Metabolic and Proteomic Perspectives of Augmentation of Nutritional Contents and Plant Defense in Vigna unguiculata

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
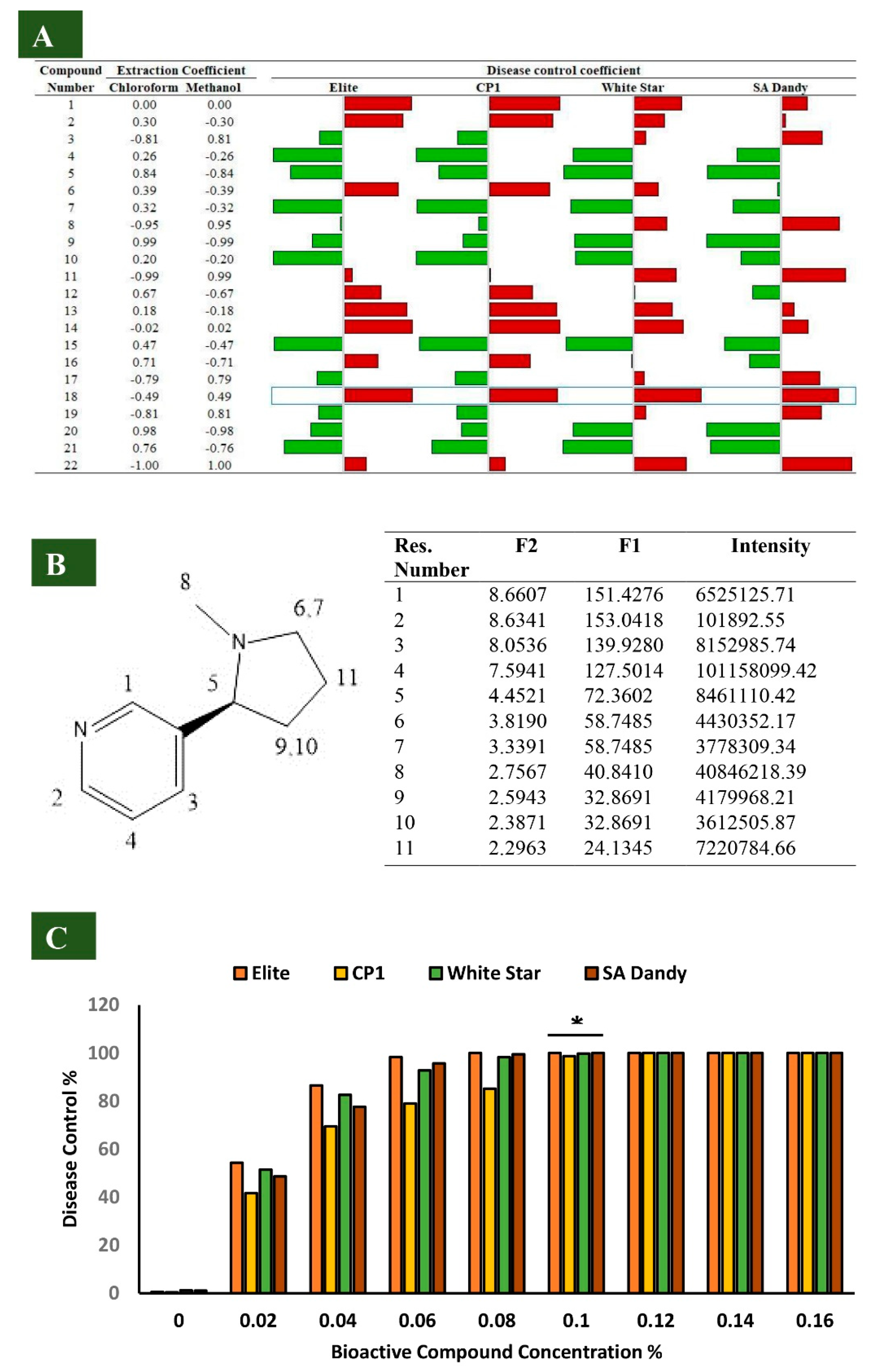
3.1. Disease Incidence (%) of Charcoal Rot and Percentage Disease Control by Alstonia Extracts
3.2. Identification of Alstonia Metabolites in Bioactive Sub-Fractions
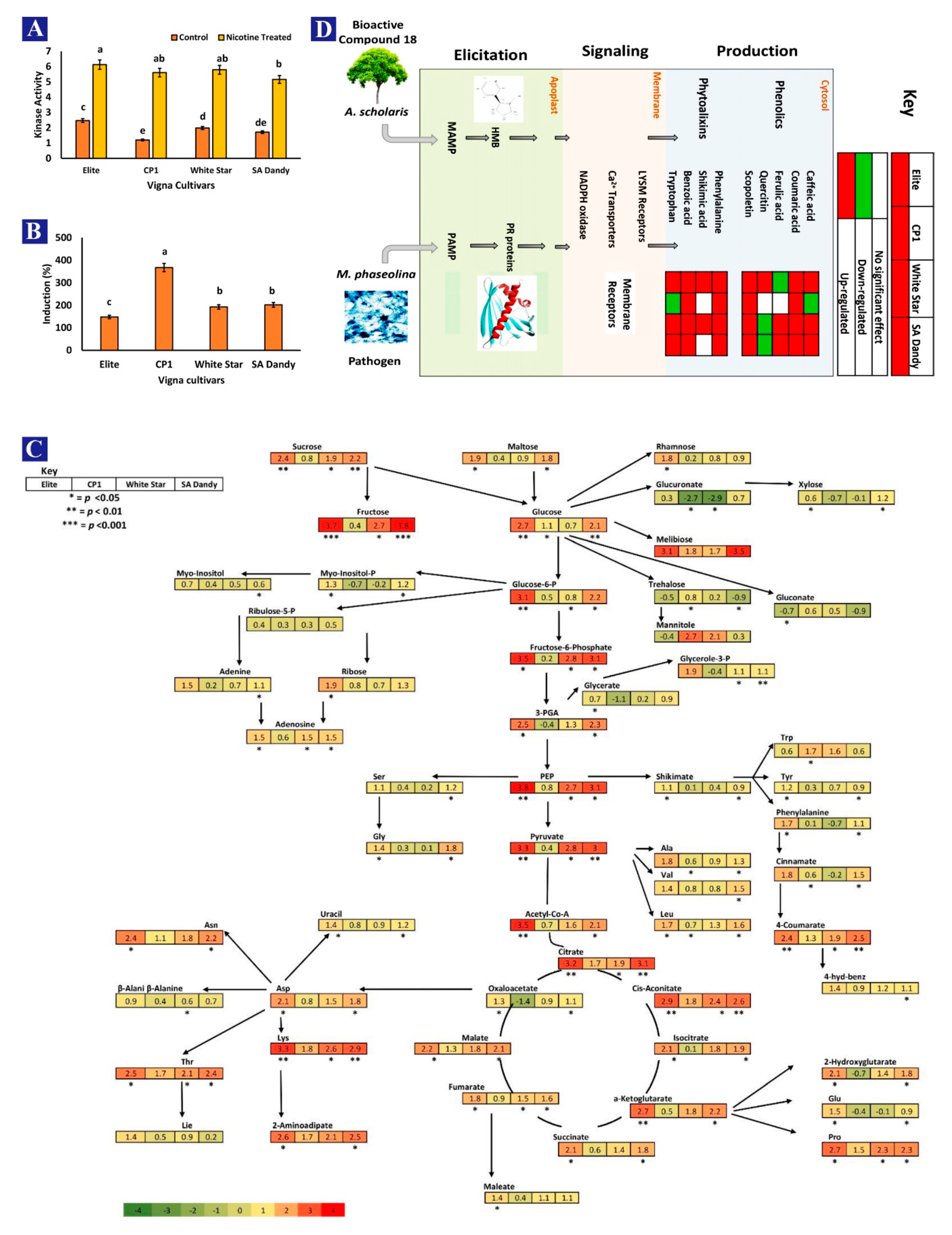
3.3. Nicotine Effect on Nutritional Contents, Physiological Factors, Enzymes, and Proline Contents
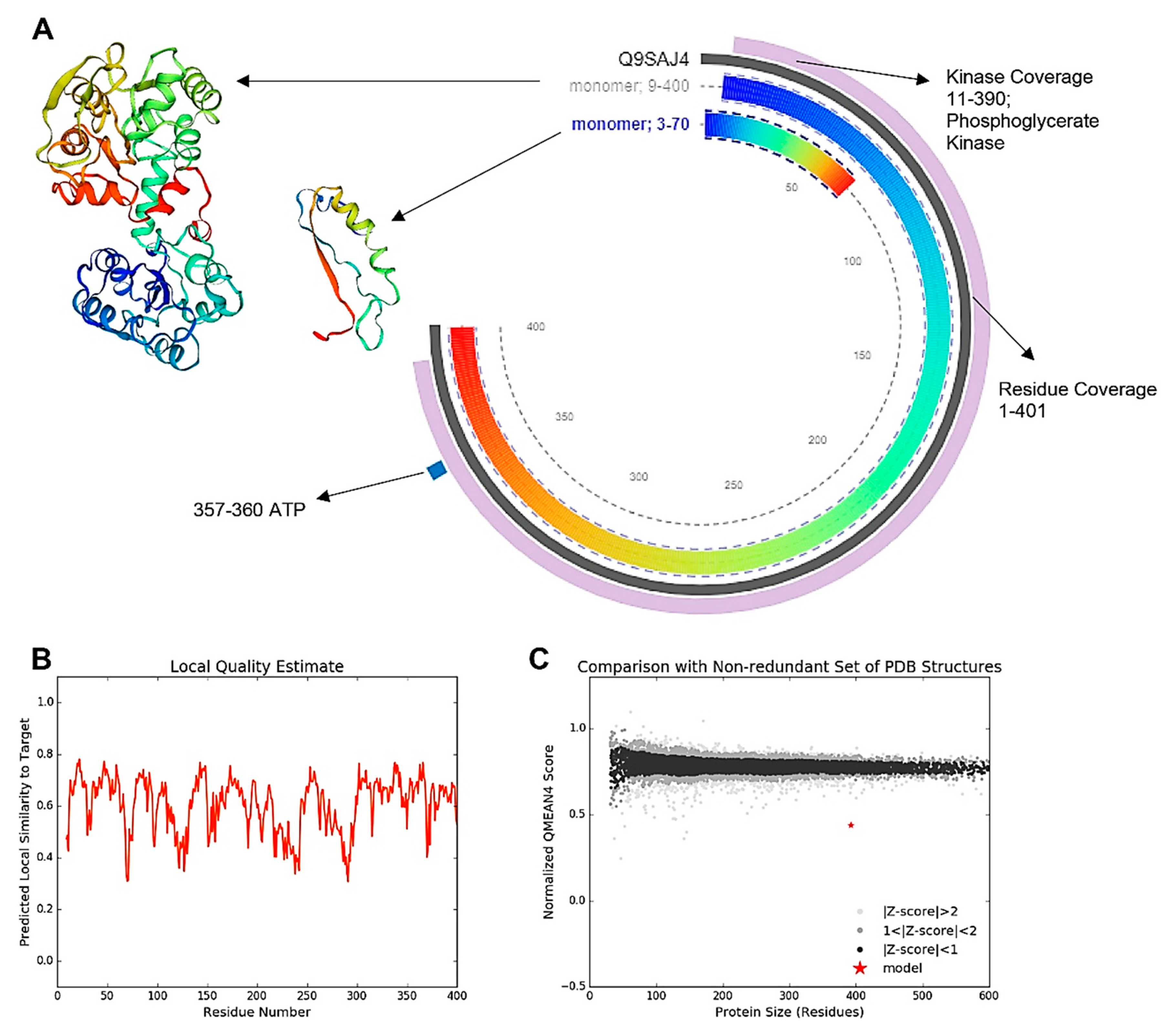
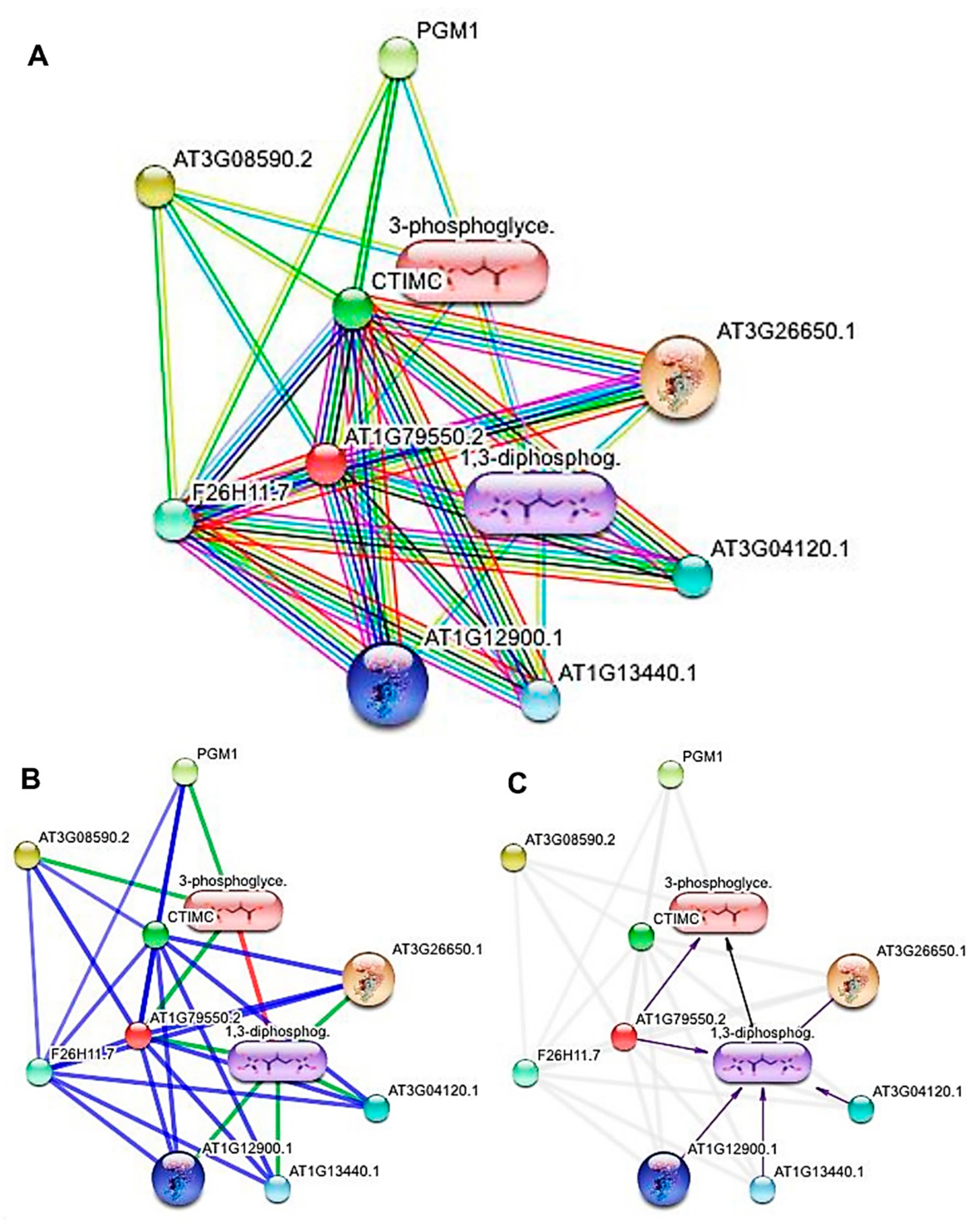
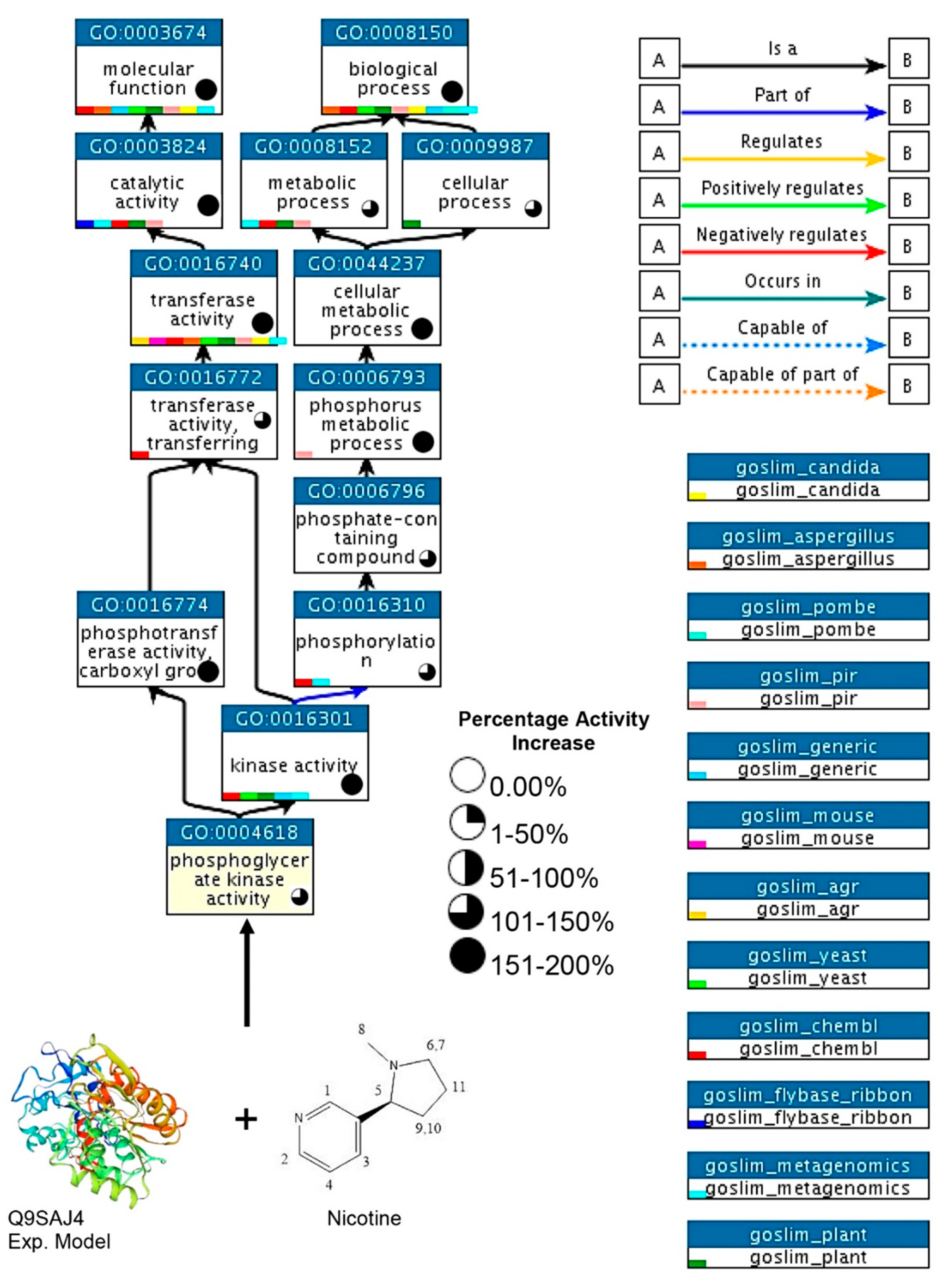
3.4. Modeling of the Most Active Protein and Its Interactions
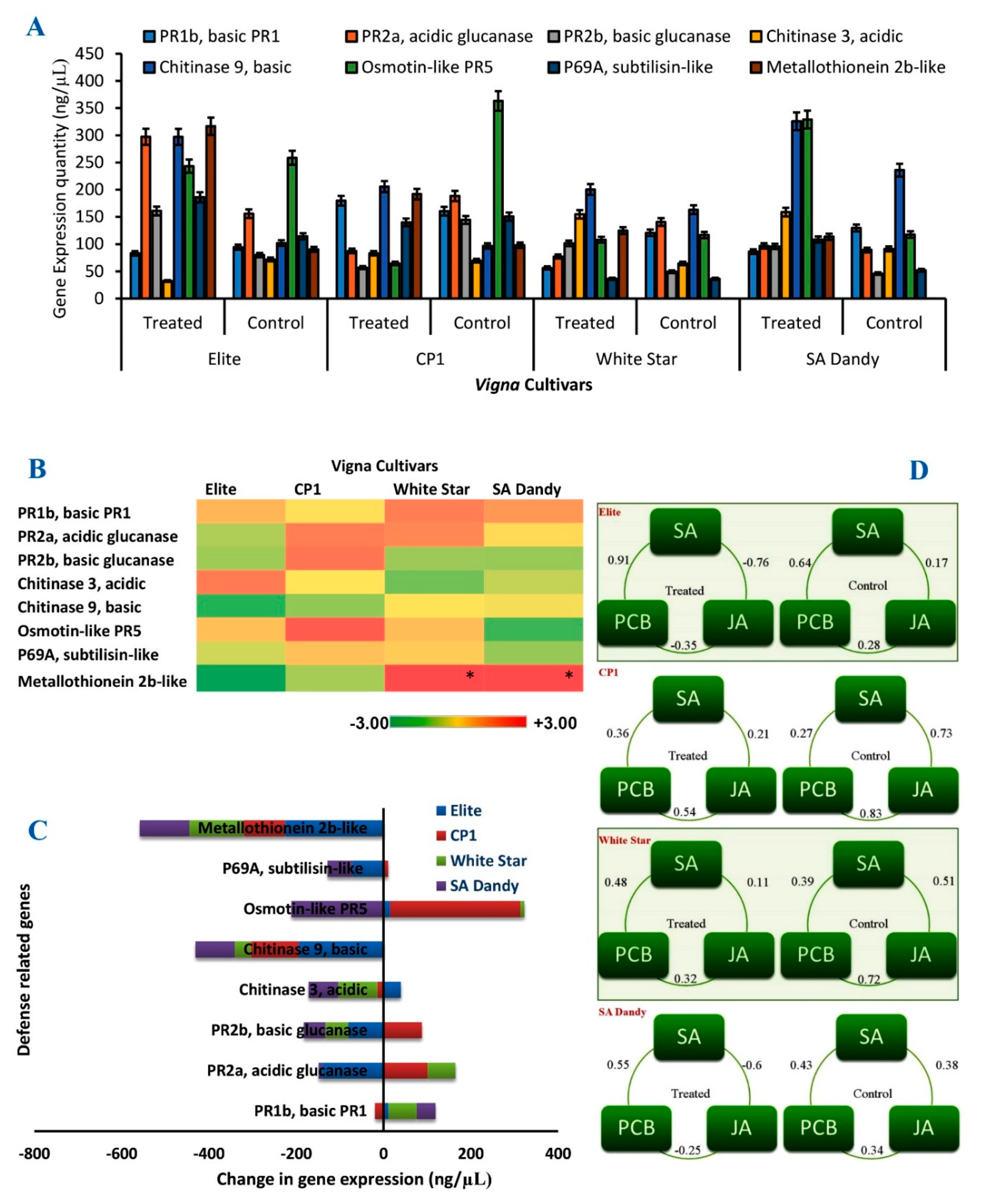
3.5. Expression of Defense-Related Genes in V. unguiculata
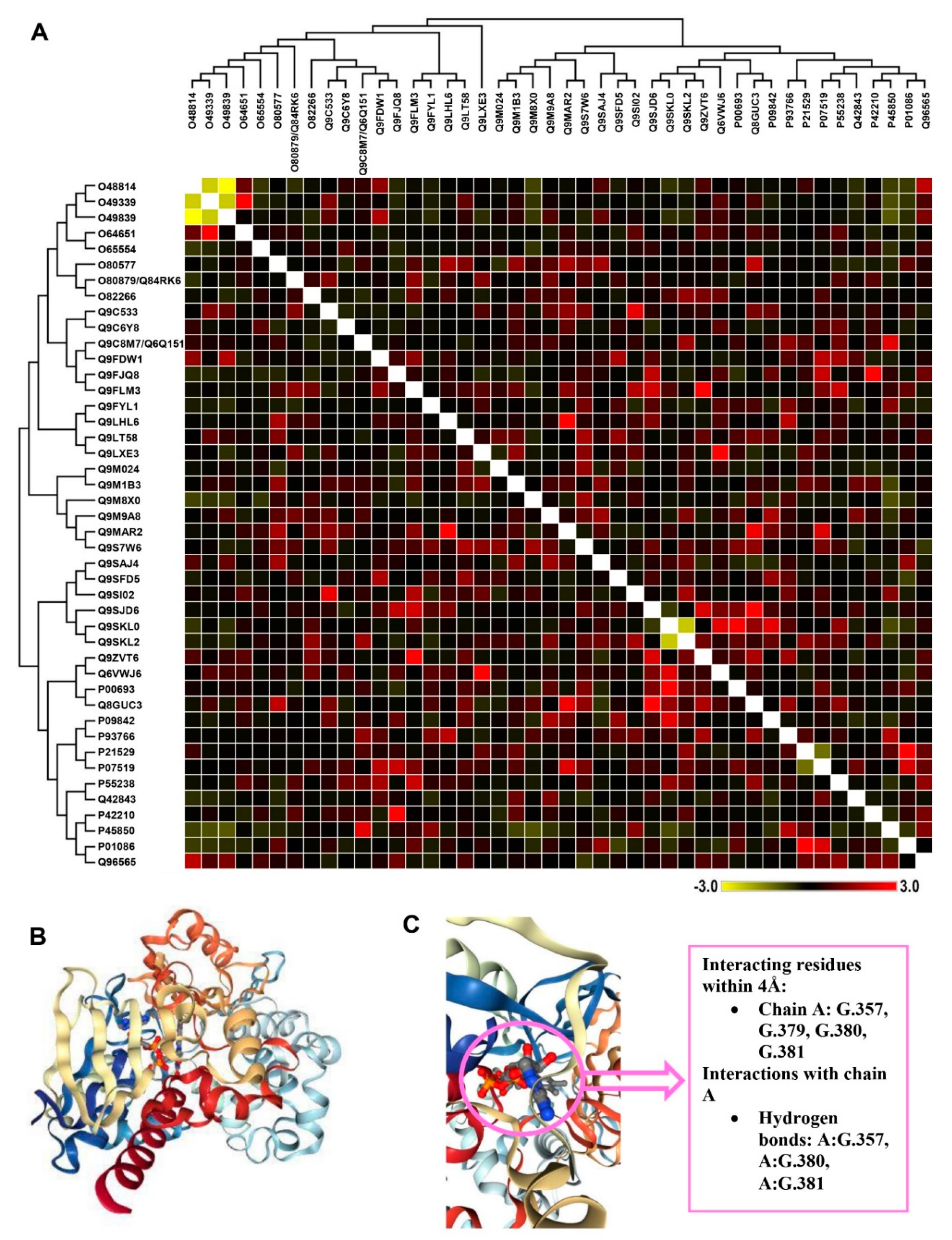
3.6. Correlation between A. scholaris Extracts and Defense-Related Pathways
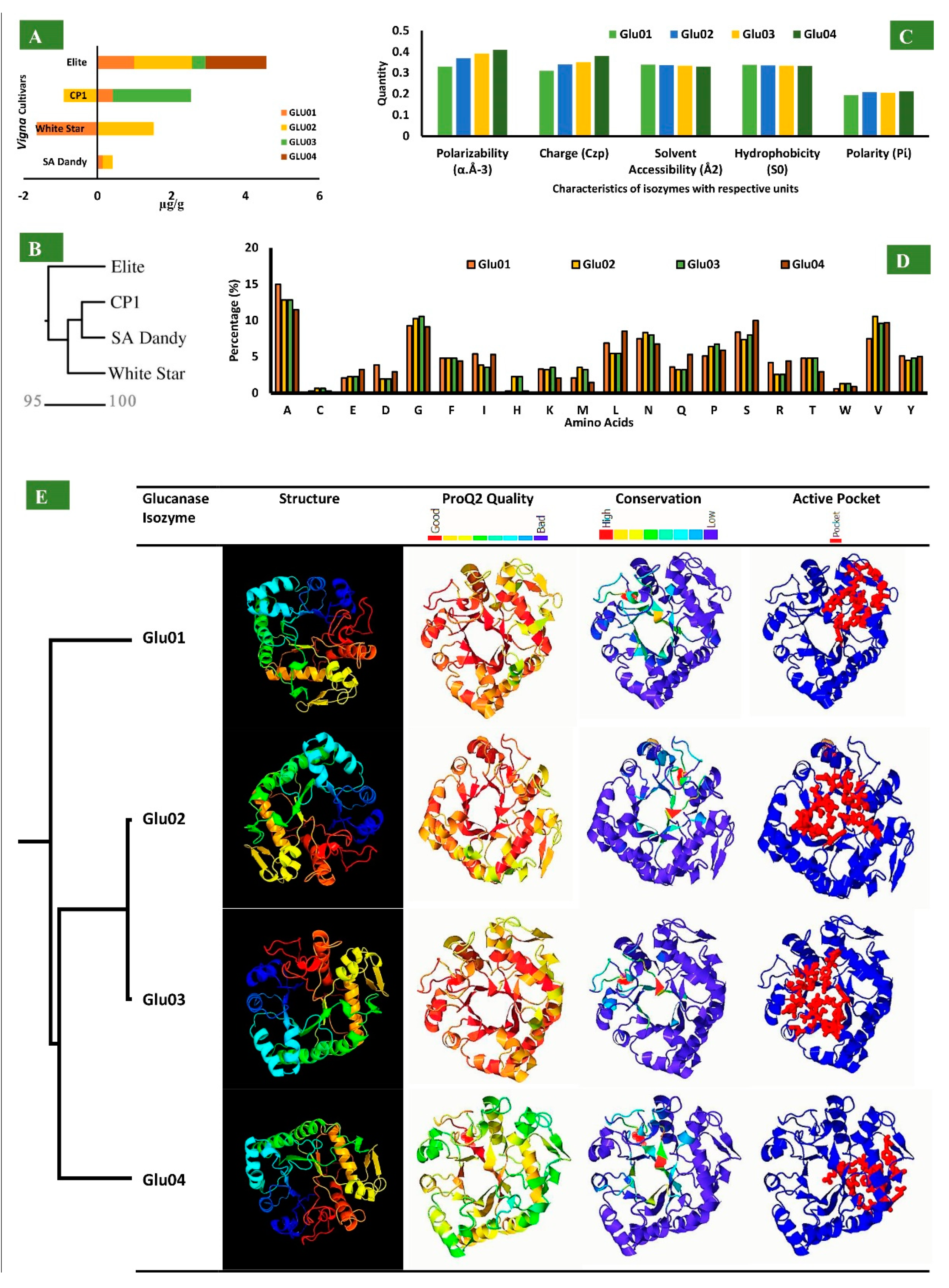
3.7. Glucanase Isozymes and Their Physiochemical Analysis
3.8. Physiological Interactions
3.9. Activity of PGK3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Greenhalgh, P. The Market Potential for Bambara Groundnut; NRI, Chattam Maritime: Kent, UK, 2000. [Google Scholar]
