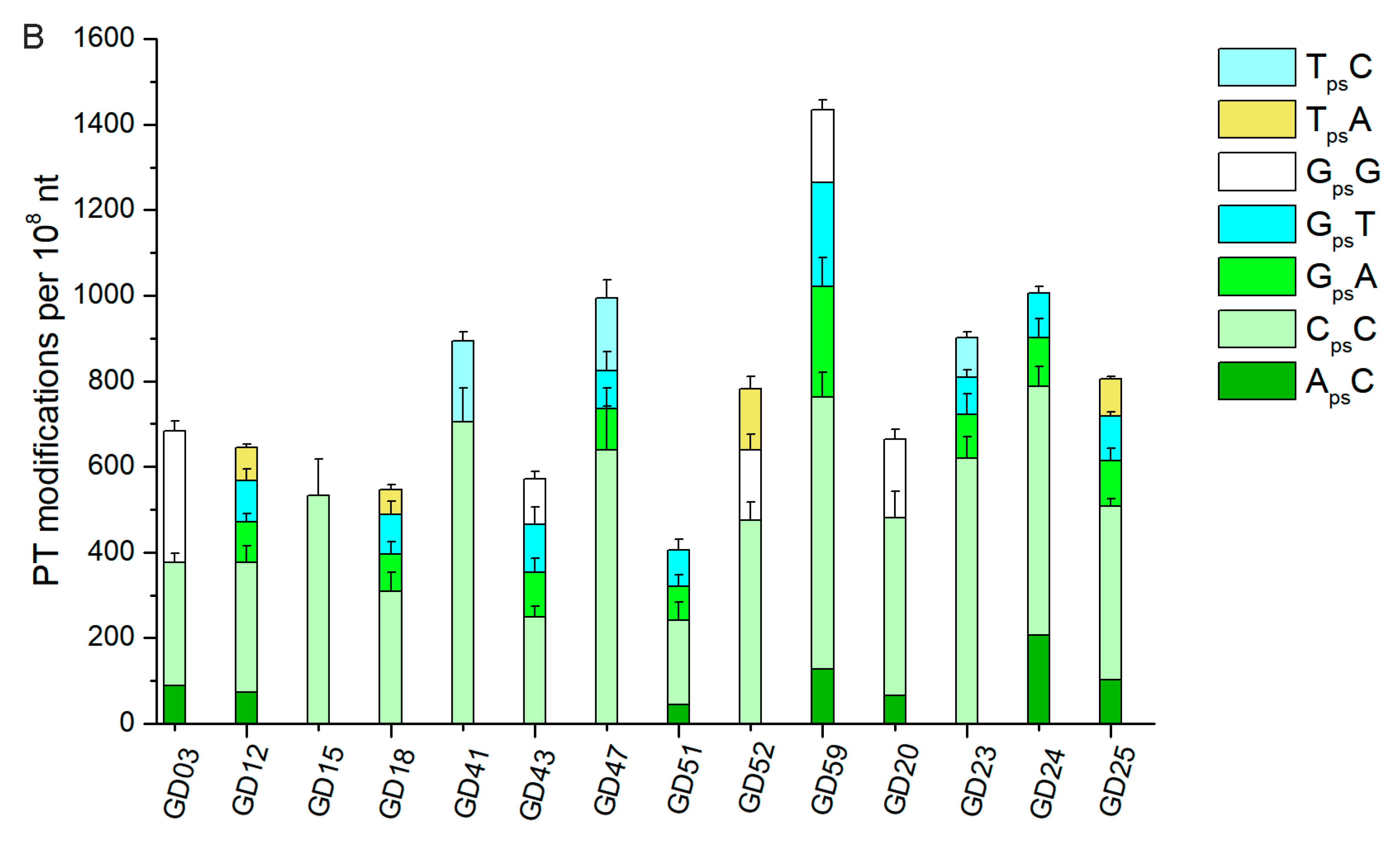
3.5. The Abundance of PT Modifications Varied Among Individuals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Chen, S.; Xu, T.; Taghizadeh, K.; Wishnok, J.S.; Zhou, X.; You, D.; Deng, Z.; Dedon, P.C. Phosphorothioation of DNA in bacteria by dnd genes. Nat. Chem. Biol. 2007, 3, 709–710. [Google Scholar] [CrossRef]
- Zhou, X.; He, X.; Liang, J.; Li, A.; Xu, T.; Kieser, T.; Helmann, J.D.; Deng, Z. A novel DNA modification by sulphur. Mol. Microbiol. 2005, 57, 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Chen, C.; DeMott, M.S.; Cheng, Q.; Clark, T.A.; Xiong, X.; Zheng, X.; Butty, V.; Levine, S.S.; Yuan, G.; et al. Genomic mapping of phosphorothioates reveals partial modification of short consensus sequences. Nat. Commun. 2014, 5, 3951. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Wu, G.; Wei, Y.; Liu, L.; Zhang, Y.; Su, R.; Jiang, X.; Li, M.; Gao, H.; Tian, X.; et al. SspABCD-SspE is a phosphorothioation-sensing bacterial defence system with broad anti-phage activities. Nat. Microbiol. 2020, 5, 917–928. [Google Scholar] [CrossRef] [PubMed]
- You, D.; Wang, L.; Yao, F.; Zhou, X.; Deng, Z. A novel DNA modification by sulfur: DndA is a NifS-like cysteine desulfurase capable of assembling DndC as an iron-sulfur cluster protein in Streptomyces lividans. Biochemistry 2007, 46, 6126–6133. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Xiong, W.; Yang, Y.; Li, F.; Zhou, X.; Wang, Z.; Deng, Z.; Liang, J. A novel target of IscS in Escherichia coli: Participating in DNA phosphorothioation. PLoS ONE 2012, 7, e51265. [Google Scholar] [CrossRef]
- Pu, T.; Mei, Z.; Zhang, W.; Liang, W.J.; Zhou, X.; Liang, J.; Deng, Z.; Wang, Z. An in vitro DNA phosphorothioate modification reaction. Mol. Microbiol. 2019, 113, 452–463. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Zheng, T.; Kong, L.; Zhu, S.; Sun, Y.; Deng, Z.; Yang, L.; You, D. Quantitative mapping of DNA phosphorothioatome reveals phosphorothioate heterogeneity of low modification frequency. PLoS Genet. 2019, 15, e1008026. [Google Scholar] [CrossRef]
- Xiong, L.; Liu, S.; Chen, S.; Xiao, Y.; Zhu, B.; Gao, Y.; Zhang, Y.; Chen, B.; Luo, J.; Deng, Z.; et al. A new type of DNA phosphorothioation-based antiviral system in archaea. Nat. Commun. 2019, 10, 1688. [Google Scholar] [CrossRef]
- Xu, T.; Yao, F.; Zhou, X.; Deng, Z.; You, D. A novel host-specific restriction system associated with DNA backbone S-modification in Salmonella. Nucleic Acids Res. 2010, 38, 7133–7141. [Google Scholar] [CrossRef]
- Tong, T.; Chen, S.; Wang, L.; Tang, Y.; Ryu, J.Y.; Jiang, S.; Wu, X.; Chen, C.; Luo, J.; Deng, Z.; et al. Occurrence, evolution, and functions of DNA phosphorothioate epigenetics in bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E2988–E2996. [Google Scholar] [CrossRef] [PubMed]
