Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Rat Brain Lipidomics
2.1.1. Lipid Annotation and Identification
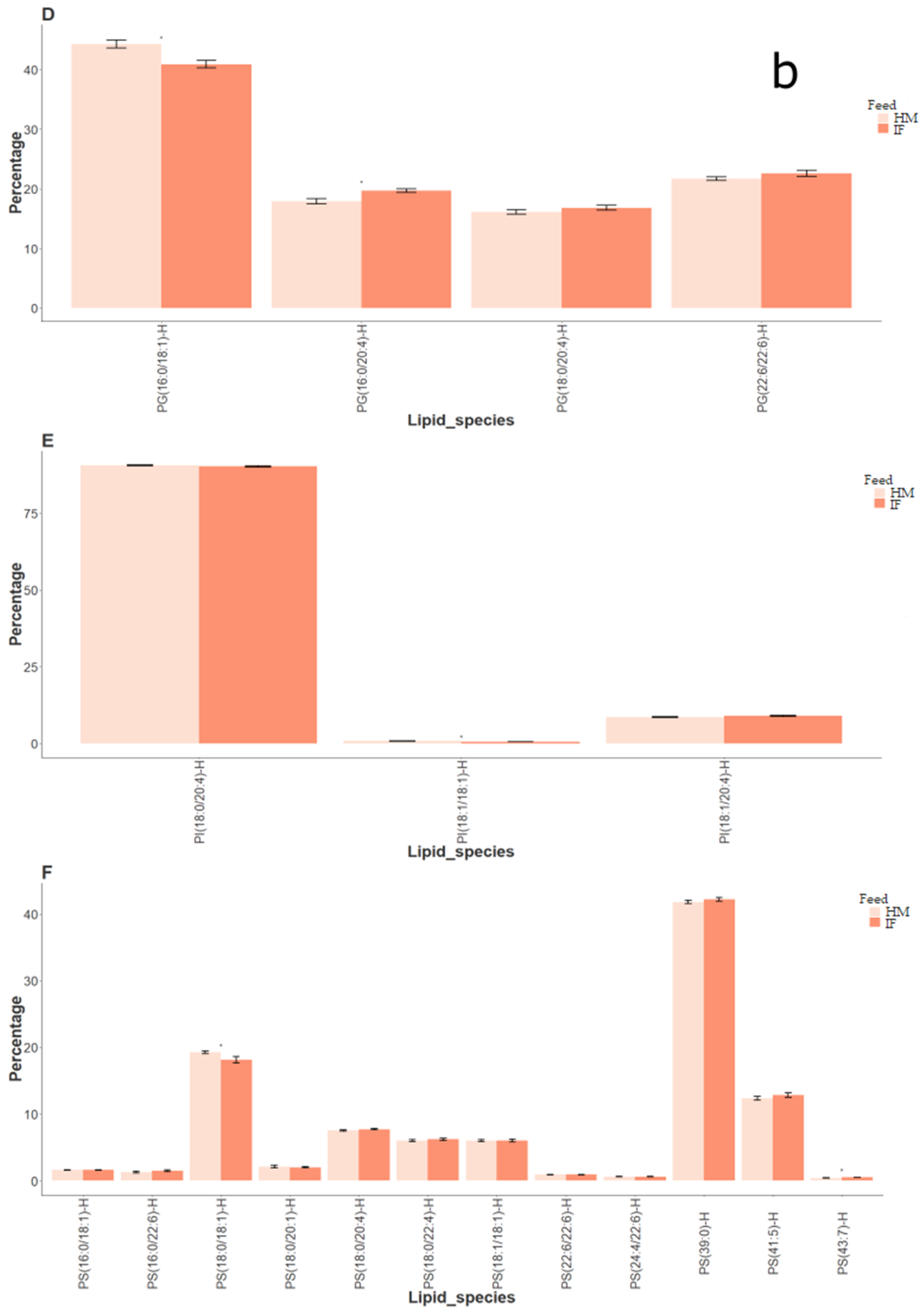
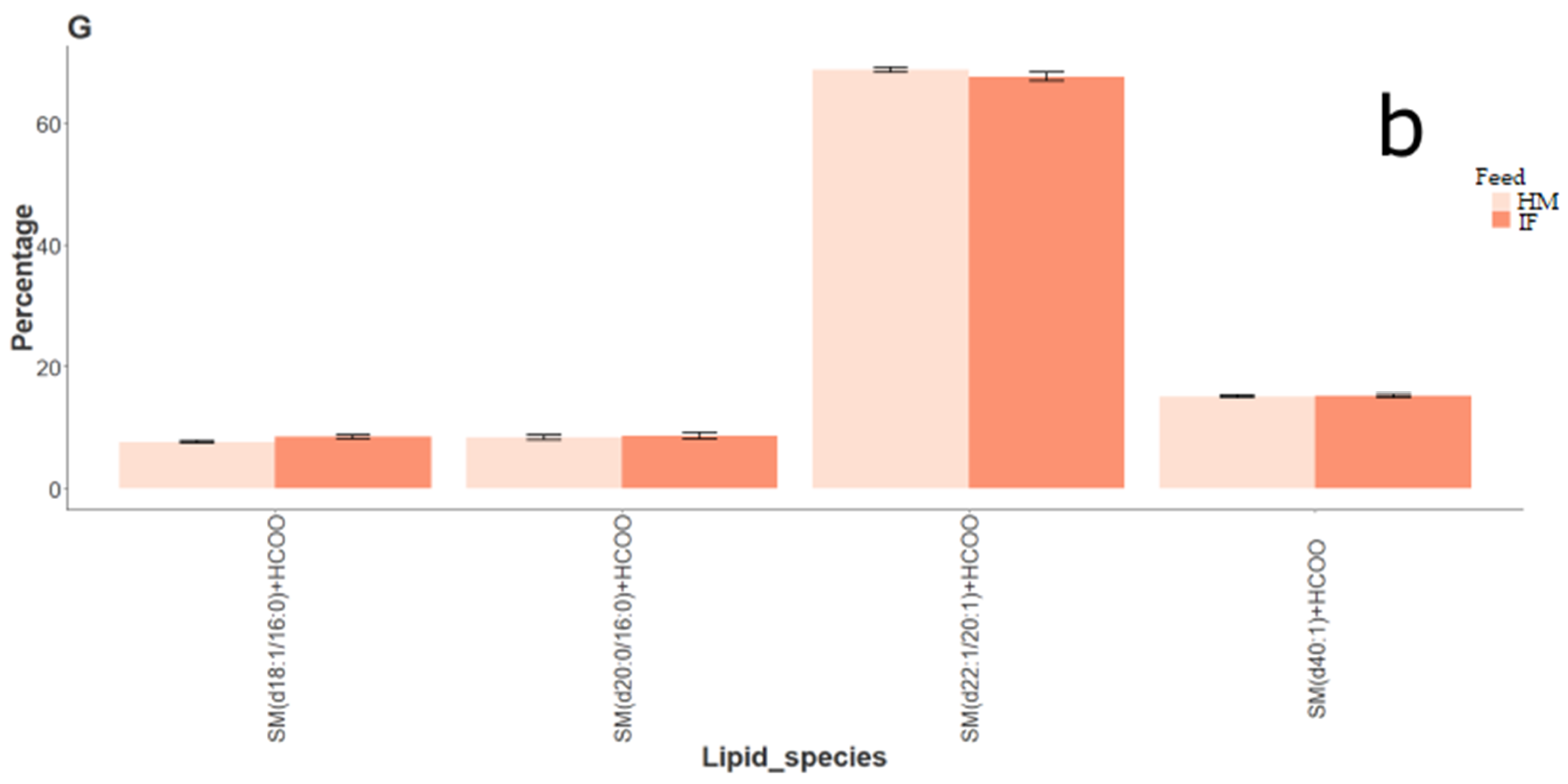
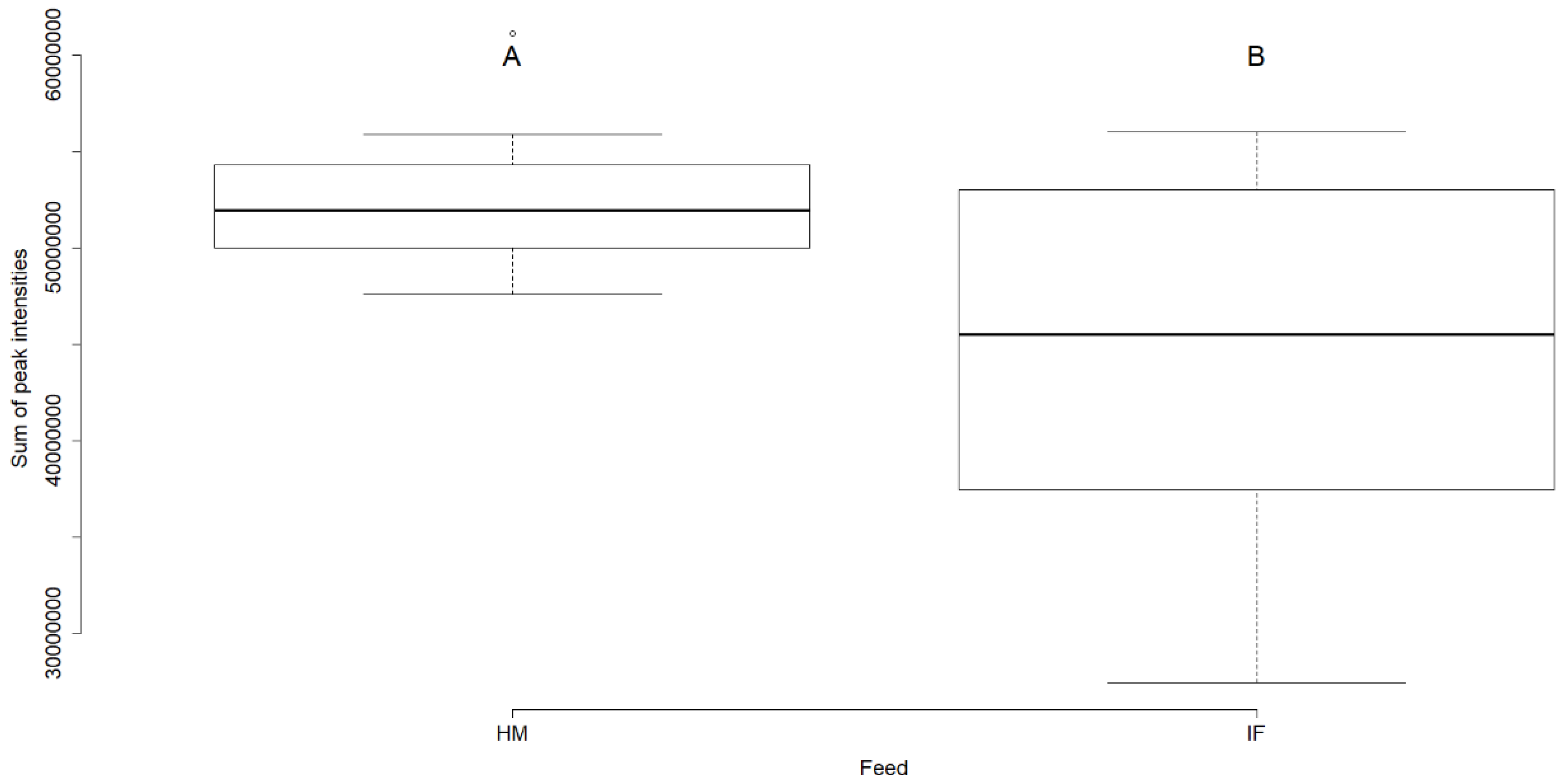
2.1.2. Lipid Composition of Rat Brain Samples (Human Milk vs. Infant Formula)
2.2. Human Milk and Infant Formula Composition
2.2.1. Fatty Acid Composition
2.2.2. Triacylglycerol (TAG) Composition
2.2.3. Phospholipid Composition
3. Discussion
3.1. Rat Brain Lipidomics
3.2. Human Milk and Infant Formula Lipid Composition
4. Materials and Methods
4.1. Sample Collection
4.2. Rat Growth Conditions, Treatment, and Ethics
4.3. Rat Brain Lipidomics
4.4. UHPLC-MS Conditions
4.4.1. Run Sequence
4.4.2. Data Analysis
4.4.3. Lipid Annotation and Identification
4.4.4. Chemometrics
4.5. Human Milk and Infant Formula Composition
4.5.1. Fatty Acid Analyses
4.5.2. PL Analysis of Milk Samples
4.5.3. TAG Analyses
4.6. Ethics Statement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yum, J. The effects of breast milk versus infant formulae on cognitive development. J. Dev. Disabil. 2007, 13, 135–164. [Google Scholar]
- Koletzko, B.; Rodriguez-Palmero, M.; Demmelmair, H.; Fidler, N.; Jensen, R.; Sauerwald, T. Physiological aspects of human milk lipids. Early Hum. Dev. 2001, 65, S3–S18. [Google Scholar] [CrossRef]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef]
- Reynolds, A. Breastfeeding and brain development. Pediatr. Clin. N. Am. 2001, 48, 159–171. [Google Scholar] [CrossRef]
- Isaacs, E.B.; Fischl, B.R.; Quinn, B.T.; Chong, W.K.; Gadian, D.G.; Lucas, A. Impact of breast milk on intelligence quotient, brain size, and white matter development. Pediatr. Res. 2010, 67, 357. [Google Scholar] [CrossRef] [PubMed]
