A Computational Approach with Biological Evaluation: Combinatorial Treatment of Curcumin and Exemestane Synergistically Regulates DDX3 Expression in Cancer Cell Lines

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Pharmacophore Generation
2.2. Common Feature Pharmacophore Generation
2.3. Receptor Based Pharmacophore Generation
2.4. Validation of the Pharmacophore Models
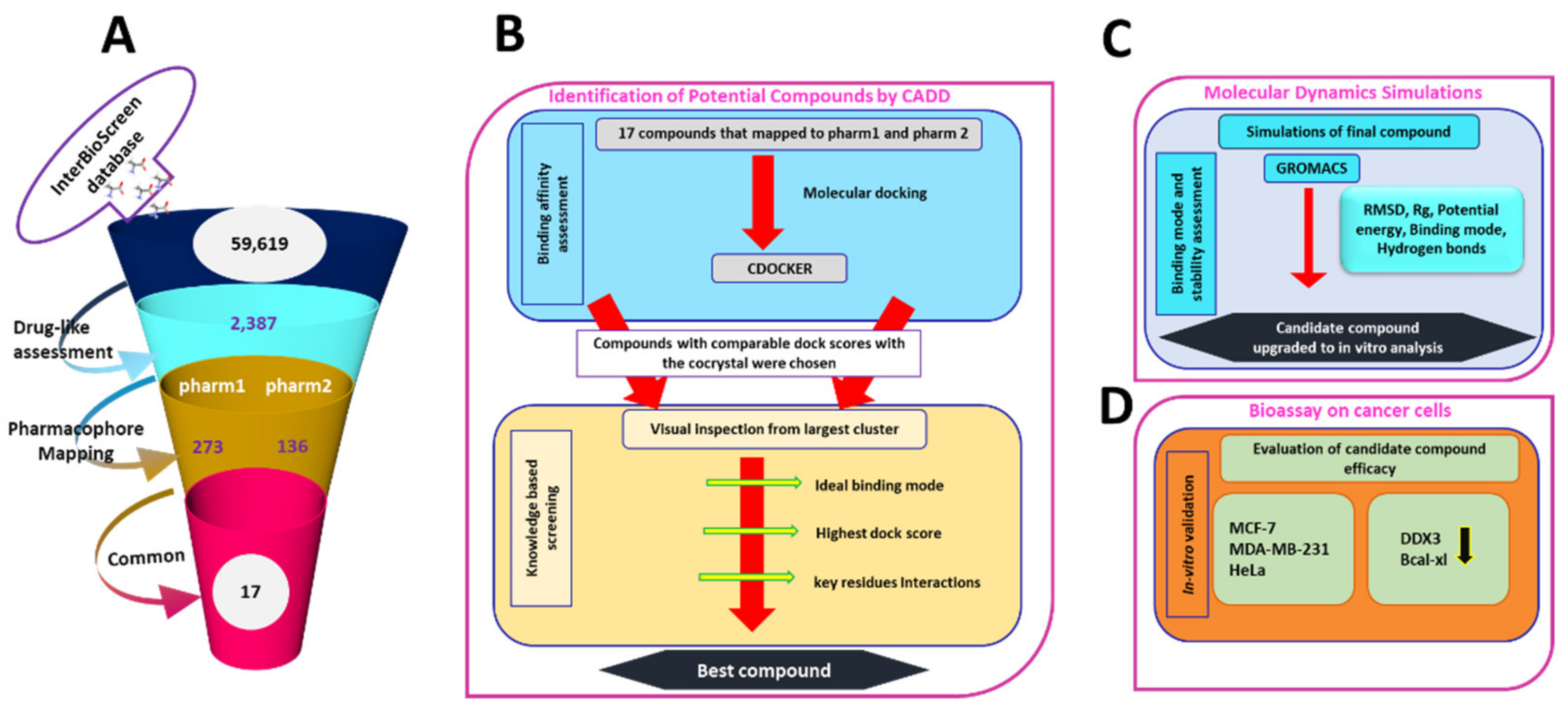
2.5. Drug-Like Database Formulation from InterBioScreen Database
2.6. Virtual Screening of InterBioScreen Database Using Pharm1 and Pharm2
2.7. Molecular Docking Studies
2.8. Molecular Dynamics Simulation Studies
2.9. In Vitro Bioassay Validation
2.9.1. Procurement of Cell Lines and Culture
2.9.2. Procurement of Compounds and Reagent
2.10. Cell Viability Assay
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. Pharmacophore Generation
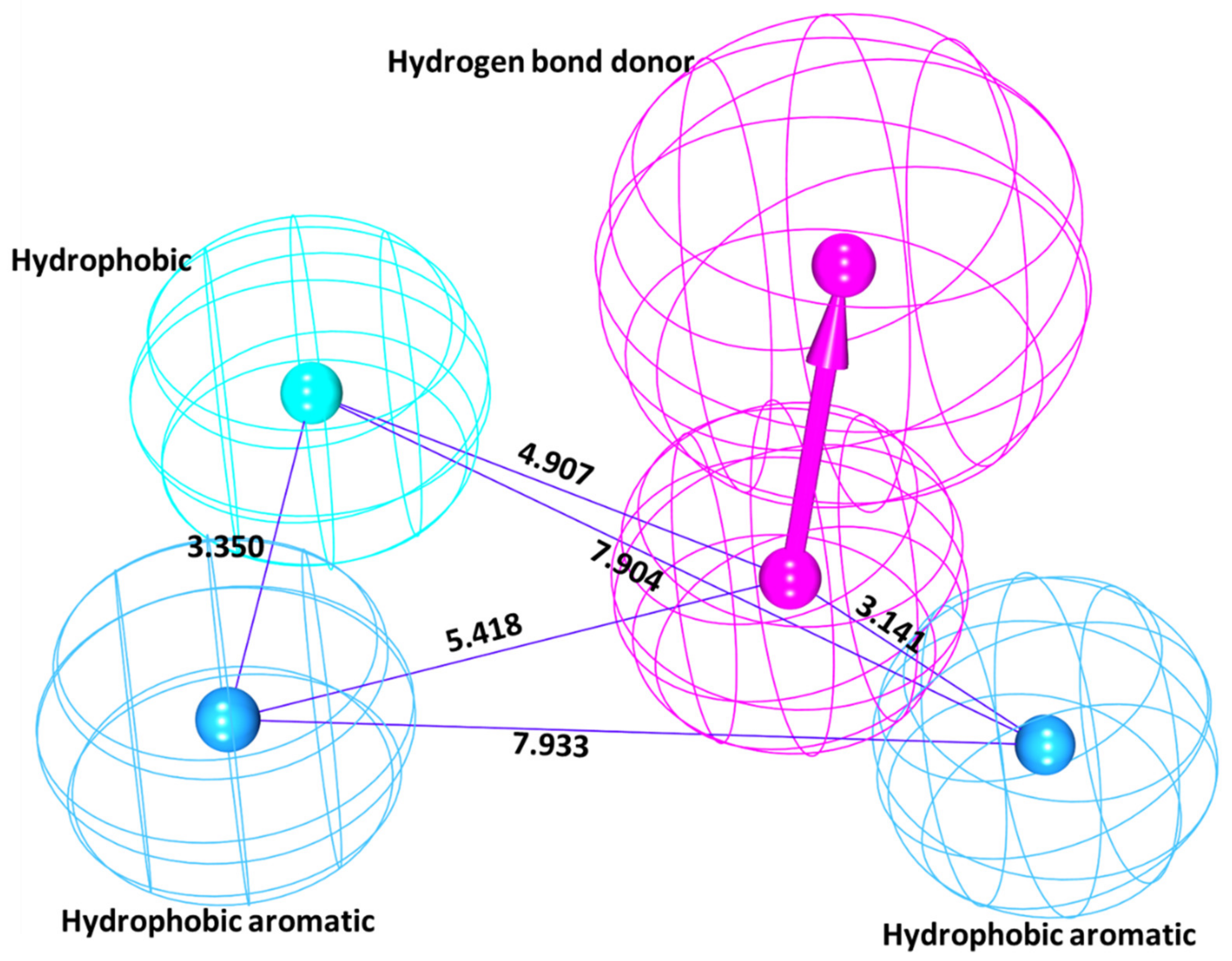
3.1.1. Common Feature Pharmacophore Generation
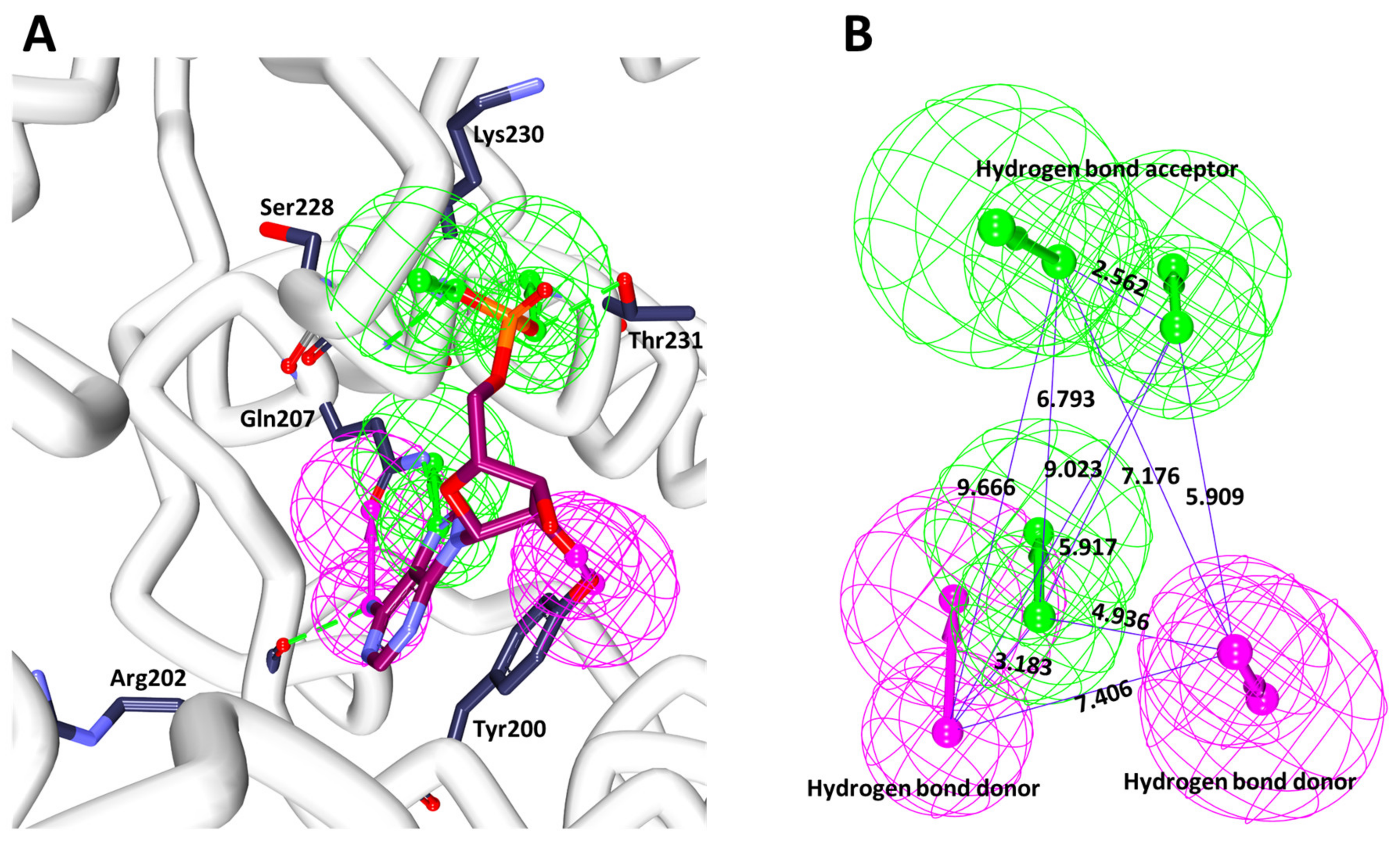
3.1.2. Receptor-Based Pharmacophore Generation
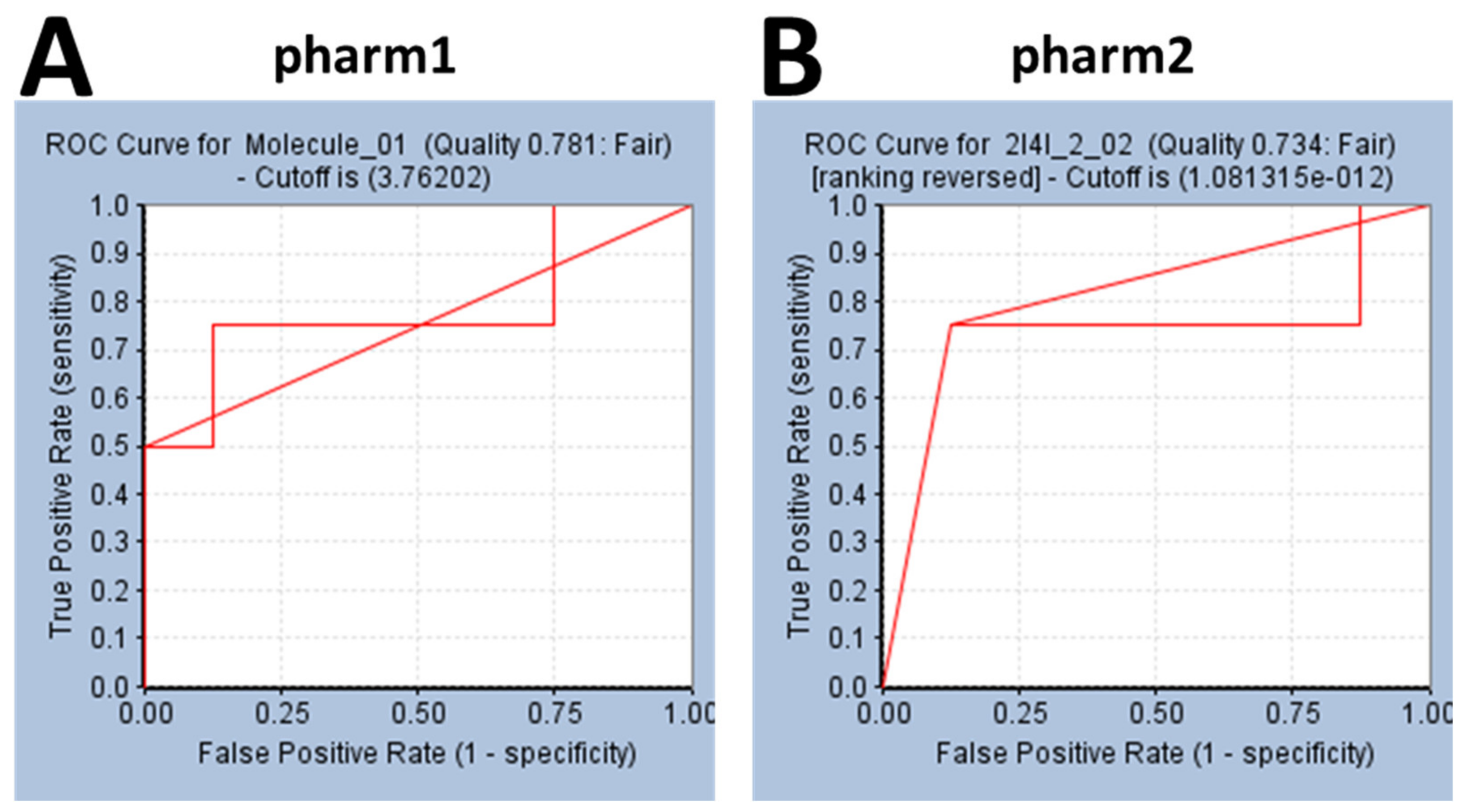
3.2. Validation of the Pharmacophore Models
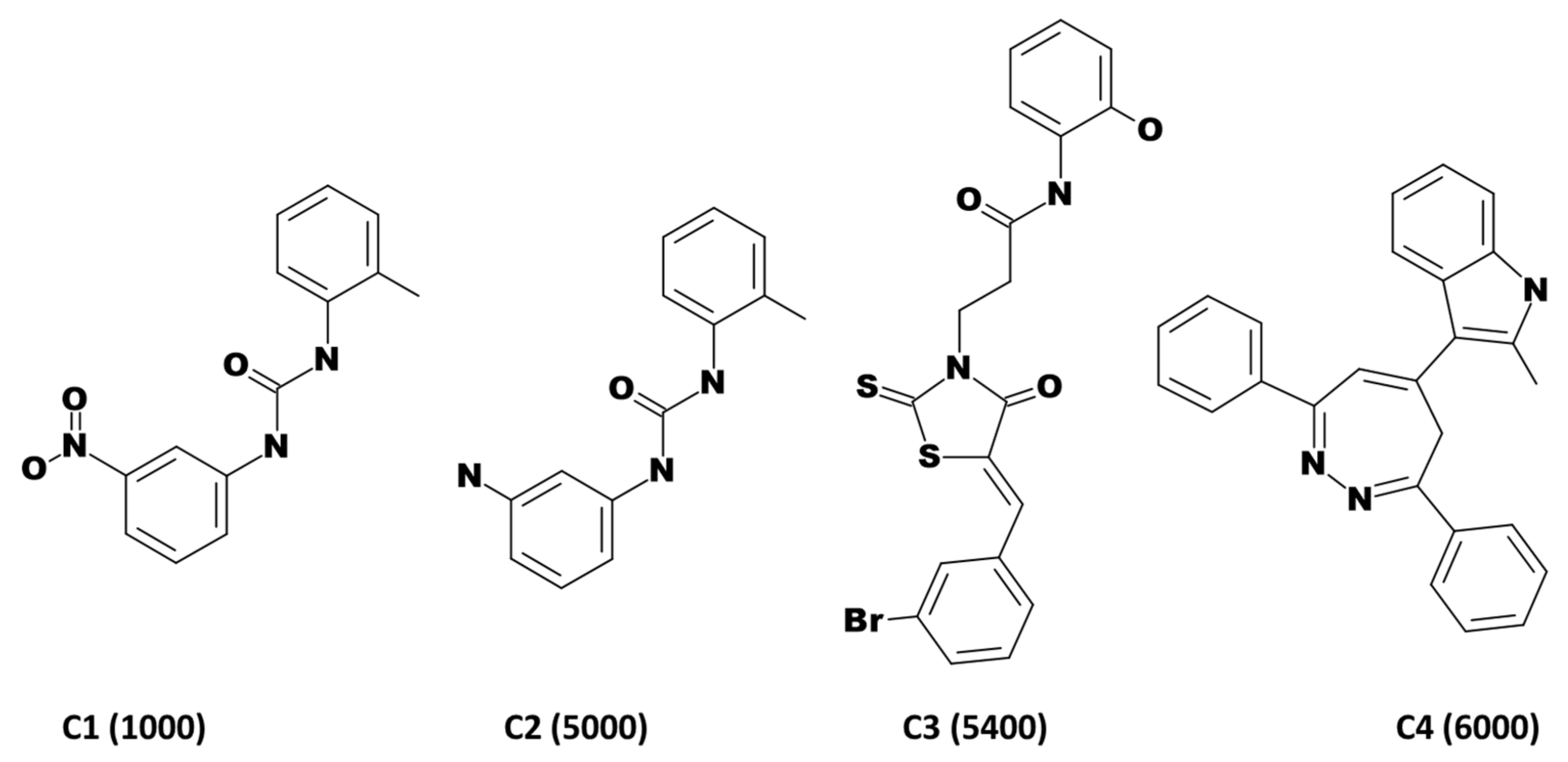
3.3. Virtual Screening of InterBioScreen Database
3.4. Molecular Docking Studies
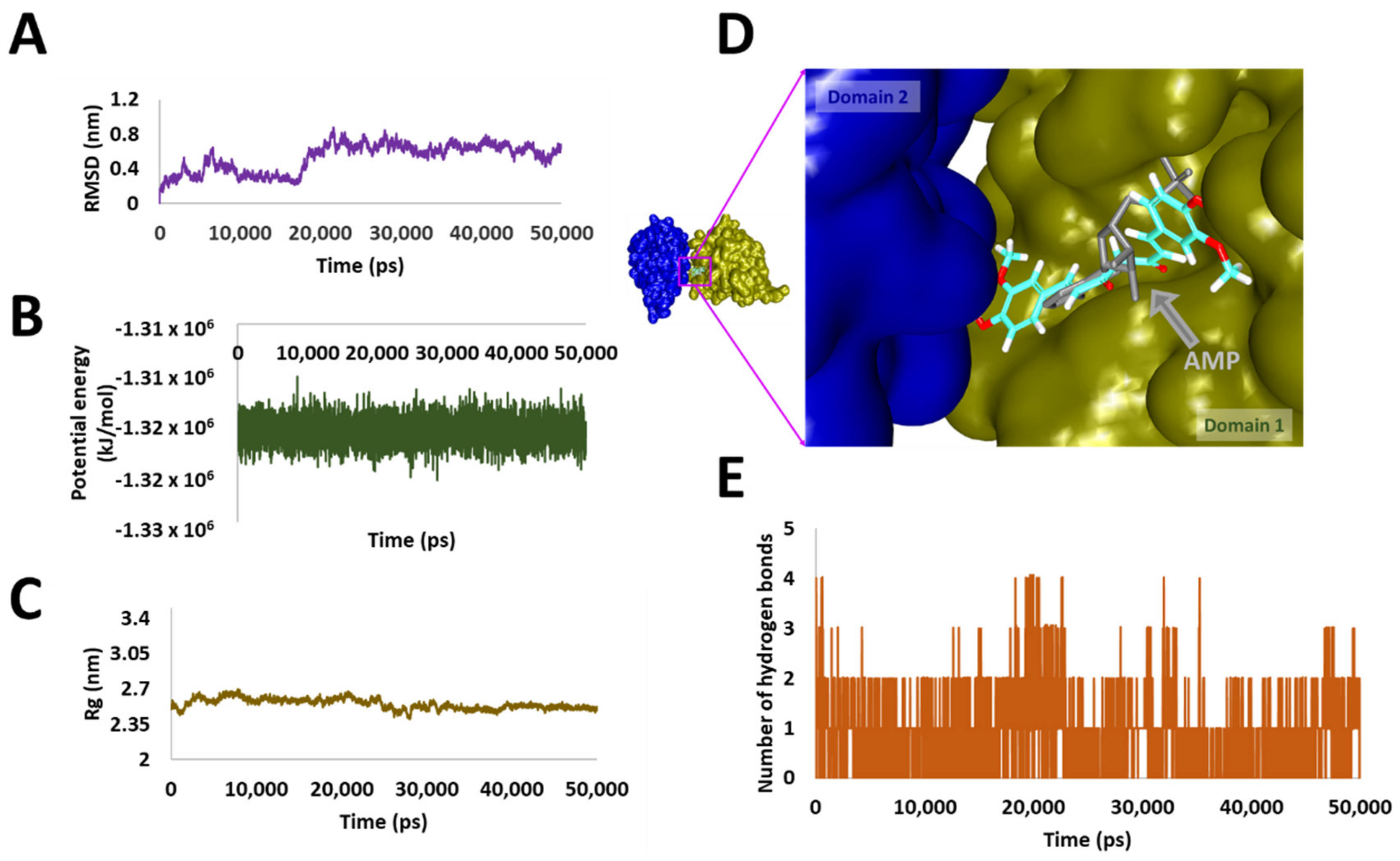
3.5. Molecular Dynamics Simulation Analysis
3.6. Stability Analysis
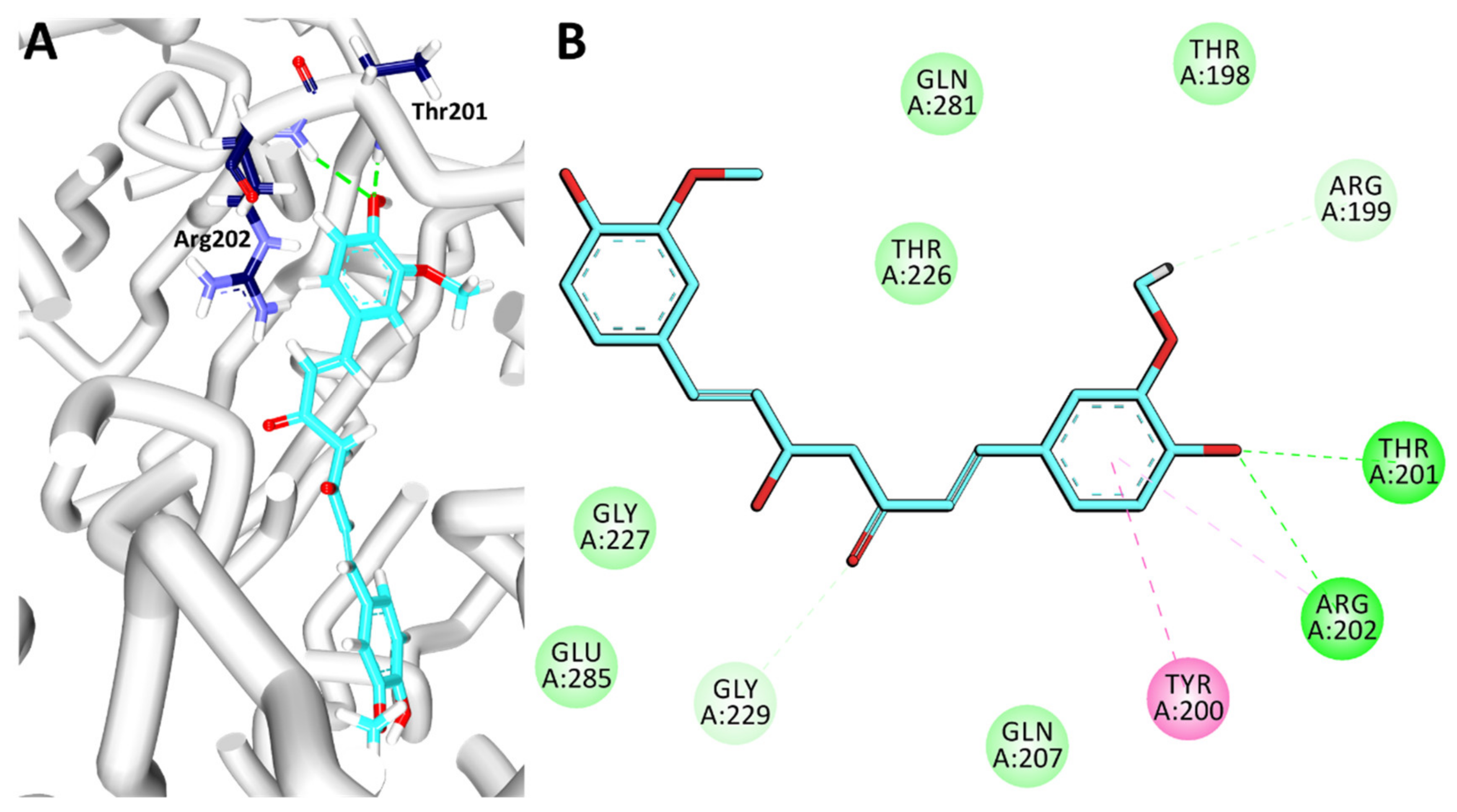
3.7. Binding Mode Analysis and Intermolecular Interactions
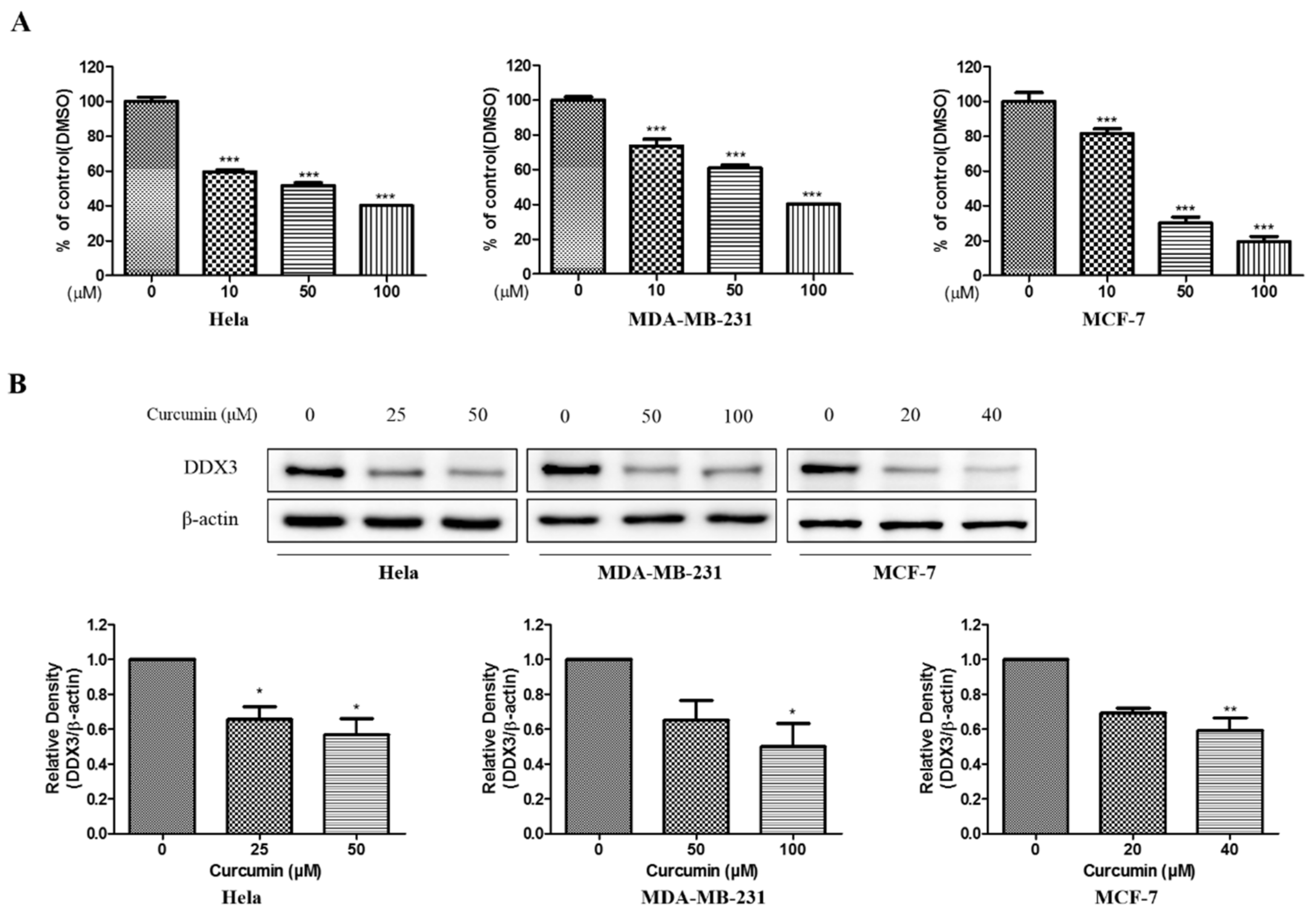
3.8. Bioassay Validation of Curcumin as Potential DDX3 Inhibitor
3.8.1. Anti-Proliferative Effect and Inhibition of DDX3 Protein Expression of Curcumin
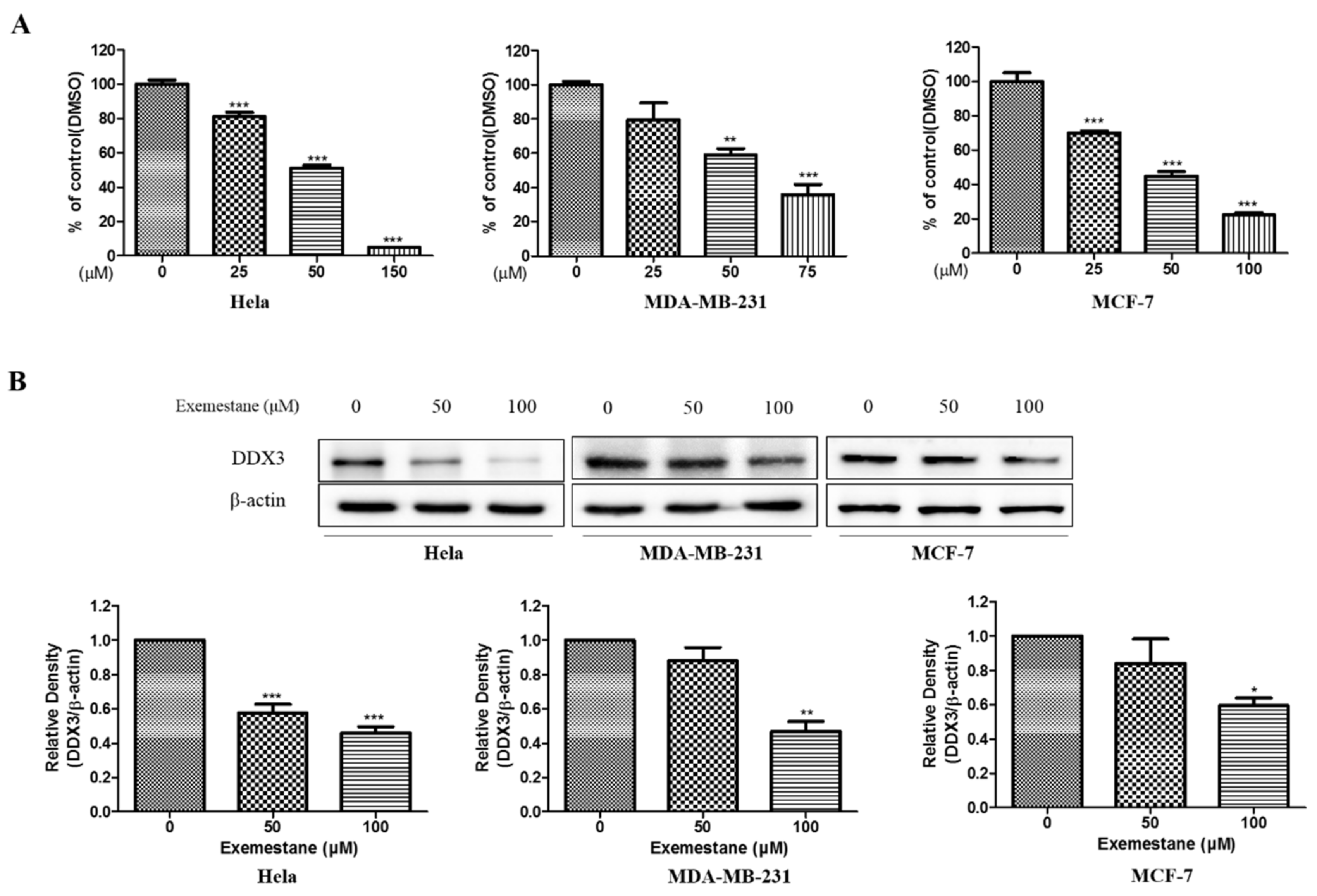
3.8.2. Anti-Proliferative Effect and Inhibition of DDX3 Protein Level on Exemestane
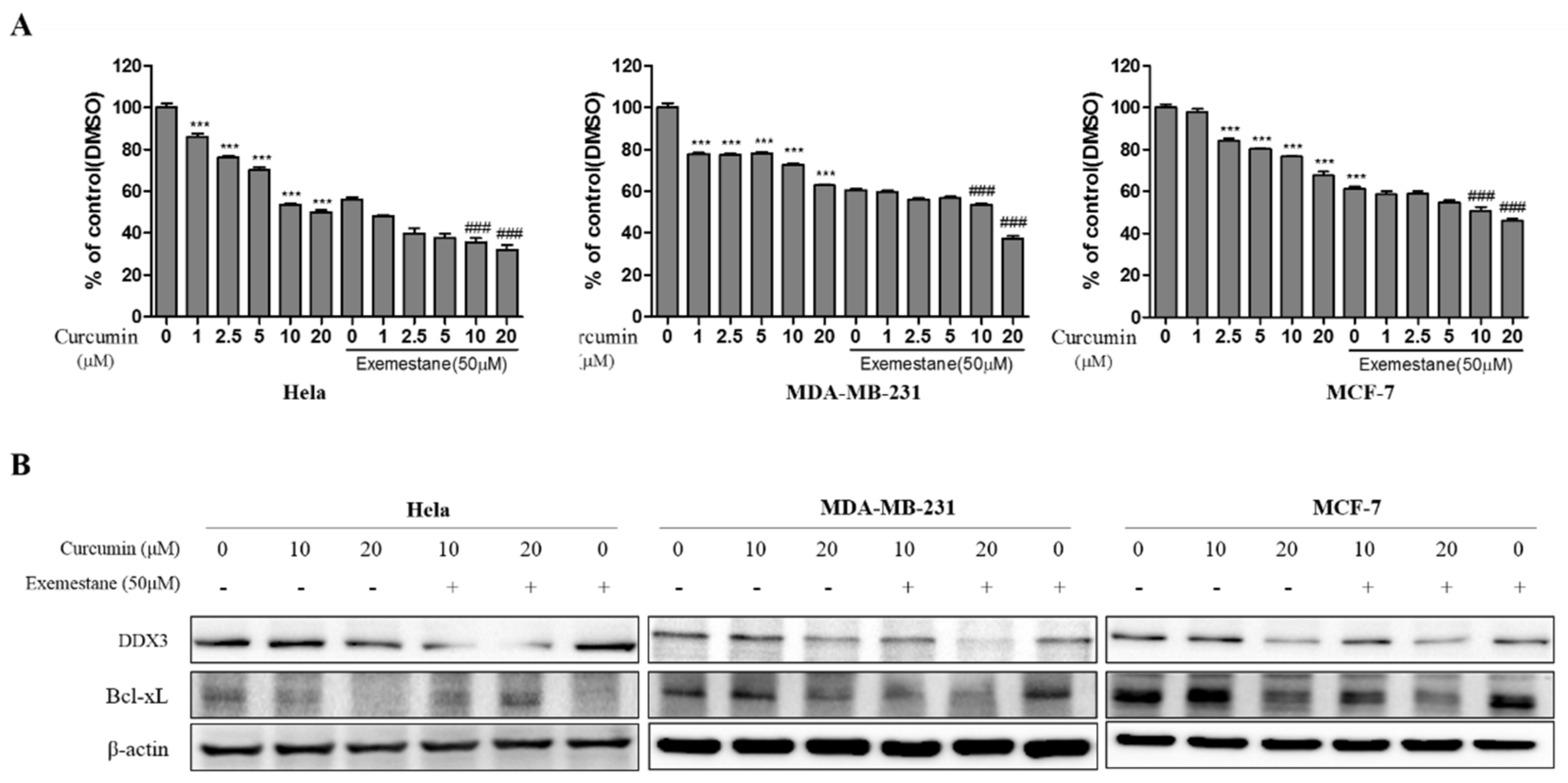
3.9. Exemestane is Synergistic with Curcumin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Van Heerma Voss, M.R.; Vesuna, F.; Bol, G.M.; Meeldijk, J.; Raman, A.; Offerhaus, G.J.; Buerger, H.; Patel, A.H.; van der Wall, E.; van Diest, P.J.; et al. Nuclear DDX3 expression predicts poor outcome in colorectal and breast cancer. Onco. Targets. Ther. 2017, 10, 3501–3513. [Google Scholar] [CrossRef]
