
ExoPhot: The Photon Absorption Rate as a New Metric for Quantifying the Exoplanetary Photosynthetic Activity Fitness

 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Photosynthetic Feasibility Metrics
2.2. Stellar Spectral Types
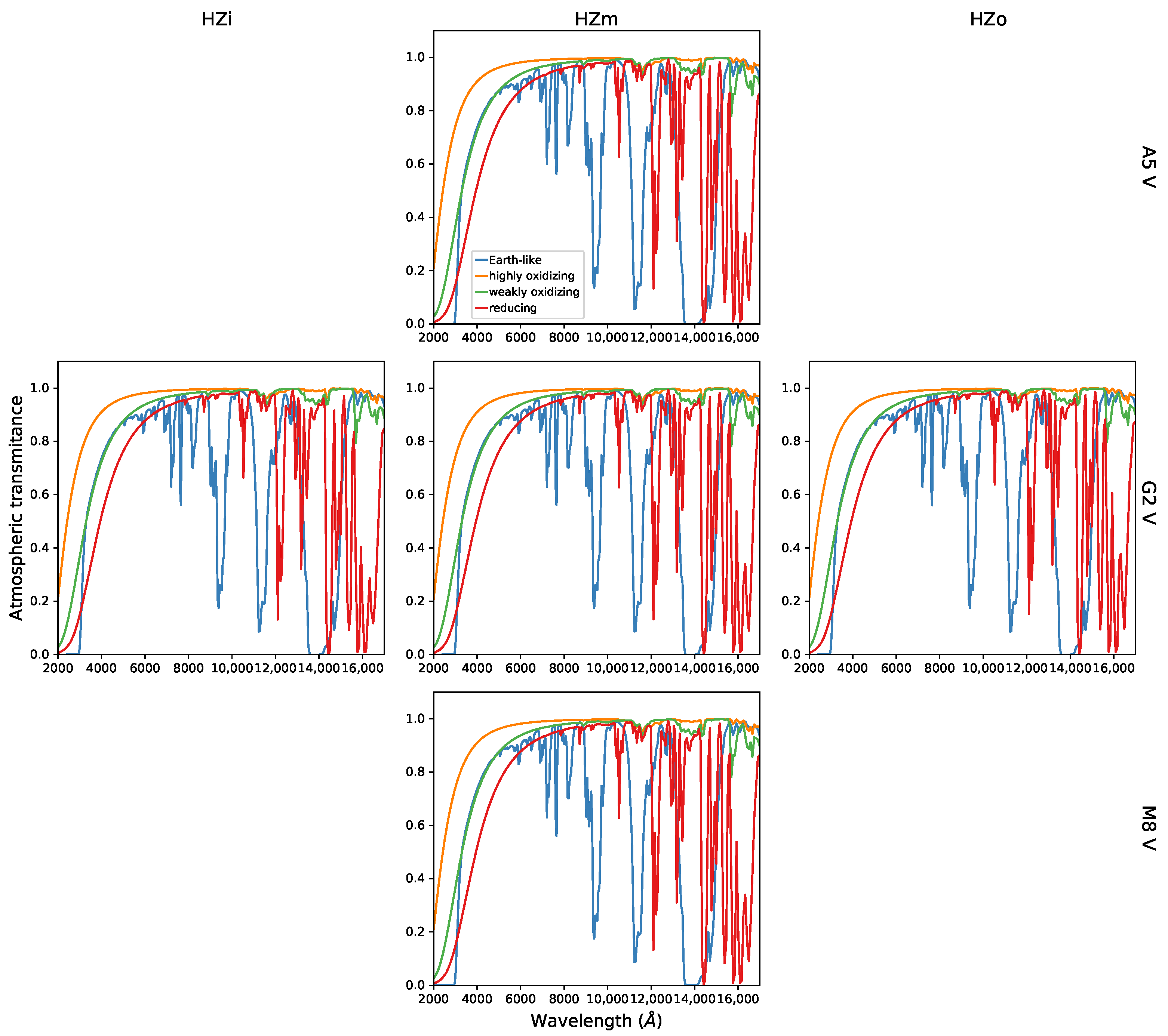
2.3. Modelling of Exoplanets Atmospheres
2.4. Calculation of Absorption Cross-Section of Pigments
2.5. Code and Data Availability
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Chl a | chlorophyll a |
| Chl b | chlorophyll b |
| BChl a | bacteriochlorophyll a |
| BChl b | bacteriochlorophyll b |
| Phot0 | synthetic photosystem 0 |
| photosynthetic photon flux density | |
| HZ | habitability zone |
| HZi | inner part of the HZ |
| HZm | middle part of the HZ |
| HZo | outer part of the HZ |
| PAR | photosynthetic active radiation |
| S | stellar irradiance |
| au | astronomical unit |
Appendix A. Figures and Tables




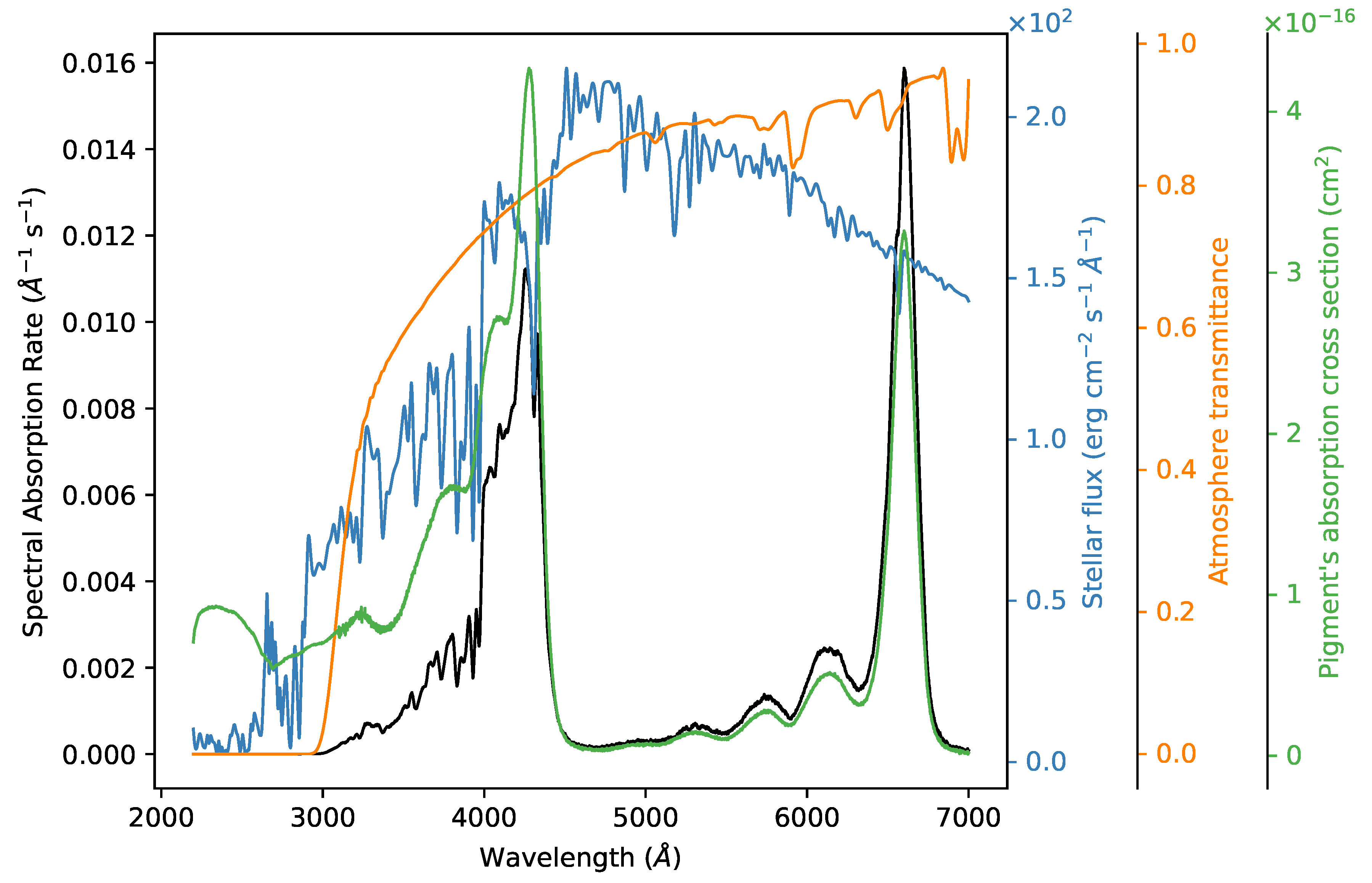{kind=link}
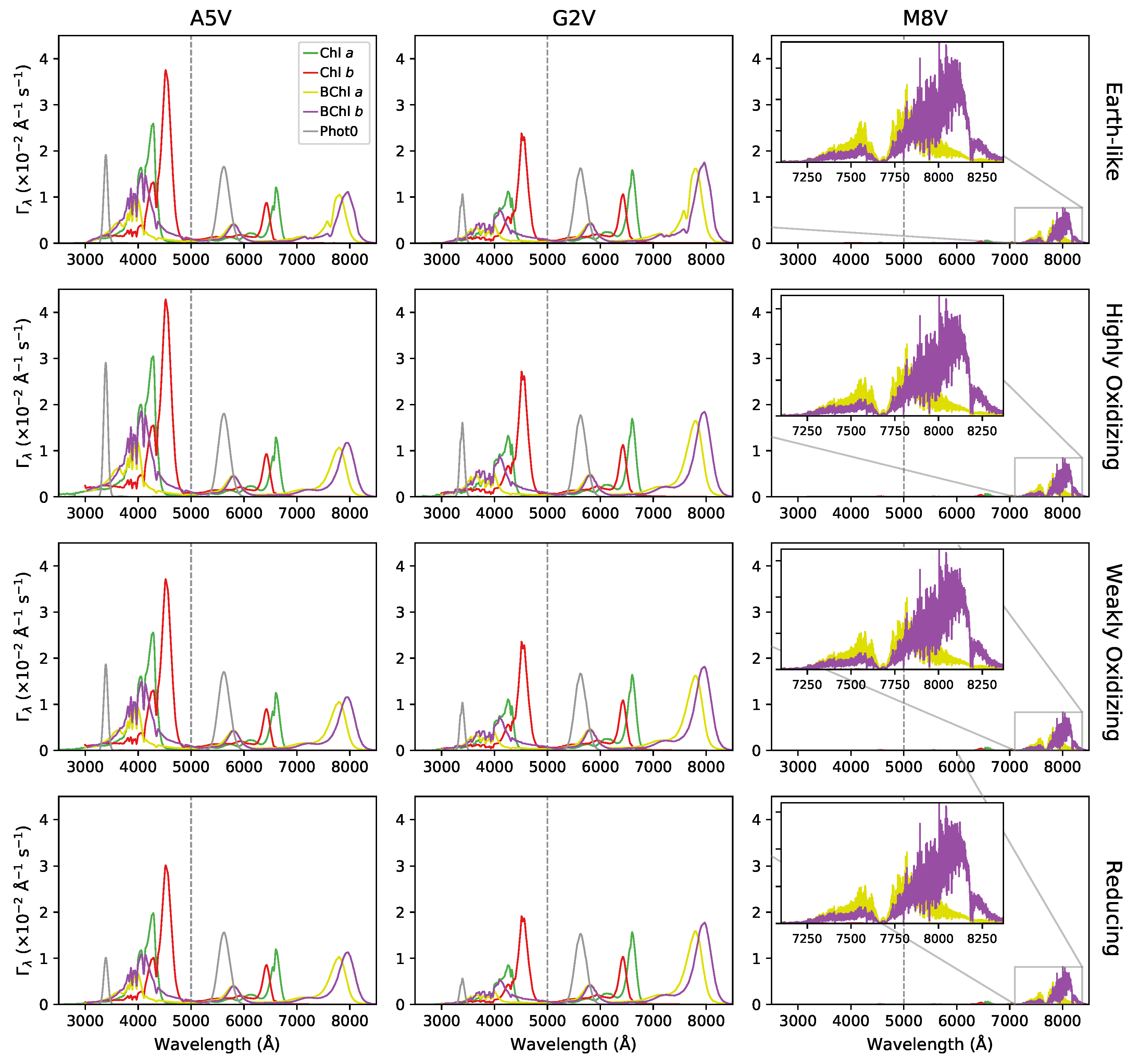{kind=link}
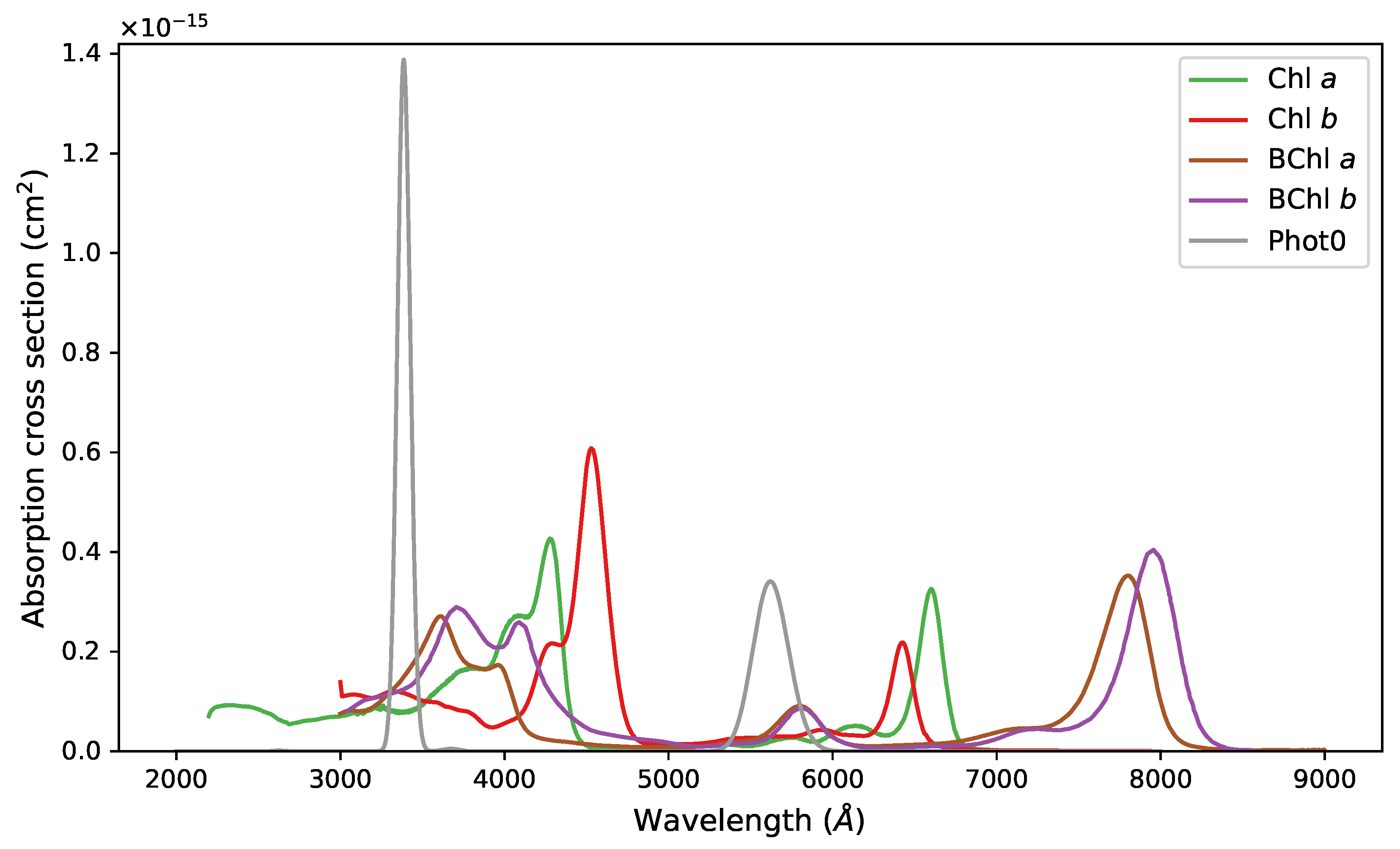{kind=link}
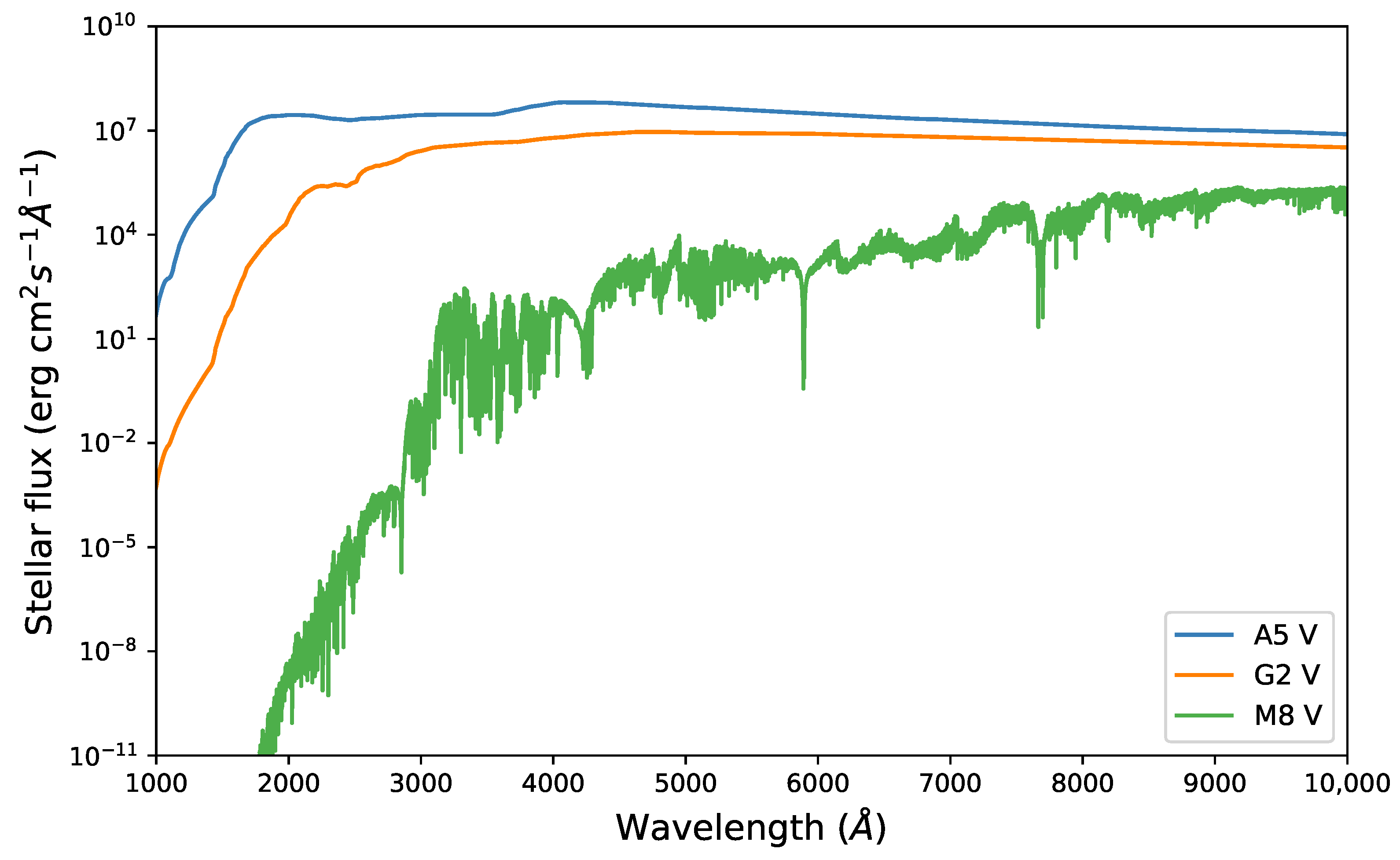{kind=link}
{kind=link}
| Spectral Type | [K] | R [] | L [] | HZi [au] | HZm [au] | HZo [au] | S [W m−2] | S [W m−2] | S [W m−2] |
|---|---|---|---|---|---|---|---|---|---|
| A5 V | 8250 | 1.70 | 12.08 | 2.50 | 3.46 | 3.91 | 2624.57 | 1370.04 | 1075.02 |
| G2 V | 5750 | 1.00 | 0.99 | 0.71 | 0.99 | 1.12 | 2624.57 | 1370.04 | 1075.02 |
| M8 V | 2500 | 0.12 | 5.2 × 10 | 0.02 | 0.02 | 0.02 | 2624.57 | 1370.04 | 1075.02 |
| Exoplanet Atmosphere | Pigment | [s−1] | [s−1] | [s−1] | ppfd [μmol photon m−2 s−1] | S [W m−2] |
|---|---|---|---|---|---|---|
| Earth- like | BChlA | 11.42 | 4.78 | 6.65 | 2400.48 | 1370.04 |
| BChlB | 15.68 | 8.92 | 6.77 | 2400.48 | 1370.04 | |
| ChlA | 13.54 | 9.79 | 3.75 | 2400.48 | 1370.04 | |
| ChlB | 16.24 | 12.96 | 3.27 | 2400.48 | 1370.04 | |
| Phot0 | 6.37 | 1.91 | 4.46 | 2400.48 | 1370.04 | |
|
Highly oxidizing | BChlA | 10.06 | 3.20 | 6.87 | 2146.82 | 1370.04 |
| BChlB | 13.31 | 6.29 | 7.02 | 2146.82 | 1370.04 | |
| ChlA | 10.78 | 7.12 | 3.66 | 2146.82 | 1370.04 | |
| ChlB | 13.21 | 10.03 | 3.18 | 2146.82 | 1370.04 | |
