High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’


Abstract

1. Introduction
1.1. The Glutaminase II Pathway in Mammalian Tissues
1.2. The Glutaminase II Pathway in Cancer Cells
2. Materials and Methods
2.1. Chemicals
2.2. Prostate Cancer Tissue Selection
2.3. Prostate Cancer Cell Lines
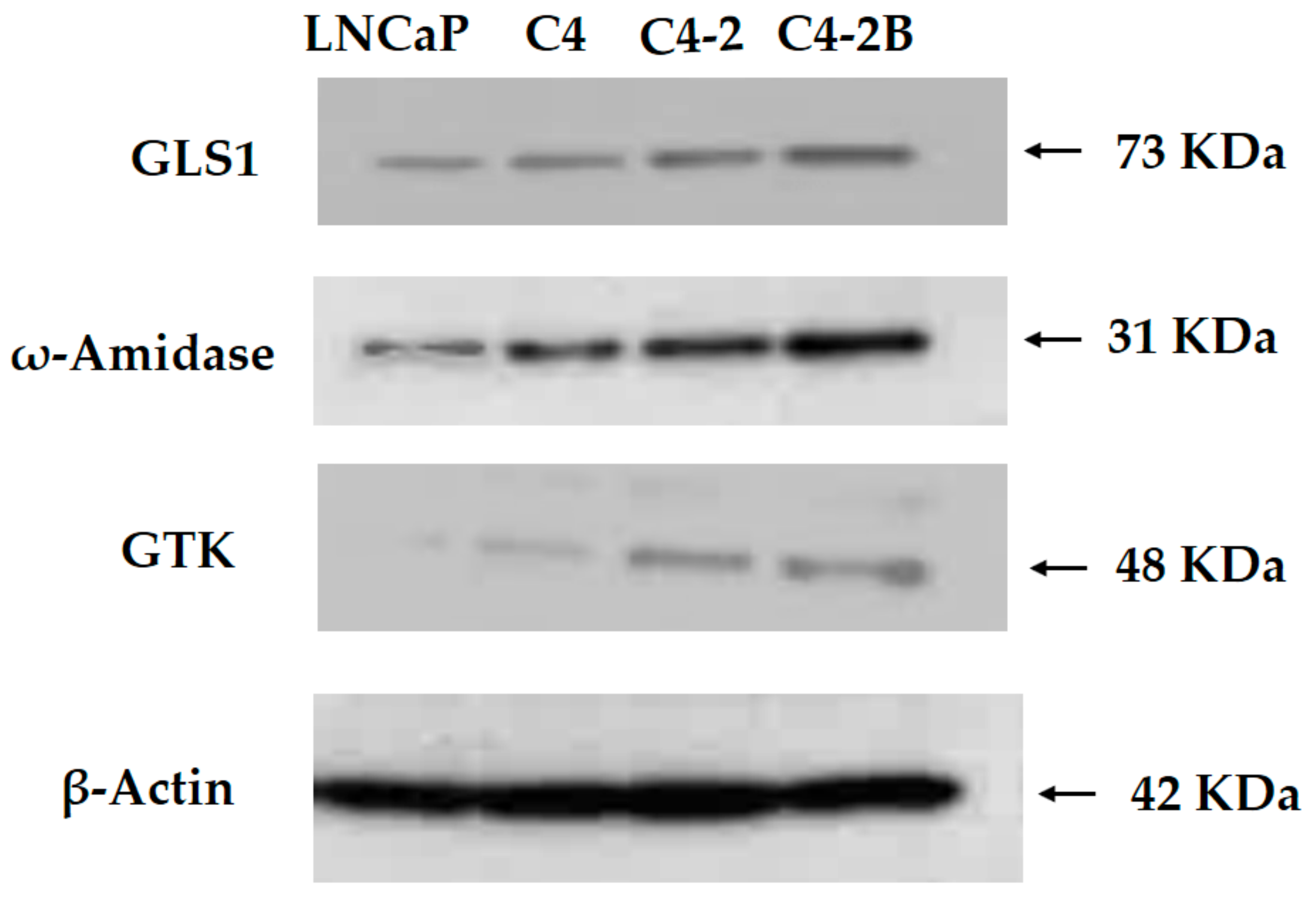
2.4. Western Blots
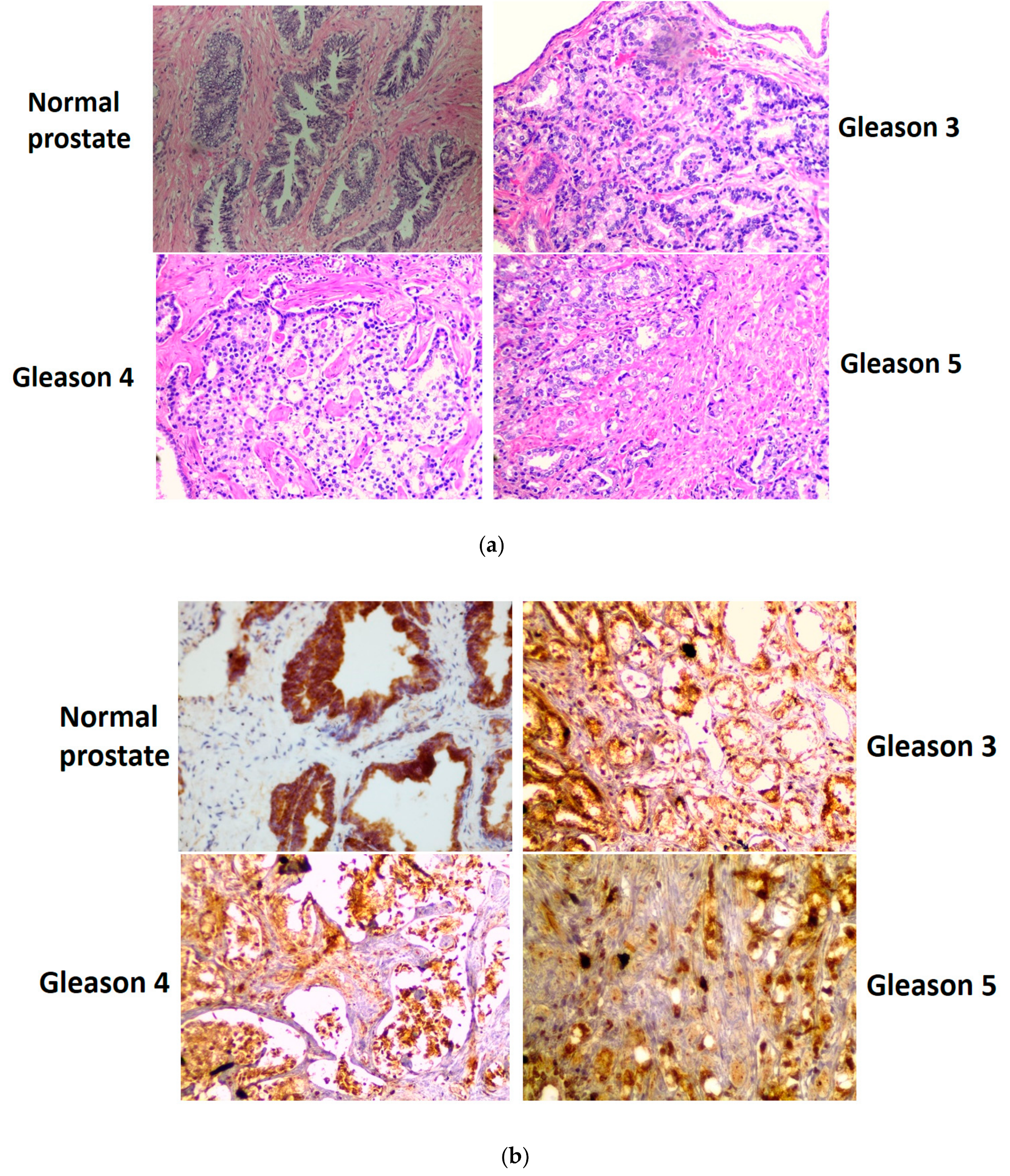
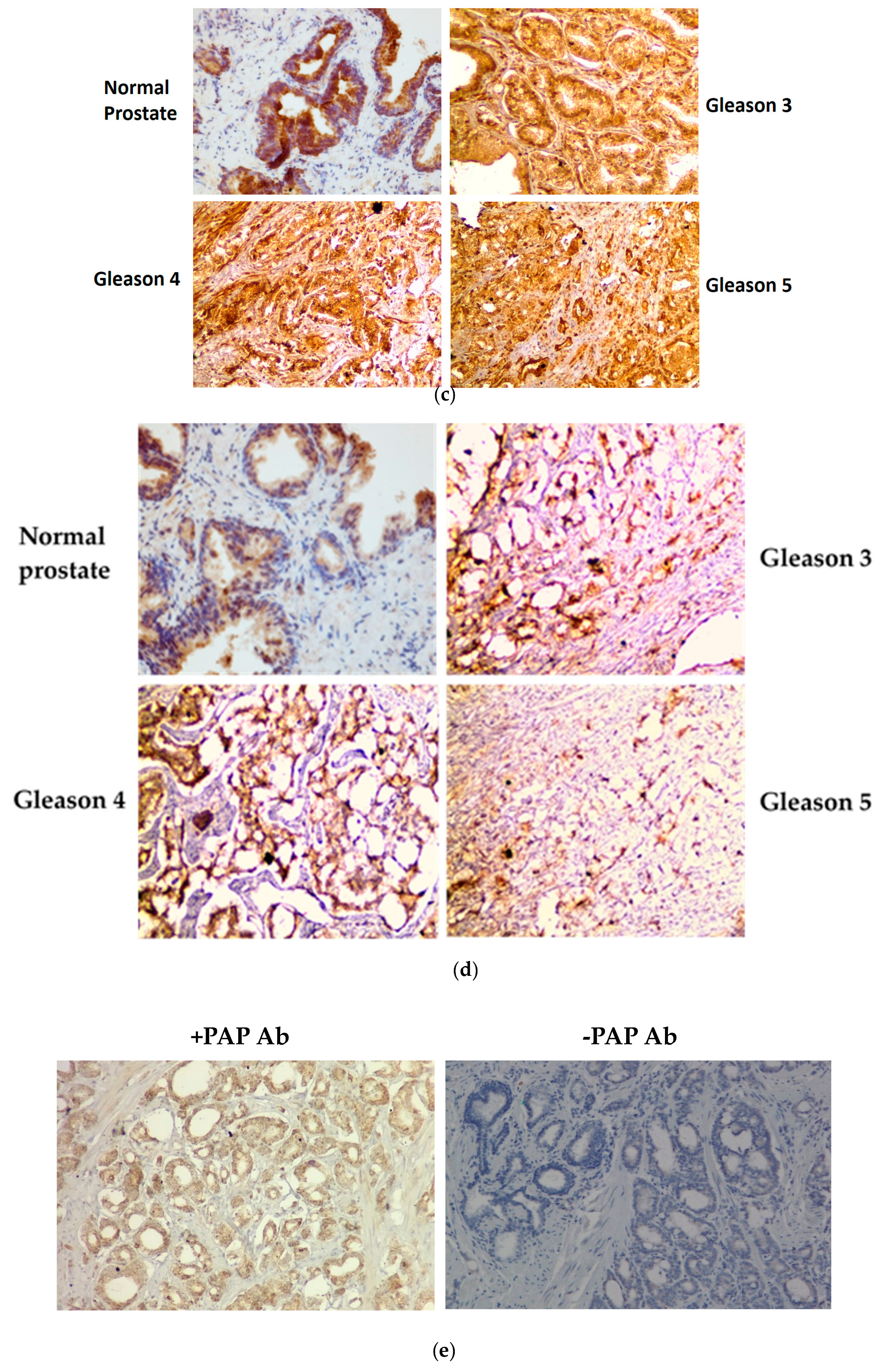
2.5. Immunohistochemistry
2.6. Preparation of Rat Tissues for Enzyme Determinations
2.7. Measurement of Transaminase Activities with l-Glutamine, MSC, and SM in Rat Tissue Homogenates
2.8. Measurement of ω-Amidase Activity in Rat Tissue Homogenates
2.9. Statistics
3. Results
3.1. Transamination of l-Glutamine, SM, and MSC Catalyzed by Preparations of Rat Liver, Kidney, and Prostate
3.2. ω-Amidase Activities in Homogenates of Rat Liver, Kidney and Prostate
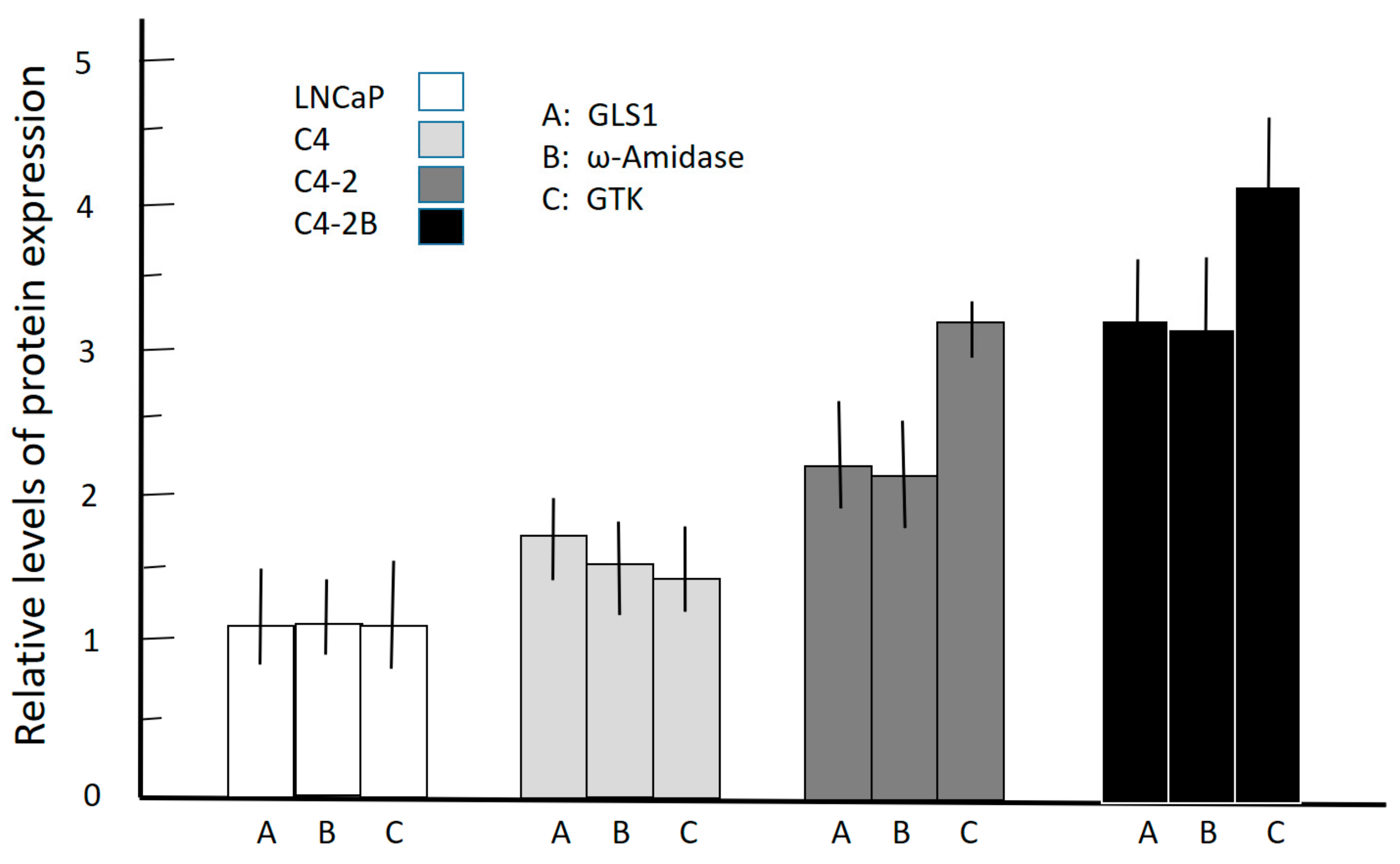
3.3. Western Blotting of Cell Lysates of Various Prostate Cancer Cells
3.4. Immunohistochemical Staining of GLS1, ω-Amidase, and GTK in Normal Human Prostate and in Cancerous Human Prostate of Increasing Aggressiveness
4. Discussion
4.1. The Glutaminase II Pathway Provides Anaplerotic a-Ketoglutarate in Normal and Cancerous Prostate while Closing the Methionine Salvage Pathway
4.2. Role of the Glutaminase II Pathway in Replenishing Citrate Carbon in Normal and Cancerous Prostate
4.3. The Glutaminase II Pathway Permits the Formation of a-Ketoglutarate from l-Glutamine under Hypoxic/Anoxic Conditions
4.4. Biological Importance of GLS1 and Glutaminase II Pathway Enzymes in the Stromal Cell Compartment of Human Prostate Cancer Cells
4.5. On the Relative Affinities Exhibited by GLS1 and GTK toward l-Glutamine and Relevance to the In Vivo Metabolism of l-Glutamine
4.6. Glutaminase II Pathway Inhibitors May Offer Potentially Significant Therapeutic Opportunities
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cooper, A.J.L.; Shurubor, Y.I.; Dorai, T.; Pinto, J.T.; Isakova, E.P.; Deryabina, Y.I.; Denton, T.T.; Krasnikov, B.F. ω-Amidase: An underappreciated, but important enzyme in L-glutamine and L-asparagine metabolism; relevance to sulfur and nitrogen metabolism, tumor biology and hyperammonemic diseases. Amino Acids 2016, 48, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.L.; Meister, A. Comparative studies of glutamine transaminases from rat tissues. Comp. Biochem. Physiol. 1981, 69, 137–145. [Google Scholar] [CrossRef]
