Atypical Antipsychotics and the Human Skeletal Muscle Lipidome

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Patient Population
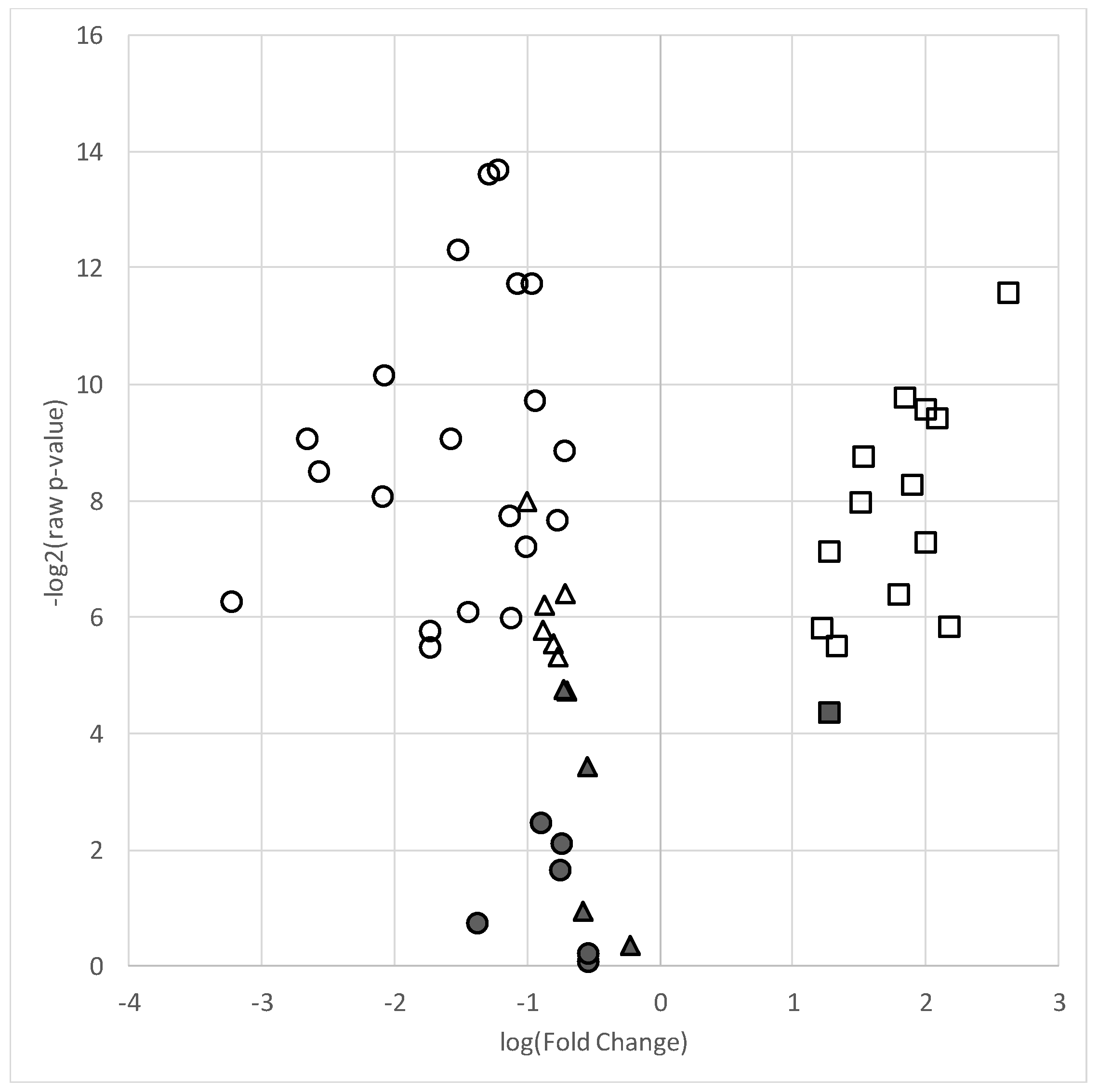
2.2. Lipidomic Analyses of Skeletal Muscle
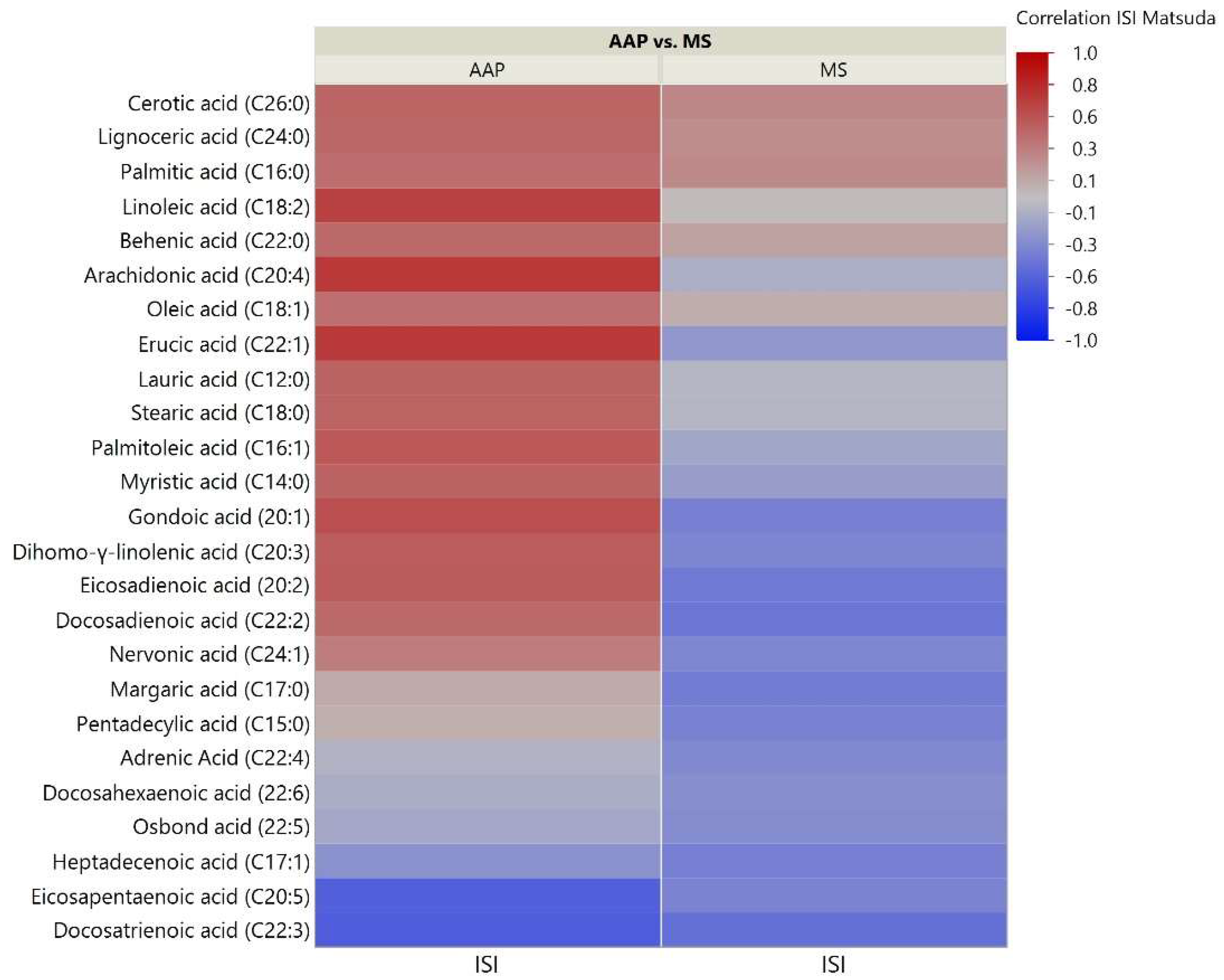
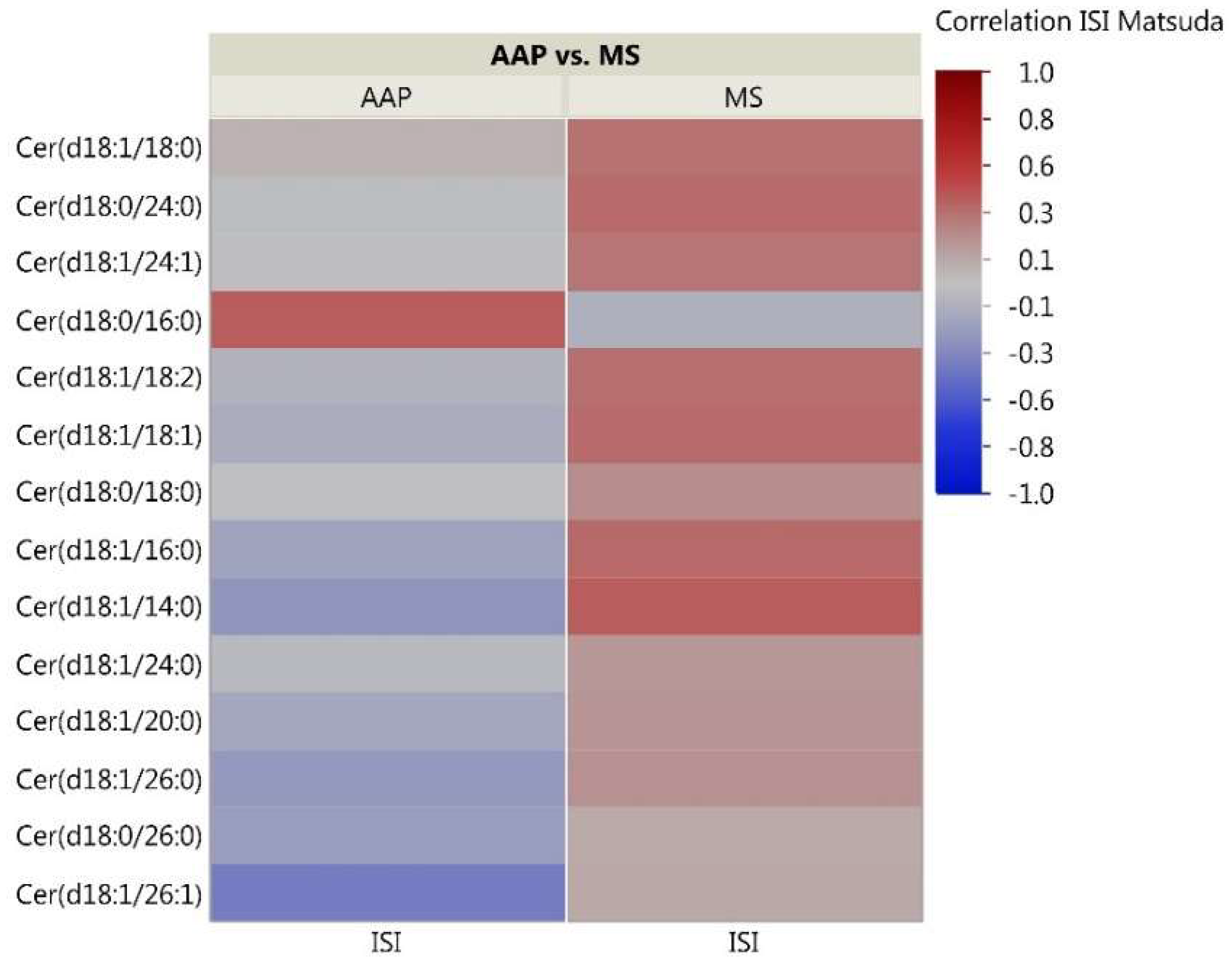
2.3. Skeletal Muscle Lipidomic Correlation to Insulin Sensitivity
3. Discussion
3.1. Atypical Antipsychotics May Cause Lipid Specific Changes in the Human Skeletal Muscle
3.2. Skeletal Muscle Lipids Correlate with AAP-Induced Insulin Resistance
3.3. Limitations
4. Materials and Methods
4.1. Subject Recruitment, Inclusion Criteria, and Clinical Assessment
4.2. Lipidomic Analysis of Biopsied Skeletal Muscle
4.2.1. Lipidomic Classes Analyzed
4.2.2. Total Fatty Acid Analysis
4.2.3. Phosphatidylcholine Analysis
4.2.4. Ceramide Analysis
4.2.5. Lipidomic Data Acquisition, Processing and Identification
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of Open Access Journals |
| TLA | three-letter acronym |
| LD | linear dichroism |
| AAP | atypical antipsychotic |
| TFA | total fatty acid |
| PC | phosphatidylcholine |
| CER | ceramide |
References
- Kessler, R.C.; Berglund, P.; Demler, O.; Jin, R.; Merikangas, K.R.; Walters, E.E. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch. Gen. Psychiatry 2005, 62, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, M.; Greene, M.; Guerin, A.; Touya, M.; Wu, E. The economic burden of bipolar I disorder in the United States in 2015. J. Affect. Disord. 2018, 226, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.E. Social and economic burden of mood disorders. Biol. Psychiat. 2003, 54, 208–215. [Google Scholar] [CrossRef]
- Ferrari, A.J.; Stockings, E.; Khoo, J.P.; Erskine, H.E.; Degenhardt, L.; Vos, T.; Whiteford, H.A. The prevalence and burden of bipolar disorder: findings from the Global Burden of Disease Study 2013. Bipolar Disord. 2016, 18, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Keck, P.E., Jr.; McElroy, S.L.; Strakowski, S.M.; Stanton, S.P.; Kizer, D.L.; Balistreri, T.M.; Bennett, J.A.; Tugrul, K.C.; West, S.A. Factors associated with pharmacologic noncompliance in patients with mania. J. Clin. Psychiatry 1996, 57, 292–297. [Google Scholar] [PubMed]
- Miasso, A.I.; do Carmo, B.P.; Tirapelli, C.R. Bipolar affective disorder: pharmacotherapeutic profile and adherence to medication. Rev. Esc. Enferm USP 2012, 46, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Montes, J.M.; Maurino, J.; de Dios, C.; Medina, E. Suboptimal treatment adherence in bipolar disorder: impact on clinical outcomes and functioning. Patient Prefer. Adher. 2013, 7, 89–94. [Google Scholar] [CrossRef]
- Haddad, P.M.; Brain, C.; Scott, J. Nonadherence with antipsychotic medication in schizophrenia: Challenges and management strategies. Patient Relat. Outcome Meas. 2014, 5, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Haddad, P.M.; Sharma, S.G. Adverse effects of atypical antipsychotics: Differential risk and clinical implications. CNS Drugs 2007, 21, 911–936. [Google Scholar] [CrossRef] [PubMed]