| Phot0 | 5.23 | 1.01 | 4.22 | 2146.82 | 1370.04 | |
| Weakly oxidizing | BChlA | 11.93 | 4.76 | 7.17 | 2442.12 | 1370.04 |
| BChlB | 16.14 | 8.83 | 7.31 | 2442.12 | 1370.04 | |
| ChlA | 13.76 | 9.88 | 3.88 | 2442.12 | 1370.04 | |
| ChlB | 16.32 | 12.92 | 3.40 | 2442.12 | 1370.04 | |
| Phot0 | 6.47 | 1.87 | 4.60 | 2442.12 | 1370.04 | |
| Reducing | BChlA | 13.77 | 6.39 | 7.38 | 2676.98 | 1370.04 |
| BChlB | 18.80 | 11.30 | 7.51 | 2676.98 | 1370.04 | |
| ChlA | 16.77 | 12.73 | 4.04 | 2676.98 | 1370.04 | |
| ChlB | 19.10 | 15.54 | 3.56 | 2676.98 | 1370.04 | |
| Phot0 | 7.78 | 2.90 | 4.88 | 2676.98 | 1370.04 |
| HZ | Exoplanet Atmosphere | Pigment | [s−1] | [s−1] | [s−1] | ppfd [μmol photon m−2 s−1] | S [W m−2] |
|---|---|---|---|---|---|---|---|
| HZi | Earth- like | BChlA | 22.05 | 4.73 | 17.32 | 4052.63 | 2624.57 |
| HZi | BChlB | 26.94 | 8.71 | 18.24 | 4052.63 | 2624.57 | |
| HZi | ChlA | 17.63 | 8.91 | 8.72 | 4052.63 | 2624.57 | |
| HZi | ChlB | 21.35 | 14.39 | 6.96 | 4052.63 | 2624.57 | |
| HZi | Phot0 | 10.42 | 2.07 | 8.35 | 4052.63 | 2624.57 | |
| HZi | Highly oxidizing | BChlA | 21.16 | 3.15 | 18.00 | 3741.48 | 2624.57 |
| HZi | BChlB | 25.19 | 6.17 | 19.02 | 3741.48 | 2624.57 | |
| HZi | ChlA | 15.02 | 6.47 | 8.56 | 3741.48 | 2624.57 | |
| HZi | ChlB | 18.02 | 11.23 | 6.79 | 3741.48 | 2624.57 | |
| HZi | Phot0 | 9.01 | 1.11 | 7.90 | 3741.48 | 2624.57 | |
| HZi | Weakly oxidizing | BChlA | 23.44 | 4.73 | 18.72 | 4157.44 | 2624.57 |
| HZi | BChlB | 28.35 | 8.64 | 19.71 | 4157.44 | 2624.57 | |
| HZi | ChlA | 17.98 | 8.94 | 9.04 | 4157.44 | 2624.57 | |
| HZi | ChlB | 21.59 | 14.35 | 7.25 | 4157.44 | 2624.57 | |
| HZi | Phot0 | 10.65 | 2.04 | 8.62 | 4157.44 | 2624.57 | |
| HZi | Reducing | BChlA | 25.57 | 6.37 | 19.20 | 4480.02 | 2624.57 |
| HZi | BChlB | 31.23 | 11.06 | 20.17 | 4480.02 | 2624.57 | |
| HZi | ChlA | 20.73 | 11.34 | 9.39 | 4480.02 | 2624.57 | |
| HZi | ChlB | 24.73 | 17.16 | 7.57 | 4480.02 | 2624.57 | |
| HZi | Phot0 | 12.30 | 3.16 | 9.14 | 4480.02 | 2624.57 | |
| HZm | Earth- like | BChlA | 11.51 | 2.47 | 9.04 | 2115.69 | 1370.04 |
| HZm | BChlB | 14.07 | 4.54 | 9.52 | 2115.69 | 1370.04 | |
