Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity

 ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Material and Methods
2.1. Materials and Reagents
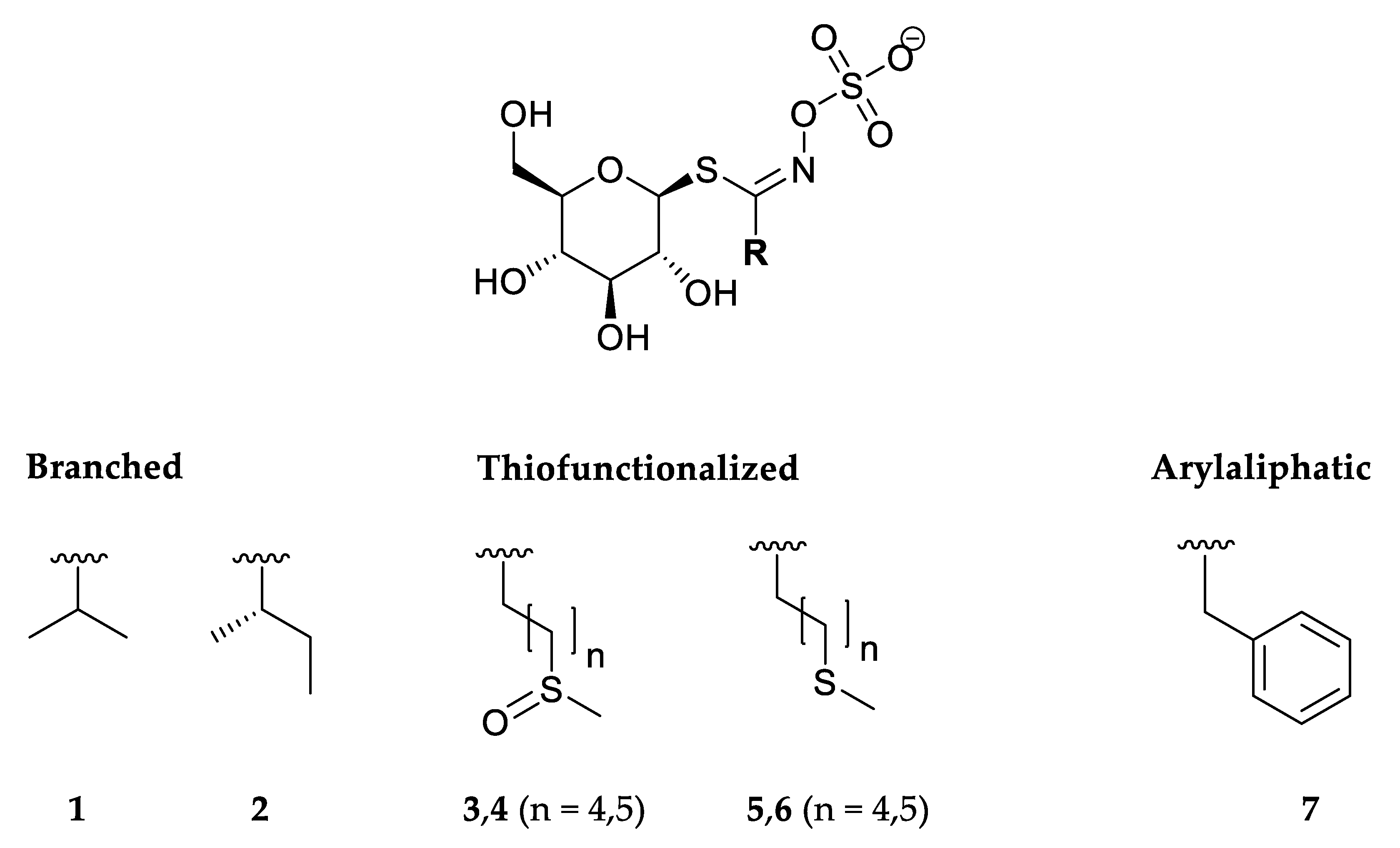
2.2. Isolation and Chemical Analysis
2.2.1. Isolation of Desulfoglucosinolates
2.2.2. HPLC-DAD Analysis of Desulfoglucosinolates
2.2.3. UHPLC-MS/MS Analysis
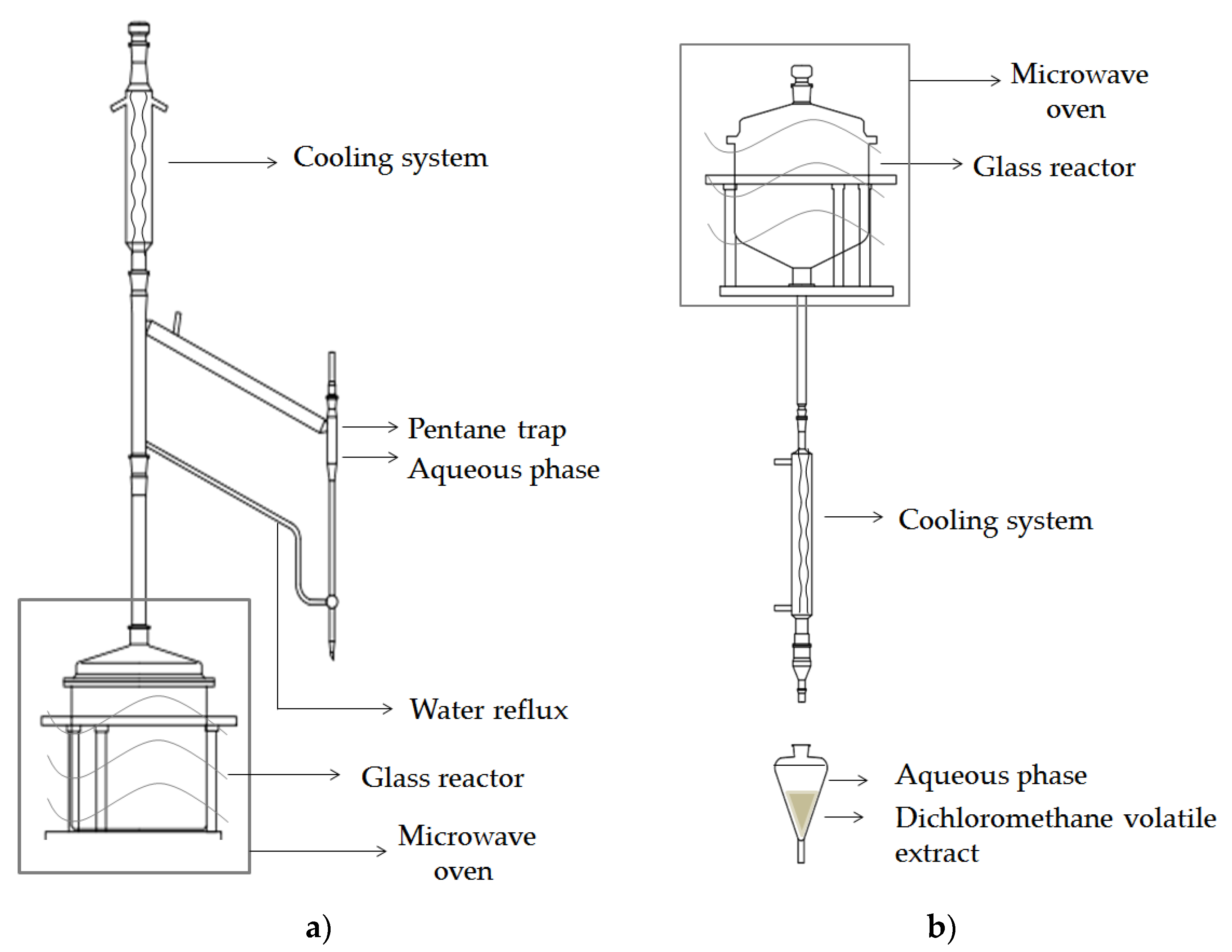
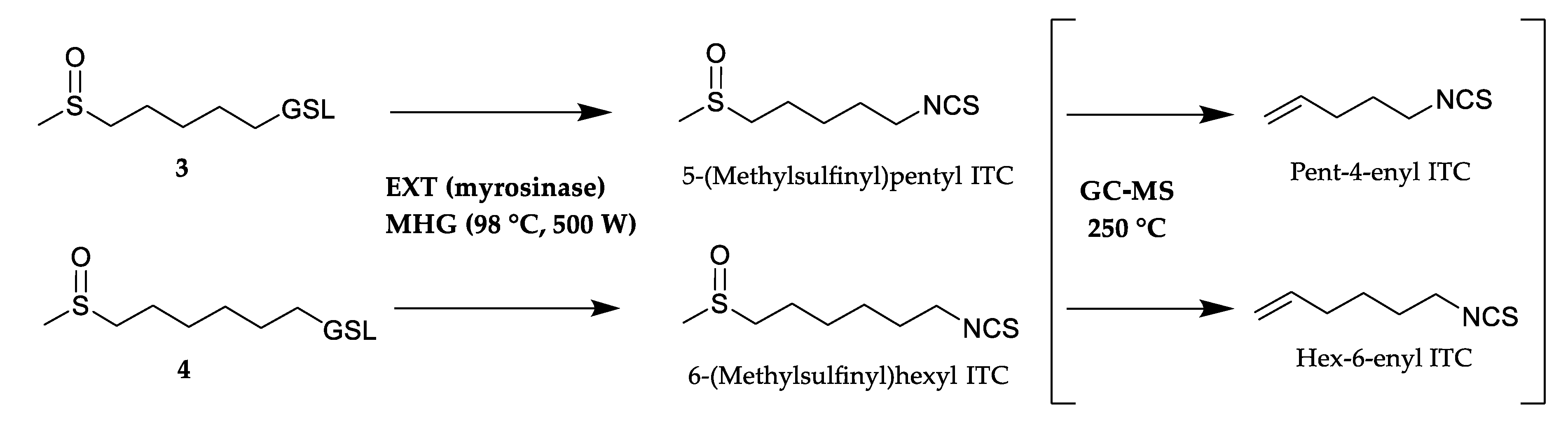
2.2.4. Isolation of Volatiles
Conventional Isolation
Microwave-Assisted Isolation
2.2.5. GC-MS Analysis
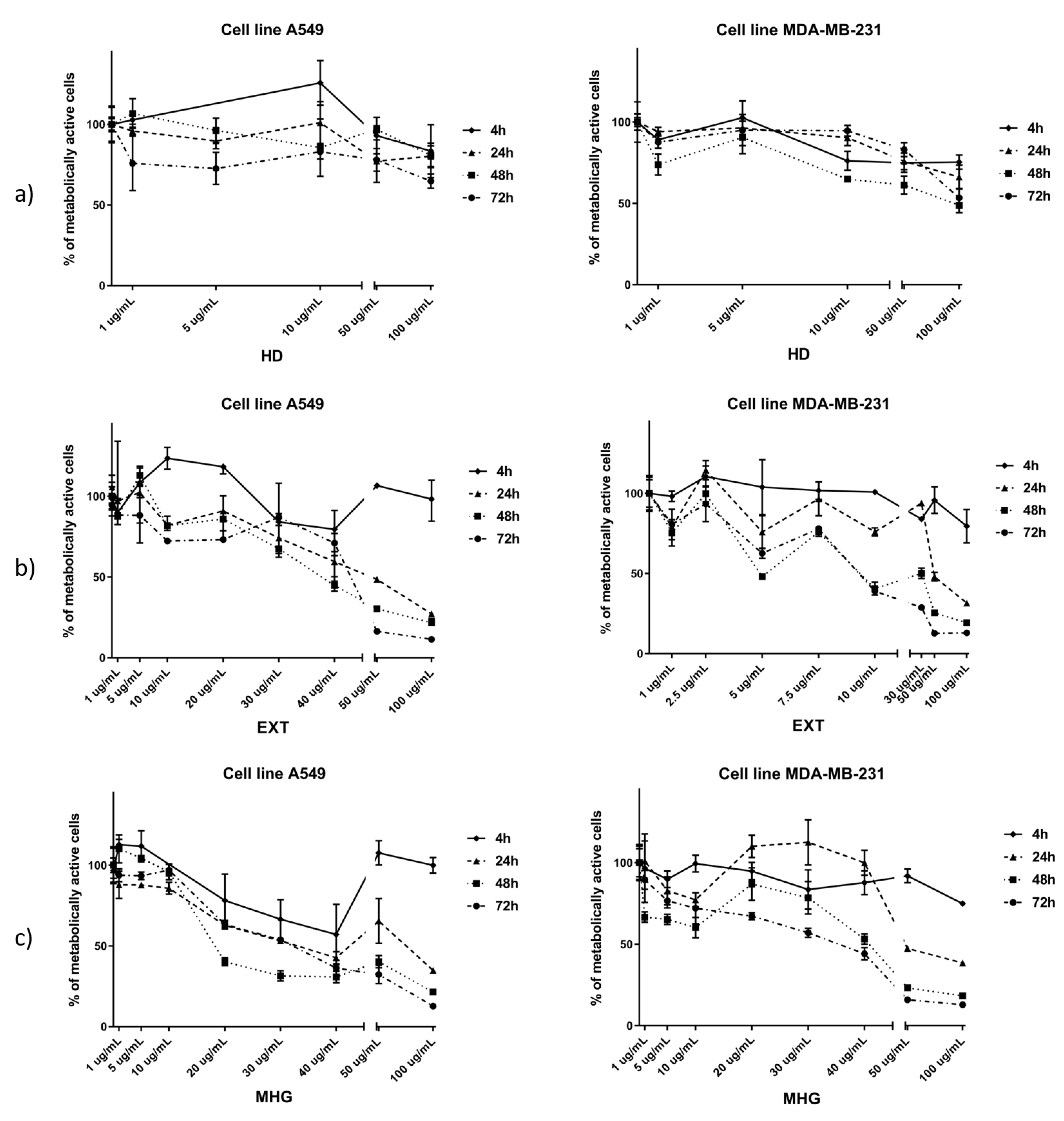
2.3. Cell Viability Assay (MTT)
3. Results and Discussion
3.1. Glucosinolates and Volatile Constituents
3.2. Cytotoxic Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel Sources and Biological Potential. In Glucosinolates; Mérillon, J.-M., Ramawat, G.K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 3–60. [Google Scholar]
- Kjær, A. isoThiocyanates XXXV. Miscellaneous isothiocyanate glucoside acetates. Acta Chem. Scand. 1959, 13, 851–852. [Google Scholar] [CrossRef]
- Danielak, R.; Borkowski, B. Biologically active compounds in seeds of crucifers Part III. Chromatographical search for glucosynolates. Dissert. Pharm. Pharmacol. 1969, 21, 563–575. [Google Scholar]
- Daxenbichler, M.E.; Spencer, G.F.; Carlson, D.G.; Rose, G.B.; Brinker, A.M.; Powell, R.G. Glucosinolate composition of seeds from 297 species of wild plants. Phytochemistry 1991, 30, 2623–2638. [Google Scholar] [CrossRef]
- Blažević, I.; Maleš, T.; Ruščić, M. Glucosinolates of Lunaria annua: Thermal, enzymatic, and chemical degradation. Chem. Nat. Compd. 2014, 49, 1154–1157. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food. Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef]