- Bol, G.M.; Xie, M.; Raman, V. DDX3, a potential target for cancer treatment. Mol. Cancer 2015, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Abdelhaleem, M. RNA helicases: Regulators of differentiation. Clin. Biochem. 2005, 38, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rifo, R.; Ohlmann, T. The role of the DEAD-box RNA helicase DDX3 in mRNA metabolism. Wiley Interdiscip. Rev. RNA 2013, 4, 369–385. [Google Scholar] [CrossRef]
- Yedavalli, V.S.R.K.; Neuveut, C.; Chi, Y.H.; Kleiman, L.; Jeang, K.T. Requirement of DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell 2004, 119, 381–392. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, S.-G.; Kim, Y.; Song, K. Assignment1 of a human putative RNA helicase gene, DDX3, to human X chromosome bands p11.3→p11.23. Cytogenet. Genome Res. 1998, 81, 178–179. [Google Scholar] [CrossRef] [PubMed]
- Fullam, A.; Schröder, M. DExD/H-box RNA helicases as mediators of anti-viral innate immunity and essential host factors for viral replication. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Heerma van Voss, M.R.; Kammers, K.; Vesuna, F.; Brilliant, J.; Bergman, Y.; Tantravedi, S.; Wu, X.; Cole, R.N.; Holland, A.; van Diest, P.J.; et al. Global Effects of DDX3 Inhibition on Cell Cycle Regulation Identified by a Combined Phosphoproteomics and Single Cell Tracking Approach. Transl. Oncol. 2018, 11, 755–763. [Google Scholar] [CrossRef]
- Botlagunta, M.; Vesuna, F.; Mironchik, Y.; Raman, A.; Lisok, A.; Winnard, P.; Mukadam, S.; Van Diest, P.; Chen, J.H.; Farabaugh, P.; et al. Oncogenic role of DDX3 in breast cancer biogenesis. Oncogene 2008, 27, 3912–3922. [Google Scholar] [CrossRef]
- Sun, M.; Song, L.; Zhou, T.; Gillespie, G.Y.; Jope, R.S. The role of DDX3 in regulating Snail. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 438–447. [Google Scholar] [CrossRef]
- Heerma van Voss, M.R.; Vesuna, F.; Bol, G.M.; Afzal, J.; Tantravedi, S.; Bergman, Y.; Kammers, K.; Lehar, M.; Malek, R.; Ballew, M.; et al. Targeting mitochondrial translation by inhibiting DDX3: A novel radiosensitization strategy for cancer treatment. Oncogene 2018, 37, 63. [Google Scholar] [CrossRef]