| HZm | ChlA | 9.20 | 4.65 | 4.55 | 2115.69 | 1370.04 | |
| HZm | ChlB | 11.14 | 7.51 | 3.63 | 2115.69 | 1370.04 | |
| HZm | Phot0 | 5.44 | 1.08 | 4.36 | 2115.69 | 1370.04 | |
| HZm | Highly oxidizing | BChlA | 11.03 | 1.64 | 9.39 | 1950.40 | 1370.04 |
| HZm | BChlB | 13.13 | 3.21 | 9.92 | 1950.40 | 1370.04 | |
| HZm | ChlA | 7.82 | 3.36 | 4.46 | 1950.40 | 1370.04 | |
| HZm | ChlB | 9.39 | 5.84 | 3.54 | 1950.40 | 1370.04 | |
| HZm | Phot0 | 4.70 | 0.57 | 4.12 | 1950.40 | 1370.04 | |
| HZm | Weakly oxidizing | BChlA | 12.23 | 2.46 | 9.77 | 2168.68 | 1370.04 |
| HZm | BChlB | 14.79 | 4.50 | 10.28 | 2168.68 | 1370.04 | |
| HZm | ChlA | 9.37 | 4.65 | 4.72 | 2168.68 | 1370.04 | |
| HZm | ChlB | 11.26 | 7.48 | 3.78 | 2168.68 | 1370.04 | |
| HZm | Phot0 | 5.55 | 1.06 | 4.50 | 2168.68 | 1370.04 | |
| HZm | Reducing | BChlA | 13.35 | 3.32 | 10.02 | 2338.10 | 1370.04 |
| HZm | BChlB | 16.30 | 5.77 | 10.53 | 2338.10 | 1370.04 | |
| HZm | ChlA | 10.82 | 5.92 | 4.90 | 2338.10 | 1370.04 | |
| HZm | ChlB | 12.90 | 8.95 | 3.95 | 2338.10 | 1370.04 | |
| HZm | Phot0 | 6.42 | 1.65 | 4.77 | 2338.10 | 1370.04 | |
| HZo | Earth- like | BChlA | 9.03 | 1.94 | 7.10 | 1660.29 | 1075.02 |
| HZo | BChlB | 11.04 | 3.57 | 7.47 | 1660.29 | 1075.02 | |
| HZo | ChlA | 7.22 | 3.65 | 3.57 | 1660.29 | 1075.02 | |
| HZo | ChlB | 8.74 | 5.89 | 2.85 | 1660.29 | 1075.02 | |
| HZo | Phot0 | 4.27 | 0.85 | 3.42 | 1660.29 | 1075.02 | |
| HZo | Highly oxidizing | BChlA | 8.68 | 1.30 | 7.38 | 1534.67 | 1075.02 |
| HZo | BChlB | 10.33 | 2.54 | 7.79 | 1534.67 | 1075.02 | |
| HZo | ChlA | 6.17 | 2.66 | 3.51 | 1534.67 | 1075.02 | |
| HZo | ChlB | 7.40 | 4.61 | 2.79 | 1534.67 | 1075.02 | |
| HZo | Phot0 | 3.70 | 0.46 | 3.24 | 1534.67 | 1075.02 | |
| HZo | Weakly oxidizing | BChlA | 9.61 | 1.94 | 7.67 | 1704.13 | 1075.02 |
| HZo | BChlB | 11.62 | 3.55 | 8.07 | 1704.13 | 1075.02 | |
| HZo | ChlA | 7.37 | 3.67 | 3.70 | 1704.13 | 1075.02 | |
| HZo | ChlB | 8.86 | 5.89 | 2.97 | 1704.13 | 1075.02 | |
| HZo | Phot0 | 4.37 | 0.84 | 3.53 | 1704.13 | 1075.02 | |
| HZo | Reducing | BChlA | 10.48 | 2.61 | 7.87 | 1835.42 | 1075.02 |
| HZo | BChlB | 12.80 | 4.53 | 8.26 | 1835.42 | 1075.02 | |
