Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
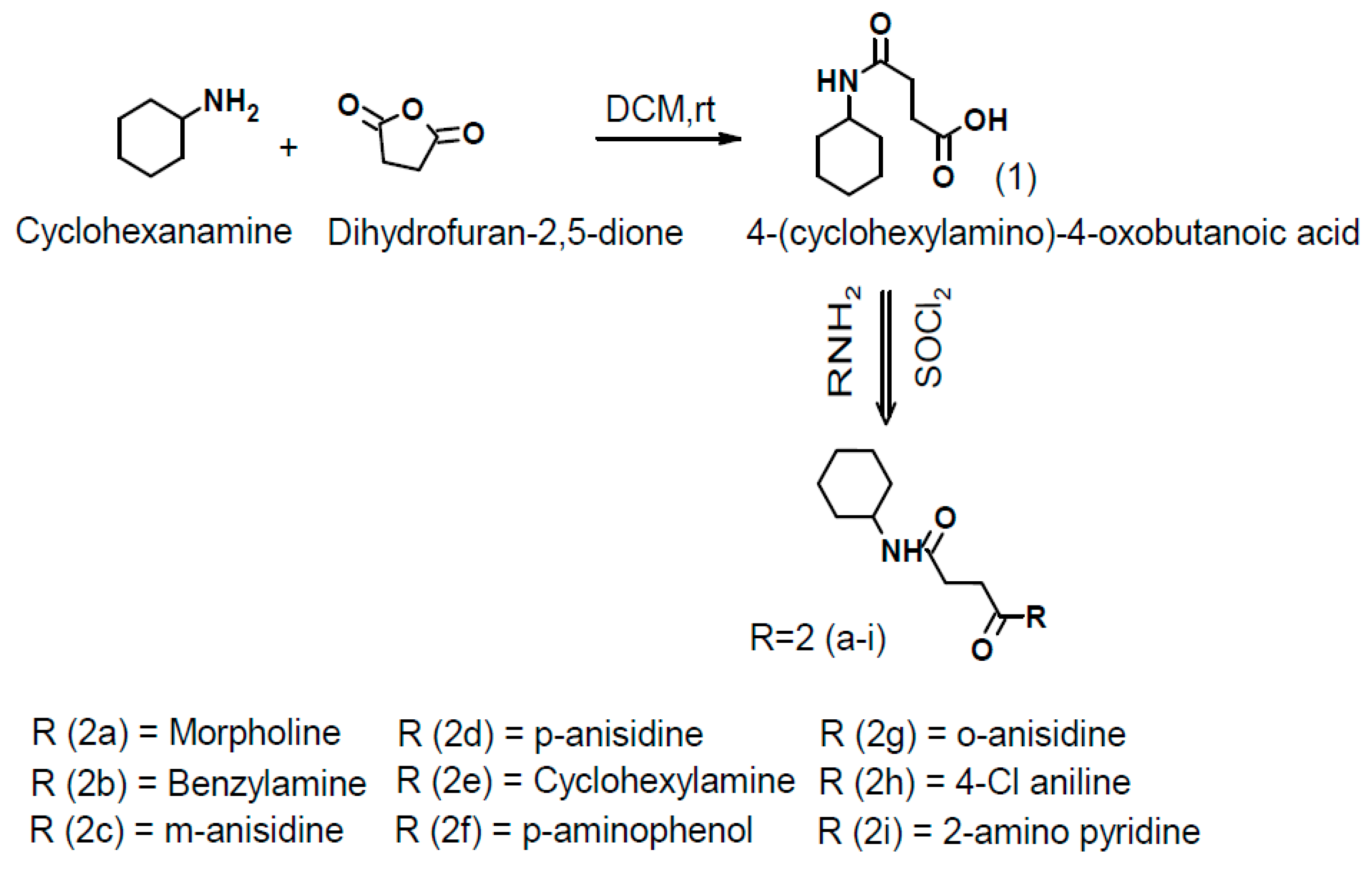
2.2. General Procedure for One-Pot Synthesis of Succinamide Derivatives(2a-2i)
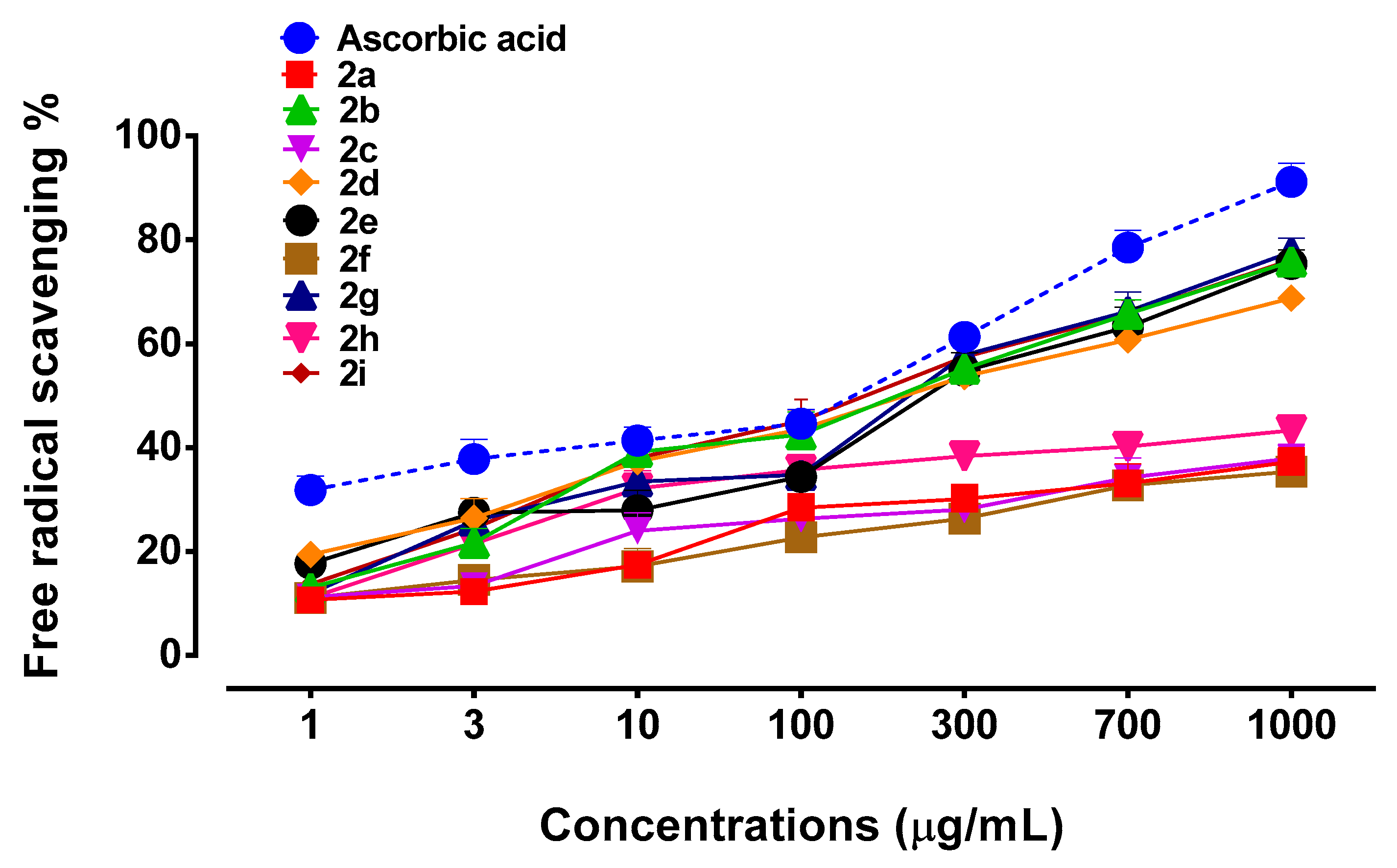
2.3. DPPH Free Radical Scavenging Assay
2.4. In-Silico Studies
2.5. Animals and Experimental Groups
2.6. Behavioural Studies
2.6.1. Y-Maze Test
2.6.2. Morris Water Maze Test (MWM)
2.7. Hematoxylin Eosin (H&E) Staining
2.8. Immunohistochemical Analysis
2.9. Assessment of Antioxidant Enzymes
2.9.1. GSH and GST Analysis
2.9.2. LPO Assay
2.9.3. Catalase Assay
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Statistical Analysis
3. Results
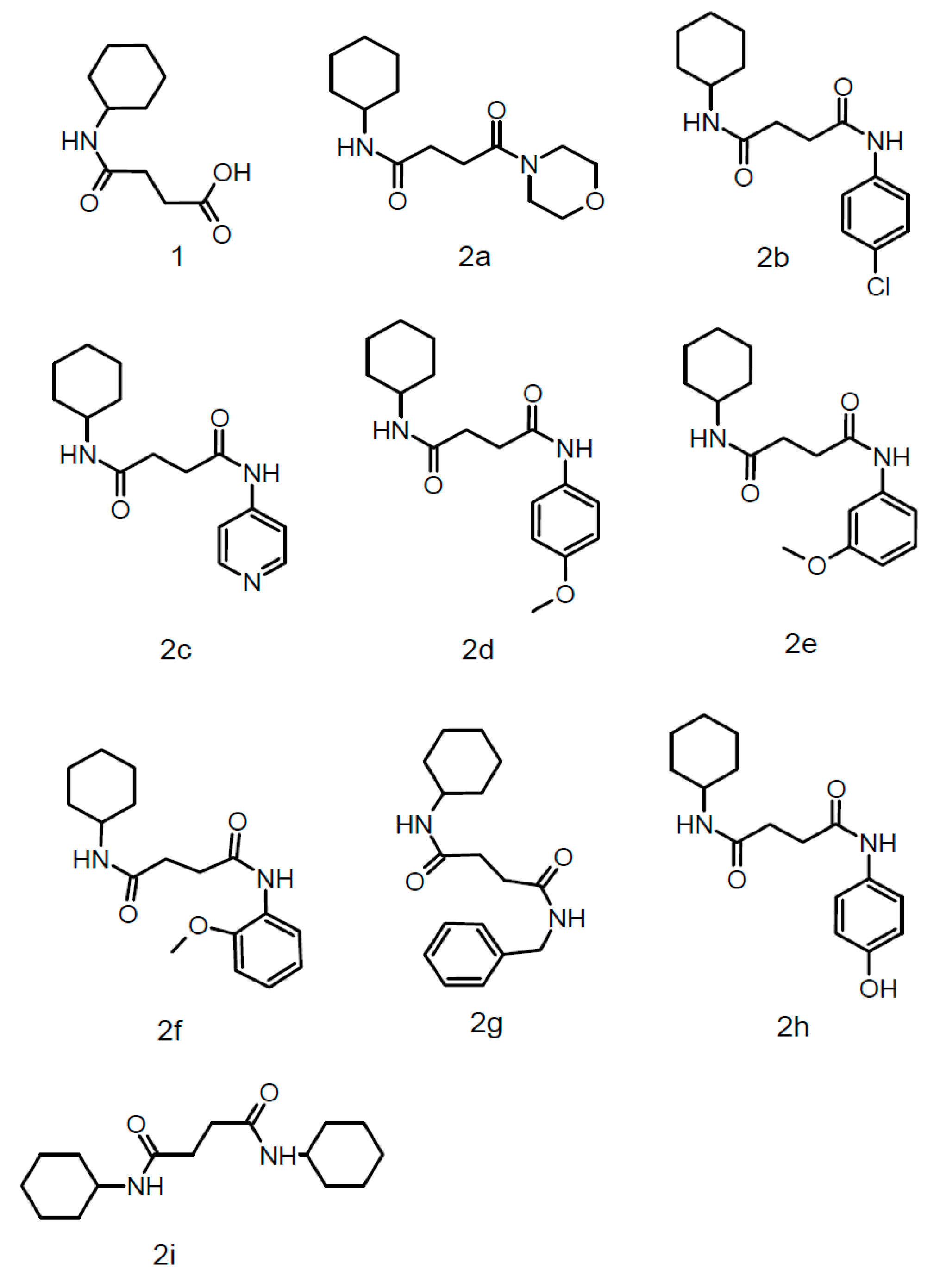
3.1. Spectral Analysis
3.1.1. 4-(Cyclohexylamine)-4-oxobutanoic acid (1)
3.1.2. N-cyclohexyl-4-(morpholin-4-yl)-4-oxobutanamide (2a)
3.1.3. N-(4-chlorophenyl)-N′-cyclohexylbutanediamide (2b)
3.1.4. N-cyclohexyl-N′-(pyridin-4-yl)butanediamide) (2c)
3.1.5. N-cyclohexyl-N′-(4-methoxyphenyl)butanediamide) (2d)
3.1.6. N-cyclohexyl-N′-(3-methoxyphenyl butanediamide) (2e)
3.1.7. N-cyclohexyl-N′-(2-methoxyphenyl)butanediamide) (2f)
3.1.8. N-benzyl-N′-cyclohexylbutanediamide (2g)
3.1.9. N-cyclohexyl-N′-(4-hydroxyphenyl)butanediamide (2h)
3.1.10. N,N′-di cyclohexyl butanediamide (2i)
3.2. Effect of Succinamide Derivatives on DPPH Free Radical Scavenging Assay
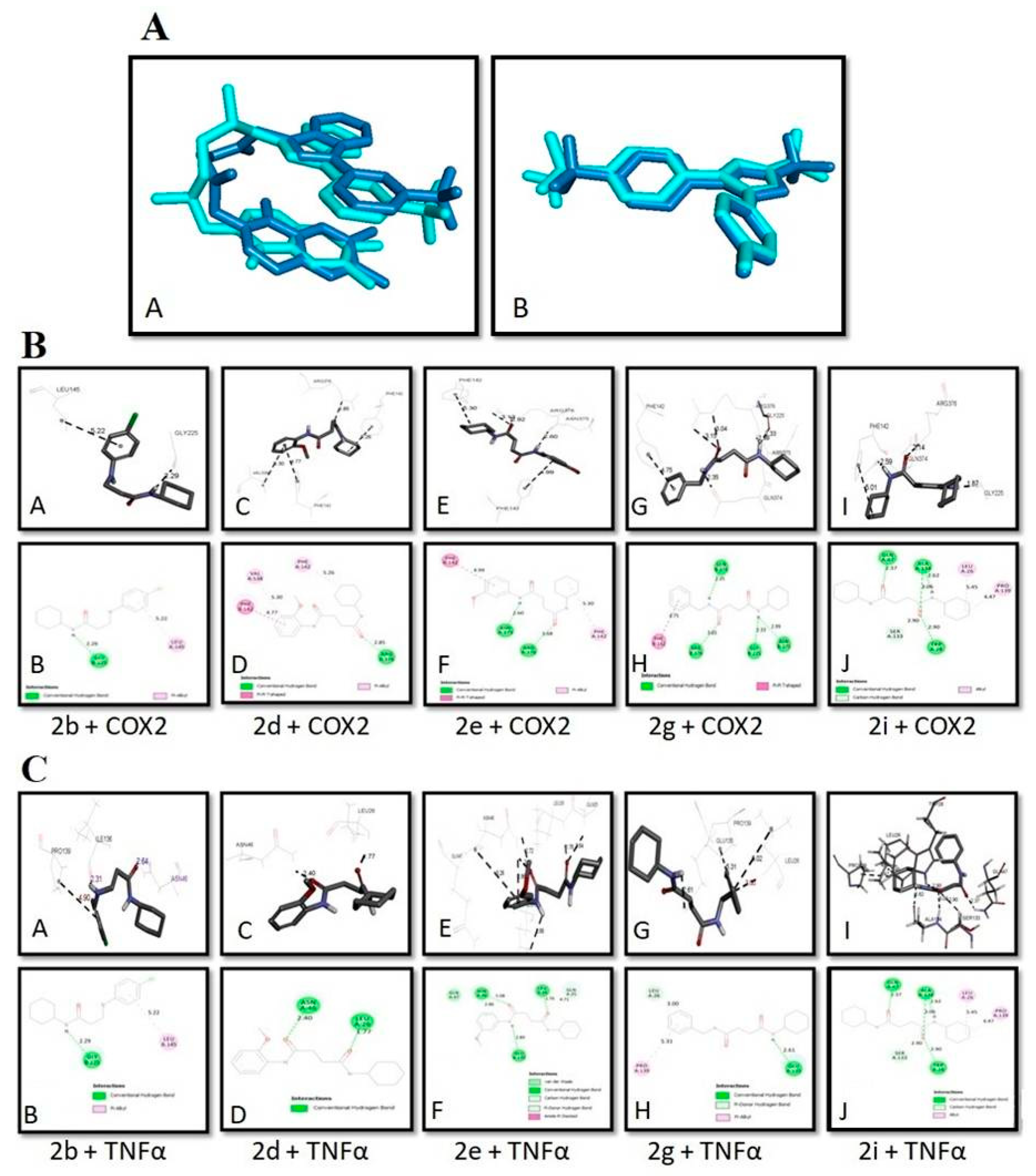
3.3. Evaluation of In-silico studies
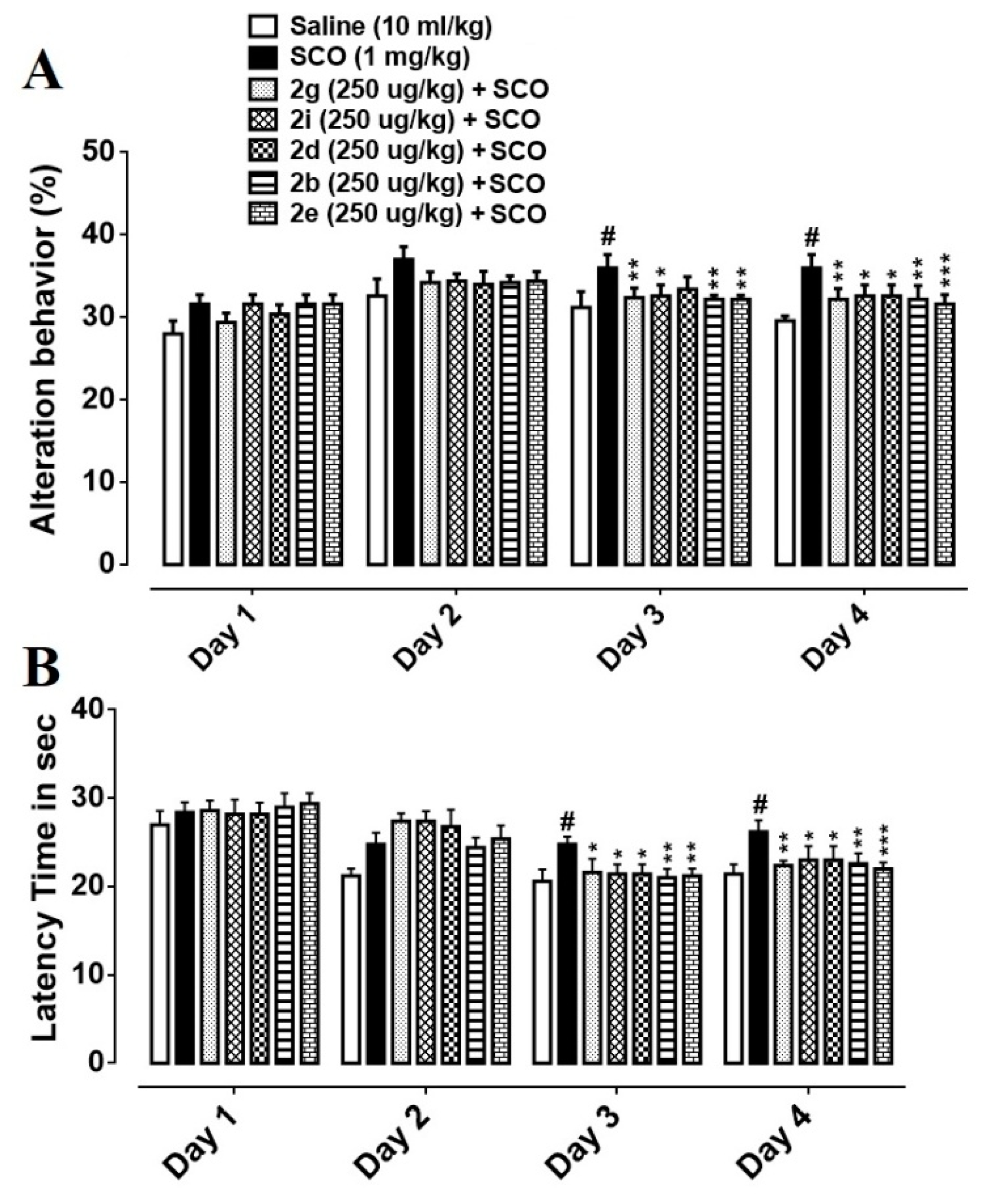
3.4. Effect of Succinamide Derivatives on Alteration Behaviour
3.5. Effect of Succinamide Derivatives on Escape Latency Time
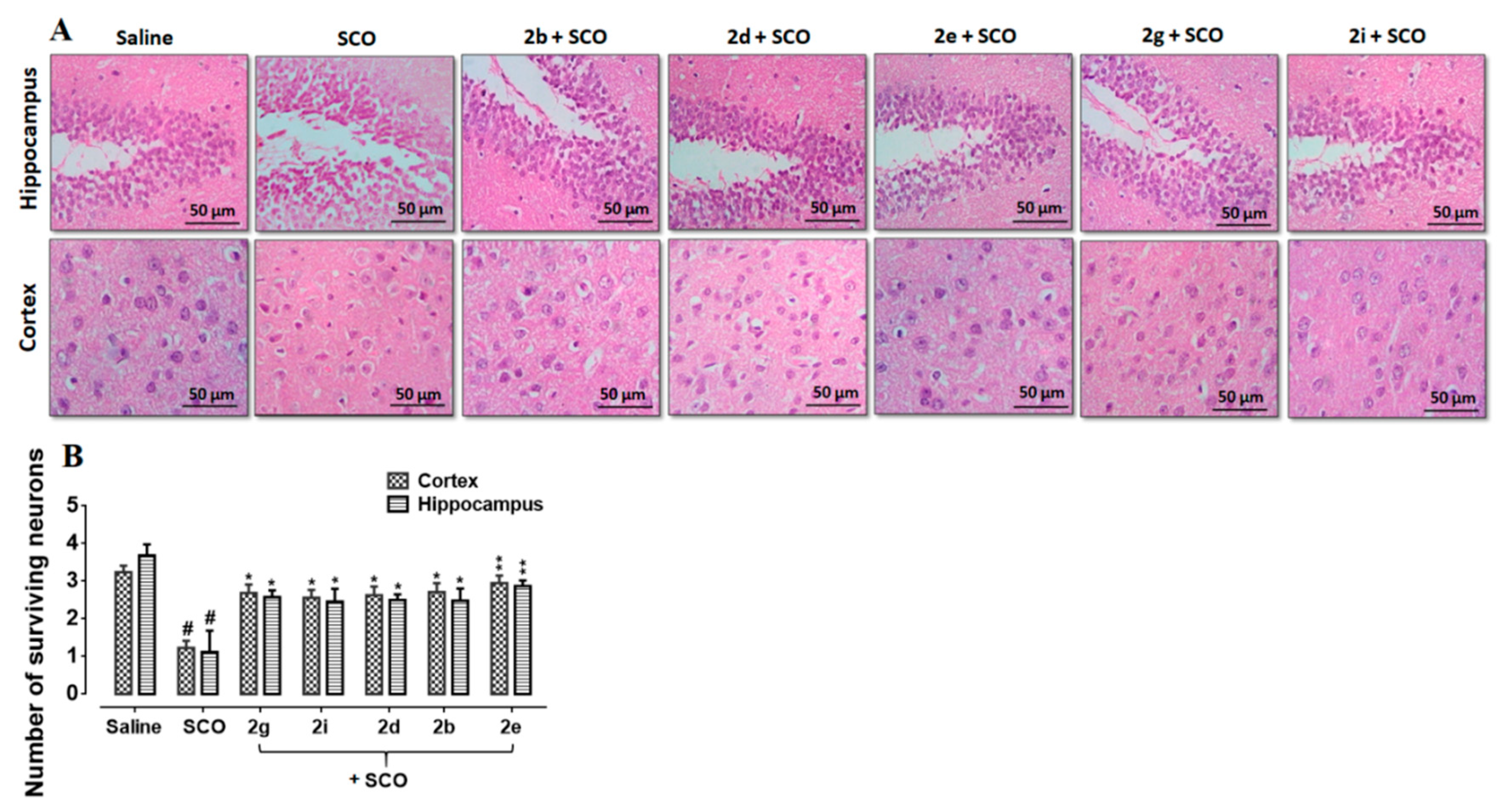
3.6. Effect of Succinamide Derivatives on Scopolamine-Induced Neurodegeneration
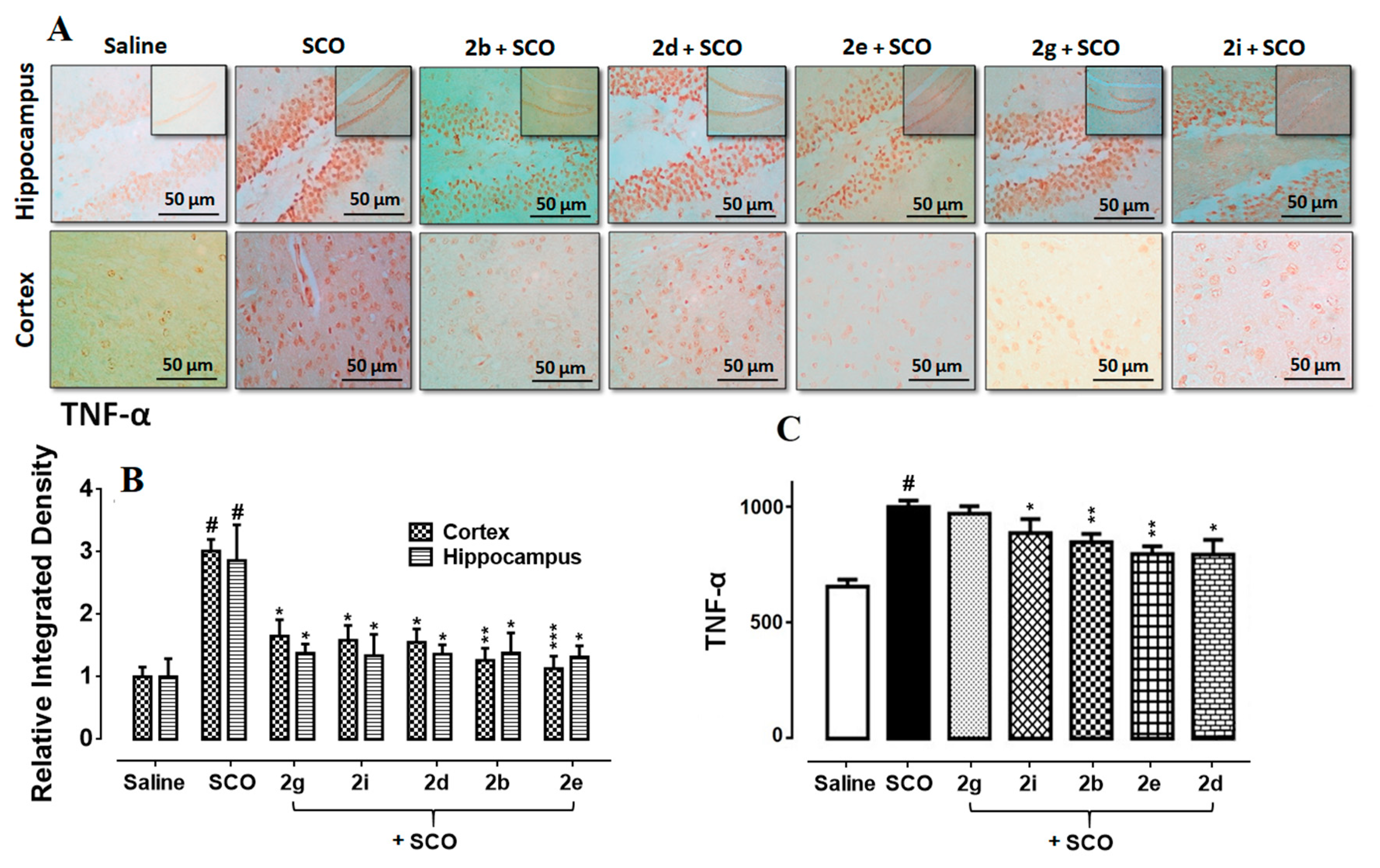