- Chen, H.H.; Yu, H.I.; Cho, W.C.; Tarn, W.Y. DDX3 modulates cell adhesion and motility and cancer cell metastasis via Rac1-mediated signaling pathway. Oncogene 2015, 34, 2790–2800. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.-C.; Chang, W.-C.; Shieh, S.-Y.; Tarn, W.-Y. DDX3 Regulates Cell Growth through Translational Control of Cyclin E1. Mol. Cell. Biol. 2010, 30, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Bol, G.M.; Vesuna, F.; Xie, M.; Zeng, J.; Aziz, K.; Gandhi, N.; Levine, A.; Irving, A.; Korz, D.; Tantravedi, S.; et al. Targeting DDX3 with a small molecule inhibitor for lung cancer therapy. EMBO Mol. Med. 2015, 7, 648–669. [Google Scholar] [CrossRef] [PubMed]
- Högbom, M.; Collins, R.; van den Berg, S.; Jenvert, R.M.; Karlberg, T.; Kotenyova, T.; Flores, A.; Hedestam, G.B.K.; Schiavone, L.H. Crystal Structure of Conserved Domains 1 and 2 of the Human DEAD-box Helicase DDX3X in Complex with the Mononucleotide AMP. J. Mol. Biol. 2007, 372, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kukhanova, M.K.; Karpenko, I.L.; Ivanov, A.V. DEAD-box RNA helicase DDX3: Functional properties and development of DDX3 inhibitors as antiviral and anticancer drugs. Molecules 2020, 25, 1015. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.N.Y.; Atkinson, S.C.; Audsley, M.D.; Heaton, S.M.; Jans, D.A.; Borg, N.A. RK-33 is a Broad-Spectrum Antiviral Agent that Targets DEAD-Box RNA Helicase DDX3X. Cells 2020, 9, 170. [Google Scholar] [CrossRef]
- Wilky, B.A.; Kim, C.; McCarty, G.; Montgomery, E.A.; Kammers, K.; Devine, L.R.; Cole, R.N.; Raman, V.; Loeb, D.M. RNA helicase DDX3: A novel therapeutic target in Ewing sarcoma. Oncogene 2016, 35, 2574–2583. [Google Scholar] [CrossRef]
- Xie, M.; Vesuna, F.; Tantravedi, S.; Bol, G.M.; Van Voss, M.R.H.; Nugent, K.; Malek, R.; Gabrielson, K.; Van Diest, P.J.; Tran, P.T.; et al. RK-33 radiosensitizes prostate cancer cells by blocking the RNA helicase DDX3. Cancer Res. 2016, 76, 6340–6350. [Google Scholar] [CrossRef]
- Samal, S.K.; Routray, S.; Veeramachaneni, G.K.; Dash, R.; Botlagunta, M. Ketorolac salt is a newly discovered DDX3 inhibitor to treat oral cancer. Sci. Rep. 2015. [Google Scholar] [CrossRef]
- Xie, M.; Vesuna, F.; Botlagunta, M.; Bol, G.M.; Irving, A.; Bergman, Y.; Hosmane, R.S.; Kato, Y.; Winnard, P.T.; Raman, V. NZ51, a ring-expanded nucleoside analog, inhibits motility and viability of breast cancer cells by targeting the RNA helicase DDX3. Oncotarget 2015, 6, 29901–29913. [Google Scholar] [CrossRef] [PubMed]
