Sphingolipid Metabolism Perturbations in Rett Syndrome

 , ,
, , 

Abstract
1. Introduction
2. Subjects and Methods
2.1. Participants and Clinical Data
2.2. Metabolic Analysis
2.3. Statistical Analyses
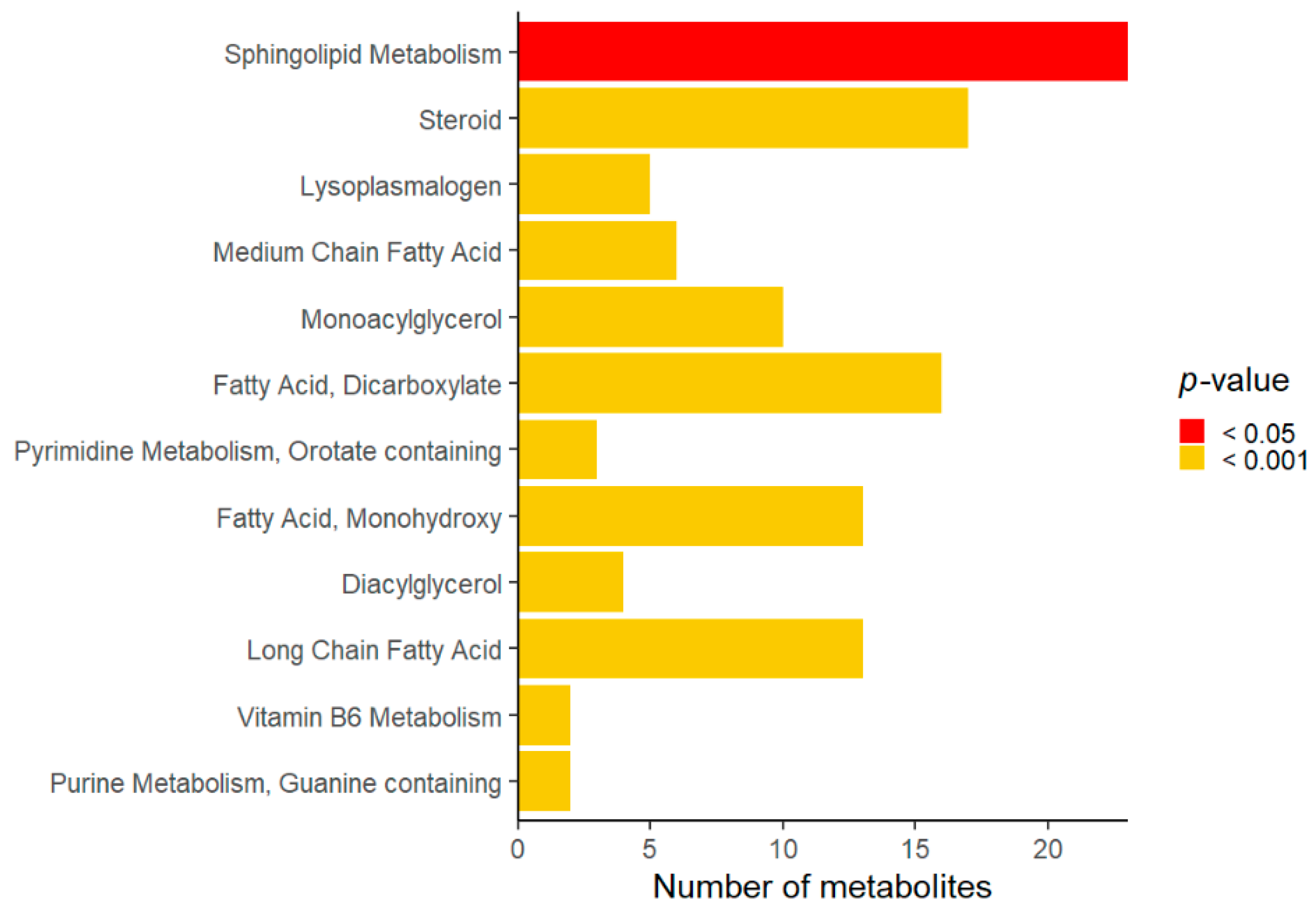
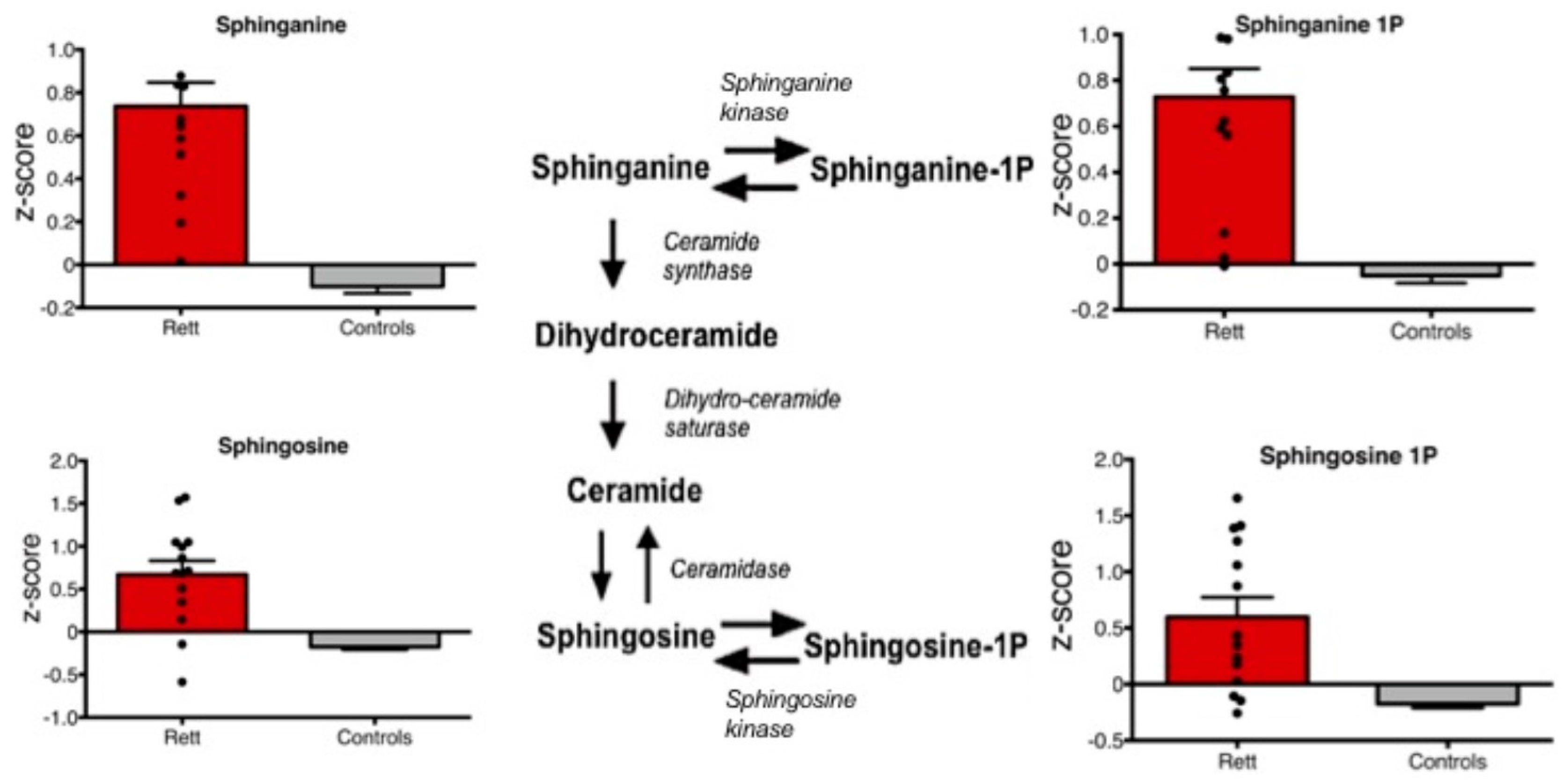
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miller, M.J.; Kennedy, A.D.; Eckhart, A.D.; Burrage, L.C.; Wulff, J.E.; Miller, L.A.D.; Milburn, M.V.; Ryals, J.A.; Beaudet, A.L.; Sun, Q.; et al. Untargeted metabolomic analysis for the clinical screening of inborn errors of metabolism. J. Inherit. Metab. Dis. 2015, 38, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Donti, T.R.; Cappuccio, G.; Hubert, L.; Neira, J.; Atwal, P.S.; Miller, M.J.; Cardon, A.L.; Sutton, V.R.; Porter, B.E.; Baumer, F.M.; et al. Diagnosis of adenylosuccinate lyase deficiency by metabolomic profiling in plasma reveals a phenotypic spectrum. Mol. Genet. Metab. Rep. 2016, 8, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Cappuccio, G.; Pinelli, M.; Alagia, M.; Donti, T.; Day-Salvatore, D.-L.; Veggiotti, P.; De Giorgis, V.; Lunghi, S.; Vari, M.S.; Striano, P.; et al. Biochemical phenotyping unravels novel metabolic abnormalities and potential biomarkers associated with treatment of GLUT1 deficiency with ketogenic diet. PLoS ONE 2017, 12, e0184022. [Google Scholar] [CrossRef] [PubMed]
- Weaving, L.S.; Ellaway, C.J.; Gécz, J.; Christodoulou, J. Rett syndrome: Clinical review and genetic update. J. Med. Genet. 2005, 42, 1–7. [Google Scholar] [CrossRef]
- Banerjee, A.; Castro, J.; Sur, M. Rett syndrome: Genes, synapses, circuits, and therapeutics. Front. Psychiatry 2012, 3, 34. [Google Scholar] [CrossRef]
- Amir, R.E.; Van den Veyver, I.B.; Wan, M.; Tran, C.Q.; Francke, U.; Zoghbi, H.Y. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat. Genet. 1999, 23, 185–188. [Google Scholar] [CrossRef]