- Aikins, S.H.M.; Afuakwa, J.J. Growth and dry matter yield responses of cowpea to different sowing depths. ARPN J. Agric. Biol. Sci. 2008, 3, 50–54. [Google Scholar]
- Kausar, T.; Sher, H.S.I.; Ainee, A.; Norin, S. Preparation and quality evaluation of carrot and carrot pomace jam. J. Innov. Biol-Res. 2017, 1, 8–12. [Google Scholar]
- Shao, A.; Drewnowski, A.; Willcox, D.C.; Krämer, L.; Lausted, C.; Eggersdorfer, M.; Mathers, J.; Bell, J.D.; Randolph, R.K.; Witkamp, R.; et al. Optimal nutrition and the ever-changing dietary landscape: A conference report. Eur. J. Nutr. 2017, 56 (Suppl. 1), S1–S21. [Google Scholar] [CrossRef] [PubMed]
- Sogari, G.; Velez-Argumedo, C.; Gómez, M.I.; Mora, C. College Students and Eating Habits: A Study Using an Ecological Model for Healthy Behavior. Nutrients 2018, 10, 1823. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Mason-D’Croz, D.; Robinson, S. Food system consequences of a fungal disease epidemic in a major crop. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150467. [Google Scholar] [CrossRef]
- Roberts, P.A.; Matthews, W.C.; Ehlers, J.D. Root-Knot Nematode Resistant Cowpea Cover Crops in Tomato Production Systems. Agron. J. 2005, 97, 1626–1635. [Google Scholar] [CrossRef]