- Uauy, R.; Peirano, P. Breast is Best: Human Milk is the Optimal Food for Brain Development. Am. J. Clin. Nutr. 1999, 70, 433–434. [Google Scholar] [CrossRef][Green Version]
- Ilcol, Y.O.; Ozbek, R.; Hamurtekin, E.; Ulus, I.H. Choline status in newborns, infants, children, breast-feeding women, breast-fed infants and human breast milk. J. Nutr. Biochem. 2005, 16, 489–499. [Google Scholar] [CrossRef]
- Sullivan, S.; Schanler, R.J.; Kim, J.H.; Patel, A.L.; Trawoger, R.; Kiechl-Kohlendorfer, U.; Chan, G.M.; Blanco, C.L.; Abrams, S.; Cotten, C.M.; et al. An exclusively human milk-based diet is associated with a lower rate of necrotizing enterocolitis than a diet of human milk and bovine milk-based products. J. Pediatr. 2010, 156, 562–567. [Google Scholar] [CrossRef]
- Cavkll, B. Gastric emptying in infants fed human milk or infant formula. Acta Paediatr. 1981, 70, 639–641. [Google Scholar] [CrossRef]
- Mehrotra, V.; Sehgal, S.K.; Bangale, N.R. Fat structure and composition in human milk and infant formulas: Implications in infant health. Clin. Epidemiol. Glob. Health 2019, 7, 153–159. [Google Scholar] [CrossRef]
- Lopez, C. Milk fat globules enveloped by their biological membrane: Unique colloidal assemblies with a specific composition and structure. Curr. Opin. Colloid Interface Sci. 2011, 16, 391–404. [Google Scholar] [CrossRef]
- Carrie, I.; Clement, M.; De Javel, D.; Frances, H.; Bourre, J.M. Specific phospholipid fatty acid composition of brain regions in mice: Effects of n–3 polyunsaturated fatty acid deficiency and phospholipid supplementation. J. Lipid Res. 2000, 41, 465–472. [Google Scholar] [PubMed]
- Timby, N.; Domellof, E.; Hernell, O.; Lonnerdal, B.; Domellof, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, M.A.; Araujo, W.M.C.; Borgo, L.A.; Alencar, E.R. Lipid profile of different infant formulas for infants. PLoS ONE 2017, 12, e0177812. [Google Scholar] [CrossRef] [PubMed]
