Interactions between Muscle and Bone—Where Physics Meets Biology

 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
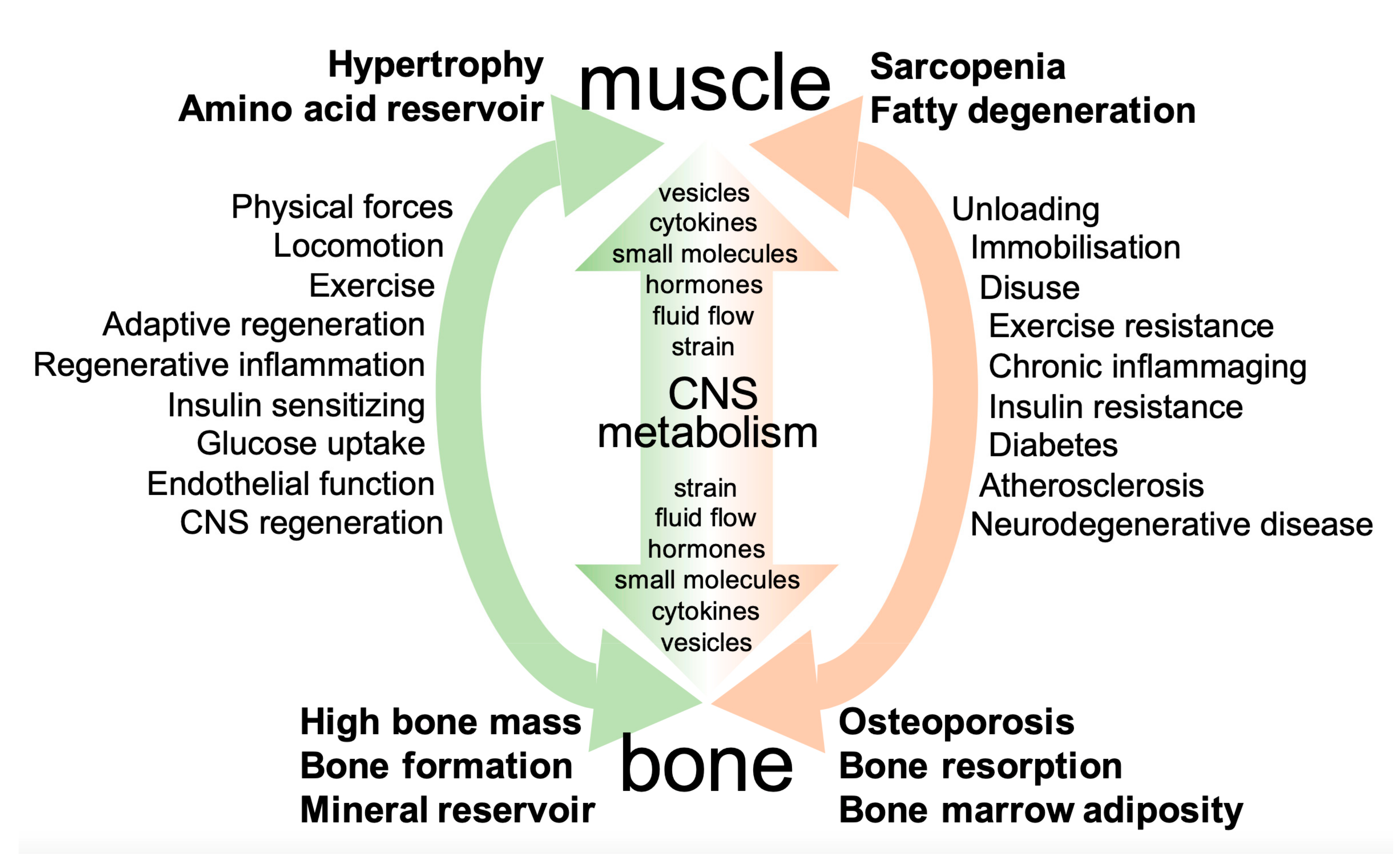
2. Principles of Interaction between Muscle and Bone
2.1. Physical Forces
2.1.1. Forces Generated by Exercise, Locomotion, and External Vibration
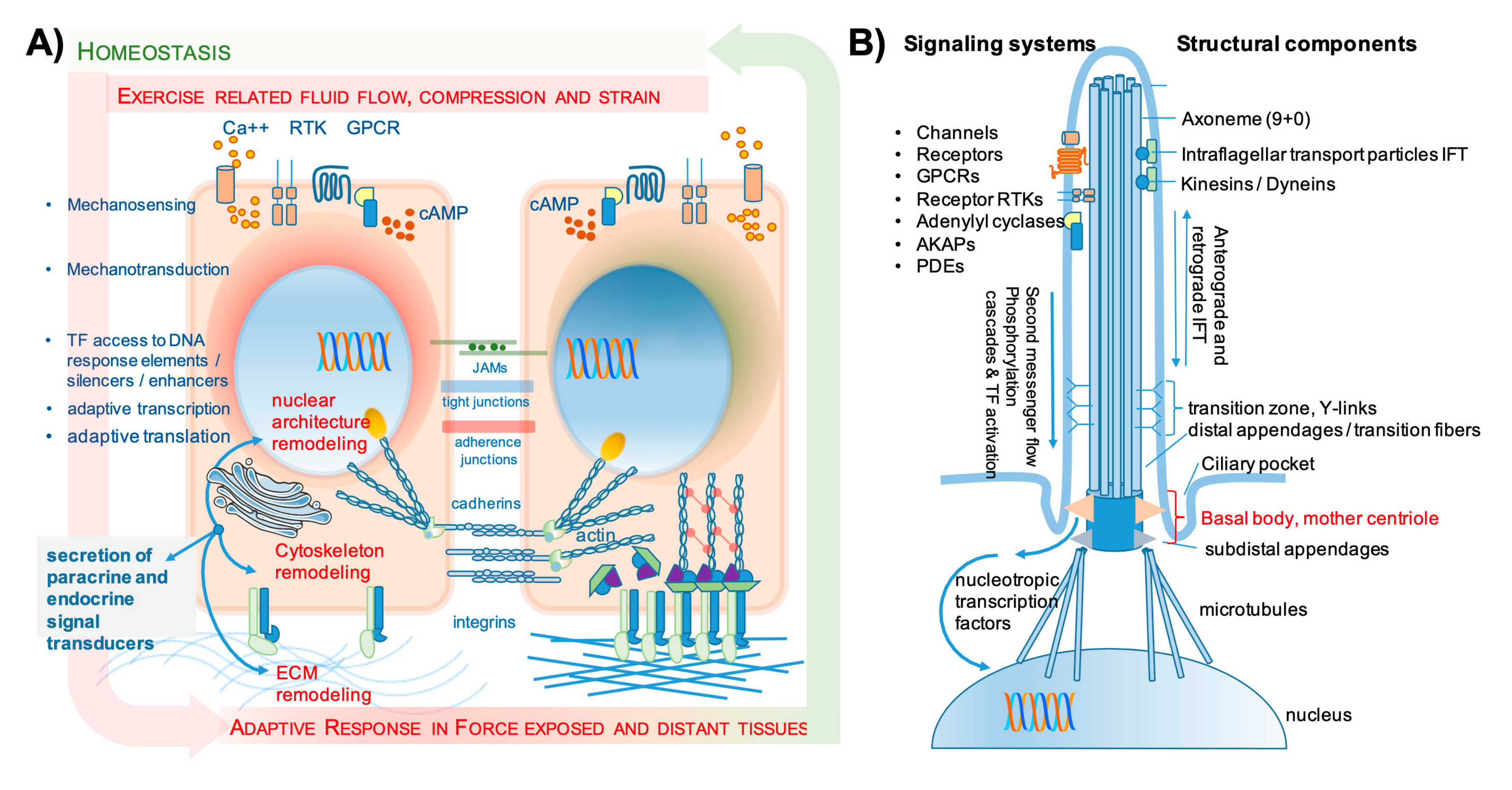
2.1.2. Mechanosensing and Mechanotransduction
2.2. Systemic Mutual Interactions of Skeletal Muscle and Bone
2.2.1. Secretion and Autocrine/Paracrine/Endocrine Communication
2.2.2. Bone Secretory Products with Endocrine Functions
2.2.3. Muscle Secretory Products with Endocrine Functions
3. Common Effector Mechanisms in Muscle and Bone from Development to Postnatal Regeneration
3.1. Role of Common Musculoskeletal Precursor’s Mechanotransduction during Development
3.2. Common Effectors of Muscle and Bone Interaction Modulate Tissue Formation and Remodeling
3.2.1. Principles of Bone Formation, Maintenance, and Regeneration
3.2.2. Principles of Muscle Formation, Regeneration, and Maintenance
3.3. Influence of Tissue Resident Cell Populations on Tissue Regeneration and Mechanoadaptation–A Focus on Immune Cell Interactions and Angiogenesis
3.4. Epigenetic Changes Shape Individual Responses According to Lifestyle, Environment, and Aging
3.5. Conclusions–Two Systems of Mutual Communication between Muscle and Bone
4. Muscle and Bone Interactions in Disease—Lessons and Backflashes from Pathology
4.1. Unloading–Lessons from Bed Rest Studies and Microgravity
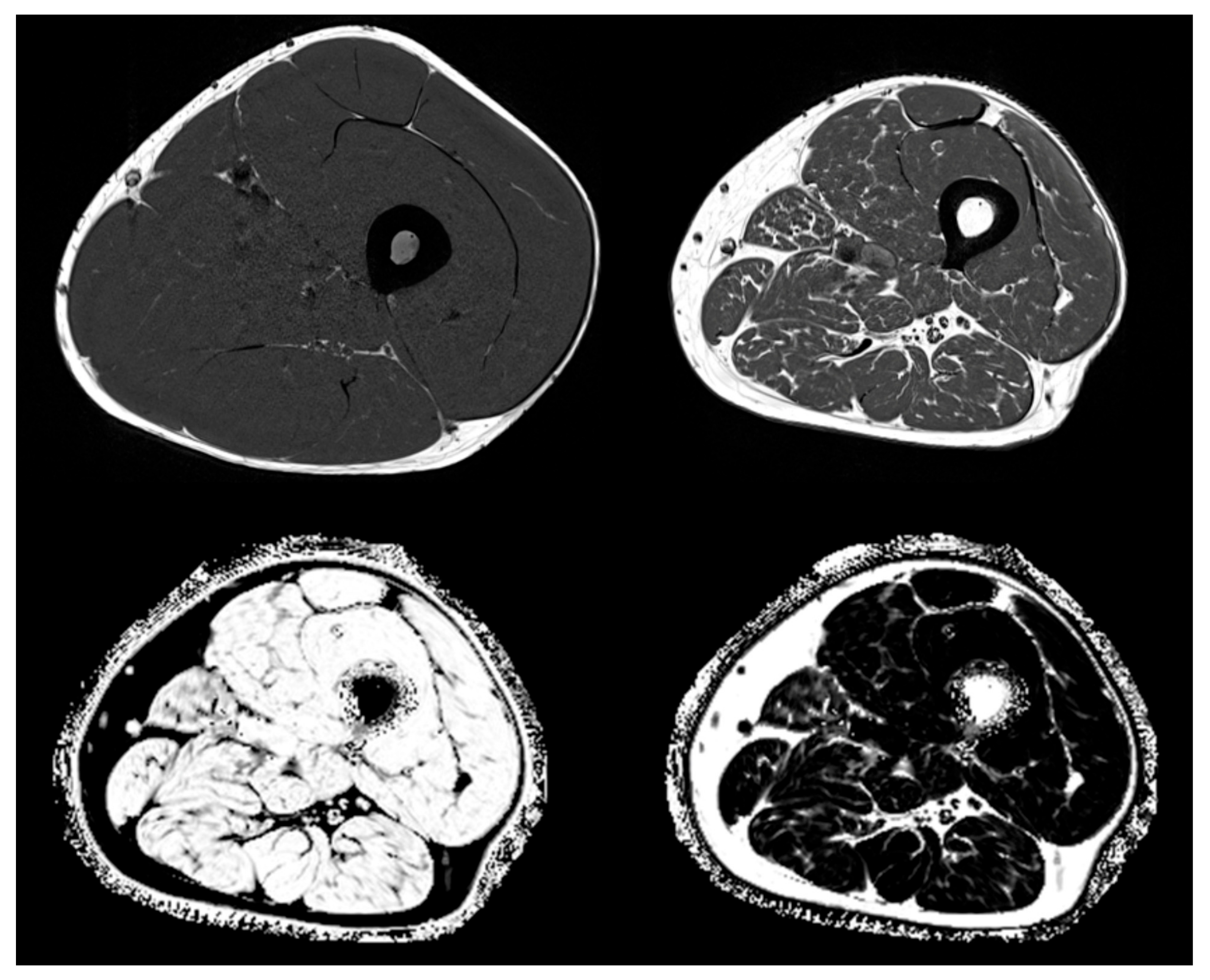
4.2. Inflammation and Fatty Degeneration—Lessons from Imaging and Structural Analyses
5. Conclusions and Perspectives
Funding
Conflicts of Interest
References
- Felsenthal, N.; Zelzer, E. Mechanical regulation of musculoskeletal system development. Dev. 2017, 144, 4271–4283. [Google Scholar] [CrossRef] [PubMed]
- Mikolajewicz, N.; Sehayek, S.; Wiseman, P.W.; Komarova, S.V. Transmission of Mechanical Information by Purinergic Signaling. Biophys. J. 2019, 116, 2009–2022. [Google Scholar] [CrossRef] [PubMed]
- Vogel, V. Unraveling the Mechanobiology of Extracellular Matrix. Annu. Rev. Physiol. 2018, 80, 353–387. [Google Scholar] [CrossRef] [PubMed]
- Ayad, N.M.; Kaushik, S.; Weaver, V.M. Tissue mechanics, an important regulator of development and disease. Philos. Trans. R. Soc. B: Boil. Sci. 2019, 374, 20180215. [Google Scholar] [CrossRef] [PubMed]
- Nassari, S.; Duprez, D.; Fournier-Thibault, C. Non-myogenic Contribution to Muscle Development and Homeostasis: The Role of Connective Tissues. Front. Cell Dev. Boil. 2017, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, D.; Versaevel, M.; Bruyère, C.; Alaimo, L.; Luciano, M.; Vercruysse, E.; Procès, A.; Gabriele, S. Innovative Tools for Mechanobiology: Unraveling Outside-In and Inside-Out Mechanotransduction. Front. Bioeng. Biotechnol. 2019, 7, 162. [Google Scholar] [CrossRef]
- Moro, A.; Driscoll, T.P.; Boraas, L.C.; Armero, W.; Kasper, D.M.; Baeyens, N.; Jouy, C.; Mallikarjun, V.; Swift, J.; Ahn, S.J.; et al. MicroRNA-dependent regulation of biomechanical genes establishes tissue stiffness homeostasis. Nat. 2019, 21, 348–358. [Google Scholar] [CrossRef]
- Uroz, M.; Wistorf, S.; Serra-Picamal, X.; Conte, V.; Sales-Pardo, M.; Roca-Cusachs, P.; Guimera, R.; Trepat, X. Regulation of cell cycle progression by cell–cell and cell–matrix forces. Nat. 2018, 20, 646–654. [Google Scholar] [CrossRef]
- Nachury, M.V.; Mick, D. Establishing and regulating the composition of cilia for signal transduction. Nat. Rev. Mol. Cell Boil. 2019, 20, 389–405. [Google Scholar] [CrossRef]
- Duncan, R.L. Transduction of mechanical strain in bone. ASGSB Bull. Publ. Am. Soc. Gravit. Space Boil. 1995, 8, 49–62. [Google Scholar]
- Duncan, R.L.; Turner, C.H. Mechanotransduction and the functional response of bone to mechanical strain. Calcif. Tissue Int. 1995, 57, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Pagnotti, G.M.; Styner, M.; Uzer, G.; Patel, V.S.; Wright, L.E.; Ness, K.K.; Guise, T.A.; Rubin, J.; Rubin, C.T. Combating osteoporosis and obesity with exercise: leveraging cell mechanosensitivity. Nat. Rev. Endocrinol. 2019, 15, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Stolzenberg, N.; Belavy, D.L.; Rawer, R.; Felsenberg, D. Whole-body vibration versus proprioceptive training on postural control in post-menopausal osteopenic women. Gait Posture 2013, 38, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Blottner, D.; Salanova, M.; Püttmann, B.; Schiffl, G.; Felsenberg, D.; Buehring, B.; Rittweger, J.; Buehring, B.; Rittweger, J. Human skeletal muscle structure and function preserved by vibration muscle exercise following 55 days of bed rest. Graefe’s Arch. Clin. Exp. Ophthalmol. 2006, 97, 261–271. [Google Scholar] [CrossRef]
- Haffner-Luntzer, M.; Lackner, I.; Liedert, A.; Fischer, V.; Ignatius, A. Effects of low-magnitude high-frequency vibration on osteoblasts are dependent on estrogen receptor α signaling and cytoskeletal remodeling. Biochem. Biophys. Res. Commun. 2018, 503, 2678–2684. [Google Scholar] [CrossRef]
- Razi, H.; I Birkhold, A.; Weinkamer, R.; Duda, G.N.; Willie, B.M.; Checa, S. Aging Leads to a Dysregulation in Mechanically Driven Bone Formation and Resorption. J. Bone Miner. Res. 2015, 30, 1864–1873. [Google Scholar] [CrossRef]
- Wehrle, E.; Liedert, A.; Heilmann, A.; Wehner, T.; Bindl, R.; Fischer, L.; Haffner-Luntzer, M.; Jakob, F.; Schinke, T.; Amling, M.; et al. The impact of low-magnitude high-frequency vibration on fracture healing is profoundly influenced by the oestrogen status in mice. Dis. Model. Mech. 2014, 8, 93–104. [Google Scholar] [CrossRef]
- Van Helvert, S.; Storm, C.; Friedl, P. Mechanoreciprocity in cell migration. Nat. 2017, 20, 8–20. [Google Scholar] [CrossRef]
- Iskratsch, T.; Wolfenson, H.; Sheetz, M.P. Appreciating force and shape — the rise of mechanotransduction in cell biology. Nat. Rev. Mol. Cell Boil. 2014, 15, 825–833. [Google Scholar] [CrossRef]