- Baliga, M.S. Alstonia scholaris Linn R Br in the Treatment and Prevention of Cancer: Past, Present, and Future. Integr. Cancer Ther. 2010, 9, 261–269. [Google Scholar] [CrossRef]
- Pratap, B.; Chakraborthy, G.S.; Mogha, N. Complete aspects of Alstonia scholaris. Int. J. Pharm. Tech. Res. 2013, 5, 17–26. [Google Scholar]
- Huang, K.; Mellor, E.K.; Paul, S.N.; Lawson, M.J.; Mackey, A.J.; Timko, M.P. Global changes in gene expression during compatible and incompatible interactions of cowpea (Vigna unguiculata L.) with the root parasitic angiosperm Striga gesnerioides. BMC Genom. 2012, 13, 402. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef]
- Gupta, P.; Ravi, I.; Sharma, V. Induction of β-1,3-glucanase and chitinase activity in the defense response of Eruca sativa plants against the fungal pathogen Alternaria brassicicola. J. Plant Interact. 2013, 8, 155–161. [Google Scholar] [CrossRef]
- Pareek, S.S.; Ravi, I.; Sharma, V. Induction of β-1,3-glucanase and chitinase in Vigna aconitifolia inoculated with Macrophomina phaseolina. J. Plant Interact. 2014, 9, 434–439. [Google Scholar] [CrossRef][Green Version]