- Leucht, S.; Cipriani, A.; Spineli, L.; Mavridis, D.; Orey, D.; Richter, F.; Samara, M.; Barbui, C.; Engel, R.R.; Geddes, J.R.; et al. Comparative efficacy and tolerability of 15 antipsychotic drugs in schizophrenia: A multiple-treatments meta-analysis. Lancet 2013, 382, 951–962. [Google Scholar] [CrossRef]
- Correll, C.U.; Detraux, J.; De Lepeleire, J.; De Hert, M. Effects of antipsychotics, antidepressants and mood stabilizers on risk for physical diseases in people with schizophrenia, depression and bipolar disorder. World Psychiatry 2015, 14, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, H.A. Atypical antipsychotic-induced metabolic side effects: Insights from receptor-binding profiles. Mol. Psychiatry 2008, 13, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ballon, J.S.; Pajvani, U.; Freyberg, Z.; Leibel, R.L.; Lieberman, J.A. Molecular pathophysiology of metabolic effects of antipsychotic medications. Trends Endocrinol. Metab. 2014, 25, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Boyda, H.N.; Tse, L.; Procyshyn, R.M.; Honer, W.G.; Barr, A.M. Preclinical models of antipsychotic drug-induced metabolic side effects. Trends Pharmacol. Sci. 2010, 31, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Lett, T.A.; Wallace, T.J.; Chowdhury, N.I.; Tiwari, A.K.; Kennedy, J.L.; Muller, D.J. Pharmacogenetics of antipsychotic-induced weight gain: Review and clinical implications. Mol. Psychiatry 2012, 17, 242–266. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, K.J.; Evans, S.J.; Wiese, K.M.; Ellingrod, V.L. An Untargeted Metabolomics Analysis of Antipsychotic Use in Bipolar Disorder. Clin. Transl. Sci. 2015, 5, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, K.J.; Goodrich, J.M.; Dolinoy, D.C.; Ellingrod, V.L. Gene-specific DNA methylation may mediate atypical antipsychotic-induced insulin resistance. Bipolar Disord. 2016, 18, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; McEvoy, J.; Baillie, R.; Lee, D.; Yao, J.; Doraiswamy, P.; Krishnan, K. Metabolomic mapping of atypical antipsychotic effects in schizophrenia. Mol. Psychiatry 2007, 12, 934–945. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, J.; Baillie, R.A.; Zhu, H.; Buckley, P.; Keshavan, M.S.; Nasrallah, H.A.; Dougherty, G.G.; Yao, J.K.; Kaddurah-Daouk, R. Lipidomics reveals early metabolic changes in subjects with schizophrenia: Effects of atypical antipsychotics. PLoS ONE 2013, 8, e68717. [Google Scholar] [CrossRef] [PubMed]
- Jaros, J.A.; Rahmoune, H.; Wesseling, H.; Leweke, F.M.; Ozcan, S.; Guest, P.C.; Bahn, S. Effects of olanzapine on serum protein phosphorylation patterns in patients with schizophrenia. Proteom. Clin. Appl. 2015, 9, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.A.; George, R.C.; Danielson, P.E.; Nelson, P.A.; Warren, A.J.; Lo, D.; Sutcliffe, J.G. Antipsychotic drug treatment alters expression of mRNAs encoding lipid metabolism-related proteins. Mol. Psychiatry 2003, 8, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Pillinger, T.; D’Ambrosio, E.; McCutcheon, R.; O, D.H. Is psychosis a multisystem disorder? A meta-review of central nervous system, immune, cardiometabolic, and endocrine alterations in first-episode psychosis and perspective on potential models. Mol. Psychiatry 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Tripathy, D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2009, 32 (Suppl. 2), 157–163. [Google Scholar] [CrossRef]