- Cooper, A.J.L. Glutamine aminotransferases and ω-amidases. In Glutamine and Glutamate in Mammals; Kvamme, E., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1988; Volume 1, pp. 33–52. [Google Scholar]
- Pinto, J.T.; Krasnikov, B.F.; Alcutt, S.; Jones, M.E.; Dorai, T.; Villar, M.T.; Artigues, A.; Li, J.; Cooper, A.J.L. Kynurenine aminotransferase III and glutamine transaminase L are identical enzymes that have cysteine S-conjugate β-lyase activity and can transaminate L-selenomethionine. J. Biol. Chem. 2014, 289, 30950–30961. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. Preparation of enzymatic reactions of the keto analogues of asparagine and glutamine. J. Biol. Chem. 1953, 200, 571–589. [Google Scholar]
- Mardashev, S.R.; Lerman, M.I.; Benyumovich, M.S. Glutamine transaminase in brain tissue preparations and in cells of a human strain of dedifferentiated astrocytoma. Fed. Proc. Transl. Suppl. 1963, 22, 976–977. [Google Scholar]
- Cooper, A.J.L.; Dorai, T.; Dorai, B.; Krasnikov, B.F.; Li, J.; Hallen, A.; Pinto, J.T. Role of glutamine transaminases in nitrogen, sulfur, selenium and 1-carbon metabolism: Glutamine transaminases in normal and cancer cells. In Glutamine in Clinical Nutrition (Nutrition and Health Series); Rajendram, R., Preedy, V.R., Patel, V.B., Bendich, A., Eds.; Humana Press: New York, NY, USA, 2015; pp. 37–54. [Google Scholar]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, 6340. [Google Scholar] [CrossRef]
- Udupa, S.; Nguyen, S.; Hoang, G.; Nguyen, T.; Quinones, A.; Pham, K.; Asaka, R.; Nguyen, K.; Zhang, C.; Elgogary, A.; et al. Upregulation of the glutaminase ii pathway contributes to glutamate production upon glutaminase 1 inhibition in pancreatic cancer. Proteomics 2019. [Google Scholar] [CrossRef]
- Nian, H.; Bisson, W.H.; Dashwood, W.M.; Pinto, J.T.; Dashwood, R.H. α-Keto acid metabolites of organoselenium compounds inhibit histone deacetylase activity in human colon cancer cells. Carcinogenesis 2009, 30, 1416–1423. [Google Scholar] [CrossRef]