- Burčul, F.; Generalić Mekinić, I.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzyme Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Maravić, A.; Čikeš Čulić, V.; Montaut, S.; Rollin, P. Antimicrobial and cytotoxic activities of Lepidium latifolium L. Hydrodistillate, extract and its major sulfur volatile allyl isothiocyanate. Chem. Biodivers. 2019, 16, e180066. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Ranjan, A.; Kim, S.-H.; Srivastava, S.K. Chapter five—Mechanisms of the anticancer effects of isothiocyanates. In The enzymes, mechanism of the anticancer effect of phytochemicals; Bathaie, S.Z., Fuyuhiko, T., Eds.; Elsevier Inc., Academic press: London, UK, 2015; Volume 37, pp. 111–137. [Google Scholar]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Burčul, F.; Ljubenkov, I.; Ruščić, M.; Generalić Mekinić, I. Bunias erucago L.: Glucosinolate profile and in vitro biological potential. Molecules 2019, 24, 741. [Google Scholar] [CrossRef]
- Grosser, K.; van Dam, N.M.A. A straightforward method for glucosinolate extraction and analysis with high-pressure liquid chromatography (HPLC). J. Vis. Exp. 2017, 121, e55425. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef]
- Wathelet, J.-P.; Iori, R.; Leoni, O.; Quinsac, A.; Palmieri, S.; Rollin, P. Guidelines for glucosinolate analysis in green tissues used for biofumigation. Agroindustria 2004, 3, 257–266. [Google Scholar]