- Abdul-Ghani, M.A.; DeFronzo, R.A. Pathogenesis of insulin resistance in skeletal muscle. J. Biomed. Biotechnol. 2010, 2010, 476279. [Google Scholar] [CrossRef] [PubMed]
- Engl, J.; Laimer, M.; Niederwanger, A.; Kranebitter, M.; Starzinger, M.; Pedrini, M.T.; Fleischhacker, W.W.; Patsch, J.R.; Ebenbichler, C.F. Olanzapine impairs glycogen synthesis and insulin signaling in L6 skeletal muscle cells. Mol. Psychiatry 2005, 10, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, K.J.; Seyoum, B.; Dass, S.; Sanders, E.; Mallisho, A.; Yi, Z. Association of Protein Kinase B (AKT) DNA Hypermethylation with Maintenance Atypical Antipsychotic Treatment in Patients with Bipolar Disorder. Pharmacotherapy 2018. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; McEvoy, J.; Baillie, R.; Zhu, H.; Yao, J.K.; Nimgaonkar, V.L.; Buckley, P.F.; Keshavan, M.S.; Georgiades, A.; Nasrallah, H.A. Impaired plasmalogens in patients with schizophrenia. Psychiatry Res. 2012, 198, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Oresic, M.; Tang, J.; Seppanen-Laakso, T.; Mattila, I.; Saarni, S.E.; Saarni, S.I.; Lonnqvist, J.; Sysi-Aho, M.; Hyotylainen, T.; Perala, J.; et al. Metabolome in schizophrenia and other psychotic disorders: A general population-based study. Genome Med. 2011, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Oresic, M.; Seppanen-Laakso, T.; Sun, D.; Tang, J.; Therman, S.; Viehman, R.; Mustonen, U.; van Erp, T.G.; Hyotylainen, T.; Thompson, P.; et al. Phospholipids and insulin resistance in psychosis: A lipidomics study of twin pairs discordant for schizophrenia. Genome Med. 2012, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Aquino, A.; Alexandrino, G.L.; Guest, P.C.; Augusto, F.; Gomes, A.F.; Murgu, M.; Steiner, J.; Martins-de-Souza, D. Blood-Based Lipidomics Approach to Evaluate Biomarkers Associated with Response to Olanzapine, Risperidone, and Quetiapine Treatment in Schizophrenia Patients. Front. Psychiatry 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.M.; Yeomans, L.; McHugh, C.; Kraal, A.Z.; Flowers, S.A.; Rothberg, A.E.; Karnovsky, A.; Das, A.; Ellingrod, V.L.; Stringer, K.A. Atypical Antipsychotic Exposure May Not Differentiate Metabolic Phenotypes of Patients with Schizophrenia. Pharmacotherapy 2018, 6, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.; Prabakaran, S.; Whitfield, P.; Major, H.; Leweke, F.M.; Koethe, D.; McKenna, P.; Bahn, S. High throughput lipidomic profiling of schizophrenia and bipolar disorder brain tissue reveals alterations of free fatty acids, phosphatidylcholines, and ceramides. J. Proteome Res. 2008, 7, 4266–4277. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Xu, W.; Hong, Q.; Xiao, C.; Yang, L.; Ma, Z.; Wang, Y.; Tan, H.; Tang, X.; Gao, Y. Rapid comparison of metabolites in humans and rats of different sexes using untargeted UPLC-TOFMS and an in-house software platform. Eur. J. Mass Spectrom. 