- Krasnikov, B.F.; Nostramo, R.; Pinto, J.T.; Cooper, A.J.L. Assay and purification of ω-amidase/Nit2, a ubiquitously expressed putative tumor suppressor that catalyzes the deamidation of the alpha-keto acid analogues of glutamine and asparagine. Anal. Biochem. 2009, 391, 144–150. [Google Scholar] [CrossRef]
- Dorai, T.; Dutcher, J.P.; Dempster, D.W.; Wiernik, P.H. Therapeutic potential of curcumin in prostate cancer--V: Interference with the osteomimetic properties of hormone refractory C4-2B prostate cancer cells. Prostate 2004, 60, 1–17. [Google Scholar] [CrossRef]
- Koeneman, K.S.; Yeung, F.; Chung, L.W.K. Osteomimetic properties of prostate cancer cells: A hypothesis supporting the predilection of prostate cancer metastasis and growth in the bone environment. Prostate 1999, 39, 246–261. [Google Scholar] [CrossRef]
- Thalmann, G.N.; Sikes, R.A.; Wu, T.T.; Degeorges, A.; Chang, S.M.; Ozen, M.; Pathak, S.; Chung, L.W. LNCaP progression model of human prostate cancer: Androgen-independence and osseous metastasis. Prostate 2000, 44, 91–103. [Google Scholar] [CrossRef]
- Cooper, A.J.L.; Meister, A. Isolation and properties of a new glutamine transaminase from rat kidney. J. Biol. Chem. 1974, 249, 2554–2561. [Google Scholar] [PubMed]
- Pinto, J.T.; Van Raamsdonk, J.M.; Leavitt, B.R.; Hayden, M.R.; Jeitner, T.M.; Thaler, H.T.; Krasnikov, B.F.; Cooper, A.J.L. Treatment of YAC128 mice and their wild-type littermates with cystamine does not lead to its accumulation in plasma or brain: Implications for the treatment of Huntington disease. J. Neurochem. 2005, 94, 1087–1101. [Google Scholar] [CrossRef]
- Pinto, J.T.; Khomenko, T.; Szabo, S.; McLaren, G.D.; Denton, T.T.; Krasnikov, B.F.; Jeitner, T.M.; Cooper, A.J.L. Measurement of sulfur-containing compounds involved in the metabolism and transport of cysteamine and cystamine. Regional differences in cerebral metabolism. J. Chromatogr. B 2009, 877, 3434–3441. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cui, H. Targeting glutamine induces apoptosis: A cancer therapy approach. Int. J. Mol. Sci. 2015, 16, 22830–22855. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Le, A. Glutamine metabolism in cancer. Adv. Exp. Med. Biol. 2018, 1063, 13–32. [Google Scholar]