- Williamson, S.L.; Christodoulou, J. Rett syndrome: New clinical and molecular insights. Eur. J. Hum. Genet. 2006, 14, 896–903. [Google Scholar] [CrossRef]
- Clarke, A.J.; Abdala Sheikh, A.P. A perspective on ‘cure’ for Rett syndrome. Orphanet J. Rare Dis. 2018, 13, 44. [Google Scholar] [CrossRef]
- Buchovecky, C.M.; Turley, S.D.; Brown, H.M.; Kyle, S.M.; McDonald, J.G.; Liu, B.; Pieper, A.A.; Huang, W.; Katz, D.M.; Russell, D.W.; et al. A suppressor screen in Mecp2 mutant mice implicates cholesterol metabolism in Rett syndrome. Nat. Genet. 2013, 45, 1013–1020. [Google Scholar] [CrossRef]
- Lane, J.B.; Lee, H.-S.; Smith, L.W.; Cheng, P.; Percy, A.K.; Glaze, D.G.; Neul, J.L.; Motil, K.J.; Barrish, J.O.; Skinner, S.A.; et al. Clinical severity and quality of life in children and adolescents with Rett syndrome. Neurology 2011, 77, 1812–1818. [Google Scholar] [CrossRef]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef] [PubMed]
- Vashi, N.; Justice, M.J. Treating Rett syndrome: From mouse models to human therapies. Mamm Genome 2019, 30, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Vogel Ciernia, A.; Yasui, D.H.; Pride, M.C.; Durbin-Johnson, B.; Noronha, A.B.; Chang, A.; Knotts, T.A.; Rutkowsky, J.R.; Ramsey, J.J.; Crawley, J.N.; et al. MeCP2 isoform e1 mutant mice recapitulate motor and metabolic phenotypes of Rett syndrome. Hum. Mol. Genet. 2018, 27, 4077–4093. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Alessio, N.; Capasso, S.; Di Bernardo, G.; Melone, M.A.B.; Peluso, G.; Galderisi, U. Senescence Phenomena and Metabolic Alteration in Mesenchymal Stromal Cells from a Mouse Model of Rett Syndrome. Int. J. Mol. Sci. 2019, 20, 2508. [Google Scholar] [CrossRef]
- Segatto, M.; Trapani, L.; Di Tunno, I.; Sticozzi, C.; Valacchi, G.; Hayek, J.; Pallottini, V. Cholesterol metabolism is altered in Rett syndrome: A study on plasma and primary cultured fibroblasts derived from patients. PLoS ONE 2014, 9, e104834. [Google Scholar] [CrossRef]