2015, 21, 801–821. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Ishikawa, M.; Murayama, M.; Urata, M.; Senoo, Y.; Toyoshima, K.; Kumagai, Y.; Maekawa, K.; Saito, Y. Effects of sex, age, and fasting conditions on plasma lipidomic profiles of fasted Sprague-Dawley rats. PLoS ONE 2014, 9, e112266. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Maekawa, K.; Kinchen, J.M.; Tanaka, R.; Kumagai, Y.; Saito, Y. Gender- and Age-Associated Differences in Serum Metabolite Profiles among Japanese Populations. Biol. Pharm. Bull. 2016, 39, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Vassalle, C.; Sciarrino, R.; Bianchi, S.; Battaglia, D.; Mercuri, A.; Maffei, S. Sex-related differences in association of oxidative stress status with coronary artery disease. Fertil. Steril. 2012, 97, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Lundsgaard, A.M.; Kiens, B. Gender differences in skeletal muscle substrate metabolism-molecular mechanisms and insulin sensitivity. Front. Endocrinol. 2014, 5, 195. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M.G.; Agellon, L.B. Sex differences in lipid metabolism and metabolic disease risk. Biochem. Cell Biol. 2012, 90, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; Jandacek, R.; Rider, T.; Tso, P.; Cole-Strauss, A.; Lipton, J.W. Differential effects of antipsychotic medications on polyunsaturated fatty acid biosynthesis in rats: Relationship with liver delta6-desaturase expression. Schizophr. Res. 2011, 129, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Yang, x.; Sun, L.; Zhao, A.; Hu, x.; Qing, Y.; Jiang, J.; Yang, C.; Xu, T.; Wang, P.; Liu, J.; et al. Serum fatty acid patterns in patients with schizophrenia: A targeted metabonomics study. Transl. Psychiatry 2017, 7, e1176. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, V.L.; Vary, T.C.; Ilkayeva, O.; Wenner, B.R.; Maresca, K.P.; Joyal, J.L.; Breazeale, S.; Elich, T.D.; Lang, C.H.; Lynch, C.J. Atypical antipsychotics rapidly and inappropriately switch peripheral fuel utilization to lipids, impairing metabolic flexibility in rodents. Schizophr. Bull. 2012, 38, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, V.L.; Singareddy, R.; Mauger, D.; Lynch, C.J. A double blind, placebo-controlled, randomized crossover study of the acute metabolic effects of olanzapine in healthy volunteers. PLoS ONE 2011, 6, e22662. [Google Scholar] [CrossRef] [PubMed]
- Vidarsdottir, S.; de Leeuw van Weenen, J.E.; Frolich, M.; Roelfsema, F.; Romijn, J.A.; Pijl, H. Effects of olanzapine and haloperidol on the metabolic status of healthy men. J Clin. Endocrinol. Metab. 2010, 95, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, K.J.; Seyoum, B.; Mallisho, A.; Burghardt, P.R.; Kowluru, R.A.; Yi, Z. Atypical antipsychotics, insulin resistance and weight; A meta-analysis of healthy volunteer studies. Prog. Neuropsychopharmacol. Biol. Psychiat. 2018, 83, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Maras, A.; Petroianu, G.; Braus, D.F.; Scheuer, L.; Gattaz, W.F. Effects of antipsychotic treatment on membrane phospholipid metabolism in schizophrenia. J. Neural. Transm. 2001, 108, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.; Thomas, E.A. Sphingolipid abnormalities in psychiatric disorders: A missing link in pathology? Front. Biosci. 2011, 16, 1797–1810. [Google Scholar] [CrossRef]