- Bernfeld, E.; Foster, D.A. Glutamine as an essential amino acid for KRas-driven cancer cells. Trends Endocrinol. Metab. 2019, 30, 357–368. [Google Scholar] [CrossRef]
- Darmaun, D.; Matthews, D.E.; Bier, D.M. Glutamine and glutamine kinetics in humans. Am. J. Physiol. 1986, 251, E117–E126. [Google Scholar]
- Williams-Ashman, H.G.; Pegg, A.E.; Lockwood, D.H. Mechanisms and regulation of polyamine and putrescine biosynthesis in male genital glands and other tissues of mammals. Adv. Enzyme Regul. 1969, 7, 291–323. [Google Scholar] [CrossRef]
- Pegg, A.E.; Lockwood, D.H.; Williams-Ashman, H.G. Concentrations of putrescine and polyamines and their enzymic synthesis during androgen-induced prostatic growth. Biochem. J. 1970, 117, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Williams-Ashman, H.G.; Canellakis, Z.N. Polyamines in mammalian biology and medicine. Perspect. Biol. Med. 1979, 22, 421–453. [Google Scholar] [CrossRef] [PubMed]
- Wray, J.W.; Abeles, R.H. The methionine salvage pathway in Klebsiella pneumoniae and rat liver. Identification and characterization of two novel dioxygenases. J. Biol. Chem. 1995, 270, 3147–3153. [Google Scholar] [CrossRef] [PubMed]
- Albers, E. Metabolic characteristics and importance of the universal methionine salvage pathway recycling methionine from 5’-methylthioadenosine. IUBMB Life 2009, 61, 1132–1142. [Google Scholar] [CrossRef]
- Backlund, P.S., Jr.; Chang, C.P.; Smith, R.A. Identification of 2-keto-4-methylthiobutyrate as an intermediate compound in methionine synthesis from 5’-methylthioadenosine. J. Biol. Chem. 1982, 257, 4196–4202. [Google Scholar]
- Cooper, A.J.L.; Meister, A. Isolation and properties of a highly purified glutamine transaminase. Biochemistry 1972, 11, 661–671. [Google Scholar]
- Han, Q.; Li, J.; Li, J. pH dependence, substrate specificity and inhibition of human kynurenine aminotransferase I. Eur. J. Biochem. 2004, 271, 4804–4814. [Google Scholar] [CrossRef]
- Han, Q.; Robinson, H.; Cai, T.; Tagle, D.A.; Li, J. Biochemical and structural properties of mouse kynurenine aminotransferase III. Mol. Cell. Biol 2009, 29, 784–793. [Google Scholar] [CrossRef]
- Bistulfi, G.; Affronti, H.C.; Foster, B.A.; Karasik, E.; Gillard, B.; Morrison, C.; Mohler, J.; Phillips, J.G.; Smiraglia, D.J. The essential role of methylthioadenosine phosphorylase in prostate cancer. Oncotarget 2016, 7, 14380–14393. [Google Scholar] [CrossRef]