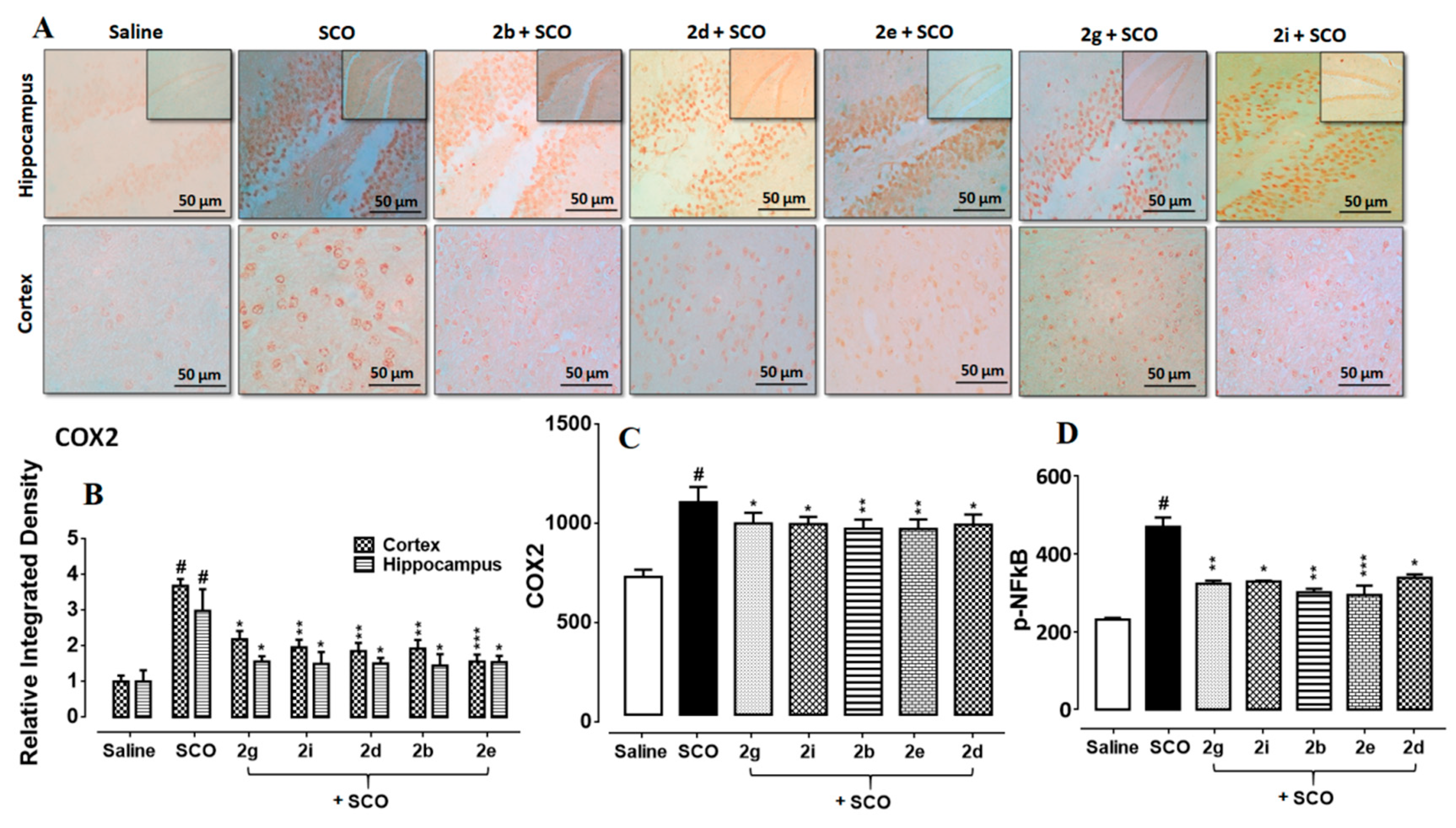
3.7. Succinamide Derivatives Mediated Downregulation of Neuroinflammation
3.8. Succinamide Derivatives Attenuated the Scopolamine-Induced Inflammatory Mediators
3.9. Effect of Succinamide Derivatives on Antioxidant Enzymes
3.10. Effect of Succinamide Derivatives on Scopolamine-Induced Lipid Peroxidation (LPO)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Imran, M.; AlKury, L.T.; Nadeem, H.; Shah, F.A.; Abbas, M.; Naz, S.; Khan, A.-U.; Li, S. Benzimidazole Containing Acetamide Derivatives Attenuate Neuroinflammation and Oxidative Stress in Ethanol-Induced Neurodegeneration. Biomolecules 2020, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Nuri, M.; Huda, T.; Hong, Y.H.; Ming, L.C.; MohdJoffry, S.; Othman, M.F.; Neoh, C.F. Knowledge on Alzheimer’s disease among public hospitals and health clinics pharmacists in the State of Selangor, Malaysia. Front. Pharmacol. 2017, 8, 739. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Liu, G.-P.; AlKury, L.T.; Zeb, A.; Abbas, M.; Li, T.; Yang, X.; Liu, F.; Jiang, Y.; Li, S. Melatonin protects MCAO-induced neuronal loss via NR2A mediated prosurvival pathways. Front. Pharmacol. 2019, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tocchetti, C.G.; Krieg, T.; Moens, A.L. Oxidative and nitrosative stress in the maintenance of myocardial function. Free Radic. Biol. Med. 2012, 53, 1531–1540. [Google Scholar] [CrossRef]
- Al Kury, L.T.; Zeb, A.; Abidin, Z.U.; Irshad, N.; Malik, I.; Alvi, A.M.; Khalil, A.A.K.; Ahmad, S.; Faheem, M.; Khan, A.-U. Neuroprotective effects of melatonin and celecoxib against ethanol-induced neurodegeneration: A computational and pharmacological approach. Drug Des. Devel. Ther. 2019, 13, 2715. [Google Scholar] [CrossRef]