- Basoli, F.; Giannitelli, S.M.; Gori, M.; Mozetic, P.; Bonfanti, A.; Trombetta, M.; Rainer, A. Biomechanical Characterization at the Cell Scale: Present and Prospects. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Chakraborty, S.; Banerjee, S.; Raina, M.; Haldar, S. Force-Directed “Mechanointeractome” of Talin–Integrin. Biochem. 2019, 58, 4677–4695. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, R.; Vassalli, M.; Viti, F. Flow-induced mechanotransduction in skeletal cells. Biophys. Rev. 2019, 11, 729–743. [Google Scholar] [CrossRef]
- Le Roux, A.-L.; Quiroga, X.; Walani, N.; Arroyo, M.; Roca-Cusachs, P. The plasma membrane as a mechanochemical transducer. Philos. Trans. R. Soc. B: Boil. Sci. 2019, 374, 20180221. [Google Scholar] [CrossRef] [PubMed]
- Lemke, S.; Schnorrer, F. Mechanical forces during muscle development. Mech. Dev. 2017, 144, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Costell, M.; Fässler, R. Integrin activation by talin, kindlin and mechanical forces. Nat. 2019, 21, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Boil. 2016, 215, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Rozo, M.; Li, L.; Fan, C.-M. Targeting β1-integrin signaling enhances regeneration in aged and dystrophic muscle in mice. Nat. Med. 2016, 22, 889–896. [Google Scholar] [CrossRef]
- Plotkin, L.; Davis, H.M.; Cisterna, B.A.; Sáez, J.C. Connexins and Pannexins in Bone and Skeletal Muscle. Curr. Osteoporos. Rep. 2017, 15, 326–334. [Google Scholar] [CrossRef]
- Shen, H.; Grimston, S.; Civitelli, R.; Thomopoulos, S. Deletion of connexin43 in osteoblasts/osteocytes leads to impaired muscle formation in mice. J. Bone Miner. Res. 2015, 30, 596–605. [Google Scholar] [CrossRef]
- Donnaloja, F.; Jacchetti, E.; Soncini, M.; Raimondi, M.T. Mechanosensing at the Nuclear Envelope by Nuclear Pore Complex Stretch Activation and Its Effect in Physiology and Pathology. Front. Physiol. 2019, 10, 896. [Google Scholar] [CrossRef]
- Maurer, M.; Lammerding, J. The Driving Force: Nuclear Mechanotransduction in Cellular Function, Fate, and Disease. Annu. Rev. Biomed. Eng. 2019, 21, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Bakker, A.D.; Wu, G.; Klein-Nulend, J.; Jaspers, R.T. Physicochemical Niche Conditions and Mechanosensing by Osteocytes and Myocytes. Curr. Osteoporos. Rep. 2019, 17, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Hinton, P.V.; Rackard, S.M.; Kennedy, O.D. In Vivo Osteocyte Mechanotransduction: Recent Developments and Future Directions. Curr. Osteoporos. Rep. 2018, 16, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Wackerhage, H.; Schoenfeld, B.J.; Hamilton, D.L.; Lehti, M.; Hulmi, J. Stimuli and sensors that initiate skeletal muscle hypertrophy following resistance exercise. J. Appl. Physiol. 2019, 126, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Dewulf, M.; Köster, D.V.; Sinha, B.; De Lesegno, C.V.; Chambon, V.; Bigot, A.; Bensalah, M.; Negroni, E.; Tardif, N.; Podkalicka, J.; et al. Dystrophy-associated caveolin-3 mutations reveal that caveolae couple IL6/STAT3 signaling with mechanosensing in human muscle cells. Nat. Commun. 2019, 10, 1974. [Google Scholar] [CrossRef]
- Ferreira, R.R.; Fukui, H.; Chow, R.; Vilfan, A.; Vermot, J. The cilium as a force sensor-myth versus reality. J. Cell Sci. 2019, 132, jcs213496. [Google Scholar] [CrossRef]
- Moore, E.R.; Jacobs, C.R. The primary cilium as a signaling nexus for growth plate function and subsequent skeletal development. J. Orthop. Res. 2017, 36, 533–545. [Google Scholar] [CrossRef]
- Katsianou, M.A.; Skondra, F.G.; Gargalionis, A.N.; Piperi, C.; Basdra, E.K. The role of transient receptor potential polycystin channels in bone diseases. Ann. Transl. Med. 2018, 6, 246. [Google Scholar] [CrossRef]
- Piperi, C.; Basdra, E.K. Polycystins and mechanotransduction: From physiology to disease. World J. Exp. Med. 2015, 5, 200–205. [Google Scholar] [CrossRef]
- Shi, W.; Ma, Z.; Zhang, G.; Wang, C.; Jiao, Z. Novel functions of the primary cilium in bone disease and cancer. Cytoskelet. 2019, 76, 233–242. [Google Scholar] [CrossRef]
- Kwon, R.Y.; Temiyasathit, S.; Tummala, P.; Quah, C.C.; Jacobs, C.R. Primary cilium-dependent mechanosensing is mediated by adenylyl cyclase 6 and cyclic AMP in bone cells. FASEB J. 2010, 24, 2859–2868. [Google Scholar] [CrossRef] [PubMed]
- Marican, N.H.J.; Cruz-Migoni, S.B.; Borycki, A.-G. Asymmetric Distribution of Primary Cilia Allocates Satellite Cells for Self-Renewal. Stem Cell Rep. 2016, 6, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dynlacht, B.D. The regulation of cilium assembly and disassembly in development and disease. Dev. 2018, 145, dev151407. [Google Scholar] [CrossRef] [PubMed]
- Mammoto, T.; Mammoto, A.; Ingber, D.E. Mechanobiology and Developmental Control. Annu. Rev. Cell Dev. Boil. 2013, 29, 27–61. [Google Scholar] [CrossRef]
- Arvind, V.; Huang, A.H. Mechanobiology of limb musculoskeletal development. Ann. New York Acad. Sci. 2017, 1409, 18–32. [Google Scholar] [CrossRef]
- Petridou, N.; Spiró, Z.; Heisenberg, C.-P. Multiscale force sensing in development. Nat. 2017, 19, 581–588. [Google Scholar] [CrossRef]
- Suckale, J.; Solimena, M. The insulin secretory granule as a signaling hub. Trends Endocrinol. Metab. 2010, 21, 599–609. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Boil. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Vakilian, M.; Tahamtani, Y.; Ghaedi, K. A review on insulin trafficking and exocytosis. Gene 2019, 706, 52–61. [Google Scholar] [CrossRef]
- Bonnemaison, M.L.; Eipper, B.A.; Mains, R.E. Role of Adaptor Proteins in Secretory Granule Biogenesis and Maturation. Front. Endocrinol. 2013, 4, 101. [Google Scholar] [CrossRef] [PubMed]
- Seefried, L.; Müller-Deubert, S.; Krug, M.; Youssef, A.; Schütze, N.; Ignatius, A.; Jakob, F.; Ebert, R.; Mueller-Deubert, S.; Schuetze, N. Dissection of mechanoresponse elements in promoter sites of the mechanoresponsive CYR61 gene. Exp. Cell Res. 2017, 354, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Amanatullah, D.F.; Lu, J.; Hecht, J.; Posey, K.; Yik, J.; Di Cesare, P.E.; Haudenschild, D. Identification of a 3Kbp Mechanoresponsive Promoter Region in the Human Cartilage Oligomeric Matrix Protein Gene. Tissue Eng. Part A 2012, 18, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Müller-Deubert, S.; Seefried, L.; Krug, M.; Jakob, F.; Ebert, R. Epidermal growth factor as a mechanosensitizer in human bone marrow stromal cells. Stem Cell Res. 2017, 24, 69–76. [Google Scholar] [CrossRef]