- Checa, A.; Bedia, C.; Jaumot, J. Lipidomic data analysis: Tutorial, practical guidelines and applications. Anal. Chim. Acta 2015, 885, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Roy, N.C.; Guo, Y.; Jia, H.; Ryan, L.; Samuelsson, L.; Thomas, A.; Plowman, J.; Clerens, S.; Day, L.; et al. Human Breast Milk and Infant Formulas Differentially Modify the Intestinal Microbiota in Human Infants and Host Physiology in Rats. J. Nutr. 2016, 146, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Wu, X.; Feng, Y. Fragmentation patterns of five types of phospholipids by ultra-high-performance liquid chromatography electrospray ionization quadrupole time-of-flight tandem mass spectrometry. Anal. Methods 2016, 8, 1319–1332. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Char, D.; Sheard, N.F. Choline, phosphatidylcholine and sphingomyelin in human and bovine milk and infant formulas. J. Nutr. 1986, 116, 50–58. [Google Scholar] [CrossRef]
- Velagapudi, V.R.; Hezaveh, R.; Reigstad, C.S.; Gopalacharyulu, P.; Yetukuri, L.; Islam, S.; Felin, J.; Perkins, R.; Boren, J.; Oresic, M.; et al. The gut microbiota modulates host energy and lipid metabolism in mice. J. Lipid Res. 2010, 51, 1101–1112. [Google Scholar] [CrossRef]
- Muller, C.P.; Reichel, M.; Muhle, C.; Rhein, C.; Gulbins, E.; Kornhuber, J. Brain membrane lipids in major depression and anxiety disorders. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 1052–1065. [Google Scholar] [CrossRef]
- Rosenberg, A.; Stern, N. Changes in sphingosine and fatty acid components of the gangliosides in developing rat and human brain. J. Lipid Res. 1966, 7, 122–131. [Google Scholar] [PubMed]
- Murphy, S.A.; Nicolaou, A. Lipidomics applications in health, disease and nutrition research. Mol. Nutr. Food Res. 2013, 57, 1336–1346. [Google Scholar] [CrossRef] [PubMed]