- Shah, F.-A.; Park, D.-J.; Koh, P.-O. Identification of proteins differentially expressed by quercetin treatment in a middle cerebral artery occlusion model: A proteomics approach. Neurochem. Res. 2018, 43, 1608–1623. [Google Scholar] [CrossRef]
- Shah, F.-A.; Kury, L.-A.; Li, T.; Zeb, A.; Koh, P.-O.; Liu, F.; Zhou, Q.; Hussain, I.; Khan, A.-U.; Jiang, Y.; et al. Polydatin Attenuates Neuronal Loss via Reducing Neuroinflammation and Oxidative Stress in Rat MCAO Models. Front. Pharmacol. 2019, 26, 663. [Google Scholar] [CrossRef]
- Lenzlinger, P.M.; Hans, V.H.; Joller-Jemelka, H.I.; Trentz, O.; Morganti-Kossmann, M.C.; Kossmann, T. Markers for cell-mediated immune response are elevated in cerebrospinal fluid and serum after severe traumatic brain injury in humans. J. Neurotrauma 2001, 18, 479–489. [Google Scholar] [CrossRef]
- Ali, T.; Rehman, S.U.; Shah, F.A.; Kim, M.O. Acute dose of melatonin via Nrf2 dependently prevents acute ethanol-induced neurotoxicity in the developing rodent brain. J. Neuroinflamm. 2018, 15, 119. [Google Scholar] [CrossRef]
- Aloisi, F. Immune function of microglia. Glia 2001, 36, 165–179. [Google Scholar] [CrossRef]
- Ishola, I.O.; Adeyemi, O.O.; Agbaje, E.O.; Tota, S.; Shukla, R. Combretum mucronatum and Capparis thonningii prevent scopolamine-induced memory deficit in mice. Pharm. Biol. 2013, 51, 49–57. [Google Scholar] [CrossRef]
- Fan, Y.; Hu, J.; Li, J.; Yang, Z.; Xin, X.; Wang, J.; Ding, J.; Geng, M. Effect of acidic oligosaccharide sugar chain on scopolamine-induced memory impairment in rats and its related mechanisms. Neurosci. Lett. 2005, 374, 222–226. [Google Scholar] [CrossRef]
- Jawaid, T.; Shakya, A.K.; Siddiqui, H.H.; Kamal, M. Evaluation of Cucurbita maxima extract against scopolamine-induced amnesia in rats: Implication of tumour necrosis factor alpha. Z. Naturforsch. C J. Biosci. 2014, 69, 407–417. [Google Scholar] [CrossRef]