| HZo | ChlA | 8.49 | 4.65 | 3.85 | 1835.42 | 1075.02 | |
| HZo | ChlB | 10.13 | 7.03 | 3.10 | 1835.42 | 1075.02 | |
| HZo | Phot0 | 5.04 | 1.30 | 3.74 | 1835.42 | 1075.02 |
| Exoplanet Atmosphere | Pigment | [s−1] | [s−1] | [s−1] | ppfd [μmol photon m−2 s−1] | S [W m−2] |
|---|---|---|---|---|---|---|
| Earth- like | BChlA | 0.95 | 1.42 × 10−3 | 0.94 | 16.48 | 1370.04 |
| BChlB | 1.63 | 3.59 × 10−3 | 1.62 | 16.48 | 1370.04 | |
| ChlA | 0.07 | 2.20 ×10−3 | 0.07 | 16.48 | 1370.04 | |
| ChlB | 0.06 | 1.46 × 10−2 | 0.05 | 16.48 | 1370.04 | |
| Phot0 | 0.02 | 2.99 × 10−4 | 0.02 | 16.48 | 1370.04 | |
| Highly oxidizing | BChlA | 0.98 | 1.09 × 10−3 | 0.98 | 16.07 | 1370.04 |
| BChlB | 1.75 | 2.84 × 10−3 | 1.75 | 16.07 | 1370.04 | |
| ChlA | 0.07 | 1.67 × 10−3 | 0.07 | 16.07 | 1370.04 | |
| ChlB | 0.06 | 1.18 × 10−2 | 0.04 | 16.07 | 1370.04 | |
| Phot0 | 0.02 | 1.56 × 10−4 | 0.02 | 16.07 | 1370.04 | |
| Weakly oxidizing | BChlA | 1.01 | 1.41 × 10−3 | 1.01 | 17.17 | 1370.04 |
| BChlB | 1.79 | 3.57 × 10−3 | 1.79 | 17.17 | 1370.04 | |
| ChlA | 0.07 | 2.17 × 10−3 | 0.07 | 17.17 | 1370.04 | |
| ChlB | 0.06 | 1.45 × 10−2 | 0.05 | 17.17 | 1370.04 | |
| Phot0 | 0.02 | 2.93 × 10−4 | 0.02 | 17.17 | 1370.04 | |
| Reducing | BChlA | 1.03 | 1.72 × 10−3 | 1.03 | 17.98 | 1370.04 |
| BChlB | 1.82 | 4.20 × 10−3 | 1.82 | 17.98 | 1370.04 | |
| ChlA | 0.08 | 2.62 × 10−3 | 0.07 | 17.98 | 1370.04 | |
| ChlB | 0.07 | 1.67 × 10−2 | 0.05 | 17.98 | 1370.04 | |
| Phot0 | 0.02 | 4.64 × 10−4 | 0.02 | 17.98 | 1370.04 |
| 1 | Further information on the ExoPhot project, data, and processing details can be found at http://github.com/ExoPhotProject (accessed on 22 November 2022). |
| 2 | http://svo2.cab.inta-csic.es/theory/vosa/ (accessed on 22 November 2022). |
| 3 | https://psg.gsfc.nasa.gov/ (accessed on 22 November 2022). |
| 4 | http://github.com/ExoPhotProject (accessed on 22 November 2022). |
References
- Cockell, C.S. ‘Astrobiology’ and the ethics of new science. Interdiscip. Sci. Rev. 2001, 26, 90–96. [Google Scholar] [CrossRef]
- Mayor, M.; Queloz, D. A Jupiter-mass companion to a solar-type star. Nature 1995, 378, 355–359. [Google Scholar] [CrossRef]