- Aidoud, N.; Delplanque, B.; Baudry, C.; Garcia, C.; Moyon, A.; Balasse, L.; Guillet, B.; Antona, C.; Darmaun, D.; Fraser, K.; et al. A combination of lipidomics, MS imaging, and PET scan imaging reveals differences in cerebral activity in rat pups according to the lipid quality of infant formulas. FASEB J. 2018, 32, 4776–4790. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Dietary (n-3) fatty acids and brain development. J. Nutr. 2007, 137, 855–859. [Google Scholar] [CrossRef]
- Sala Vila, A.; Castellote-Bargallo, A.I.; Rodriguez-Palmero-Seuma, M.; Lopez-Sabater, M.C. High.-performance liquid chromatography with evaporative light-scattering detection for the determination of phospholipid classes in human milk, infant formulas and phospholipid sources of long-chain polyunsaturated fatty acids. J. Chromatogr. A 2003, 1008, 73–80. [Google Scholar] [CrossRef]
- Farquharson, J.; Cockburn, F.; Patrick, W.A.; Jamieson, E.C.; Logan, R.W. Infant cerebral cortex phospholipid fatty-acid composition and diet. Lancet 1992, 340, 810–813. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Holman, J.D.; Tabb, D.L.; Mallick, P. Employing ProteoWizard to convert raw mass spectrometry data. Curr. Protoc. Bioinform. 2014, 46, 13.24.1–13.24.9. [Google Scholar]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Gr. Stat. 1996, 5, 299–314. [Google Scholar]
- Dunn, W.B.; Wilson, I.D.; Nicholls, A.W.; Broadhurst, D. The importance of experimental design and QC samples in large-scale and MS-driven untargeted metabolomic studies of humans. Bioanalysis 2012, 4, 2249–2264. [Google Scholar] [CrossRef] [PubMed]