- Ellens, K.W.; Richardson, L.G.; Frelin, O.; Collins, J.; Ribeiro, C.L.; Hsieh, Y.F.; Mullen, R.T.; Hanson, A.D. Evidence that glutamine transaminase and ω-amidase potentially act in tandem to close the methionine salvage cycle in bacteria and plants. Phytochemistry 2015, 113, 160–169. [Google Scholar] [CrossRef]
- Kline, E.E.; Treat, E.G.; Averna, T.A.; Davis, M.S.; Smith, A.Y.; Sillerud, L.O. Citrate concentrations in human seminal fluid and expressed prostatic fluid determined via 1H nuclear magnetic resonance spectroscopy outperform prostate specific antigen in prostate cancer detection. J. Urol. 2006, 176, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Costello, L.C.; Franklin, R.B.A. Comprehensive review of the role of zinc in normal prostate function and metabolism; and its implications in prostate cancer. Arch. Biochem. Biophys. 2016, 611, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Eidelman, E.; Twum-Ampofo, J.; Ansari, J.; Siddiqui, M.M. The metabolic phenotype of prostate cancer. Front. Oncol. 2017, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Costello, L.C.; Franklin, R.B.; Feng, P. Mitochondrial function, zinc, and intermediary metabolism relationships in normal prostate and prostate cancer. Mitochondrion 2005, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Gao, L.; Wu, G.; Shen, G.; Xie, S.; Wen, H.; Yang, J.; Zhou, Y.; Tu, Z.; Qian, W. Elevated expression of glutaminase confers glucose utilization via glutaminolysis in prostate cancer. Biochem. Biophys. Res. Commun. 2015, 456, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.L.; Abraham, D.G.; Gelbard, A.S.; Lai, J.C.K.; Petito, C.K. High activities of glutamine transaminase K (dichlorovinylcysteine β-lyase) and ω-amidase in the choroid plexus of rat brain. J. Neurochem. 1993, 61, 1731–1741. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.W.; Qin, C.; Schaeffer, V.H.; Stevens, J.L. Immunohistochemical localization of glutamine transaminase K, a rat kidney cysteine conjugate β-lyase, and the relationship to the segment specificity of cysteine conjugate nephrotoxicity. Mol. Pharmacol. 1988, 34, 621–627. [Google Scholar]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef]