- Dasgupta, I.; Mccollum, D. Control of cellular responses to mechanical cues through YAP/TAZ regulation. J. Boil. Chem. 2019, 294, 17693–17706. [Google Scholar] [CrossRef]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell Boil. 2017, 18, 758–770. [Google Scholar] [CrossRef]
- Zheng, Y.; Pan, D. The Hippo Signaling Pathway in Development and Disease. Dev. Cell 2019, 50, 264–282. [Google Scholar] [CrossRef]
- Ziouti, F.; Ebert, R.; Rummler, M.; Krug, M.; Müller-Deubert, S.; Lüdemann, M.; Jakob, F.; Willie, B.M.; Jundt, F. NOTCH Signaling Is Activated through Mechanical Strain in Human Bone Marrow-Derived Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019. [Google Scholar] [CrossRef]
- Bonewald, L.F. Use it or lose it to age: A review of bone and muscle communication. Bone 2019, 120, 212–218. [Google Scholar] [CrossRef]
- Lee, J.H.; Jun, H.-S. Role of Myokines in Regulating Skeletal Muscle Mass and Function. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, S.J.; Han, S.A.; Lee, S.H.; Koh, J.-M.; Hamrick, M.W.; Kim, B.-J. The effects of myokines on osteoclasts and osteoblasts. Biochem. Biophys. Res. Commun. 2019, 517, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Pourteymour, S.; Eckardt, K.; Holen, T.; Langleite, T.; Lee, S.; Jensen, J.; Birkeland, K.; Drevon, C.; Hjorth, M. Global mRNA sequencing of human skeletal muscle: Search for novel exercise-regulated myokines. Mol. Metab. 2017, 6, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Formigli, L.; Sassoli, C.; Squecco, R.; Bini, F.; Martinesi, M.; Chellini, F.; Luciani, G.; Sbrana, F.; Zecchi-Orlandini, S.; Francini, F.; et al. Regulation of transient receptor potential canonical channel 1 (TRPC1) by sphingosine 1-phosphate in C2C12 myoblasts and its relevance for a role of mechanotransduction in skeletal muscle differentiation. J. Cell Sci. 2009, 122, 1322–1333. [Google Scholar] [CrossRef] [PubMed]
- Brotto, M.; Bonewald, L. Bone and muscle: Interactions beyond mechanical. Bone 2015, 80, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Pierucci, F.; Zecchi-Orlandini, S.; Meacci, E. Sphingosine 1-Phosphate (S1P)/ S1P Receptor Signaling and Mechanotransduction: Implications for Intrinsic Tissue Repair/Regeneration. Int. J. Mol. Sci. 2019, 20, 5545. [Google Scholar] [CrossRef]
- Furber, K.L.; Backlund, P.S.; Yergey, A.L.; Coorssen, J.R. Unbiased Thiol-Labeling and Top-Down Proteomic Analyses Implicate Multiple Proteins in the Late Steps of Regulated Secretion. Proteomes 2019, 7, 34. [Google Scholar] [CrossRef]
- Gerber, S.H.; Südhof, T.C. Molecular determinants of regulated exocytosis. Diabetes 2002, 51, S3–S11. [Google Scholar] [CrossRef]
- Park, J.J.; Loh, Y.P. How peptide hormone vesicles are transported to the secretion site for exocytosis. Mol. Endocrinol. 2008, 22, 2583–2595. [Google Scholar] [CrossRef]
- Takamori, S.; Holt, M.; Stenius, K.; Lemke, E.A.; Grønborg, M.; Riedel, D.; Urlaub, H.; Schenck, S.; Brügger, B.; Ringler, P.; et al. Molecular Anatomy of a Trafficking Organelle. Cell 2006, 127, 831–846. [Google Scholar] [CrossRef]
- Bittel, D.C.; Jaiswal, J.K. Contribution of Extracellular Vesicles in Rebuilding Injured Muscles. Front. Physiol. 2019, 10, 828. [Google Scholar] [CrossRef]
- Lauritzen, H.P.M.M.; Brandauer, J.; Schjerling, P.; Koh, H.-J.; Treebak, J.T.; Hirshman, M.F.; Galbo, H.; Goodyear, L.J. Contraction and AICAR Stimulate IL-6 Vesicle Depletion From Skeletal Muscle Fibers In Vivo. Diabetes 2013, 62, 3081–3092. [Google Scholar] [CrossRef]
- Morrell, A.E.; Brown, G.N.; Robinson, S.T.; Sattler, R.L.; Baik, A.D.; Zhen, G.; Cao, X.; Bonewald, L.F.; Jin, W.; Kam, L.C.; et al. Mechanically induced Ca2+ oscillations in osteocytes release extracellular vesicles and enhance bone formation. Bone Res. 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Murray, L.M.; Krasnodembskaya, A. Concise Review: Intercellular Communication Via Organelle Transfer in the Biology and Therapeutic Applications of Stem Cells. STEM CELLS 2018, 37, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Trovato, E.; Di Felice, V.; Barone, R. Extracellular Vesicles: Delivery Vehicles of Myokines. Front. Physiol. 2019, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Annibalini, G.; Contarelli, S.; Lucertini, F.; Guescini, M.; Maggio, S.; Ceccaroli, P.; Gervasi, M.; Marini, C.F.; Fardetti, F.; Grassi, E.; et al. Muscle and Systemic Molecular Responses to a Single Flywheel Based Iso-Inertial Training Session in Resistance-Trained Men. Front. Physiol. 2019, 10, 554. [Google Scholar] [CrossRef] [PubMed]
- Moser, S.C.; Van Der Eerden, B. Osteocalcin—A Versatile Bone-Derived Hormone. Front. Endocrinol. 2019, 9. [Google Scholar] [CrossRef]
- Kim, J.A.; Roh, E.; Hong, S.-H.; Lee, Y.-B.; Kim, N.H.; Yoo, H.J.; Seo, J.A.; Kim, N.H.; Kim, S.G.; Baik, S.H.; et al. Association of serum sclerostin levels with low skeletal muscle mass: The Korean Sarcopenic Obesity Study (KSOS). Bone 2019, 128, 115053. [Google Scholar] [CrossRef]
- Ghardashi-Afousi, A.; Davoodi, M.; Hesamabadi, B.K.-; Asvadi-Fard, M.; Bigi, M.A.B.; Izadi, M.R.; Gaeini, A.A. Improved carotid intima-media thickness-induced high-intensity interval training associated with decreased serum levels of Dkk-1 and sclerostin in type 2 diabetes. J. Diabetes its Complicat. 2019, 34, 107469. [Google Scholar] [CrossRef]
- Picke, A.-K.; Sylow, L.; Møller, L.L.V.; Kjøbsted, R.; Schmidt, F.N.; Steejn, M.W.; Salbach-Hirsch, J.; Hofbauer, C.; Blüher, M.; Saalbach, A.; et al. Differential effects of high-fat diet and exercise training on bone and energy metabolism. Bone 2018, 116, 120–134. [Google Scholar] [CrossRef]
- Adhikary, S.; Choudhary, D.; Tripathi, A.K.; Karvande, A.; Ahmad, N.; Kothari, P.; Trivedi, R.; Ahmad, A.K.N. FGF-2 targets sclerostin in bone and myostatin in skeletal muscle to mitigate the deleterious effects of glucocorticoid on musculoskeletal degradation. Life Sci. 2019, 229, 261–276. [Google Scholar] [CrossRef]
- Li, G.; Zhang, L.; Wang, D.; Aiqudsy, L.; Jiang, J.X.; Xu, H.; Shang, P. Muscle-bone crosstalk and potential therapies for sarco-osteoporosis. J. Cell. Biochem. 2019, 120, 14262–14273. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Li, J.; Yi, Z. The role of the fibroblast growth factor family in bone-related diseases. Chem. Boil. Drug Des. 2019, 94, 1740–1749. [Google Scholar] [CrossRef] [PubMed]