- Heerma van Voss, M.R.; Schrijver, W.A.M.E.; ter Hoeve, N.D.; Hoefnagel, L.D.; Manson, Q.F.; van der Wall, E.; Raman, V.; van Diest, P.J. The prognostic effect of DDX3 upregulation in distant breast cancer metastases. Clin. Exp. Metastasis 2017, 34, 85–92. [Google Scholar] [CrossRef] [PubMed]
- He, T.Y.; Wu, D.W.; Lin, P.L.; Wang, L.; Huang, C.C.; Chou, M.C.; Lee, H. DDX3 promotes tumor invasion in colorectal cancer via the CK1ε/Dvl2 axis. Sci. Rep. 2016, 6, 21483. [Google Scholar] [CrossRef] [PubMed]
- Van Voss, M.R.H.; Vesuna, F.; Trumpi, K.; Brilliant, J.; Berlinicke, C.; de Leng, W.; Kranenburg, O.; Offerhaus, G.J.; Bürger, H.; van der Wall, E.; et al. Identification of the DEAD box RNA helicase DDX3 as a therapeutic target in colorectal cancer. Oncotarget 2015, 6, 28312–28326. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-S.; Zhao, Z.; Yang, Z.-N.; Xu, F.; Lu, H.-J.; Zhu, Z.-Y.; Shi, W.; Jiang, J.; Yao, P.-P.; Zhu, H.-P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef]
- Bol, G.M.; Raman, V.; van der Groep, P.; Vermeulen, J.F.; Patel, A.H.; van der Wall, E.; van Diest, P.J. Expression of the RNA Helicase DDX3 and the Hypoxia Response in Breast Cancer. PLoS ONE 2013, 8, e63548. [Google Scholar] [CrossRef]
- Lim, M.C.; Lee, M.; Shim, S.H.; Nam, E.J.; Lee, J.Y.; Kim, H.J.; Lee, Y.Y.; Lee, K.B.; Park, J.Y.; Kim, Y.H.; et al. Practice guidelines for management of cervical cancer in Korea: A Korean Society of Gynecologic Oncology Consensus Statement. J. Gynecol. Oncol. 2017, 28, e22. [Google Scholar] [CrossRef]
- Chen, X.; Lin, Y.; Gilson, M.K. The binding database: Overview and user’s guide. Biopolymers 2001, 61, 127–141. [Google Scholar] [CrossRef]
- Barnum, D.; Greene, J.; Smellie, A.; Sprague, P. Identification of common functional configurations among molecules. J. Chem. Inf. Comput. Sci. 1996, 36, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Gilson, M.K.; Liu, T.; Baitaluk, M.; Nicola, G.; Hwang, L.; Chong, J. BindingDB in 2015: A public database for medicinal chemistry, computational chemistry and systems pharmacology. Nucleic Acids Res. 2016, 44, D1045–D1053. [Google Scholar] [CrossRef]
- Chen, X.; Lin, Y.; Liu, M.; Gilson, M.K. The binding database: Data management and interface design. Bioinformatics 2002, 18, 130–139. [Google Scholar] [CrossRef]