- Xian, Y.-F.; Ip, S.-P.; Mao, Q.-Q.; Su, Z.-R.; Chen, J.-N.; Lai, X.-P.; Lin, Z.-X. Honokiol improves learning and memory impairments induced by scopolamine in mice. Eur. J. Pharmacol. 2015, 760, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, S.D.; Tota, S.; Khandelwal, K.; Verma, P.; Nath, C.; Hanif, K.; Shukla, R.; Saxena, J.; Dwivedi, A.K. Protective effect of fruits of Morindacitrifolia L. on scopolamine induced memory impairment in mice: A behavioral, biochemical and cerebral blood flow study. J. Ethnopharmacol. 2012, 139, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Ali, T.; Ikram, M.; Khan, A.; Alam, S.I.; Kim, M.O. Melatonin rescue oxidative stress-mediated neuroinflammation/neurodegeneration and memory impairment in scopolamine-induced amnesia mice model. J. Neuroimmune Pharm. 2019, 14, 278–294. [Google Scholar] [CrossRef]
- Vesna, D.V.; Željko, J.V.; Nebojša, R.B.; Nataša, V.V.; Gordana, S.U.; Ivan, O.J. Quantum mechanical and spectroscopic (FT-IR, 13C, 1H NMR and UV) investigations of potent antiepileptic drug 1-(4-chlorophenyl)-3-phenyl-succinimide. Spectrochim. Acta A Mol. Biomol. Spectrosco. 2013, 117, 42–53. [Google Scholar]
- Patil, M.; Rajput, S. Succinimides: Synthesis, reaction, and biological activity. Int. J. Pharm. Pharm. Sci. 2014, 6, 8–14. [Google Scholar]
- Sadiq, A.; Mahmood, F.; Ullah, F.; Ayaz, M.; Ahmad, S.; Haq, F.U.; Khan, G.; Jan, M.S. Synthesis, anticholinesterase and antioxidant potentials of ketoesters derivatives of succinimides: A possible role in the management of Alzheimer’s. Chem. Cent. J. 2015, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Michaelidou, A.; Hadjipavlou-Litina, D. Anti-inflammatory, antioxidant and analgesic amides. J. Enzym. Inhib. Med. Chem. 2003, 18, 537–544. [Google Scholar] [CrossRef]