- Mouro, F.M.; Miranda-Lourenço, C.; Sebastião, A.M.; Diógenes, M.J. From Cannabinoids and Neurosteroids to Statins and the Ketogenic Diet: New Therapeutic Avenues in Rett Syndrome? Front. Neurosci. 2019, 13, 680. [Google Scholar] [CrossRef]
- Liebhaber, G.M.; Riemann, E.; Baumeister, F.A.M. Ketogenic diet in Rett syndrome. J. Child. Neurol. 2003, 18, 74–75. [Google Scholar] [CrossRef]
- Chin, E.W.M.; Marcy, G.; Yoon, S.-I.; Ma, D.; Rosales, F.J.; Augustine, G.J.; Goh, E.L.K. Choline Ameliorates Disease Phenotypes in Human iPSC Models of Rett Syndrome. NeuroMolecular Med. 2016, 18, 364–377. [Google Scholar] [CrossRef]
- Wang, H.; Liang, S.; Wang, M.; Gao, J.; Sun, C.; Wang, J.; Xia, W.; Wu, S.; Sumner, S.J.; Zhang, F.; et al. Potential serum biomarkers from a metabolomics study of autism. J. Psychiatry Neurosci. 2016, 41, 27–37. [Google Scholar] [CrossRef]
- Di Pardo, A.; Maglione, V. Sphingolipid Metabolism: A New Therapeutic Opportunity for Brain Degenerative Disorders. Front. Neurosci. 2018, 12, 249. [Google Scholar] [CrossRef]
- Deogracias, R.; Yazdani, M.; Dekkers, M.P.J.; Guy, J.; Ionescu, M.C.S.; Vogt, K.E.; Barde, Y.-A. Fingolimod, a sphingosine-1 phosphate receptor modulator, increases BDNF levels and improves symptoms of a mouse model of Rett syndrome. Proc. Natl. Acad. Sci. USA 2012, 109, 14230–14235. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Subject | Age (years) | MECP2 Variant a | Seizures | Drugs | Clinical Severity Score |
|---|---|---|---|---|---|
| 1 | 6 | c.1116_1201del86 | - | Vitamin D | 6 |
| 2 | 13 | c.455C>G (p.Pro152Arg) | + | Clonazepam, Carbamazepine, Acetazolamide, Lansoprazole, Vitamin D | 58 |
| 3 | 12 | c.455C>G (p.Pro152Arg) | + | Carbamazepine, Lansoprazole | 57 |
| 4 | 10 | c.1164_1189del26 | - | N-acetyl-cysteine | 6 |