- Blažević, I.; Radonić, A.; Skočibušić, M.; De Nicola, G.R.; Montaut, S.; Iori, R.; Rollin, P.; Mastelić, J.; Zekić, M.; Maravić, A. Glucosinolate profiling and antimicrobial screening of Aurinia leucadea (Brassicaceae). Chem. Biodivers. 2011, 8, 2310–2321. [Google Scholar] [CrossRef]
- Kjær, A.; Ohashi, M.; Wilson, J.M.; Djerassi, C. Mass spectra of isothiocyanates. Acta Chem. Scand. 1963, 17, 2143–2154. [Google Scholar] [CrossRef]
- Spencer, G.F.; Daxenbichler, M.E. Gas chromatography-mass spectrometry of nitriles, isothiocyanates and oxazolidinethiones derived from cruciferous glucosinolates. J. Sci. Food Agric. 1980, 31, 359–367. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Berhow, M.A. Glucosinolate hydrolysis products from various plant sources: pH effects, isolation, and purification. Ind. Crops Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Al-Gendy, A.A.; Nematallah, K.A.; Zaghloul, S.S.; Ayoub, N.A. Glucosinolates profile, volatile constituents, antimicrobial, and cytotoxic activities of Lobularia libyca. Pharm. Biol. 2016, 54, 3257–3263. [Google Scholar] [CrossRef]
- Olsen, C.E.; Huang, X.-C.; Hansen, C.I.C.; Cipollini, D.; Ørgaard, M.; Matthes, A.; Geu-Flores, F.; Koch, M.A.; Agerbirk, N. Glucosinolate diversity within a phylogenetic framework of the tribe Cardamineae (Brassicaceae) unrevealed with HPLC-MS/MS and NMR-based analytical distinction of 70 desulfoglucosinolates. Phytochemistry 2016, 132, 33–56. [Google Scholar] [CrossRef]
- Chiang, W.C.K.; Pusateri, D.J.; Leitz, R.E.A. Gas chromatography/mass spectrometry for the determination of sulforaphane and sulforaphane nitrile in Broccoli. J. Agric. Food Chem. 1998, 46, 1018–1021. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; De Nicola, G.R.; Rollin, P. Long-chain glucosinolates from Arabis turrita: Enzymatic and non-enzymatic degradations. Nat. Prod. Commun. 2015, 10, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Shinoda, S.; Yamori, T.; Sawaki, S.; Nagata, I.; Ryoyama, K.; Fuke, Y. Selective sensitivity to wasabi-derived 6-(methylsulfinyl)hexyl isothiocyanate of human breast cancer and melanoma cell lines studied in vitro. Cancer Detect. Prev. 2005, 29, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Fuke, Y.; Hishinuma, M.; Namikawa, M.; Oishi, Y.; Matsuzaki, T. Wasabi-derived 6-(methylsulfinyl)hexyl isothiocyanate induces apoptosis in human breast cancer by possible involvement of the NF-κB pathways. Nutr. Cancer 2014, 66, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Morroni, F.; Sita, G.; Tarozzi, A.; Cantelli-Forti, G.; Hrelia, P. Neuroprotection by 6-(methylsulfinyl)hexyl isothiocyanate in a 6-hydroxydopamine mouse model of Parkinson׳s disease. Brain Res. 2014, 1589, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Trio, P.Z.; Fujisaki, S.; Tanigawa, S.; Hisanaga, A.; Sakao, K.; Hou, D.-X. DNA microarray highlights Nrf2-mediated neuron protection targeted by Wasabi-derived isothiocyanates in IMR-32 cells. Gene Regul. Syst. Biol. 2016, 10, 73–83. [Google Scholar] [CrossRef]
- Yano, S.; Wu, S.; Sakao, K.; Hou, D.-X. Wasabi 6-(methylsulfinyl)hexyl isothiocyanate induces apoptosis in human colorectal cancer cells through p53-independent mitochondrial dysfunction pathway. BioFactors 2018, 44, 361–368. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Huang, Y.-C.; Tsai, T.-H.; Liao, H.-F. Effect of wasabi component 6-(methylsulfinyl)hexyl isothiocyanate and derivatives on human pancreatic cancer cells. Evidence-Based Compl. Alt. Med. 2014, 494739. [Google Scholar] [CrossRef]
