

In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of Bioactive Metabolites Extracted from Kiwi and Its By-Products

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Bioactive Compounds from Juice and By-Products
2.3. Platelet Aggregometry Biological Assays
2.4. FT-IR Based Structural Analysis of TAC Extracts from Lemna Minor and Nelumbo Nucifera
2.5. Fatty Acid Composition and Polar Lipids’ Structural Analysis by Liquid Chromatography-Mass Spectrometry (LC-MS)
2.6. Assessment of Total Phenolic Content, Carotenoid Content, and Antioxidant Activities
2.6.1. Sample Preparation for Further Analysis
2.6.2. Total Phenolic Content (TPC) Analysis
2.6.3. Total Antioxidant Activity (TAA) Evaluation
2.6.4. Total Carotenoid Content (TCC) Analysis
2.6.5. Statistical Analysis
3. Results
3.1. Yield of Extraction
3.2. Total Phenolic and Carotenoid Content of Kiwi and Its By-Products
3.3. Antioxidant Activities
3.4. FT-IR
3.5. LC-MSAnalysis of the TAC Extracts from Kiwi Juice and Its By-Products—Fatty Acid Composition and Structural Elucidation of Polar Lipids
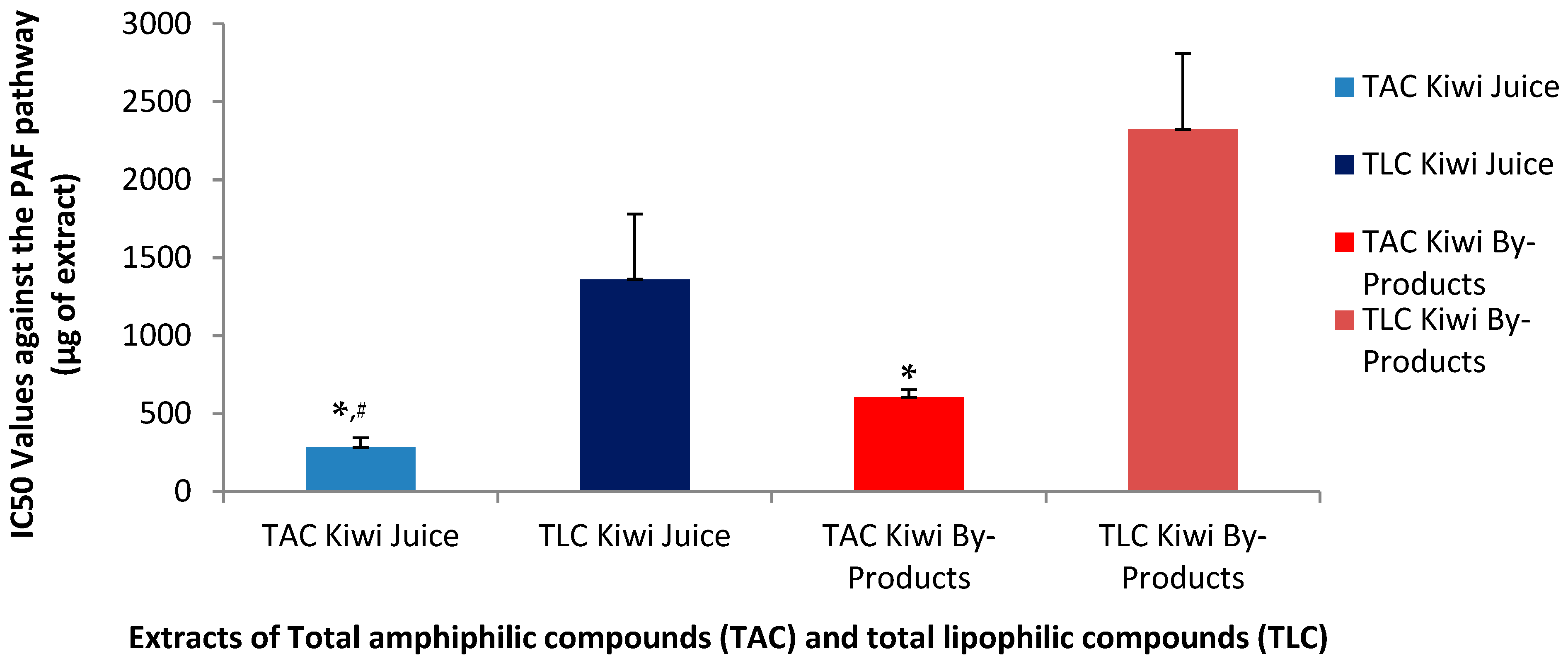
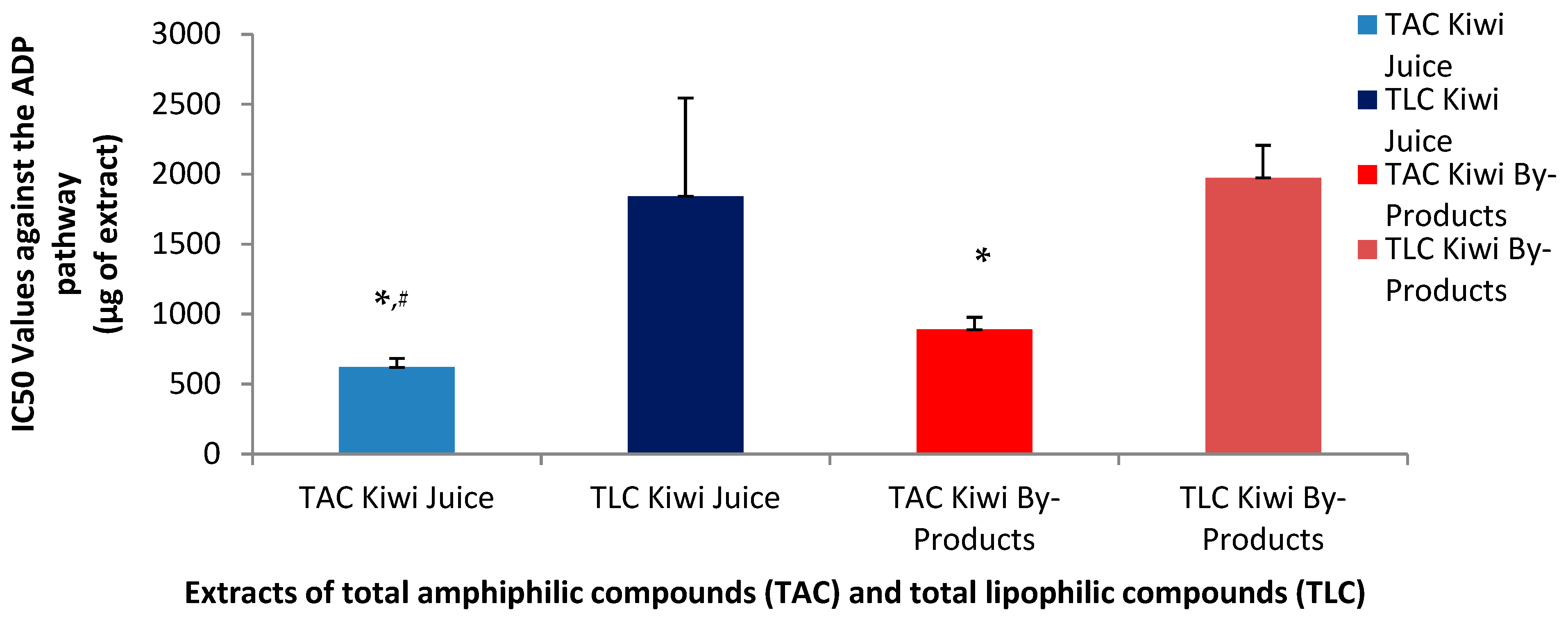
3.6. Anti-Inflammatory and Anti-Platelet Properties of Kiwi
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moysidou, A.M.; Cheimpeloglou, K.; Koutra, S.I.; Finos, M.A.; Ofrydopoulou, A.; Tsoupras, A. A Comprehensive Review on the Antioxidant and Anti-Inflammatory Bioactives of Kiwi and Its By-Products for Functional Foods and Cosmetics with Health-Promoting Properties. Appl. Sci. 2024, 14, 5990. [Google Scholar] [CrossRef]
- Cassano, A.; Donato, L.; Conidi, C.; Drioli, E. Recovery of bioactive compounds in kiwifruit juice by ultrafiltration. Innov. Food Sci. Emerg. Technol. 2008, 9, 556–562. [Google Scholar] [CrossRef]
- Khiralla, G.; Ali, H.M. Bioavailability and antioxidant potentials of fresh and pasteurized kiwi juice before and after in vitro gastrointestinal digestion. J. Food Sci. Technol. 2020, 57, 4277–4285. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, F.; Carpena, M.; Fraga-Corral, M.; Echave, J.; Riaz Rajoka, M.S.; Barba, F.J.; Cao, H.; Xiao, J.; Prieto, M.A.; Simal-Gandara, J. Valorization of kiwi agricultural waste and industry by-products by recovering bioactive compounds and applications as food additives: A circular economy model. Food Chem. 2022, 370, 131315. [Google Scholar] [CrossRef]