- Avin, K.G.; Vallejo, J.A.; Chen, N.X.; Wang, K.; Touchberry, C.; Brotto, M.; Dallas, S.L.; Moe, S.M.; Wacker, M.J. Fibroblast growth factor 23 does not directly influence skeletal muscle cell proliferation and differentiation or ex vivo muscle contractility. Am. J. Physiol. Metab. 2018, 315, E594–E604. [Google Scholar] [CrossRef] [PubMed]
- Edmonston, D.; Wolf, M. FGF23 at the crossroads of phosphate, iron economy and erythropoiesis. Nat. Rev. Nephrol. 2019, 16, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Ramos-Molina, B.; Lick, A.N.; Prideaux, M.; Albornoz, V.; Bonewald, L.; Lindberg, I. Posttranslational processing of FGF23 in osteocytes during the osteoblast to osteocyte transition. Bone 2015, 84, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Long, D.; Li, J.; Yu, R.; Song, Y.; Fang, J.; Yang, X.; Zhou, S.; Huang, S.; Zhao, Z. Extracellular vesicles in bone and tooth: A state-of-art paradigm in skeletal regeneration. J. Cell. Physiol. 2019, 234, 14838–14851. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an Endocrine Organ: Focus on Muscle-Derived Interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef]
- Romagnoli, C.; Pampaloni, B.; Brandi, M. Muscle endocrinology and its relation with nutrition. Aging Clin. Exp. Res. 2019, 31, 783–792. [Google Scholar] [CrossRef]
- Le Bihan, M.-C.; Bigot, A.; Jensen, S.; Dennis, J.L.; Rogowska-Wrzesinska, A.; Lainé, J.; Gache, V.; Furling, D.; Jensen, O.N.; Voit, T.; et al. In-depth analysis of the secretome identifies three major independent secretory pathways in differentiating human myoblasts. J. Proteom. 2012, 77, 344–356. [Google Scholar] [CrossRef]
- E Terry, E.; Zhang, X.; Hoffmann, C.; Hughes, L.D.; A Lewis, S.; Li, J.; Wallace, M.J.; A Riley, L.; Douglas, C.M.; A Gutierrez-Monreal, M.; et al. Transcriptional profiling reveals extraordinary diversity among skeletal muscle tissues. eLife 2018, 7, e34613. [Google Scholar] [CrossRef]
- Ebert, R.; Benisch, P.; Krug, M.; Zeck, S.; Meißner-Weigl, J.; Steinert, A.; Rauner, M.; Hofbauer, L.; Jakob, D.F.; Steinert, A. Acute phase serum amyloid A induces proinflammatory cytokines and mineralization via toll-like receptor 4 in mesenchymal stem cells. Stem Cell Res. 2015, 15, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.; Yao, Z.; Goodman, S. Inflammation, fracture and bone repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Sass, F.A.; Fuchs, M.; Pumberger, M.; Geissler, S.; Duda, G.N.; Perka, C.; Schmidt-Bleek, K. Immunology Guides Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2018, 19, 835. [Google Scholar] [CrossRef] [PubMed]
- Stenina-Adognravi, O.; Plow, E.F. Thrombospondin-4 in tissue remodeling. Matrix Boil. 2019, 300–313. [Google Scholar] [CrossRef]
- Lavin, K.M.; Perkins, R.K.; Jemiolo, B.; Raue, U.; Trappe, S.W.; Trappe, T.A. Effects of aging and lifelong aerobic exercise on basal and exercise-induced inflammation. J. Appl. Physiol. 2019, 128, 87–99. [Google Scholar] [CrossRef]
- Brylka, L.J.; Schinke, T. Chemokines in Physiological and Pathological Bone Remodeling. Front. Immunol. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Hafen, B.; Wiesner, S.; Schlegelmilch, K.; Keller, A.; Seefried, L.; Ebert, R.; Walles, T.; Jakob, F.; Schütze, N. Physical contact between mesenchymal stem cells and endothelial precursors induces distinct signatures with relevance to the very early phase of regeneration. J. Cell. Biochem. 2018, 119, 9122–9140. [Google Scholar] [CrossRef]
- Herrmann, M.; Jakob, F. Bone Marrow Niches for Skeletal Progenitor Cells and their Inhabitants in Health and Disease. Curr. Stem Cell Res. Ther. 2019, 14, 305–319. [Google Scholar] [CrossRef]
- Robey, P. “Mesenchymal stem cells”: Fact or fiction, and implications in their therapeutic use. F1000Research 2017, 6, 524. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Remoli, C.; Giannicola, G.; Kogler, G.; Liedtke, S.; Cossu, G.; Serafini, M.; Sampaolesi, M.; Tagliafico, E.; et al. No Identical “Mesenchymal Stem Cells” at Different Times and Sites: Human Committed Progenitors of Distinct Origin and Differentiation Potential Are Incorporated as Adventitial Cells in Microvessels. Stem Cell Rep. 2016, 6, 897–913. [Google Scholar] [CrossRef]
- Chijimatsu, R.; Saito, T. Mechanisms of synovial joint and articular cartilage development. Cell. Mol. Life Sci. 2019, 76, 3939–3952. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, R.; Bressan, M.; Herzlinger, D. Paraxial mesoderm contributes stromal cells to the developing kidney. Dev. Boil. 2009, 329, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.; Lewis, P.; Przepiorski, A.; Sander, V. Turning mesoderm into kidney. Semin. Cell Dev. Boil. 2019, 91, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Limbert, C.; Ebert, R.; Path, G.; Niu, X.; Bretzel, G.; Seufert, J.; Jakob, F. Comparison of stemness and endocrine differentiation potential of human pancreatic islet derived and human bone-marrow derived stromal cells. J. Stem Cells Regen. Med. 2007, 2. [Google Scholar]
- Rux, D.R.; Wellik, D.M. Hox genes in the adult skeleton: Novel functions beyond embryonic development. Dev. Dyn. 2017, 246, 310–317. [Google Scholar] [CrossRef]
- Friedl, P.; Mayor, R. Tuning Collective Cell Migration by Cell–Cell Junction Regulation. Cold Spring Harb. Perspect. Boil. 2017, 9, a029199. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Swift, J. The consequences of ageing, progeroid syndromes and cellular senescence on mechanotransduction and the nucleus. Exp. Cell Res. 2019, 378, 98–103. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Serrano, M. Cellular senescence: from physiology to pathology. Nat. Rev. Mol. Cell Boil. 2014, 15, 482–496. [Google Scholar] [CrossRef]
- Czarkwiani, A.; Yun, M. Out with the old, in with the new: senescence in development. Curr. Opin. Cell Boil. 2018, 55, 74–80. [Google Scholar] [CrossRef]