- Wu, K.-M.; Liao, H.-F.; Chi, C.-W.; Kou, Y.R.; Chen, Y.-J. Wasabi compound 6-(methylsulfinyl) hexyl isothiocyanate induces cell death with coexisting mitotic arrest and autophagy in human chronic myelogenous leukemia K562 cells. Biomolecules 2019, 9, 774. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucosinolate (Trivial Name) | tR (min) | Content (μmol/g DW) | [M + Na]+ | |
|---|---|---|---|---|
| Branched | ||||
| 1 | Isopropyl GSL (Glucoputranjivin) | 3.80 | 14.60 ± 0.59 | 304 |
| 2 | sec-Butyl GSL (Glucocochlearin) | 5.34 | 0.41 ± 0.11 | 318 |
| Sulfur-containing | ||||
| 3 | 5-(Methylsulfinyl)pentyl GSL (Glucoalyssin) | 3.90 | 1.74 ± 0.13 | 394 |
| 4 | 6-(Methylsulfinyl)hexyl GSL (Glucohesperin) | 5.05 | 3.54 ± 0.21 | 408 |
| 5 | 5-(Methylsulfanyl)pentyl GSL (Glucoberteroin) | 7.61 | Tr | 378 |
| 6 | 6-(Methylsulfanyl)hexyl GSL (Glucolesquerellin) | 9.11 | Tr | 392 |
| Arylaliphatic | ||||
| 7 | Benzyl GSL (Glucotropaeolin) | 6.06 | 1.54 ± 0.31 | 352 |
| Total (μmol/g DW) | 21.83 ± 1.35 | |||
| Parent Glucosinolate/ Identified Compound | RI | Conventional Techniques | Microwave-Assisted Isolation | ||
|---|---|---|---|---|---|
| HD | EXT | MAD | MHG | ||
| Glucoputranjivin | |||||
| Isopropyl ITC a,b,c | 858 | 92.13 | 35.56 | 87.03 | 88.02 |
| Glucocochlearin | |||||
| sec-Butyl ITC a,b,c | 952 | 4.59 | 0.39 | 4.11 | 3.08 |
| Glucoalyssin | |||||
| 5-(Methylsulfinyl)pentyl ITC (alyssin) a,c | 2005 | - | 3.30 | - | - |
| Pent-4-enyl ITC a,c, ⸸ | 1093 | - | 24.34 | - | 1.56 |