- Parsons, H.M.; Ekman, D.R.; Collette, T.W.; Viant, M.R. Spectral relative standard deviation: A practical benchmark in metabolomics. Analyst 2009, 134, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, R.; Ishikawa, M. Precise and global identification of phospholipid molecular species by an Orbitrap mass spectrometer and automated search engine Lipid Search. J. Chromatogr. A 2010, 1217, 4229–4239. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef]
- Christie, W.W. Preparation of ester derivatives of fatty acids for chromatographic analysis. Adv. Lipid Methodol. 1993, 2, e111. [Google Scholar]
- Reis, M.G.; Roy, N.C.; Bermingham, E.N.; Ryan, L.; Bibiloni, R.; Young, W.; Krause, L.; Berger, B.; North, M.; Stelwagen, K.; et al. Impact of Dietary Dairy Polar Lipids on Lipid Metabolism of Mice Fed a High-Fat Diet. J. Agric. Food Chem. 2013, 61, 2729–2738. [Google Scholar] [CrossRef]
- Hamilton, R.J.; Hamilton, S. Lipid Analysis: A practical Approach; IRL Press: Oxford, UK, 1992. [Google Scholar]
- Murphy, R.C. Mass Spectrometry of Lipids; Springer US: New York, NY, USA; Philadelphia, PA, USA, 1993. [Google Scholar]
- Teng, F.; Reis, M.G.; Yang, L.; Ma, Y.; Day, L. In-depth lipidomic analysis of tri-, di-, and mono-acylglycerols released from milk fat after in vitro digestion. Food Chem. 2019, 297, 124976. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid (% of Total FA) | Human Milk | Infant Formula |
|---|---|---|
| C4:0 | 0.38 | |
| C6:0 | 0.34 | |
| C8:0 | 1.26 | |
| C10:0 | 1.13 | 1.21 |
| C12:0 | 4.26 | 7.25 |
| C14:0 | 4.02 | 4.10 |
| C14:1 | 0.12 | |
| C15:0 | 0.08 | 0.18 |
| C16:0 | 22.40 | 23.78 |
| C16:1 | 1.87 | 0.27 |
| C17:0 | 0.18 | 0.14 |
| C18:0 | 5.96 | 4.68 |
| C18:1 n9 | 30.41 | 34.03 |
| C18:1 n11 | 1.75 | 0.74 |
| C18:2 n6 | 23.44 | 17.52 |
| C18:3 n6 | 0.17 | |
| C18:3 n3 | 1.53 | 1.80 |
| C20:0 | 0.19 | 0.29 |
| C20:1 | 0.44 | 0.22 |
| C20:2 | 0.43 | |
| C20:3 n3 | 0.17 | 0.24 |