- Sharir, A.; Stern, T.; Rot, C.; Shahar, R.; Zelzer, E. Muscle force regulates bone shaping for optimal load-bearing capacity during embryogenesis. Dev. 2011, 138, 3247–3259. [Google Scholar] [CrossRef]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: from human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Main, R.P.; Shefelbine, S.J.; Meakin, L.; Silva, M.J.; Van Der Meulen, M.C.; Willie, B.M. Murine Axial Compression Tibial Loading Model to Study Bone Mechanobiology: Implementing the Model and Reporting Results. J. Orthop. Res. 2019, 38, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Calle, J.; Tu, X.; Pacheco-Costa, R.; McAndrews, K.; Edwards, R.; Pellegrini, G.G.; Kuhlenschmidt, K.; Olivos, N.; Robling, A.; Peacock, M.; et al. Control of Bone Anabolism in Response to Mechanical Loading and PTH by Distinct Mechanisms Downstream of the PTH Receptor. J. Bone Miner. Res. 2016, 32, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E. Notch in skeletal physiology and disease. Osteoporos. Int. 2018, 29, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Qiu, Y.; Lin, K.C.; Kumar, A.; Placone, J.K.; Fang, C.; Wang, K.-C.; Lu, S.; Pan, M.; Hong, A.W.; et al. RAP2 mediates mechanoresponses of the Hippo pathway. Nat. 2018, 560, 655–660. [Google Scholar] [CrossRef]
- Xiang, L.; Yu, H.; Zhang, X.; Wang, B.; Yuan, Y.; Zhang, Q.; Ye, R.; Gong, P.; Wu, Y. The versatile hippo pathway in oral-maxillofacial development and bone remodeling. Dev. Boil. 2018, 440, 53–63. [Google Scholar] [CrossRef]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of Osteoclast Differentiation by Cytokine Networks. Immune Netw. 2018, 18. [Google Scholar] [CrossRef]
- Ma, Q.; Ma, Z.; Liang, M.; Luo, F.; Xu, J.; Dou, C.; Dong, S. The role of physical forces in osteoclastogenesis. J. Cell. Physiol. 2019, 234, 12498–12507. [Google Scholar] [CrossRef]
- Kim, B.-J.; Koh, J.-M. Coupling factors involved in preserving bone balance. Cell. Mol. Life Sci. 2018, 76, 1243–1253. [Google Scholar] [CrossRef]
- Alto, L.T.; Terman, J.R. Semaphorins and their Signaling Mechanisms. Breast Cancer 2017, 1493, 1–25. [Google Scholar]
- Paul, G.R.; Malhotra, A.; Müller, R. Mechanical Stimuli in the Local In Vivo Environment in Bone: Computational Approaches Linking Organ-Scale Loads to Cellular Signals. Curr. Osteoporos. Rep. 2018, 16, 395–403. [Google Scholar] [CrossRef] [PubMed]
- E Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, T.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2014, 11, 45–54. [Google Scholar] [CrossRef]
- Rauner, M.; Stein, N.; Winzer, M.; Goettsch, C.; Zwerina, J.; Schett, G.; Distler, J.H.; Albers, J.; Schulze, J.; Schinke, T.; et al. WNT5A is induced by inflammatory mediators in bone marrow stromal cells and regulates cytokine and chemokine production. J. Bone Miner. Res. 2012, 27, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Farr, J.N.; Khosla, S. Cellular senescence in bone. Bone 2019, 121, 121–133. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Sharpless, N.E. Senescence in Health and Disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef]
- Rhinn, M.; Ritschka, B.; Keyes, W.M. Cellular senescence in development, regeneration and disease. Dev. 2019, 146, dev151837. [Google Scholar] [CrossRef]
- Buckingham, M. Gene regulatory networks and cell lineages that underlie the formation of skeletal muscle. Proc. Natl. Acad. Sci. 2017, 114, 5830–5837. [Google Scholar] [CrossRef]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesisin vivoandin vitro. Dev. 2017, 144, 2104–2122. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Li, M.; He, J.; Huang, Z. Regulation of skeletal myogenesis by microRNAs. J. Cell. Physiol. 2019, 235, 87–104. [Google Scholar] [CrossRef]
- Baghdadi, M.B.; Tajbakhsh, S. Regulation and phylogeny of skeletal muscle regeneration. Dev. Boil. 2018, 433, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Dort, J.; Fabre, P.; Molina, T.; Dumont, N.A. Macrophages Are Key Regulators of Stem Cells during Skeletal Muscle Regeneration and Diseases. Stem Cells Int. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip. Rev. Syst. Boil. Med. 2019, 12, e1462. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Lewis, B.; Mourikis, P.; Fre, S. Notch signalling: Sensor and instructor of the microenvironment to coordinate cell fate and organ morphogenesis. Curr. Opin. Cell Boil. 2019, 61, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Rindom, E.; Vissing, K. Mechanosensitive Molecular Networks Involved in Transducing Resistance Exercise-Signals into Muscle Protein Accretion. Front. Physiol. 2016, 7, 547. [Google Scholar] [CrossRef] [PubMed]
- Hogarth, M.W.; Defour, A.; Lazarski, C.; Gallardo, E.; Manera, J.D.; Partridge, T.A.; Nagaraju, K.; Jaiswal, J.K. Fibroadipogenic progenitors are responsible for muscle loss in limb girdle muscular dystrophy 2B. Nat. Commun. 2019, 10, 2430. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Shan, T. The role of satellite and other functional cell types in muscle repair and regeneration. J. Muscle Res. Cell Motil. 2019, 40, 1–8. [Google Scholar] [CrossRef]
- Latroche, C.; Weiss-Gayet, M.; Muller, L.; Gitiaux, C.; Leblanc, P.; Liot, S.; Ben-Larbi, S.; Abou-Khalil, R.; Verger, N.; Bardot, P.; et al. Coupling between Myogenesis and Angiogenesis during Skeletal Muscle Regeneration Is Stimulated by Restorative Macrophages. Stem Cell Rep. 2017, 9, 2018–2033. [Google Scholar] [CrossRef]
- Varga, T.; Mounier, R.; Patsalos, A.; Gogolák, P.; Peloquin, M.; Csermely, A.; Pap, A.; Daniel, B.; Nagy, G.; Pintye, E.; et al. Macrophage PPARγ, a Lipid Activated Transcription Factor Controls the Growth Factor GDF3 and Skeletal Muscle Regeneration. Immun. 2016, 45, 1038–1051. [Google Scholar] [CrossRef]
- Jain, N.; Moeller, J.; Vogel, V. Mechanobiology of Macrophages: How Physical Factors Coregulate Macrophage Plasticity and Phagocytosis. Annu. Rev. Biomed. Eng. 2019, 21, 267–297. [Google Scholar] [CrossRef]