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The human SLC1A5 (ASCT2) amino acid transporter: From function to structure and role in cell biology. Front. Cell Dev. Biol. 2018, 6, 96. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, T.; Li, Z.; Wang, L.; Yuan, S.; Sun, L. The role of ASCT2 in cancer: A review. Eur. J. Pharmacol. 2018, 837, 81–87. [Google Scholar] [CrossRef]
- Windmueller, H.G.; Spaeth, A.E. Uptake and metabolism of plasma glutamine by the small intestine. J. Biol. Chem. 1974, 249, 5070–5079. [Google Scholar] [CrossRef] [PubMed]
- Brand, K.; Fekl, W.; von Hintzenstern, J.; Langer, K.; Luppa, P.; Schoerner, C. Metabolism of glutamine in lymphocytes. Metabolism 1989, 38, 29–33. [Google Scholar] [CrossRef]
- Biltz, R.M.; Letteri, J.M.; Pellegrino, E.D.; Palekar, A.; Pinkus, L.M. Glutamine metabolism in bone. Miner. Electrolyte Metab. 1983, 9, 125–131. [Google Scholar]
- Pailla, K.; Blonde-Cynober, F.; Aussel, C.; De Bandt, J.P.; Cynober, L. Branched-chain keto-acids and pyruvate in blood: Measurement by HPLC with fluorimetric detection and changes in older subjects. Clin. Chem. 2000, 46, 848–853. [Google Scholar] [PubMed]
- Steele, R.D. Blood-brain barrier transport of the α-keto acid analogs of amino acids. Fed. Proc. 1986, 45, 2060–2064. [Google Scholar] [PubMed]
- Häussinger, D.; Stehle, T.; Gerok, W. Glutamine metabolism in isolated perfused rat liver. The transamination pathway. Biol. Chem. Hoppe Seyler 1985, 366, 527–536. [Google Scholar]
- Suh, E.H.; Hackett, E.P.; Wynn, R.M.; Chuang, D.T.; Zhang, B.; Luo, W.; Sherry, A.D.; Park, J.M. In vivo assessment of increased oxidation of branched-chain amino acids in glioblastoma. Sci. Rep. 2019, 9, 340. [Google Scholar] [CrossRef]
- Dorai, T.; Shah, A.; Summers, F.; Mathew, R.; Huang, J.; Hsieh, T.-C.; Wu, J.M. NRH:quinone oxidoreductase 2 (NQO2) and glutaminase (GLS) both play a role in large extracellular vesicles (LEV) formation in preclinical LNCaP-C4-2B prostate cancer model of progressive metastasis. Prostate 2018, 78, 1181–1195. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; You, S.; Spinelli, C.; Morley, S.; Zandian, M.; Aspuria, P.J.; Cavallini, L.; Ciardiello, C.; Reis Sobreiro, M.; Morello, M.; et al. Large oncosomes contain distinct protein cargo and represent a separate functional class of tumor derived extracellular vesicles. Oncotarget 2015, 6, 11327–11341. [Google Scholar] [CrossRef]