| Glucohesperin | |||||
| 6-(Methylsulfinyl)hexyl ITC (hesperin) a,c | 2104 | - | 2.31 | - | - |
| Hex-5-enyl ITC a,c, ‡ | 1226 | - | 32.02 | - | 2.34 |
| Glucoberteroin | |||||
| 5-(Methylsulfanyl)pentyl ITC (berteroin) a,c | 1566 | 0.21 | - | - | - |
| Glucolesquerellin | |||||
| 6-(Methylsulfanyl)hexyl ITC (lesquerellin) a,c | 1714 | 0.29 | - | - | - |
| Glucotropaeolin | |||||
| Benzyl ITC a,b,c | 1395 | 0.14 | - | - | - |
| Others | |||||
| S-Methyl- methanethiosulfinate a,b,c | 1005 | - | 0.63 | - | - |
| Diisopropyl disulfide a,b,c | 1010 | - | 0.10 | 0.33 | 0.14 |
| Phenylacetaldehyde a,b,c | 1072 | - | - | - | 0.31 |
| Hexadecanoic acid a,b,c | 1980 | 1.04 | - | 0.38 | 0.82 |
| Total (%) | 98.40 | 98.66 | 92.05 | 96.26 | |
| Yield (µg/g) | 578.90 | 2331.20 | 3.80 | 47.10 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity. Biomolecules 2020, 10, 215. https://doi.org/10.3390/biom10020215
Blažević I, Đulović A, Čikeš Čulić V, Popović M, Guillot X, Burčul F, Rollin P. Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity. Biomolecules. 2020; 10(2):215. https://doi.org/10.3390/biom10020215
Chicago/Turabian StyleBlažević, Ivica, Azra Đulović, Vedrana Čikeš Čulić, Marijana Popović, Xavier Guillot, Franko Burčul, and Patrick Rollin. 2020. "Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity" Biomolecules 10, no. 2: 215. https://doi.org/10.3390/biom10020215
APA StyleBlažević, I., Đulović, A., Čikeš Čulić, V., Popović, M., Guillot, X., Burčul, F., & Rollin, P. (2020). Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity. Biomolecules, 10(2), 215. https://doi.org/10.3390/biom10020215