- Meslamani, J.; Li, J.; Sutter, J.; Stevens, A.; Bertrand, H.O.; Rognan, D. Protein-ligand-based pharmacophores: Generation and utility assessment in computational ligand profiling. J. Chem. Inf. Model. 2012, 52, 943–955. [Google Scholar] [CrossRef]
- Padariya, M.; Kalathiya, U.; Baginski, M. Docking simulations, molecular properties and admet studies of novel chromane-6,7-diol analogues as potential inhibitors of mushroom tyrosinase. Gene Ther. Mol. Biol. 2014, 16, 201–218. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2012, 64, 4–17. [Google Scholar] [CrossRef]
- Wu, G.; Robertson, D.H.; Brooks, C.L.; Vieth, M. Detailed analysis of grid-based molecular docking: A case study of CDOCKER—A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef] [PubMed]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Rampogu, S.; Lee, G.; Ravinder, D.; Woo Lee, K. Short communication for targeting natural compounds against HER2 kinase domain as potential anticancer drugs applying pharmacophore based molecular modelling approaches—Part 2. Comput. Biol. Chem. 2020, 87, 107242. [Google Scholar] [CrossRef]
- Zoete, V.; Cuendet, M.A.; Grosdidier, A.; Michielin, O. SwissParam: A fast force field generation tool for small organic molecules. J. Comput. Chem. 2011, 32, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Morris, G.M.; Lim-Wilby, M. Molecular docking. Methods Mol. Biol. 2008, 443, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided. Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K.K. Various types and management of breast cancer: An overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar] [PubMed]
- Akram, M.; Iqbal, M.; Daniyal, M.; Khan, A.U. Awareness and current knowledge of breast cancer. Biol. Res. 2017, 50, 33. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Tong, C.W.S.; Wu, M.; Cho, W.C.S.; To, K.K.W. Recent Advances in the Treatment of Breast Cancer. Front. Oncol. 2018, 8, 227. [Google Scholar] [CrossRef]
- Ahmad, A. Pathways to breast cancer recurrence. ISRN Oncol. 2013, 2013, 290568. [Google Scholar] [CrossRef]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- McGraw, S.L.; Ferrante, J.M. Update on prevention and screening of cervical cancer. World J. Clin. Oncol. 2014, 5, 744–752. [Google Scholar] [CrossRef]