- Castillo, R.I.; Rojo, L.E.; Henriquez-Henriquez, M.; Silva, H.; Maturana, A.; Villar, M.J.; Fuentes, M.; Gaspar, P.A. From Molecules to the Clinic: Linking Schizophrenia and Metabolic Syndrome through Sphingolipids Metabolism. Front. Neurosci. 2016, 10, 488. [Google Scholar] [CrossRef] [PubMed]
- Smesny, S.; Schmelzer, C.E.H.; Hinder, A.; Köhler, A.; Schneider, C.; Rudzok, M.; Schmidt, U.; Milleit, B.; Milleit, C.; Nenadic, I.; et al. Skin Ceramide Alterations in First-Episode Schizophrenia Indicate Abnormal Sphingolipid Metabolism. Schizophr. Bull. 2013, 39, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.; Head, S.R.; Gilmartin, T.J.; Dean, B.; Thomas, E.A. Evidence for Disruption of Sphingolipid Metabolism in Schizophrenia. J. Neurosci. Res. 2009, 87, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Suvitaival, T.; Mantere, O.; Kieseppa, T.; Mattila, I.; Poho, P.; Hyotylainen, T.; Suvisaari, J.; Oresic, M. Serum metabolite profile associates with the development of metabolic co-morbidities in first-episode psychosis. Transl. Psychiatry 2016, 6, e951. [Google Scholar] [CrossRef] [PubMed]
- Weston-Green, K.; Babic, I.; de Santis, M.; Pan, B.; Montgomery, M.K.; Mitchell, T.; Huang, x.F.; Nealon, J. Disrupted sphingolipid metabolism following acute clozapine and olanzapine administration. J. Biomed. Sci. 2018, 25, 40. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.A. Ceramides in insulin resistance and lipotoxicity. Prog. Lipid Res. 2006, 45, 42–72. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, M.P.; Lamon-Fava, S.; Fielding, R.A. Skeletal muscle lipid deposition and insulin resistance: Effect of dietary fatty acids and exercise. Am. J. Clin. Nutr. 2007, 85, 662–677. [Google Scholar] [CrossRef] [PubMed]
- Kitessa, S.M.; Abeywardena, M.Y. Lipid-Induced Insulin Resistance in Skeletal Muscle: The Chase for the Culprit Goes from Total Intramuscular Fat to Lipid Intermediates, and Finally to Species of Lipid Intermediates. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Funai, K.; Lodhi, I.J.; Spears, L.D.; Yin, L.; Song, H.; Klein, S.; Semenkovich, C.F. Skeletal Muscle Phospholipid Metabolism Regulates Insulin Sensitivity and Contractile Function. Diabetes 2016, 65, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Pinnamaneni, S.K.; Eo, S.J.; Cho, I.H.; Pyo, J.H.; Kim, C.K.; Sinclair, A.J.; Febbraio, M.A.; Watt, M.J. Saturated, but not n-6 polyunsaturated, fatty acids induce insulin resistance: Role of intramuscular accumulation of lipid metabolites. J. Appl. Physiol. 