- Rose, T.L.; Conceição, A.D.S.; Xavier-Filho, J.; Okorokov, L.A.; Fernandes, K.V.S.; Marty, F.; Marty-Mazars, D.; Carvalho, A.O.; Gomes, V.M. Defense Proteins from Vigna Unguiculata Seed Exudates: Characterization and Inhibitory Activity Against Fusarium Oxysporum. Plant Soil 2006, 286, 181–191. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Filho, J.H.A.; Grangeiro, T.B.; Gondim, D.M.F.; Segalin, J.; Pinto, P.M.; Carlini, C.R.R.S.; Silva, F.D.A.; Lobo, M.D.P.; Costa, J.H.; et al. Enhanced synthesis of antioxidant enzymes, defense proteins and leghemoglobin in rhizobium-free cowpea roots after challenging with Meloidogyne incognita. Proteomes 2014, 2, 527–549. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Akram, W.; Shahzadi, I.; Wang, R.; Hu, D.; Bashir, Z.; Jaleel, W.; Ahmed, S.; Tariq, W.; Li, G.; et al. Benzenedicarboxylic acid upregulates O48814 and Q9FJQ8 for improved nutritional contents of tomato and low risk of fungal attack. J. Sci. Food Agric. 2019, 99, 6139–6154. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Zahin, M.; Ahmad, I.; Owais, M.; Khan, M.S.A.; Bansal, S.S.; Farooq, S. Antifungal Activity of Medicinal Plant Extracts and Phytocompounds: A review. In Combating Fungal Infections; Ahmad, I., Owais, M., Shahid, M., Aqil, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 449–484. [Google Scholar]