- He, Y.; Zhang, D.; Yang, Y.; Wang, X.; Zhao, X.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. A double-edged function of DDX3, as an oncogene or tumor suppressor, in cancer progression (Review). Oncol. Rep. 2018, 39, 883–892. [Google Scholar] [CrossRef]
- Tantravedi, S.; Vesuna, F.; Winnard, P.T., Jr.; Martin, A.; Lim, M.; Eberhart, C.G.; Berlinicke, C.; Raabe, E.; van Diest, P.J.; Raman, V. Targeting DDX3 in Medulloblastoma Using the Small Molecule Inhibitor RK-33. Transl. Oncol. 2019, 12, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Botlagunta, M.; Kollapalli, B.; Kakarla, L.; Gajarla, S.P.; Gade, S.P.; Dadi, C.L.; Penumadu, A.; Javeed, S. In vitro anti-cancer activity of doxorubicin against human RNA helicase, DDX3. Bioinformation 2016, 12, 347–353. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Radi, M.; Falchi, F.; Garbelli, A.; Samuele, A.; Bernardo, V.; Paolucci, S.; Baldanti, F.; Schenone, S.; Manetti, F.; Maga, G.; et al. Discovery of the first small molecule inhibitor of human DDX3 specifically designed to target the RNA binding site: Towards the next generation HIV-1 inhibitors. Bioorganic Med. Chem. Lett. 2012, 22, 2094–2098. [Google Scholar] [CrossRef] [PubMed]
- Brai, A.; Ronzini, S.; Riva, V.; Botta, L.; Zamperini, C.; Borgini, M.; Trivisani, C.I.; Garbelli, A.; Pennisi, C.; Boccuto, A.; et al. Synthesis and Antiviral Activity of Novel 1,3,4-Thiadiazole Inhibitors of DDX3X. Molecules 2019, 24, 3988. [Google Scholar] [CrossRef]
- Fu, R.; Yang, P.; Li, Z.; Liu, W.; Amin, S.; Li, Z. Avenanthramide A triggers potent ROS-mediated anti-tumor effects in colorectal cancer by directly targeting DDX3. Cell Death Dis. 2019, 10, 593. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Nagesh, P.K.B.; Jaggi, M.; Chauhan, S.C. Therapeutic Applications of Curcumin Nanoformulations. AAPS J. 2015, 17, 1341–1356. [Google Scholar] [CrossRef]
- Suresh, S.; Yadav, V.R.; Suresh, A. Health Benefits and Therapeutic Applications of Curcumin. Clin. Res. Regul. Aff. 2006, 23, 191–210. [Google Scholar] [CrossRef]
- Mimeault, M.; Batra, S.K. Potential applications of curcumin and its novel synthetic analogs and nanotechnology-based formulations in cancer prevention and therapy. Chin. Med. 2011, 6, 31. [Google Scholar] [CrossRef]