| C20:4 n6 | 0.53 | 0.48 |
| C20:5 n3 | 0.06 | |
| C22:5 | 0.16 | |
| C22:6 n3 | 0.46 | 0.28 |
| C24:0 | 0.18 | |
| C24:1 | 0.05 |
| Total Carbon Chain Length (% Total TAG) | Human Milk | Infant Formula | Putative Species |
|---|---|---|---|
| C30 | 0.9 | 0.82 | C14:0C12:0C4:0/C12:0C14:0C4:0/ C8:0C12:0C10:0/ C10:0C16:0C4:0/ C8:0C18:0C4:0/ C10:0C18:1C4:0/ C10:0C18:2C6:0 |
| C32 | 0.56 | 2.21 | C16:0C10:0C6:0/C10:0C16:0C6:0/ C14:0C14:0C4:0/ C16:0C12:0C4:0/C12:0C16:0C4:0/ C10:0C18:0C4:0 |
| C34 | 0.26 | 8.23 | C16:1C14:0C4:0/ C18:1C10:0C6:0/ C16:0C14:1C4:0/ C12:0C12:0C10:0/C12:0C10:0C12:0/ C10:0C18:0C6:0/ C14:0C8:0C12:0/C8:0C14:0C12:0/ C18:0C12:0C4:0/C12:0C18:0C4:0/ C16:0C14:0C4:0 |
| C36 | 1.06 | 12.3 | C18:2C14:0C4:0/ C8:0C18:1C10:0/ C18:1C14:0C4:0/C14:0C18:1C4:0/ C16:0C14:0C6:0/ C16:0C16:0C4:0 |
| C38 | 1.78 | 15.13 | C18:1C16:1C4:0/ C18:2C16:0C4:0/ C18:1C10:0C10:0/C10:0C18:1C10:0/ C18:1C16:0C4:0/C16:0C18:1C4:0/ C12:0C16:0C10:0/ C16:0C16:0C6:0/ C16:0C18:0C4:0 |
| C40 | 3.11 | 11.88 | C18:2C18:1C4:0/ C18:1C18:1C4:0/ C18:1C12:0C10:0/ C18:1C16:0C6:0/C16:0C18:1C6:0/ C18:0C18:1C4:0/ C16:0C14:0C10:0/ C16:0C18:0C6:0/C18:0C16:0C6:0 |
| C42 | 5.48 | 8.79 | C18:1C18:1C6:0/ C14:1C14:0C14:0/C14:0C14:1C14:0/ C18:1C10:0C14:0/C10:0C18:1C14:0/ C18:1C16:0C8:0/ C18:0C18:1C6:0/ C16:0C12:0C14:0/ C16:0C10:0C16:0/C16:0C16:0C10:0 |
| C44 | 9.01 | 5.74 | C10:0C16:0C18:3/ C18:1C8:0C18:1/C18:1C18:1C8:0/ C16:0C10:0C18:2/ C16:0C10:0C18:1/ C14:0C14:0C16:0/C14:0C16:0C14:0/ C18:0C10:0C16:0 |
| C46 | 13.15 | 4.49 | C16:0C12:0C18:3/ C14:1C14:0C18:1/C14:0C14:1C18:1/ C14:0C14:0C18:2/C14:0C18:2C14:0/ C16:0C12:0C18:2/ C16:0C12:0C18:1/ C14:0C16:1C16:0/ C18:0C10:0C18:1 |
| C48 | 12.45 | 3.08 | C16:0C14:0C18:3/ C16:0C14:0C18:2/ C16:0C14:1C18:1/ C16:0C14:0C18:1/ C18:0C14:0C16:0 |