- Wendler, F.; Stamp, G.W.; Chiamas, G. Tumor-stromal cell communication: Small vesicles signal big changes. Trends Cancer 2016, 2, 326–329. [Google Scholar] [CrossRef]
- Vagner, T.; Spinelli, C.; Minciacchi, V.R.; Balaj, L.; Zandian, M.; Conley, A.; Zijlstra, A.; Freeman, M.R.; Demichelis, F.; De, S.; et al. Large extracellular vesicles carry most of the tumour DNA circulating in prostate cancer patient plasma. J. Extracell. Vesicles 2018, 7, 1505403. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, L.; Morandi, A.; Taddei, M.L.; Parri, M.; Comito, G.; Iscaro, A.; Raspollini, M.R.; Magherini, F.; Rapizzi, E.; Masquelier, J.; et al. Cancer-associated fibroblasts promote prostate cancer malignancy via metabolic rewiring and mitochondrial transfer. Oncogene 2019, 38, 5339–5355. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhuo, L.; Wang, X. Metabolic reprogramming of carcinoma-associated fibroblasts and its impact on metabolic heterogeneity of tumors. Semin. Cell Dev. Biol. 2017, 64, 125–131. [Google Scholar] [CrossRef] [PubMed]
- von Ahrens, D.; Bhagat, T.D.; Nagrath, D.; Maitra, A.; Verma, A. The role of stromal cancer-associated fibroblasts in pancreatic cancer. J. Hematol. Oncol. 2017, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Outschoorn, U.E.; Lisanti, M.P.; Sotgia, F. Catabolic cancer-associated fibroblasts transfer energy and biomass to anabolic cancer cells, fueling tumor growth. Semin. Cancer Biol. 2014, 25, 47–60. [Google Scholar] [CrossRef]
- Tajan, M.; Vousden, K.H. The quid pro quo of the tumor/stromal interaction. Cell Metab. 2016, 24, 645–646. [Google Scholar] [CrossRef][Green Version]
- Avagliano, A.; Granato, G.; Ruocco, M.R.; Romano, V.; Belviso, I.; Carfora, A.; Montagnani, S.; Arcucci, A. Metabolic reprogramming of cancer associated fibroblasts: The slavery of stromal fibroblasts. Biomed. Res. Int. 2018. [Google Scholar] [CrossRef]
- Newsholme, E.A.; Parry-Billings, M. Properties of glutamine release from muscle and its importance for the immune system. JPEN J. Parenter. Nutr. 1990, 14, 63S–67S. [Google Scholar] [CrossRef]