- Bachmeier, E.B.; Melchart, D. Therapeutic Effects of Curcumin—From Traditional Past to Present and Future Clinical Applications. Int. J. Mol. Sci. 2019, 20, 3757. [Google Scholar] [CrossRef]
- Liu, H.-T.; Ho, Y.-S. Anticancer effect of curcumin on breast cancer and stem cells. Food Sci. Hum. Wellness 2018, 7, 134–137. [Google Scholar] [CrossRef]
- Wang, X.; Hang, Y.; Liu, J.; Hou, Y.; Wang, N.; Wang, M. Anticancer effect of curcumin inhibits cell growth through miR-21/PTEN/Akt pathway in breast cancer cell. Oncol. Lett. 2017, 13, 4825–4831. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, P. Exemestane, a new steroidal aromatase inhibitor of clinical relevance. Biochim. Biophys. Acta Mol. Basis Dis. 2002, 1587, 326–337. [Google Scholar] [CrossRef]
- Miller, W.R.; Bartlett, J.; Brodie, A.M.H.; Brueggemeier, R.W.; di Salle, E.; Lonning, P.E.; Llombart, A.; Maass, N.; Maudelonde, T.; Sasano, H.; et al. Aromatase Inhibitors: Are There Differences Between Steroidal and Nonsteroidal Aromatase Inhibitors and Do They Matter? Oncologist 2008, 13, 829–837. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No. | SMILES | IC50 nM | ChEMBL ID |
|---|---|---|---|
| Compound 1 | Oc1cccc(NC(=O)CCN2C(=S)S\C(=C/c3cccc(Br)c3)\C2=O)c1 | 90,000 | CHEMBL457233 |
| Compound 2 | Oc1cccc(NC(=O)CCCN2C(=S)S\C(=C/c3cccc(Br)c3)\C2=O)c1 | 150,000 | CHEMBL456405 |
| Compound 3 | COc1ccccc1\C=C\2/SC(=S)N(CCC(=O)Nc3cccc(O)c3)C2=O | 200,000 | CHEMBL514760 |
| Compound 4 | OC(=O)c1ccccc1NC(=O)CCN2C(=S)S\C(=C/c3cccc(Br)c3)\C2=O | 200,000 | CHEMBL484400 |
| Compound 5 | Nc1c(C#N)c(nn1c2ccccc2)\C(=C\c3oc(cc3)c4ccccc4[N+](=O)[O-])\C#N | 300,000 | CHEMBL484749 |
| Compound 6 | Oc1ccc(NC(=O)CCN2C(=S)S\C(=C/c3cccc(Br)c3)\C2=O)cc1 | 300,000 | CHEMBL515669 |
| Compound 7 | Nc1c(C#N)c(nn1c2ccccc2)\C(=C\c3oc(cc3)c4cccc(Cl)c4)\C#N | 500,000 | CHEMBL500746 |
| Compound 8 | Oc1ccccc1NC(=O)CCCN2C(=S)S\C(=C/c3cccc(Br)c3)\C2=O | 500,000 | CHEMBL459179 |
| Model No. | Features * | Rank | Direct Hit | Partial Hit | Max Fit |
|---|---|---|---|---|---|
| 01 | 2HA,HYP,HBD | 31.233 | 1111 | 0000 | 4 |
| 02 | HA,HYP,HBD,RA | 30.714 | 1111 | 0000 | 4 |
| 03 | HA,HYP,HBD,RA | 30.629 | 1111 | 0000 | 4 |
| 04 | HA,2HYP,HBD | 28.033 | 1111 | 0000 | 4 |
| 05 | HA,2HYP,HBD | 28.033 | 1111 | 0000 | 4 |