2006, 100, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Tonks, K.T.; Coster, A.C.; Christopher, M.J.; Chaudhuri, R.; Xu, A.; Gagnon-Bartsch, J.; Chisholm, D.J.; James, D.E.; Meikle, P.J.; Greenfield, J.R.; et al. Skeletal muscle and plasma lipidomic signatures of insulin resistance and overweight/obesity in humans. Obesity (Silver Spring, Md.) 2016, 24, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Broskey, N.T.; Obanda, D.N.; Burton, J.H.; Cefalu, W.T.; Ravussin, E. Skeletal muscle ceramides and daily fat oxidation in obesity and diabetes. Metab. Clin. Exp. 2018, 82, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H. Lipotoxicity in the pathogenesis of obesity-dependent NIDDM. Genetic and clinical implications. Diabetes 1995, 44, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Amati, F. Revisiting the diacylglycerol-induced insulin resistance hypothesis. Obes. Rev. 2012, 13 (Suppl. 2), 40–50. [Google Scholar] [CrossRef]
- Erion, D.M.; Shulman, G.I. Diacylglycerol-mediated insulin resistance. Nat. Med. 2010, 16, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Clore, J.N.; Li, J.; Gill, R.; Gupta, S.; Spencer, R.; Azzam, A.; Zuelzer, W.; Rizzo, W.B.; Blackard, W.G. Skeletal muscle phosphatidylcholine fatty acids and insulin sensitivity in normal humans. Am. J. Physiol. Renal. Physiol. 1998, 275, E665–670. [Google Scholar] [CrossRef]
- Timmers, S.; Schrauwen, P.; de Vogel, J. Muscular diacylglycerol metabolism and insulin resistance. Physiol. Behav. 2008, 94, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.; Fransen, A.; Janssen, J.; van Os, J.; Drukker, M. Almost all antipsychotics result in weight gain: A meta-analysis. PLoS ONE 2014, 9, e94112. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, A.M.; Gonzalez-Blanco, L.; Garcia-Rizo, C.; Fernandez-Egea, E.; Miller, B.; Arroyo, M.B.; Kirkpatrick, B. Meta-analysis of glucose tolerance, insulin, and insulin resistance in antipsychotic-naive patients with nonaffective psychosis. Schizophr. Res. 2017, 179, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Pillinger, T.; Beck, K.; Gobjila, C.; Donocik, J.G.; Jauhar, S.; Howes, O.D. Impaired Glucose Homeostasis in First-Episode Schizophrenia: A Systematic Review and Meta-analysis. JAMA Psychiatry 2017, 74, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Spertus, J.; Horvitz-Lennon, M.; Abing, H.; Normand, S.L. Risk of weight gain for specific antipsychotic drugs: A meta-analysis. NPJ Schizophr. 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiat. 1998, 59, 22–33. [Google Scholar]
- Shanely, R.A.; Zwetsloot, K.A.; Triplett, N.T.; Meaney, M.P.; Farris, G.E.; Nieman, D.C. Human skeletal muscle biopsy procedures using the modified Bergstrom technique. J. Vis. Exp. 2014, 51812. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.A.; Heckert, A.; Ulmer, C.Z.; Jones, C.M.; Koelmel, J.P.; Abdullah, L.; Ahonen, L.; Alnouti, Y.; Armando, A.M.; Asara, J.M.; et al. Harmonizing lipidomics: NIST interlaboratory comparison exercise for lipidomics using SRM 1950-Metabolites in Frozen Human Plasma. J. Lipid. Res. 2017, 58, 2275–2288. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid. Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.T.; Maddipati, K.R.; Taatjes, D.J.; Jena, B.P. Neuronal porosome lipidome. J. Cell. Mol. Med. 2014, 18, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Volden, P.A. Mammary Adipose Tissue-derived Lysophospholipids Promote Estrogen Receptor-negative Mammary Epithelial Cell Proliferation. Cancer Prev. Res. (Phila). 