- Kellner, S.; DeMott, M.S.; Cheng, C.P.; Russell, B.S.; Cao, B.; You, D.; Dedon, P.C. Oxidation of phosphorothioate DNA modifications leads to lethal genomic instability. Nat. Chem. Biol. 2017, 13, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Takano, E.; Breitling, R. Detecting sequence homology at the gene cluster level with MultiGeneBlast. Mol. Biol. Evol. 2013, 30, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Dawid, S.; Barenkamp, S.J.; St Geme, J.W. Variation in expression of the Haemophilus influenzae HMW adhesins: A prokaryotic system reminiscent of eukaryotes. Proc. Natl. Acad. Sci. USA 1999, 96, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.B.; Almeida, M.; Juncker, A.S.; Rasmussen, S.; Li, J.; Sunagawa, S.; Plichta, D.R.; Gautier, L.; Pedersen, A.G.; Le Chatelier, E.; et al. Identification and assembly of genomes and genetic elements in complex metagenomic samples without using reference genomes. Nat. Biotechnol. 2014, 32, 822–828. [Google Scholar] [CrossRef]
- Wang, L.; Chen, S.; Vergin, K.L.; Giovannoni, S.J.; Chan, S.W.; DeMott, M.S.; Taghizadeh, K.; Cordero, O.X.; Cutler, M.; Timberlake, S.; et al. DNA phosphorothioation is widespread and quantized in bacterial genomes. Proc. Natl. Acad. Sci. USA 2011, 108, 2963–2968. [Google Scholar] [CrossRef]
- Cheng, Q.; Cao, B.; Yao, F.; Li, J.; Deng, Z.; You, D. Regulation of DNA phosphorothioate modifications by the transcriptional regulator DptB in Salmonella. Mol. Microbiol. 2015, 97, 1186–1194. [Google Scholar] [CrossRef]
- Aagaard, K.; Petrosino, J.; Keitel, W.; Watson, M.; Katancik, J.; Garcia, N.; Patel, S.; Cutting, M.; Madden, T.; Hamilton, H.; et al. The Human Microbiome Project strategy for comprehensive sampling of the human microbiome and why it matters. FASEB J. 2013, 27, 1012–1022. [Google Scholar] [CrossRef]
- He, X.; Ou, H.Y.; Yu, Q.; Zhou, X.; Wu, J.; Liang, J.; Zhang, W.; Rajakumar, K.; Deng, Z. Analysis of a genomic island housing genes for DNA S-modification system in Streptomyces lividans 66 and its counterparts in other distantly related bacteria. Mol. Microbiol. 2007, 65, 1034–1048. [Google Scholar] [CrossRef]
- Yao, F.; Xu, T.; Zhou, X.; Deng, Z.; You, D. Functional analysis of spfD gene involved in DNA phosphorothioation in Pseudomonas fluorescens Pf0-1. FEBS Lett. 2009, 583, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zheng, X.; Cheng, Q.; Yao, F.; Zheng, T.; Ramesh Babu, I.; Zhou, H.; Dedon, P.; You, D. In vitro analysis of phosphorothioate modification of DNA reveals substrate recognition by a multiprotein complex. Sci. Rep. 2015, 5, 12513. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.Y.; He, X.; Shao, Y.; Tai, C.; Rajakumar, K.; Deng, Z. dndDB: A database focused on phosphorothioation of the DNA backbone. PLoS ONE 2009, 4, e5132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Jiang, P.; Cao, B.; Cheng, Q.; Kong, L.; Zheng, X.; Hu, Q.; You, D. DndEi Exhibits Helicase Activity Essential for DNA Phosphorothioate Modification and ATPase Activity Strongly Stimulated by DNA Substrate with a GAAC/GTTC Motif. J. Biol. Chem. 2016, 291, 1492–1500. [Google Scholar] [CrossRef]
- Chen, C.; Wang, L.; Chen, S.; Wu, X.; Gu, M.; Chen, X.; Jiang, S.; Wang, Y.; Deng, Z.; Dedon, P.C.; et al. Convergence of DNA methylation and phosphorothioation epigenetics in bacterial genomes. Proc. Natl. Acad. Sci. USA 2017, 114, 4501–4506. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.E. 2001 Fred Griffith review lecture. Immigration control of DNA in bacteria: Self versus non-self. Microbiology 2002, 148, 3–20. [Google Scholar] [CrossRef]
- Murray, N.E. Type I restriction systems: Sophisticated molecular machines (a legacy of Bertani and Weigle). Microbiol. Mol. Biol. Rev. 2000, 64, 412–434. [Google Scholar] [CrossRef]
- Ho, W.S.; Ou, H.Y.; Yeo, C.C.; Thong, K.L. The dnd operon for DNA phosphorothioation modification system in Escherichia coli is located in diverse genomic islands. BMC Genom. 2015, 16, 199. [Google Scholar] [CrossRef][Green Version]
- Zhang, Y.; Yakrus, M.A.; Graviss, E.A.; Williams-Bouyer, N.; Turenne, C.; Kabani, A.; Wallace, R.J., Jr. Pulsed-field gel electrophoresis study of Mycobacterium abscessus isolates previously affected by DNA degradation. J. Clin. Microbiol. 2004, 42, 5582–5587. [Google Scholar] [CrossRef][Green Version]
- Xie, X.; Liang, J.; Pu, T.; Xu, F.; Yao, F.; Yang, Y.; Zhao, Y.L.; You, D.; Zhou, X.; Deng, Z.; et al. Phosphorothioate DNA as an antioxidant in bacteria. Nucleic Acids Res. 2012, 40, 9115–9124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Parts | Queried by dnd Genes | Queried by ssp Genes |
|---|---|---|
| Gut system | Escherichia coli MS 45-1 | Dysgonomonas mossii DSM 22836 |
| Escherichia coli MS 117-3 | Alistipes indistinctus YIT 12060 | |
| Enterobacter cloacae NCTC 9394 | Prevotella HGA0225 | |
| Escherichia coli SE11 | Bacteroides dorei 5_1_36/D4 | |
| Lachnospiraceae bacterium 6_1_37FAA | Bacteroides 1_1_14 | |
| Desulfovibrio piger ATCC 29098 | Roseburia inulinivorans DSM 16841 | |
| Helicobacter bilis ATCC 43879 | Roseburia intestinalis M50/1 | |
| Paraprevotella xylaniphila YIT 11841 | Roseburia intestinalis L1-82 | |
| Lachnospiraceae bacterium 2_1_58FAA | Eubacterium ventriosum ATCC 27560 | |
| Clostridium asparagiforme DSM 15981 | Eubacterium siraeum V10Sc8a | |
| Peptoclostridium difficile 70-100-2010 | Megamonas funiformis YIT 11815 | |