- Kasting, J.F.; Whitmire, D.P.; Reynolds, R.T. Habitable Zones around Main Sequence Stars. Icarus 1993, 101, 108–128. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C.; Thompson, W.R.; Carlson, R.; Gurnett, D.; Hord, C. A search for life on Earth from the Galileo spacecraft. Nature 1993, 365, 715–721. [Google Scholar] [CrossRef]
- Des Marais, D.J.; Harwit, M.O.; Jucks, K.W.; Kasting, J.F.; Lin, D.N.C.; Lunine, J.I.; Schneider, J.; Seager, S.; Traub, W.A.; Woolf, N.J. Remote Sensing of Planetary Properties and Biosignatures on Extrasolar Terrestrial Planets. Astrobiology 2002, 2, 153–181. [Google Scholar] [CrossRef]
- Schwieterman, E.W.; Kiang, N.Y.; Parenteau, M.N.; Harman, C.E.; DasSarma, S.; Fisher, T.M.; Arney, G.N.; Hartnett, H.E.; Reinhard, C.T.; Olson, S.L.; et al. Exoplanet Biosignatures: A Review of Remotely Detectable Signs of Life. Astrobiology 2018, 18, 663–708. [Google Scholar] [CrossRef]
- Quanz, S.P.; Ottiger, M.; Fontanet, E.; Kammerer, J.; Menti, F.; Dannert, F.; Gheorghe, A.; Absil, O.; Airapetian, V.S.; Alei, E.; et al. Large Interferometer For Exoplanets (LIFE). Astron. Astrophys. 2022, 664, A21. [Google Scholar] [CrossRef]
- Hall, D.O.; Rao, K. Photosynthesis; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; John Wiley & Sons: Chichester, UK, 2021. [Google Scholar]
- Gale, J.; Wandel, A. The potential of planets orbiting red dwarf stars to support oxygenic photosynthesis and complex life. Int. J. Astrobiol. 2017, 16, 1–9. [Google Scholar] [CrossRef]
- Hu, R.; Seager, S.; Bains, W. Photochemistry in Terrestrial Exoplanet Atmospheres. I. Photochemistry Model and Benchmark Cases. Astrophys. J. 2012, 761, 166. [Google Scholar] [CrossRef]
- Trifonov, T.; Caballero, J.; Morales, J.; Seifahrt, A.; Ribas, I.; Reiners, A.; Bean, J.; Luque, R.; Parviainen, H.; Pallé, E.; et al. A nearby transiting rocky exoplanet that is suitable for atmospheric investigation. Science 2021, 371, 1038–1041. [Google Scholar] [CrossRef]