- Leggio, A.; Belsito, E.; De Luca, G.; DiGioia, M.; Leotta, V.; Romio, E.; Siciliano, C.; Liguori, A. One-pot synthesis of amides from carboxylic acids activated using thionyl chloride. RSC Adv. 2016, 6, 34468–34475. [Google Scholar] [CrossRef]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Volkamer, A.; Kuhn, D.; Grombacher, T.; Rippmann, F.; Rarey, M. Combining global and local measures for structure-based druggability predictions. J. Chem. Inf. Model. 2012, 52, 360–372. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Dowling, C.; Allen, N.J. Mice Lacking Glypican 4 Display Juvenile Hyperactivity and Adult Social Interaction Deficits. Brain Plast. 2018, 4, 197–209. [Google Scholar] [CrossRef]
- Kim, D.H.; Hung, T.M.; Bae, K.H.; Jung, J.W.; Lee, S.; Yoon, B.H.; Cheong, J.H.; Ko, K.H.; Ryu, J.H. Gomisin A improves scopolamine-induced memory impairment in mice. Eur. J. Pharmacol. 2006, 542, 129–135. [Google Scholar] [CrossRef]
- Kang, J.B.; Park, D.J.; Shah, M.A.; Kim, M.O.; Koh, P.O. Lipopolysaccharide induces neuroglia activation and NF-κB activation in cerebral cortex of adult mice. Lab. Anim. Res. 2019, 35, 19. [Google Scholar]
- Shah, F.-A.; Zeb, A.; Ali, T.; Muhammad, T.; Faheem, M.; Alam, S.-I.; Saeed, K.; Koh, P.-O.; Lee, K.-W.; Kim, M.-O. Identification of Proteins Differentially Expressed in the Striatum by Melatonin in a Middle Cerebral Artery Occlusion Rat Model-a Proteomic and in silico Approach. Front. Neurosci. 2018, 12, 888. [Google Scholar] [CrossRef]