| Bacteroides 2_1_33B | Mitsuokella multacida DSM 20544 | |
| Bacteroides xylanisolvens XB1A | Butyrivibrio fibrisolvens 16/4 | |
| Clostridium citroniae WAL-17108 | ||
| Faecalibacterium prausnitzii M21/2 | ||
| Fusobacterium necrophorum | ||
| funduliforme 1_1_36S | ||
| Oral system | Neisseria 020 | Prevotella salivae F0493 |
| Neisseria subflava NJ9703 N | Prevotella F0055 | |
| Neisseria bacilliformis ATCC BAA-1200 | Prevotella tannerae ATCC 51259 | |
| Eikenella corrodens ATCC 23834 | ||
| Lachnospiraceae bacterium F0431 | ||
| Selenomonas CM52 | ||
| Urogenital system | Prevotella amnii CRIS 21A-A | Prevotella oralis ATCC 33269 |
| Mycobacterium parascrofulaceum ATCC BAA-614 | Prevotella denticola CRIS 18C-A | |
| Prevotella bivia JCVIHMP010 | ||
| Skin | Acinetobacter baumannii 6014059 |
| PT-Linked Dinucleotides | Precursor Ion | Product Ion | z1 | z2 | z3 | 1488 | 1489 | 1494 | 1493 |
|---|---|---|---|---|---|---|---|---|---|
| d(CpsG) | 573 | 152 | Y | Y | N | ||||
| d(CpsC) | 533 | 112 | Y | Y | Y | Y | |||
| d(GpsG) | 613 | 152 | Y | Y | Y | Y | |||
| d(CpsA) | 557 | 136 | Y * | Y * | Y * | Y * | |||
| d(CpsT) | 548 | 112 | N ** | N ** | Y | N ** | N | Y | |
| d(ApsG) | 597 | 136 | Y | Y | |||||
| d(TpsG) | 588 | 152 | Y | Y | Y | Y | |||
| d(GpsA) | 597 | 136 | Y | Y | |||||
| d(GpsC) | 573 | 112 | N | Y | N | Y | |||
| d(GpsT) | 588 | 152 | Y | N | Y | Y | N | Y | |
| d(ApsA) | 581 | 136 | |||||||
| d(TpsA) | 572 | 136 | Y | Y | N | ||||
| d(ApsC) | 557 | 112 | Y * | Y * | Y * | Y * | |||
| d(TpsC) | 548 | 112 | N ** | Y | N | Y | |||
| d(ApsT) | 572 | 136 | Y | Y | Y | Y | N | ||
| d(TpsT) | 563 | 127 | |||||||
| d(GpsA) Sp | 597 | 136 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Kong, L.; Wu, G.; Cao, B.; Pang, X.; Deng, Z.; Dedon, P.C.; Zhang, C.; You, D. DNA Phosphorothioate Modifications Are Widely Distributed in the Human Microbiome. Biomolecules 2020, 10, 1175. https://doi.org/10.3390/biom10081175
Sun Y, Kong L, Wu G, Cao B, Pang X, Deng Z, Dedon PC, Zhang C, You D. DNA Phosphorothioate Modifications Are Widely Distributed in the Human Microbiome. Biomolecules. 2020; 10(8):1175. https://doi.org/10.3390/biom10081175
Chicago/Turabian StyleSun, Yihua, Lingxin Kong, Guojun Wu, Bo Cao, Xiaoyan Pang, Zixin Deng, Peter C. Dedon, Chenhong Zhang, and Delin You. 2020. "DNA Phosphorothioate Modifications Are Widely Distributed in the Human Microbiome" Biomolecules 10, no. 8: 1175. https://doi.org/10.3390/biom10081175
APA StyleSun, Y., Kong, L., Wu, G., Cao, B., Pang, X., Deng, Z., Dedon, P. C., Zhang, C., & You, D. (2020). DNA Phosphorothioate Modifications Are Widely Distributed in the Human Microbiome. Biomolecules, 10(8), 1175. https://doi.org/10.3390/biom10081175