- García de la Concepción, J.; Cerdán, L.; Marcos-Arenal, P.; Burillo-Villalobos, M.; Fonseca-Bonilla, N.; Lizcano-Vaquero, R.; Ángeles López-Cayuela, M.; Gómez, F.; Caballero, J.A. Phot0, a plausible primeval pigment on Earth and rocky exoplanets. Phys. Chem. Chem. Phys. 2022, 24, 16979–16987. [Google Scholar] [CrossRef] [PubMed]
- Berdyugina, S.V.; Kuhn, J.R.; Harrington, D.M.; Šantl Temkiv, T.; Messersmith, E.J. Remote sensing of life: Polarimetric signatures of photosynthetic pigments as sensitive biomarkers. Int. J. Astrobiol. 2016, 15, 45–56. [Google Scholar] [CrossRef]
- Kolokolova, L.; Hough, J.H.; Levasseur-Regourd, A.C. Polarimetry of Stars and Planetary Systems; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Kiang, N.Y.; Segura, A.; Tinetti, G.; Govindjee; Blankenship, R.E.; Cohen, M.; Siefert, J.; Crisp, D.; Meadows, V.S. Spectral Signatures of Photosynthesis. II. Coevolution with Other Stars And The Atmosphere on Extrasolar Worlds. Astrobiology 2007, 7, 252–274. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, Y.; Umemura, M.; Shoji, M.; Kayanuma, M.; Yabana, K.; Shiraishi, K. Light absorption efficiencies of photosynthetic pigments: The dependence on spectral types of central stars. Int. J. Astrobiol. 2015, 14, 505–510. [Google Scholar] [CrossRef]
- Ritchie, R.J.; Larkum, A.W.; Ribas, I. Could photosynthesis function on Proxima Centauri b? Int. J. Astrobiol. 2018, 17, 147–176. [Google Scholar] [CrossRef]
- Mullan, D.J.; Bais, H.P. Photosynthesis on a Planet Orbiting an M Dwarf: Enhanced Effectiveness during Flares. Astrophys. J. 2018, 865, 101. [Google Scholar] [CrossRef]
- Lehmer, O.R.; Catling, D.C.; Parenteau, M.N.; Hoehler, T.M. The Productivity of Oxygenic Photosynthesis around Cool, M Dwarf Stars. Astrophys. J. 2018, 859, 171. [Google Scholar] [CrossRef]
- Lehmer, O.R.; Catling, D.C.; Parenteau, M.N.; Kiang, N.Y.; Hoehler, T.M. The Peak Absorbance Wavelength of Photosynthetic Pigments Around Other Stars From Spectral Optimization. Front. Astron. Space Sci. 2021, 8, 689441. [Google Scholar] [CrossRef]
- Lingam, M.; Loeb, A. Photosynthesis on habitable planets around low-mass stars. Mon. Not. RAS 2019, 485, 5924–5928. Available online: https://academic.oup.com/mnras/article-pdf/485/4/5924/28271824/stz847.pdf (accessed on 22 November 2022).