- Khan, A.; Shah, B.; Naveed, M.; Shah, F.A.; Atiq, A.; Khan, N.U.; Kim, Y.S.; Khan, S. Matrine ameliorates anxiety and depression-like behaviour by targeting hyperammonemia-induced neuroinflammation and oxidative stress in CCl4 model of liver injury. Neurotoxicology 2019, 72, 38–50. [Google Scholar] [CrossRef]
- Hadwan, M.H. Simple spectrophotometric assay for measuring catalase activity in biological tissues. BMC Biochem. 2018, 19, 7. [Google Scholar] [CrossRef]
- Nakajima, K.; Tohyama, Y.; Kohsaka, S.; Kurihara, T. Protein kinase Cα requirement in the activation of p38 mitogen-activated protein kinase, which is linked to the induction of tumor necrosis factor α in lipopolysaccharide-stimulated microglia. Neurochem. Int. 2004, 44, 205–214. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Eikelenboom, P.; Bate, C.; VanGool, W.; Hoozemans, J.; Rozemuller, J.; Veerhuis, R.; Williams, A. Neuroinflammation in Alzheimer’s disease and prion disease. Glia 2002, 40, 232–239. [Google Scholar] [CrossRef]
- Shah, F.-A.; Li, T.; Kury, L.-T.-A.; Zeb, A.; Khatoon, S.; Liu, G.; Yang, X.; Liu, F.; Yao, H.; Khan, A.-U.; et al. Pathological Comparisons of the Hippocampal Changes in the Transient and Permanent Middle Cerebral Artery Occlusion Rat Models. Front. Neurol. 2019, 10, 1178. [Google Scholar] [CrossRef] [PubMed]
- Goverdhan, P.; Sravanthi, A.; Mamatha, T. Neuroprotective effects of meloxicam and selegiline in scopolamine-induced cognitive impairment and oxidative stress. Int. J. Alzheimers Dis. 2012, 2012, 974013. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.K.; Pritchard, J.; Dave, H. Cyclic carboxylic monoimides. Chem. Rev. 1970, 70, 439–469. [Google Scholar] [CrossRef]
- Husain, A.; Ahuja, P.; Ahmad, A.; Khan, S.A. Synthesis, Biological Evaluation and Pharmacokinetic Studies of Mefenamic Acid-N-Hydroxymethylsuccinimide Ester Prodrug as Safer NSAID. Med. Chem. 2016, 12, 585–591. [Google Scholar] [CrossRef]