- Javanmardi, J.; Stushnoff, C.; Locke, E.; Vivanco, J.M. Antioxidant activity and total phenolic content of Iranian Ocimum accessions. Food Chem. 2003, 83, 547–550. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, Isolation, and Identification of Bioactive Compounds from Plant Extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Khan, K.A.; Shoaib, A.; Awan, Z.A.; Basit, A.; Hussain, M. Macrophomina phaseolina alters the biochemical pathway in Vigna radiata chastened by Zn2+ and FYM to improve plant growth. J. Plant Interact. 2018, 13, 131–140. [Google Scholar] [CrossRef]
- Sharma, I.; Kumari, N.; Sharma, V. Defense gene expression in Sorghum bicolor against Macrophomina phaseolina in leaves and roots of susceptible and resistant cultivars. J. Plant Interact. 2014, 9, 315–323. [Google Scholar] [CrossRef]
- Ahmad, A.; Akram, W.; Yasin, N.A.; Khan, W.U.; Noreen, S.; Ashraf, Y.; Awan, S.A. An insight into the associated microflora of Parthenium hysterophorus L. J. Innov. Bio-Res. 2017, 1, 1–7. [Google Scholar]
- Voet, J.G.; Voet, C.W.P.D. Fundamentals of Biochemistry. In Life at the Molecular Level, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; Volume 499. [Google Scholar]