- Lingam, M.; Loeb, A. Constraints on Aquatic Photosynthesis for Terrestrial Planets around Other Stars. Astrophys. J. 2020, 889, l15. [Google Scholar] [CrossRef]
- Covone, G.; Ienco, R.M.; Cacciapuoti, L.; Inno, L. Efficiency of the oxygenic photosynthesis on Earth-like planets in the habitable zone. Mon. Not. RAS 2021, 505, 3329–3335. Available online: https://academic.oup.com/mnras/article-pdf/505/3/3329/38673779/stab1357.pdf (accessed on 22 November 2022).
- Lingam, M.; Balbi, A.; Mahajan, S.M. Excitation Properties of Photopigments and Their Possible Dependence on the Host Star. Astrophys. J. 2021, 921, L41. [Google Scholar] [CrossRef]
- Mondragón-Solórzano, G.; Sandoval-Lira, J.; Nochebuena, J.; Cisneros, G.A.; Barroso-Flores, J. Electronic Structure Effects Related to the Origin of the Remarkable Near-Infrared Absorption of Blastochloris viridis’ Light Harvesting 1-Reaction Center Complex. J. Chem. Theory Comput. 2022, 18, 4555–4564. [Google Scholar] [CrossRef] [PubMed]
- McCree, K. Test of current definitions of photosynthetically active radiation against leaf photosynthesis data. Agric. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- Madronich, S. Photodissociation in the atmosphere: 1. Actinic flux and the effects of ground reflections and clouds. J. Geophys. Res. Atmos. 1987, 92, 9740–9752. Available online: https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/JD092iD08p09740 (accessed on 22 November 2022).
- McCree, K. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 328. [Google Scholar] [CrossRef]
- Desidera, S. Properties of Hypothetical Planetary Systems around the Brown Dwarf Gliese 229B. Publ. ASP 1999, 111, 1529–1538. [Google Scholar] [CrossRef]
- Caballero, J.A. Formation, Evolution and Multiplicity of Brown Dwarfs and Giant Exoplanets. Astrophys. Space Sci. Proc. 2010, 14, 79–90. [Google Scholar] [CrossRef]
- Gillon, M.; Jehin, E.; Lederer, S.M.; Delrez, L.; de Wit, J.; Burdanov, A.; Van Grootel, V.; Burgasser, A.J.; Triaud, A.H.M.J.; Opitom, C.; et al. Temperate Earth-sized planets transiting a nearby ultracool dwarf star. Nature 2016, 533, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Gillon, M.; Triaud, A.H.M.J.; Demory, B.O.; Jehin, E.; Agol, E.; Deck, K.M.; Lederer, S.M.; de Wit, J.; Burdanov, A.; Ingalls, J.G.; et al. Seven temperate terrestrial planets around the nearby ultracool dwarf star TRAPPIST-1. Nature 2017, 542, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister, M.; Dreizler, S.; Ribas, I.; Reiners, A.; Caballero, J.A.; Bauer, F.F.; Béjar, V.J.S.; González-Cuesta, L.; Herrero, E.; Lalitha, S.; et al. The CARMENES search for exoplanets around M dwarfs. Two temperate Earth-mass planet candidates around Teegarden’s Star. Astron. Astrophys. 2019, 627, A49. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, H.; Caballero, J.A.; Cifuentes, C.; Piro, A.L.; Barnes, R. Exomoons in the Habitable Zones of M Dwarfs. Astrophys. J. 2019, 887, 261. [Google Scholar] [CrossRef]
- Scalo, J.; Kaltenegger, L.; Segura, A.G.; Fridlund, M.; Ribas, I.; Kulikov, Y.N.; Grenfell, J.L.; Rauer, H.; Odert, P.; Leitzinger, M.; et al. M Stars as Targets for Terrestrial Exoplanet Searches And Biosignature Detection. Astrobiology 2007, 7, 85–166. [Google Scholar] [CrossRef] [PubMed]
- Caballero, J.A.; González-Álvarez, E.; Brady, M.; Trifonov, T.; Ellis, T.G.; Dorn, C.; Cifuentes, C.; Molaverdikhani, K.; Bean, J.L.; Boyajian, T.; et al. A detailed analysis of the Gl 486 planetary system. A&A 2022, 665, A120. [Google Scholar] [CrossRef]
- Castelli, F.; Kurucz, R.L. New Grids of ATLAS9 Model Atmospheres. In Modelling of Stellar Atmospheres, Poster Contributions, Proceedings of the 210th Symposium of the International Astronomical Union, Uppsala University, Uppsala, Sweden, 17–21 June 2002; Piskunov, N., Weiss, W.W., Gray, D.F., Piskunov, N., Weiss, W.W., Gray, D.F., Eds.; Published on behalf of the IAU by the Astronomical Society of the Pacific; Cambridge University Press: Cambridge, UK, 2003; Volume 210, p. A20. Available online: https://arxiv.org/abs/astro-ph/0405087 (accessed on 22 November 2022).