- Arya, S.; Kumar, S.; Rani, R.; Kumar, N.; Roy, P.; Sondhi, S.M. Synthesis, anti-inflammatory, and cytotoxicity evaluation of 9, 10-dihydroanthracene-9, 10-α, β-succinimide and bis-succinimide derivatives. Med. Chem. Res. 2013, 22, 4278–4285. [Google Scholar] [CrossRef]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine induced impairment of spontaneous alternation behavior by antagonist but not inverse agonist and beta-carboline. Psychopharmacology 1988, 94, 491–495. [Google Scholar] [CrossRef]
- Morris, R.G. Development of a water maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Barnes, C.A.; Danysz, W.; Parsons, C.G. Effects of the uncompetitive NMDA receptor antagonist memantine on hippocampal long-term potentiation, short-term exploratory modulation and spatial memory in awake, freely moving rats. Eur. J. Neurosci. 1996, 8, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Blokland, A.; Geraerts, E.; Been, M. A detailed analysis of rats’ spatial memory in a probe trial of a Morris task. Behav. Brain Res. 2004, 154, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Vallés, S.L.; Blanco, A.M.; Pascual, M.; Guerri, C. Chronic ethanol treatment enhances inflammatory mediators and cell death in the brain and in astrocytes. Brain Pathol. 2004, 14, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Loeches, S.; Pascual-Lucas, M.; Blanco, A.M.; Sanchez-Vera, I.; Guerri, C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J. Neurosci. Res. 2010, 30, 8285–8295. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Energies (kcal/mol) | ||
|---|---|---|
| Compounds | COX2 | TNFα |
| 2b | −6.8 | −5.5 |
| 2d | −7.6 | −6.1 |
| 2e | −7.8 | −6.0 |
| 2g | −7.9 | −5.7 |
| 2i | −6.4 | −5.9 |