- Botman, D.; Tigchelaar, W.; Van Noorden, C.J. Determination of phosphate-activated glutaminase activity and its kinetics in mouse tissues using metabolic mapping (quantitative enzyme histochemistry). J. Histochem. Cytochem. 2014, 62, 813–826. [Google Scholar] [CrossRef]
- Song, M.; Kim, S.H.; Im, C.Y.; Hwang, H.J. Recent Development of Small Molecule Glutaminase Inhibitors. Curr. Top. Med. Chem. 2018, 18, 432–443. [Google Scholar] [CrossRef]
- Ogier, G.; Chantepie, J.; Deshayes, C.; Chantegrel, B.; Charlot, C.; Doutheau, A.; Quash, G. Contribution of 4-methylthio-2-oxobutanoate and its transaminase to the growth of methionine dependent cells in culture: Effect of transaminase inhibitors. Biochem. Pharmacol. 1993, 45, 1631–1644. [Google Scholar] [CrossRef]
- Quash, G.; Roch, A.M.; Charlot, C.; Chantepie, J.; Thomas, V.; Hamedi-Sangsari, F.; Vila, J. 4-methyl 2-oxobutanoate transaminase: A specific target for antiproliferative agents. Bull. Cancer 2004, 91, E61–E79. [Google Scholar] [PubMed]
- Nematollahi, A.; Sun, G.; Jayawickrama, G.S.; Church, W.B. Kynurenine aminotransferase isozyme inhibitors. A Review. Int. J. Mol. Sci. 2016, 17, 946. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gln-KMB (T) | MSC-KMB (T) | SM-KMB (T) | ω-Amidase | |
|---|---|---|---|---|
| Liver | 0.28 ± 0.08 | 0.23 ± 0.02 | 0.48 ± 0.08 | 97 ± 11 |
| Kidney | 0.30 ± 0.02 | 0.31 ± 0.07 | 0.43 ± 0.02 | 102 ± 20 |
| Prostate | 0.81 ± 0.08 | 0.81 ± 0.07 | 0.90 ± 0.05 | 66 ± 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorai, T.; Dorai, B.; Pinto, J.T.; Grasso, M.; Cooper, A.J.L. High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’. Biomolecules 2020, 10, 2. https://doi.org/10.3390/biom10010002
Dorai T, Dorai B, Pinto JT, Grasso M, Cooper AJL. High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’. Biomolecules. 2020; 10(1):2. https://doi.org/10.3390/biom10010002
Chicago/Turabian StyleDorai, Thambi, Bhuvaneswari Dorai, John T. Pinto, Michael Grasso, and Arthur J. L. Cooper. 2020. "High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’" Biomolecules 10, no. 1: 2. https://doi.org/10.3390/biom10010002
APA StyleDorai, T., Dorai, B., Pinto, J. T., Grasso, M., & Cooper, A. J. L. (2020). High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’. Biomolecules, 10(1), 2. https://doi.org/10.3390/biom10010002