| 06 | 2HA,HBD | 27.918 | 1111 | 0000 | 3 |
| 07 | 2HYP,HBD,RA | 27.514 | 1111 | 0000 | 4 |
| 08 | HA,HBD,RA | 27.470 | 1111 | 0000 | 3 |
| 09 | HA,HBD,RA | 27.438 | 1111 | 0000 | 3 |
| 10 | 2HYP,HBD,RA | 27.429 | 1111 | 0000 | 4 |
| Model No. | Number of Features | Feature Set * |
|---|---|---|
| 01 | 5 | HBA,HBA,HBD,HBD,RA |
| 02 | 5 | HBA,HBA,HBA,HBD,HBD |
| 03 | 5 | HBA,HBA,HBD,HBD,RA |
| 04 | 5 | HBA,HBA,HBA,HBD,HBD |
| 05 | 5 | HBA,HBA,HBD,HBD,RA |
| 06 | 5 | HBA,HBA,HBD,HBD,RA |
| 07 | 5 | HBA,HBA,HBD,HBD,RA |
| 08 | 5 | HBA,HBA,HBD,HBD,RA |
| 09 | 5 | HBA,HBA,HBD,HBD,RA |
| 10 | 5 | HBA,HBA,HBD,HBD,RA |
| Compound | Hydrogen Bonds | π-π Stacked | van der Waals Interactions |
|---|---|---|---|
| Curcumin | Thr201: HN-O3 (2.2 Å) | Tyr200 | Thr198, Gln207, Thr226, Gly227, Gly229, Gln281, Glu285 |
| Arg202: HN-O3 (2.3 Å) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rampogu, S.; Kim, S.M.; Son, M.; Baek, A.; Park, C.; Lee, G.; Kim, Y.; Kim, G.S.; Kim, J.H.; Lee, K.W. A Computational Approach with Biological Evaluation: Combinatorial Treatment of Curcumin and Exemestane Synergistically Regulates DDX3 Expression in Cancer Cell Lines. Biomolecules 2020, 10, 857. https://doi.org/10.3390/biom10060857
Rampogu S, Kim SM, Son M, Baek A, Park C, Lee G, Kim Y, Kim GS, Kim JH, Lee KW. A Computational Approach with Biological Evaluation: Combinatorial Treatment of Curcumin and Exemestane Synergistically Regulates DDX3 Expression in Cancer Cell Lines. Biomolecules. 2020; 10(6):857. https://doi.org/10.3390/biom10060857
Chicago/Turabian StyleRampogu, Shailima, Seong Min Kim, Minky Son, Ayoung Baek, Chanin Park, Gihwan Lee, Yumi Kim, Gon Sup Kim, Ju Hyun Kim, and Keun Woo Lee. 2020. "A Computational Approach with Biological Evaluation: Combinatorial Treatment of Curcumin and Exemestane Synergistically Regulates DDX3 Expression in Cancer Cell Lines" Biomolecules 10, no. 6: 857. https://doi.org/10.3390/biom10060857
APA StyleRampogu, S., Kim, S. M., Son, M., Baek, A., Park, C., Lee, G., Kim, Y., Kim, G. S., Kim, J. H., & Lee, K. W. (2020). A Computational Approach with Biological Evaluation: Combinatorial Treatment of Curcumin and Exemestane Synergistically Regulates DDX3 Expression in Cancer Cell Lines. Biomolecules, 10(6), 857. https://doi.org/10.3390/biom10060857