- Dick, S.A.; Macklin, J.; Nejat, S.; Momen, A.; Clemente-Casares, X.; AlThagafi, M.G.; Chen, J.; Kantores, C.; Hosseinzadeh, S.; Aronoff, L.; et al. Self-renewing resident cardiac macrophages limit adverse remodeling following myocardial infarction. Nat. Immunol. 2018, 20, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Schultze, J.L. Myocardial infarction cell by cell. Nat. Immunol. 2018, 20, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Bozec, A.; Soulat, D. Latest perspectives on macrophages in bone homeostasis. Pflügers Archiv - European Journal of Physiology 2017, 469, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.L.; T’Jonck, W.; Martens, L.; Todorov, H.; Sichien, D.; Soen, B.; Bonnardel, J.; De Prijck, S.; Vandamme, N.; Cannoodt, R.; et al. The Transcription Factor ZEB2 Is Required to Maintain the Tissue-Specific Identities of Macrophages. Immun. 2018, 49, 312–325. [Google Scholar] [CrossRef]
- Zhao, Y.; Zou, W.; Du, J.; Zhao, Y. The origins and homeostasis of monocytes and tissue-resident macrophages in physiological situation. J. Cell. Physiol. 2018, 233, 6425–6439. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, H.; Liu, Y.; Adams, S.; Eilken, H.; Stehling, M.; Corada, M.; Dejana, E.; Zhou, B.; Adams, R.H. Endothelial cells are progenitors of cardiac pericytes and vascular smooth muscle cells. Nat. Commun. 2016, 7, 12422. [Google Scholar] [CrossRef]
- Chopra, H.; Hung, M.K.; Kwong, D.L.; Zhang, C.F.; Pow, E.H.N. Insights into Endothelial Progenitor Cells: Origin, Classification, Potentials, and Prospects. Stem Cells Int. 2018, 2018, 1–24. [Google Scholar] [CrossRef]
- Kusumbe, A.P.; Ramasamy, S.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nat. 2014, 507, 323–328. [Google Scholar] [CrossRef]
- Ramasamy, S.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nat. 2014, 507, 376–380. [Google Scholar] [CrossRef]
- Sivaraj, K.K.; Adams, R.H. Blood vessel formation and function in bone. Dev. 2016, 143, 2706–2715. [Google Scholar] [CrossRef]
- Verma, M.; Asakura, Y.; Murakonda, B.S.R.; Pengo, T.; Latroche, C.; Chazaud, B.; McLoon, L.K.; Asakura, A. Muscle Satellite Cell Cross-Talk with a Vascular Niche Maintains Quiescence via VEGF and Notch Signaling. Cell Stem Cell 2018, 23, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.J.; Iruela-Arispe, M.L. NOTCH regulation of the endothelial cell phenotype. Curr. Opin. Hematol. 2018, 25, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Wosczyna, M.N.; Rando, T.A. A Muscle Stem Cell Support Group: Coordinated Cellular Responses in Muscle Regeneration. Dev. Cell 2018, 46, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, V.; Puri, P. Shaping Gene Expression by Landscaping Chromatin Architecture: Lessons from a Master. Mol. Cell 2018, 71, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C. Vitamin D Genomics: From In Vitro to In Vivo. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C.; Seuter, S.; Nurmi, T.; Tuomainen, T.-P.; Virtanen, J.K.; Neme, A. In vivo response of the human epigenome to vitamin D: A Proof-of-principle study. J. Steroid Biochem. Mol. Boil. 2018, 180, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Buskermolen, J.; Van Der Meijden, K.; Furrer, R.; Mons, D.-J.; Van Essen, H.W.; Heijboer, A.C.; Lips, P.; Jaspers, R.T.; Bravenboer, N. Effects of different training modalities on phosphate homeostasis and local vitamin D metabolism in rat bone. PeerJ 2019, 7, e6184. [Google Scholar] [CrossRef]
- Sahu, A.; Mamiya, H.; Shinde, S.N.; Cheikhi, A.; Winter, L.L.; Vo, N.V.; Stolz, D.; Roginskaya, V.; Tang, W.Y.; Croix, C.S.; et al. Age-related declines in α-Klotho drive progenitor cell mitochondrial dysfunction and impaired muscle regeneration. Nat. Commun. 2018, 9, 4859. [Google Scholar] [CrossRef]
- Del Real, A.; Riancho-Zarrabeitia, L.; López-Delgado, L.; Riancho, J.A. Epigenetics of Skeletal Diseases. Curr. Osteoporos. Rep. 2018, 16, 246–255. [Google Scholar] [CrossRef]
- Fernandez-Rebollo, E.; Eipel, M.; Seefried, L.; Hoffmann, P.; Strathmann, K.; Jakob, F.; Wagner, W. Primary Osteoporosis Is Not Reflected by Disease-Specific DNA Methylation or Accelerated Epigenetic Age in Blood. J. Bone Miner. Res. 2017, 33, 356–361. [Google Scholar] [CrossRef]
- Jakob, D.F.; Ebert, R.; Fernandez-Rebollo, E.; Bischof, O.; Wagner, W. Response to Letter to the Editor: Epigenetic Aging in Osteoporosis. J. Bone Miner. Res. 2018, 33, 1904–1905. [Google Scholar] [CrossRef] [PubMed]
- Benisch, P.; Schilling, T.; Klein-Hitpass, L.; Frey, S.P.; Seefried, L.; Raaijmakers, N.; Krug, M.; Regensburger, M.; Zeck, S.; Schinke, T.; et al. The Transcriptional Profile of Mesenchymal Stem Cell Populations in Primary Osteoporosis Is Distinct and Shows Overexpression of Osteogenic Inhibitors. PLOS ONE 2012, 7, e45142. [Google Scholar] [CrossRef] [PubMed]
- Axsom, J.E.; Libonati, J.R. Impact of parental exercise on epigenetic modifications inherited by offspring: A systematic review. Physiol. Rep. 2019, 7, e14287. [Google Scholar] [CrossRef] [PubMed]
- McGee, S.L.; Hargreaves, M. Epigenetics and Exercise. Trends Endocrinol. Metab. 2019, 30, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Koedijk, J.B.; Van Rijswijk, J.; Oranje, W.A.; Bergh, J.V.D.; Bours, S.P.; Savelberg, H.H.; Schaper, N.C. Sedentary behaviour and bone health in children, adolescents and young adults: a systematic review-supplementary presentation. Osteoporos. Int. 2017, 28, 3075–3076. [Google Scholar] [CrossRef] [PubMed]
- Winnard, A.; Scott, J.; Waters, N.; Vance, M.; Caplan, N. Effect of Time on Human Muscle Outcomes During Simulated Microgravity Exposure Without Countermeasures—Systematic Review. Front. Physiol. 2019, 10, 1046. [Google Scholar] [CrossRef]
- Rittweger, J.; Frost, H.; Schiessl, H.; Ohshima, H.; Alkner, B.; Tesch, P.; Felsenberg, D. Muscle atrophy and bone loss after 90 days’ bed rest and the effects of flywheel resistive exercise and pamidronate: Results from the LTBR study. Bone 2005, 36, 1019–1029. [Google Scholar] [CrossRef]
- Belavy, D.L.; Baecker, N.; Armbrecht, G.; Beller, G.; Buehlmeier, J.; Frings-Meuthen, P.; Rittweger, J.; Roth, H.J.; Heer, M.; Felsenberg, D. Serum sclerostin and DKK1 in relation to exercise against bone loss in experimental bed rest. J. Bone Miner. Metab. 2015, 34, 354–365. [Google Scholar] [CrossRef]
- Sun, Z.; Cao, X.; Zhang, Z.; Hu, Z.; Zhang, L.; Wang, H.; Zhou, H.; Li, D.; Zhang, S.; Xie, M.-J. Simulated microgravity inhibits L-type calcium channel currents partially by the up-regulation of miR-103 in MC3T3-E1 osteoblasts. Sci. Rep. 2015, 5, 8077. [Google Scholar] [CrossRef]