- Auerbach, G.; Huber, R.; Grättinger, M.; Zaiss, K.; Schurig, H.; Jaenicke, R.; Jacob, U. Closed structure of phosphoglycerate kinase from Thermotoga maritima reveals the catalytic mechanism and determinants of thermal stability. Structure 1997, 5, 1475–1483. [Google Scholar] [CrossRef]
- Vas, M.; Varga, A.; Graczer, E. Insight into the mechanism of domain movements and their role in enzyme function: Example of 3-phosphoglycerate kinase. Curr. Protein Pept. Sci. 2010, 11, 118–147. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, P.; Chaloin, L.; Roy, B.; Barman, T.; Bowler, M.W.; Lionne, C. Interaction of Human 3-Phosphoglycerate Kinase with Its Two Substrates: Is Substrate Antagonism a Kinetic Advantage? J. Mol. Biol. 2011, 409, 742–757. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Filippova, E.V.; Tkaczuk, K.L.; Dworzynski, P.; Chruszcz, M.; Porebski, P.J.; Wawrzak, Z.; Onopriyenko, O.; Kudritska, M.; Grimshaw, S.; et al. Crystal structures of putative phosphoglycerate kinases from B. anthracis and C. jejuni. J. Struct. Funct. Genom. 2012, 13, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Doxey, A.C.; Yaish, M.W.F.; Moffatt, B.A.; Griffith, M.; McConkey, B.J. Functional divergence in the Arabidopsis beta-1,3-glucanase gene family inferred by phylogenetic reconstruction of expression states. Mol. Biol. Evol. 2007, 24, 1045–1055. [Google Scholar] [CrossRef]