| 5 | 16 | c.908_1143del236+1159-1170del12 | - | Carbamazepine, Insulin | 48 |
| 6 | 15 | c.763C>T (p.Arg255Ter) | + | Valproate, Lamotrigine, Melatonin | 47 |
| 7 | 7 | c.1151_1352del202 | + | Vitamin D | 28 |
| 8 | 11 | c.397C>T (p.Arg133Cys) | + | Valproate | 36 |
| 9 | 3 | c.502C>T (p.Arg168Ter) | - | Carnitine | 48 |
| 10 | 17 | c.763C>T (p.Arg255Ter) | - | Carbamazepine | 43 |
| 11 | 29 | c.808C>T (p.Arg270Ter) | - | Carnitine, Carbamazepine, Pentoxifylline, Chlorpromazine | 37 |
| 12 | 13 | c.502C>T (p.Arg168Ter) | - | Valproate | 12 |
| 13 | 21 | c.397C>T (p.Arg133Cys) | - | Valproate, Topiramate, Prazosin | 12 |
| 14 | 29 | c.473C>T (p.Thr158Met) | - | Phenobarbital | 16 |
| Super-Pathway | Compounds in Super-Pathway |
|---|---|
| Lipid | 305 *** |
| Amino Acid | 147 ** |
| Cofactor and Vitamin | 17 * |
| Xenobiotics | 71 |
| Energy | 8 |
| Peptide | 34 |
| Carbohydrate | 22 |
| Nucleotide | 28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappuccio, G.; Donti, T.; Pinelli, M.; Bernardo, P.; Bravaccio, C.; Elsea, S.H.; Brunetti-Pierri, N. Sphingolipid Metabolism Perturbations in Rett Syndrome. Metabolites 2019, 9, 221. https://doi.org/10.3390/metabo9100221
Cappuccio G, Donti T, Pinelli M, Bernardo P, Bravaccio C, Elsea SH, Brunetti-Pierri N. Sphingolipid Metabolism Perturbations in Rett Syndrome. Metabolites. 2019; 9(10):221. https://doi.org/10.3390/metabo9100221
Chicago/Turabian StyleCappuccio, Gerarda, Taraka Donti, Michele Pinelli, Pia Bernardo, Carmela Bravaccio, Sarah H. Elsea, and Nicola Brunetti-Pierri. 2019. "Sphingolipid Metabolism Perturbations in Rett Syndrome" Metabolites 9, no. 10: 221. https://doi.org/10.3390/metabo9100221
APA StyleCappuccio, G., Donti, T., Pinelli, M., Bernardo, P., Bravaccio, C., Elsea, S. H., & Brunetti-Pierri, N. (2019). Sphingolipid Metabolism Perturbations in Rett Syndrome. Metabolites, 9(10), 221. https://doi.org/10.3390/metabo9100221