- Gao, Y.; Arfat, Y.; Wang, H.; Goswami, N. Muscle Atrophy Induced by Mechanical Unloading: Mechanisms and Potential Countermeasures. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Salanova, M.; Gelfi, C.; Moriggi, M.; Vasso, M.; Viganò, A.; Minafra, L.; Bonifacio, G.; Schiffl, G.; Gutsmann, M.; Felsenberg, D.; et al. Disuse deterioration of human skeletal muscle challenged by resistive exercise superimposed with vibration: evidence from structural and proteomic analysis. FASEB J. 2014, 28, 4748–4763. [Google Scholar] [CrossRef] [PubMed]
- Salanova, M.; Gambara, G.; Moriggi, M.; Vasso, M.; Ungethuem, U.; Belavy, D.L.; Felsenberg, D.; Cerretelli, P.; Gelfi, C.; Blottner, D. Vibration mechanosignals superimposed to resistive exercise result in baseline skeletal muscle transcriptome profiles following chronic disuse in bed rest. Sci. Rep. 2015, 5, 17027. [Google Scholar] [CrossRef] [PubMed]
- Vilchinskaya, N.A.; Krivoi, I.I.; Shenkman, B.S. AMP-Activated Protein Kinase as a Key Trigger for the Disuse-Induced Skeletal Muscle Remodeling. Int. J. Mol. Sci. 2018, 19, 3558. [Google Scholar] [CrossRef] [PubMed]
- Timmer, L.T.; Hoogaars, W.M.H.; Jaspers, R.T. The Role of IGF-1 Signaling in Skeletal Muscle Atrophy. Advances in Experimental Medicine and Biology 2018, 109–137. [Google Scholar]
- Tian, F.; Wang, Y.; Bikle, D.D. IGF-1 signaling mediated cell-specific skeletal mechano-transduction. J. Orthop. Res. 2017, 36, 576–583. [Google Scholar] [CrossRef]
- Wuest, S.; Gantenbein, B.; Ille, F.; Egli, M. Electrophysiological experiments in microgravity: lessons learned and future challenges. npj Microgravity 2018, 4, 7. [Google Scholar] [CrossRef]
- Schmidt-Bleek, K.; Schell, H.; Schulz, N.; Hoff, P.; Perka, C.; Buttgereit, F.; Volk, H.-D.; Lienau, J.; Duda, G.N. Inflammatory phase of bone healing initiates the regenerative healing cascade. Cell Tissue Research 2011, 347, 567–573. [Google Scholar] [CrossRef]
- Buckinx, F.; Landi, F.; Cesari, M.; Fielding, R.A.; Visser, M.; Engelke, K.; Maggi, S.; Dennison, E.M.; Al-Daghri, N.M.; Allepaerts, S.; et al. Pitfalls in the measurement of muscle mass: a need for a reference standard. J. Cachex- Sarcopenia Muscle 2018, 9, 269–278. [Google Scholar] [CrossRef]
- Baumgartner, R.N.; Koehler, K.M.; Gallagher, D.; Romero, L.; Heymsfield, S.; Ross, R.R.; Garry, P.J.; Lindeman, R.D. Epidemiology of sarcopenia among the elderly in New Mexico. Am. J. Epidemiology 1998, 147, 755–763. [Google Scholar] [CrossRef]
- Kyle, U.G.; Bosaeus, I.; De Lorenzo, A.D.; Deurenberg, P.; Elia, M.; Gómez, J.M.; Heitmann, B.L.; Kent-Smith, L.; Melchior, J.-C.; Pirlich, M.; et al. Bioelectrical impedance analysis?part I: review of principles and methods. Clin. Nutr. 2004, 23, 1226–1243. [Google Scholar] [CrossRef]
- Karampinos, D.; Baum, T.; Nardo, L.; Alizai, H.; Yu, H.; Carballido-Gamio, J.; Yap, S.P.; Shimakawa, A.; Link, T.M.; Majumdar, S. Characterization of the regional distribution of skeletal muscle adipose tissue in type 2 diabetes using chemical shift-based water/fat separation. J. Magn. Reson. Imaging 2011, 35, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Grimm, A.; Meyer, H.; Nickel, M.D.; Nittka, M.; Raithel, E.; Chaudry, O.; Friedberger, A.; Uder, M.; Kemmler, W.; Engelke, K.; et al. Repeatability of Dixon magnetic resonance imaging and magnetic resonance spectroscopy for quantitative muscle fat assessments in the thigh. J. Cachex- Sarcopenia Muscle 2018, 9, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Wu, E.X. Diffusion-weighted magnetic resonance spectroscopy: A novel approach to investigate intramyocellular lipids. Magn. Reson. Med. 2011, 66, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Boesch, C.; Slotboom, J.; Hoppeler, H.; Kreis, R. In vivo determination of intra-myocellular lipids in human muscle by means of localized 1H-MR-spectroscopy. Magn. Reson. Med. 1997, 37, 484–493. [Google Scholar] [CrossRef]
- Engelke, K.; Museyko, O.; Wang, L.; Laredo, J.-D. Quantitative analysis of skeletal muscle by computed tomography imaging-State of the art. J. Orthop. Transl. 2018, 15, 91–103. [Google Scholar] [CrossRef]
- Calella, P.; Gallè, F.; Fornelli, G.; Liguori, G.; Valerio, G. Type 1 diabetes and body composition in youth: A systematic review. Diabetes/Metabolism Res. Rev. 2019, 36, e3211. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Hsu, C.-Y.; Liu, J.-F. Effects of dietary and exercise intervention on weight loss and body composition in obese postmenopausal women. Menopause 2018, 25, 772–782. [Google Scholar] [CrossRef]
- Hopkins, J.; Sawyer, M. A review of body composition and pharmacokinetics in oncology. Expert Rev. Clin. Pharmacol. 2017, 10, 947–956. [Google Scholar] [CrossRef]
- Tannir, H.; Itani, L.; Kreidieh, D.; El Masri, D.; Traboulsi, S.; El Ghoch, M. Body composition in adolescents and young adults with anorexia nervosa: A clinical review. Curr. Rheumatol. Rev. 2019, 15, 1. [Google Scholar] [CrossRef]
- Addison, O.; Drummond, M.J.; LaStayo, P.C.; Dibble, L.E.; Wende, A.R.; McClain, D.; Marcus, R. Intramuscular fat and inflammation differ in older adults: the impact of frailty and inactivity. J. Nutr. Heal. Aging 2014, 18, 532–538. [Google Scholar] [CrossRef]
- Addison, O.; Marcus, R.; LaStayo, P.C.; Ryan, A.S. Intermuscular Fat: A Review of the Consequences and Causes. Int. J. Endocrinol. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Loenneke, J.P.; Buckner, S.L.; Dankel, S.J.; Abe, T. Exercise-Induced Changes in Muscle Size do not Contribute to Exercise-Induced Changes in Muscle Strength. Sports Med. 2019, 49, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.E.; Bucarey, J.L.; Espinosa, A. Muscle Lipid Metabolism: Role of Lipid Droplets and Perilipins. J. Diabetes Res. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachex- Sarcopenia Muscle 2017, 9, 3–19. [Google Scholar] [CrossRef] [PubMed]
- A McGregor, R.; Cameron-Smith, D.; Poppitt, S. It is not just muscle mass: a review of muscle quality, composition and metabolism during ageing as determinants of muscle function and mobility in later life. Longev. Heal. 2014, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Malkov, S.; Cawthon, P.M.; Peters, K.W.; A Cauley, J.; A Murphy, R.; Visser, M.; Wilson, J.; Harris, T.; Satterfield, S.; Cummings, S.; et al. Hip Fractures Risk in Older Men and Women Associated With DXA-Derived Measures of Thigh Subcutaneous Fat Thickness, Cross-Sectional Muscle Area, and Muscle Density. J. Bone Miner. Res. 2015, 30, 1414–1421. [Google Scholar] [CrossRef]