2016, 9, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Hellmuth, C.; Weber, M.; Koletzko, B.; Peissner, W. Nonesterified Fatty Acid Determination for Functional Lipidomics: Comprehensive Ultrahigh Performance Liquid Chromatography–Tandem Mass Spectrometry Quantitation, Qualification, and Parameter Prediction. Anal. Chem. 2012, 84, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Fauland, A.; Trötzmüller, M.; Eberl, A.; Afiuni-Zadeh, S.; Köfeler, H.; Guo, x.; Lankmayr, E. An improved SPE method for fractionation and identification of phospholipids. J. Sep. Sci. 2013, 36, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Ejsing, C.S.; Duchoslav, E.; Sampaio, J.; Simons, K.; Bonner, R.; Thiele, C.; Ekroos, K.; Shevchenko, A. Automated identification and quantification of glycerophospholipid molecular species by multiple precursor ion scanning. Anal. Chem. 2006, 78, 6202–6214. [Google Scholar] [CrossRef] [PubMed]
- Bielawski, J.; Szulc, Z.M.; Hannun, Y.A.; Bielawska, A. Simultaneous quantitative analysis of bioactive sphingolipids by high-performance liquid chromatography-tandem mass spectrometry. Methods 2006, 39, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 289–300. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atypical Antipsychotic (n = 15) | Mood Stabilizer (n = 15) | |
|---|---|---|
| Age (years) | 43.0 ± 14.9 | 45.3 ± 13.0 |
| Sex (% female) | 61 | 57 |
| Race (% Caucasian/% African-American) | 57/34 | 71/28 |
| WHR | 1.00 ± 0.10 | 1.02 ± 0.04 |
| BMI (mg/kg2) | 32.0 ± 8.9 | 33.5 ± 6.3 |
| Insulin Sensitivity Index * | 5.20 ± 3.8 | 12.4 ± 6.90 |
| Cholesterol (mg/dL) | 173 ± 40.9 | 178 ± 38.9 |
| LDL (mg/dL) | 96.1 ± 34.1 | 103 ± 41.3 |
| HDL (mg/dL) | 57.4 ± 23.6 | 47.2 ± 15.8 |
| Triglycerides (mg/dL) | 95.3 ± 46.7 | 107 ± 53.8 |
| Antipsychotic Type (%quetiapine/%risperidone/ %olanzapine/%lurasidone) | 50/ 25/ 12.5/ 12.5 | NA |
| Mood Stabilizer (%lamotrigine/%lithium/%valproic acid). | NA | 40/33/27 |
| Lipid | Raw Fold Change a | Log2 Fold Change a | Raw p-Value | FDR q-Value |
|---|---|---|---|---|
| Total Fatty Acids | ||||
| Lauric acid (C12:0) | 0.41 | −1.28 | 8.19 × 10−5 | 1.07 × 10−3 * |
| Myristic acid (C14:0) | 0.59 | −0.76 | 5.74 × 10−3 | 9.29 × 10−3 * |
| Pentadecylic acid (C15:0) | 0.61 | −0.70 | 2.24 × 10−3 | 5.72 × 10−3 * |
| Palmitic acid (C16:0) | 0.50 | −1.0 | 6.9 × 10−3 | 0.01 * |
| Palmitoleic acid (C16:1) | 0.30 | −1.72 | 0.02 | 0.03 * |
| Margaric acid (C17:0) | 0.52 | −0.96 | 3.32 × 10−4 | 1.56 × 10−3 * |
| Heptadecenoic acid (C17:1) | 0.54 | −0.89 | 0.18 | 0.22 |
| Stearic acid (C18:0) | 0.53 | −0.92 | 1.24 × 10−3 | 4.5 × 10−3 * |
| Oleic acid (C18:1) | 0.30 | −1.7 | 0.02 | 0.03 * |
| Linoleic acid (C18:2) | 0.37 | −1.44 | 0.02 | 0.02 * |
| Arachidic acid (C20:0) | 0.43 | −1.21 | 7.66 × 19−5 | 1.07 × 10−3 * |
| Gondoic acid (C20:1) | 0.10 | −3.22 | 0.01 | 0.02 * |
| Eicosadienoic acid (C20:2) | 0.17 | −2.56 | 2.83 × 10−3 | 6.62 × 10−3 * |