| Groups | GST (µmol/mg of Protein) | GSH (µmol CDNB Conjugate/min/mg of Protein) | Catalase (µmol H2O2/min/mg of Protein) | LPO (nmol/TBARS/mg of Protein) |
|---|---|---|---|---|
| Saline | 88.25 ± 0.15 | 67.90 ± 0.56 | 165.6 ± 1.650 | 85.15 ± 0.91 |
| Scopolamine | 27.8 ± 2.89 ### | 26.78 ± 2.78 ### | 88.80 ± 3.890 ### | 123.70 ± 3.71### |
| 2g | 45.1 ± 3.8 * | 33.90 ± 1.23 * | 125.1±3.230 ** | 87.45±8.90 ** |
| 2i | 43.78 ± 3.7 * | 32.78 ± 0.90 * | 120.5 ± 8.890 * | 96.89 ± 4.56 * |
| 2d | 44.10 ± 3.78 * | 33.33 ± 0.67 * | 127.4 ± 4.900 ** | 92.78 ± 2.78 * |
| 2b | 50.13 ± 3.89 ** | 35.33 ± 0.67 ** | 130.9 ± 6.890 ** | 85.12 ± 4.89 ** |
| 2e | 55.23 ± 2.89 *** | 37.23 ± 0.78 *** | 135.9 ± 8.890 *** | 78.89 ± 6.89 *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, S.; Shah, F.A.; Naeem, K.; Nadeem, H.; Sarwar, S.; Ashraf, Z.; Imran, M.; Khan, T.; Anwar, T.; Li, S. Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration. Biomolecules 2020, 10, 443. https://doi.org/10.3390/biom10030443
Iqbal S, Shah FA, Naeem K, Nadeem H, Sarwar S, Ashraf Z, Imran M, Khan T, Anwar T, Li S. Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration. Biomolecules. 2020; 10(3):443. https://doi.org/10.3390/biom10030443
Chicago/Turabian StyleIqbal, Sumbal, Fawad Ali Shah, Komal Naeem, Humaira Nadeem, Sadia Sarwar, Zaman Ashraf, Muhammad Imran, Tariq Khan, Tayyaba Anwar, and Shupeng Li. 2020. "Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration" Biomolecules 10, no. 3: 443. https://doi.org/10.3390/biom10030443
APA StyleIqbal, S., Shah, F. A., Naeem, K., Nadeem, H., Sarwar, S., Ashraf, Z., Imran, M., Khan, T., Anwar, T., & Li, S. (2020). Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration. Biomolecules, 10(3), 443. https://doi.org/10.3390/biom10030443