| C50 | 19.5 | 7.3 | C18:1C14:0C18:2/ C16:0C16:1C18:2/ C18:1C14:0C18:1/C14:0C18:1C18:1/ C16:0C16:1C18:1/ C16:0C16:0C18:1/C16:0C18:1C16:0/ C16:0C18:0C16:0/C16:0C16:0C18:0/ |
| C52 | 30.98 | 16.82 | C16:0C18:2C18:2/C18:2C16:0C18:2/ C18:1C16:0C18:2/ C16:0C18:1C18:1/C18:1C16:0C18:1/ C18:0C16:0C18:1/ C18:0C16:0C18:0/C18:0C18:0C16:0 |
| C54 | 1.76 | 3.21 | C18:1C18:1C18:2/C18:1C18:2C18:1/ C18:1C18:1C18:1/ C18:0C18:2C18:1/ C18:1C18:0C18:1/C18:0C18:1C18:1/ C18:0C18:1C18:0/C18:0C18:0C18:1/ C18:0C18:0C18:0 |
| Phospholipid (mg/100 mL) | Human Milk | Infant Formula |
|---|---|---|
| PI | 1.13 ± 0.16 | 3.83 ± 0.56 |
| PE | 2.62 ± 0.26 | 5.10 ± 1.17 |
| PS | 1.46 ± 0.18 | 3.66 ± 0.22 |
| PC | 3.27 ± 0.53 | 8.40 ± 0.64 |
| SM | 3.39 ± 0.19 | 7.08 ± 1.30 |
| Total PL | 11.87 ± 1.32 | 28.11 ± 2.14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, M.; Subbaraj, A.K.; Fraser, K.; Qi, X.; Jia, H.; Chen, W.; Gomes Reis, M.; Agnew, M.; Day, L.; Roy, N.C.; et al. Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula. Metabolites 2019, 9, 253. https://doi.org/10.3390/metabo9110253
Su M, Subbaraj AK, Fraser K, Qi X, Jia H, Chen W, Gomes Reis M, Agnew M, Day L, Roy NC, et al. Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula. Metabolites. 2019; 9(11):253. https://doi.org/10.3390/metabo9110253
Chicago/Turabian StyleSu, Miya, Arvind K. Subbaraj, Karl Fraser, Xiaoyan Qi, Hongxin Jia, Wenliang Chen, Mariza Gomes Reis, Mike Agnew, Li Day, Nicole C. Roy, and et al. 2019. "Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula" Metabolites 9, no. 11: 253. https://doi.org/10.3390/metabo9110253
APA StyleSu, M., Subbaraj, A. K., Fraser, K., Qi, X., Jia, H., Chen, W., Gomes Reis, M., Agnew, M., Day, L., Roy, N. C., & Young, W. (2019). Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula. Metabolites, 9(11), 253. https://doi.org/10.3390/metabo9110253