| Dihomo-ү-linolenic acid (C20:3) | 0.24 | −2.08 | 3.88 × 10−3 | 8.23 × 10−3 * |
| Arachidonic acid (C20:4) | 0.34 | −1.56 | 1.93 × 10−3 | 5.49 × 10−3 * |
| Eicosapentaenoic acid (C20:5) | 0.69 | −0.53 | 0.88 | 0.92 |
| Behenic acid (C22:0) | 0.48 | −1.06 | 3.72 × 10−4 | 1.56 × 10−3 * |
| Erucic acid (C22:1) | 0.24 | −2.07 | 9.01 × 10−4 | 3.92 × 10−3 * |
| Docosadienoic acid (C22:2) | 0.16 | −2.64 | 1.94 × 10−3 | 5.49 × 10−3 * |
| Docosatrienoic acid (C22:3) | 0.69 | −0.53 | 0.96 | 0.96 |
| Adrenic Acid (C22:4) | 0.60 | −0.73 | 0.23 | 0.28 |
| Osbond acid (C22:5) | 0.60 | −0.73 | 0.32 | 0.36 |
| Docosahexaenoic acid (C22:6) | 0.39 | −1.37 | 0.60 | 0.65 |
| Lignoceric acid (C24:0) | 0.46 | −1.12 | 4.86 × 10−3 | 9.29 × 10−3 * |
| Nervonic acid (C24:1) | 0.35 | −1.51 | 2.98 × 10−4 | 1.56 × 10−3 * |
| Cerotic acid (C26:0) | 0.46 | −1.12 | 0.02 | 0.02* |
| Phosphatidylcholines | ||||
| PC 32:0 | 0.55 | −0.86 | 0.01 | 0.04 * |
| PC 32:1 | 0.69 | −0.54 | 0.09 | 0.12 |
| PC 34:1 | 0.54 | −0.88 | 0.02 | 0.04 * |
| PC 34:2 | 0.62 | −0.70 | 0.04 | 0.05 |
| PC 34:3 | 0.50 | −1.0 | 4.01 × 10−3 | 0.04 * |
| PC 36:1 | 0.61 | −0.71 | 0.01 | 0.04 * |
| PC 36:2 | 0.67 | −0.57 | 0.53 | 0.58 |
| PC 36:3 | 0.59 | −0.76 | 0.03 | 0.04 * |
| PC 36:4 | 0.58 | −0.80 | 0.02 | 0.04 * |
| PC 38:4 | 0.61 | −0.72 | 0.04 | 0.05 |
| PC 38:5 | 0.86 | −0.22 | 0.80 | 0.80 |
| Ceramides | ||||
| CER(d18:1/14:0) | 4.00 | 2.00 | 1.34 × 10−3 | 5.21 × 10−3 * |
| CER(d18:1/16:0) | 4.29 | 2.10 | 1.49 × 10−3 | 5.21 × 10−3 * |
| CER(d18:0/16:0) | 6.21 | 2.63 | 3.32 × 10−4 | 4.65 × 10−3 * |
| CER(d18:1/18:0) | 3.50 | 1.81 | 0.01 | 0.01 * |
| CER(d18:0/18:0) | 4.55 | 2.19 | 0.02 | 0.02 * |
| CER(d18:1/18:1) | 3.61 | 1.85 | 1.17 × 10−3 | 5.21 × 10−3 * |
| CER(d18:1/18:2) | 2.91 | 1.54 | 2.34 × 10−3 | 6.56 × 10−3 * |
| CER(d18:1/20:0) | 4.03 | 2.01 | 6.51 × 10−3 | 0.01 * |
| CER(d18:1/24:0) | 2.54 | 1.34 | 0.02 | 0.02 * |
| CER(d18:0/24:0) | 2.34 | 1.23 | 0.02 | 0.02 * |
| CER(d18:1/24:1) | 3.76 | 1.91 | 3.29 × 10−3 | 7.68 × 10−3 * |
| CER(d18:1/26:0) | 2.43 | 1.28 | 7.26 × 10−3 | 0.01 * |
| CER(d18:0/26:0) | 2.86 | 1.52 | 4.07 × 10−3 | 8.15 × 10−3 * |
| CER(d18:1/26:1) | 2.44 | 1.29 | 0.05 | 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burghardt, K.J.; Ward, K.M.; Sanders, E.J.; Howlett, B.H.; Seyoum, B.; Yi, Z. Atypical Antipsychotics and the Human Skeletal Muscle Lipidome. Metabolites 2018, 8, 64. https://doi.org/10.3390/metabo8040064
Burghardt KJ, Ward KM, Sanders EJ, Howlett BH, Seyoum B, Yi Z. Atypical Antipsychotics and the Human Skeletal Muscle Lipidome. Metabolites. 2018; 8(4):64. https://doi.org/10.3390/metabo8040064
Chicago/Turabian StyleBurghardt, Kyle J., Kristen M. Ward, Elani J. Sanders, Bradley H. Howlett, Berhane Seyoum, and Zhengping Yi. 2018. "Atypical Antipsychotics and the Human Skeletal Muscle Lipidome" Metabolites 8, no. 4: 64. https://doi.org/10.3390/metabo8040064
APA StyleBurghardt, K. J., Ward, K. M., Sanders, E. J., Howlett, B. H., Seyoum, B., & Yi, Z. (2018). Atypical Antipsychotics and the Human Skeletal Muscle Lipidome. Metabolites, 8(4), 64. https://doi.org/10.3390/metabo8040064