- Dyakov, Y.T.; Dzhavakhiya, V.G.; Korpela, T. Comprehensive and Molecular Phytopathology. In Chapter 10—Immune Response; Dzhavakhiya, V.G., Ozeretskovskaya, O.L., Zinovyeva, S.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 265–314. [Google Scholar]
- Vidhyasekaran, P. Concise Encyclopedia of Plant Pathology. In Chapter 34: Pathogenesis-Related Proteins; CRC Press: Boca Raton, FL, USA, 2004; pp. 523–536. [Google Scholar]
- Vosloh, D. Subcellular Compartmentation of Primary Carbon Metabolism in Mesophyll Cells of Arabidopsis thaliana. Ph.D. Thesis, University of Potsdam, Potsdam, Germany, 2011. [Google Scholar]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Huffman, R.D. Investigations into Nutritional Quality, Commercial Productivity, and Genetic Purity of Maize Grain for Organic Production Systems. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2017. Available online: https://lib.dr.iastate.edu/etd/15322 (accessed on 25 November 2019).










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Khan, T.A.; Mubeen, S.; Shahzadi, I.; Akram, W.; Saeed, T.; Bashir, Z.; Wang, R.; Alam, M.; Ahmed, S.; et al. Metabolic and Proteomic Perspectives of Augmentation of Nutritional Contents and Plant Defense in Vigna unguiculata. Biomolecules 2020, 10, 224. https://doi.org/10.3390/biom10020224
Ahmad A, Khan TA, Mubeen S, Shahzadi I, Akram W, Saeed T, Bashir Z, Wang R, Alam M, Ahmed S, et al. Metabolic and Proteomic Perspectives of Augmentation of Nutritional Contents and Plant Defense in Vigna unguiculata. Biomolecules. 2020; 10(2):224. https://doi.org/10.3390/biom10020224
Chicago/Turabian StyleAhmad, Aqeel, Tanveer Alam Khan, Samavia Mubeen, Iqra Shahzadi, Waheed Akram, Taiba Saeed, Zoobia Bashir, Rui Wang, Mufid Alam, Shakeel Ahmed, and et al. 2020. "Metabolic and Proteomic Perspectives of Augmentation of Nutritional Contents and Plant Defense in Vigna unguiculata" Biomolecules 10, no. 2: 224. https://doi.org/10.3390/biom10020224
APA StyleAhmad, A., Khan, T. A., Mubeen, S., Shahzadi, I., Akram, W., Saeed, T., Bashir, Z., Wang, R., Alam, M., Ahmed, S., Hu, D., Li, G., & Wu, T. (2020). Metabolic and Proteomic Perspectives of Augmentation of Nutritional Contents and Plant Defense in Vigna unguiculata. Biomolecules, 10(2), 224. https://doi.org/10.3390/biom10020224