- Baraffe, I.; Homeier, D.; Allard, F.; Chabrier, G. New evolutionary models for pre-main sequence and main sequence low-mass stars down to the hydrogen-burning limit. Astron. Astrophys. 2015, 577, 4–9. [Google Scholar] [CrossRef]
- Cifuentes, C.; Caballero, J.A.; Cortés-Contreras, M.; Montes, D.; Abellán, F.J.; Dorda, R.; Holgado, G.; Zapatero Osorio, M.R.; Morales, J.C.; Amado, P.J.; et al. CARMENES input catalogue of M dwarfs. Astron. Astrophys. 2020, 642, A115. [Google Scholar] [CrossRef]
- Bayo, A.; Rodrigo, C.; Barrado Y Navascués, D.; Solano, E.; Gutiérrez, R.; Morales-Calderón, M.; Allard, F. VOSA: Virtual observatory SED analyzer. An application to the Collinder 69 open cluster. Astron. Astrophys. 2008, 492, 277–287. [Google Scholar] [CrossRef]
- Villanueva, G.L.; Smith, M.D.; Protopapa, S.; Faggi, S.; Mandell, A.M. Planetary Spectrum Generator: An accurate online radiative transfer suite for atmospheres, comets, small bodies and exoplanets. J. Quant. Spectrosc. Radiat. Transf. 2018, 217, 86–104. [Google Scholar] [CrossRef]
- Kasting, J.F.; Brown, L.L. The early atmosphere as a source of biogenic compounds. In The Molecular Origins of Life: Assembling Pieces of the Puzzle; Brack, A., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 35–56. [Google Scholar] [CrossRef]
- Kasting, J.F. Runaway and moist greenhouse atmospheres and the evolution of Earth and Venus. Icarus 1988, 74, 472–494. [Google Scholar] [CrossRef]
- Taniguchi, M.; Lindsey, J.S. Absorption and Fluorescence Spectral Database of Chlorophylls and Analogues. Photochem. Photobiol. 2021, 97, 136–165. [Google Scholar] [CrossRef] [PubMed]
- Sirohiwal, A.; Berraud-Pache, R.; Neese, F.; Izsák, R.; Pantazis, D.A. Accurate Computation of the Absorption Spectrum of Chlorophyll a with Pair Natural Orbital Coupled Cluster Methods. J. Phys. Chem. B 2020, 124, 8761–8771. [Google Scholar] [CrossRef]
- Mirkovic, T.; Ostroumov, E.E.; Anna, J.M.; van Grondelle, R.; Govindjee; Scholes, G.D. Light Absorption and Energy Transfer in the Antenna Complexes of Photosynthetic Organisms. Chem. Rev. 2017, 117, 249–293. [Google Scholar] [CrossRef]
- Kurashov, V.; Ho, M.Y.; Shen, G.; Piedl, K.; Laremore, T.N.; Bryant, D.A.; Golbeck, J.H. Energy transfer from chlorophyll f to the trapping center in naturally occurring and engineered Photosystem I complexes. Photosynth. Res. 2019, 141, 151–163. [Google Scholar] [CrossRef]
- Claudi, R.; Alei, E.; Battistuzzi, M.; Cocola, L.; Erculiani, M.S.; Pozzer, A.C.; Salasnich, B.; Simionato, D.; Squicciarini, V.; Poletto, L.; et al. Super-Earths, M Dwarfs, and Photosynthetic Organisms: Habitability in the Lab. Life 2021, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Kopparapu, R.K.; Ramirez, R.; Kasting, J.F.; Eymet, V.; Robinson, T.D.; Mahadevan, S.; Terrien, R.C.; Domagal-Goldman, S.; Meadows, V.; Deshpande, R. Habitable Zones around Main-sequence Stars: New Estimates. Astrophys. J. 2013, 765, 131. [Google Scholar] [CrossRef]
- McKay, C.P. Requirements and limits for life in the context of exoplanets. Proc. Natl. Acad. Sci. USA 2014, 111, 12628–12633. Available online: https://www.pnas.org/content/111/35/12628.full.pdf (accessed on 22 November 2022). [PubMed]
- Hashimoto, G.L.; Abe, Y.; Sugita, S. The chemical composition of the early terrestrial atmosphere: Formation of a reducing atmosphere from CI-like material. J. Geophys. Res. Planets 2007, 112, 5010. [Google Scholar] [CrossRef]
- Henry, T.J.; Jao, W.C.; Subasavage, J.P.; Beaulieu, T.D.; Ianna, P.A.; Costa, E.; Méndez, R.A. The Solar Neighborhood. XVII. Parallax Results from the CTIOPI 0.9 m Program: 20 New Members of the RECONS 10 Parsec Sample. Astron. J. 2006, 132, 2360–2371. [Google Scholar] [CrossRef]
- Reylé, C.; Jardine, K.; Fouqué, P.; Caballero, J.A.; Smart, R.L.; Sozzetti, A. The 10 parsec sample in the Gaia era. A&A 2021, 650, A201. [Google Scholar] [CrossRef]


| Exopl. Atmos. | ppfd [mol photon m−2 s−1] | Pigment | [s−1] | [s−1] | [s−1] |
|---|---|---|---|---|---|
| Earth- like | 2115.69 | BChl a | 11.51 | 2.47 | 9.04 |
| BChl b | 14.07 | 4.54 | 9.52 | ||
| Chl a | 9.20 | 4.65 | 4.55 | ||
| Chl b | 11.14 | 7.51 | 3.63 | ||
| Phot0 | 6.72 | 1.33 | 5.39 | ||
| Highly oxidizing | 1950.40 | BChl a | 13.35 | 3.32 | 10.02 |
| BChl b | 16.30 | 5.77 | 10.53 | ||
| Chl a | 10.82 | 5.92 | 4.90 | ||
| Chl b | 12.90 | 8.95 | 3.95 | ||
| Phot0 | 8.00 | 2.01 | 5.99 | ||
| Weakly oxidizing | 2168.68 | BChl a | 12.23 | 2.46 | 9.77 |
| BChl b | 14.79 | 4.50 | 10.28 | ||
| Chl a | 9.37 | 4.65 | 4.72 | ||
| Chl b | 11.26 | 7.48 | 3.78 | ||
| Phot0 | 7.15 | 1.32 | 5.84 | ||
| Reducing | 2338.10 | BChl a | 11.03 | 1.64 | 9.39 |
| BChl b | 13.13 | 3.21 | 9.92 | ||
| Chl a | 7.82 | 3.36 | 4.46 | ||
| Chl b | 9.39 | 5.84 | 3.54 | ||
| Phot0 | 6.34 | 0.73 | 5.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcos-Arenal, P.; Cerdán, L.; Burillo-Villalobos, M.; Fonseca-Bonilla, N.; de la Concepción, J.G.; López-Cayuela, M.Á.; Gómez, F.; Caballero, J.A. ExoPhot: The Photon Absorption Rate as a New Metric for Quantifying the Exoplanetary Photosynthetic Activity Fitness. Universe 2022, 8, 624. https://doi.org/10.3390/universe8120624
Marcos-Arenal P, Cerdán L, Burillo-Villalobos M, Fonseca-Bonilla N, de la Concepción JG, López-Cayuela MÁ, Gómez F, Caballero JA. ExoPhot: The Photon Absorption Rate as a New Metric for Quantifying the Exoplanetary Photosynthetic Activity Fitness. Universe. 2022; 8(12):624. https://doi.org/10.3390/universe8120624
Chicago/Turabian StyleMarcos-Arenal, Pablo, Luis Cerdán, Mercedes Burillo-Villalobos, Nuria Fonseca-Bonilla, Juan García de la Concepción, María Ángeles López-Cayuela, Felipe Gómez, and José A. Caballero. 2022. "ExoPhot: The Photon Absorption Rate as a New Metric for Quantifying the Exoplanetary Photosynthetic Activity Fitness" Universe 8, no. 12: 624. https://doi.org/10.3390/universe8120624
APA StyleMarcos-Arenal, P., Cerdán, L., Burillo-Villalobos, M., Fonseca-Bonilla, N., de la Concepción, J. G., López-Cayuela, M. Á., Gómez, F., & Caballero, J. A. (2022). ExoPhot: The Photon Absorption Rate as a New Metric for Quantifying the Exoplanetary Photosynthetic Activity Fitness. Universe, 8(12), 624. https://doi.org/10.3390/universe8120624