- McLean, R.R.; Kiel, D.P.; Berry, S.D.; Broe, K.E.; Zhang, X.; Cupples, L.A.; Hannan, M.T. Lower Lean Mass Measured by Dual-Energy X-ray Absorptiometry (DXA) is Not Associated with Increased Risk of Hip Fracture in Women: The Framingham Osteoporosis Study. Calcif. Tissue Int. 2018, 103, 16–23. [Google Scholar] [CrossRef]
- Schott, A.-M.; Cormier, C.; Hans, D.; Favier, F.; Hausherr, E.; Dargent-Molina, P.; Delmas, P.D.; Ribot, C.; Sebert, J.L.; Bréart, G.; et al. How Hip and Whole-Body Bone Mineral Density Predict Hip Fracture in Elderly Women: The EPIDOS Prospective Study. Osteoporos. Int. 1998, 8, 247–254. [Google Scholar] [CrossRef]
- Zaslavsky, O.; Li, W.; Going, S.; Datta, I.; Snetselaar, L.; Zelber-Sagi, S. Association between body composition and hip fractures in older women with physical frailty. Geriatr. Gerontol. Int. 2016, 17, 898–904. [Google Scholar] [CrossRef]
- Lang, T.F.T.; A Cauley, J.; Tylavsky, F.; Bauer, U.; Cummings, S.; Harris, T.B. Computed Tomographic Measurements of Thigh Muscle Cross-Sectional Area and Attenuation Coefficient Predict Hip Fracture: The Health, Aging, and Body Composition Study. J. Bone Miner. Res. 2009, 25, 513–519. [Google Scholar] [CrossRef]
- Lang, T.F.T.; Koyama, A.; Li, C.; Li, J.; Lu, Y.; Saeed, I.; Gazze, E.; Keyak, J.; Harris, T.; Cheng, X. Pelvic body composition measurements by quantitative computed tomography: Association with recent hip fracture. Bone 2008, 42, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Mühlberg, A.; Museyko, O.; Laredo, J.-D.; Engelke, K. A reproducible semi-automatic method to quantify the muscle-lipid distribution in clinical 3D CT images of the thigh. PLOS ONE 2017, 12, e0175174. [Google Scholar] [CrossRef] [PubMed]
- Daguet, E.; Jolivet, E.; Bousson, V.; Boutron, C.; Dahmen, N.; Bergot, C.; Vicaut, E.; Laredo, J. Fat Content of Hip Muscles. J. Bone Jt. Surgery-American Vol. 2011, 93, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Mühlberg, A.; Museyko, O.; Bousson, V.; Pottecher, P.; Laredo, J.-D.; Engelke, K. Three-dimensional Distribution of Muscle and Adipose Tissue of the Thigh at CT: Association with Acute Hip Fracture. Radiol. 2019, 290, 426–434. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inflammatory Phase Upregulated Genes after 45′ Exercise | Upregulated Genes 2 h after 45′ Exercise | Upregulated Genes after 12 Weeks of Intermittent Exercise |
|---|---|---|
| Pourteymour et al. [62] | ||
| Interleukin 6 | Interleukin 6 receptor | Secreted frizzled-related protein 5 |
| Interleukin 8 | Colony stimulating factor 3 receptor | Secreted frizzled-related protein 2 |
| Interleukin 1, beta | TNF receptor superfamily member 8 | |
| Prostaglandin–endoperoxide synthase 2 | Prostaglandin I2 (prostacyclin) receptor | |
| Chemokine (C-X-C motif) ligand 1 | Tumor necrosis factor receptor | Chemokine (C-C motif) ligand 21 |
| Chemokine (C-C motif) ligand 8 | Complement component 8 | Collagen, type I, alpha 1 |
| Chemokine (C-X-C motif) ligand 2 | Plasminogen | Collagen, type III, alpha 1 |
| Chemokine (C-C motif) ligand 2 | Stanniocalcin 2 | Collagen, type IV, alpha 1 |
| Chemokine (C-X-C motif) ligand 3 | Lipocalin 10 | Collagen, type IV, alpha 2 |
| Chemokine (C-X3-C motif) ligand 1 | Lipocalin 6 | Collagen, type VI, alpha 6 |
| Leukemia inhibitory factor | Lysyl oxidase-like 2 | |
| Serum amyloid A1 | Matrix-remodeling associated 5 | |
| Serum amyloid A2 | Osteoglycin | |
| Angiopoietin-like 4 | Angiopoietin-like 4 | Biglycan |
| CYR61/CCN1 | Angiopoietin-like 2 | |
| Connective tissue growth factor/CCN2 | ||
| Vascular endothelial growth factor A | ||
| Thrombospondin 1 | Thrombospondin 1 | Thrombospondin 4 |
| Fibroblast growth factor 6 | Fibroblast growth factor 6 | Insulin-like growth factor 2 |
| Fibroblast growth factor 18 | ||
| Matrix metallopeptidase 19 | Serpin peptidase inhibitor, clade F, member 2 | |
| ADAM metallopeptidase with thrombospondin type 1 motif 4 | Serpin peptidase inhibitor, clade A, member 3 | |
| ADAM metallopeptidase with thrombospondin type 1 motif, 1 | Serpin peptidase inhibitor, clade A, member 1 | |
| ADAM metallopeptidase with thrombospondin type 1 motif 9 | ||
| VOIs | Parameters | |
|---|---|---|
| DXA | total body appendicular skeleton abdomen: estimation of SAT and VAT | lean mass (g) fat mass (g) |
| BIA | total body appendicular skeleton abdomen: estimation of SAT and VAT | estimates of lean and fat mass (g) based on sex, age, and ethnicity-specific equations |
| MRI T1 | thigh abdomen paraspinal muscle | SAT, VAT area/volume (cm2/cm3) IMAT area/volume (cm2/cm3) muscle area/volume (cm2/cm3) |
| MRI Dixon | whole body thigh paraspinal muscle | FF (%) WF (%) |
| CT | thigh abdomen paraspinal muscle | SAT, VAT area/volume (cm2/cm3) IMAT area/volume (cm2/cm3) muscle area/volume (cm2/cm3) muscle density (HU) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrmann, M.; Engelke, K.; Ebert, R.; Müller-Deubert, S.; Rudert, M.; Ziouti, F.; Jundt, F.; Felsenberg, D.; Jakob, F. Interactions between Muscle and Bone—Where Physics Meets Biology. Biomolecules 2020, 10, 432. https://doi.org/10.3390/biom10030432
Herrmann M, Engelke K, Ebert R, Müller-Deubert S, Rudert M, Ziouti F, Jundt F, Felsenberg D, Jakob F. Interactions between Muscle and Bone—Where Physics Meets Biology. Biomolecules. 2020; 10(3):432. https://doi.org/10.3390/biom10030432
Chicago/Turabian StyleHerrmann, Marietta, Klaus Engelke, Regina Ebert, Sigrid Müller-Deubert, Maximilian Rudert, Fani Ziouti, Franziska Jundt, Dieter Felsenberg, and Franz Jakob. 2020. "Interactions between Muscle and Bone—Where Physics Meets Biology" Biomolecules 10, no. 3: 432. https://doi.org/10.3390/biom10030432
APA StyleHerrmann, M., Engelke, K., Ebert, R., Müller-Deubert, S., Rudert, M., Ziouti, F., Jundt, F., Felsenberg, D., & Jakob, F. (2020). Interactions between Muscle and Bone—Where Physics Meets Biology. Biomolecules, 10(3), 432. https://doi.org/10.3390/biom10030432