- Chamorro, F.; Carpena, M.; Nuñez-Estevez, B.; Prieto, M.A.; Simal-Gandara, J. Valorization of Kiwi by-Products for the Recovery of Bioactive Compounds: Circular Economy Model. Proceedings 2020, 70, 9. [Google Scholar] [CrossRef]
- Sanz, V.; López-Hortas, L.; Torres, M.D.; Domínguez, H. Trends in kiwifruit and byproducts valorization. Trends Food Sci. Technol. 2020, 107, 401–414. [Google Scholar] [CrossRef]
- Dias, M.; Caleja, C.; Pereira, C.; Calhelha, R.C.; Kostic, M.; Sokovic, M.; Tavares, D.; Baraldi, I.J.; Barros, L.; Ferreira, I.C.F.R. Chemical composition and bioactive properties of byproducts from two different kiwi varieties. Food Res. Int. 2020, 127, 108753. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Vandorou, M.; Plakidis, C.; Tsompanidou, I.M.; Ofrydopoulou, A.; Shiels, K.; Saha, S.K.; Tsoupras, A. In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Properties of the Amphiphilic Bioactives from Greek Organic Starking Apple Juice and Its By-Products (Apple Pomace). Appl. Sci. 2025, 15, 2807. [Google Scholar] [CrossRef]
- Galanos, D.S.; Kapoulas, V.M. Isolation of polar lipids from triglyceride mixtures. J. Lipid Res. 1962, 3, 134–136. [Google Scholar] [CrossRef]
- Tsoupras, A.; Cholidis, P.; Kranas, D.; Galouni, E.A.; Ofrydopoulou, A.; Efthymiopoulos, P.; Shiels, K.; Saha, S.K.; Kyzas, G.Z.; Anastasiadou, C. Anti-Inflammatory, Antithrombotic, and Antioxidant Properties of Amphiphilic Lipid Bioactives from Shrimp. Pharmaceuticals 2025, 18, 25. [Google Scholar] [CrossRef] [PubMed]
- Vordos, N.; Giannakopoulos, S.; Vansant, E.F.; Kalaitzis, C.; Nolan, J.W.; Bandekas, D.V.; Karavasilis, I.; Mitropoulos, A.C.; Touloupidis, S. Small-angle X-ray scattering (SAXS) and nitrogen porosimetry (NP): Two novel techniques for the evaluation of urinary stone hardness. Int. Urol. Nephrol. 2018, 50, 1779–1785. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, D.; Chrysikopoulou, V.; Rampaouni, A.; Plakidis, C.; Ofrydopoulou, A.; Shiels, K.; Saha, S.K.; Tsoupras, A. Antioxidant, Antithrombotic and Anti-Inflammatory Properties of Amphiphilic Bioactives from Water Kefir Grains and Its Apple Pomace-Based Fermented Beverage. Antioxidants 2025, 14, 164. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Manousakis, V.; Koutis, N.; Zervas, G.P.; Ofrydopoulou, A.; Shiels, K.; Saha, S.K.; Tsoupras, A. In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of the Amphiphilic Bioactives Extracted from Avocado and Its By-Products. Antioxidants 2025, 14, 146. [Google Scholar] [CrossRef]
- McGhie, T.K.; Ainge, G.D. Color in fruit of the genus actinidia: Carotenoid and chlorophyll compositions. J. Agric. Food Chem. 2002, 50, 117–121. [Google Scholar] [CrossRef]
- Guthrie, F.; Wang, Y.; Neeve, N.; Quek, S.Y.; Mohammadi, K.; Baroutian, S. Recovery of Phenolic Antioxidants from Green Kiwifruit peel using Subcritical Water Extraction. Food Bioprod. Process. 2020, 122, 136–144. [Google Scholar] [CrossRef]
- Dawes, H.M.; Keene, J.B. Phenolic composition of kiwifruit juice. J. Agric. Food Chem. 1999, 47, 2398–2403. [Google Scholar] [CrossRef]
- Carbone, K.; Amoriello, T.; Iadecola, R. Exploitation of Kiwi Juice Pomace for the Recovery of Natural Antioxidants through Microwave-Assisted Extraction. Agriculture 2020, 10, 435. [Google Scholar] [CrossRef]
- Sun-Waterhouse, D.; Wen, I.; Wibisono, R.; Melton, L.; Wadhwa, S. Evaluation of the extraction efficiency for polyphenol extracts from by-products of green kiwifruit juicing. Int. J. Food Sci. Technol. 2009, 44, 2644–2652. [Google Scholar] [CrossRef]
- Deng, J.; Liu, Q.; Zhang, Q.; Zhang, C.; Liu, D.; Fan, D.; Yang, H. Comparative study on composition, physicochemical and antioxidant characteristics of different varieties of kiwifruit seed oil in China. Food Chem. 2018, 264, 411–418. [Google Scholar] [CrossRef]
- Ilie, G.-I.; Milea, Ș.-A.; Râpeanu, G.; Cîrciumaru, A.; Stănciuc, N. Sustainable Design of Innovative Kiwi Byproducts-Based Ingredients Containing Probiotics. Foods 2022, 11, 2334. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Kamphuis, M.M.; Mela, D.J.; Westerterp-Plantenga, M.S. Diacylglycerols affect substrate oxidation and appetite in humans. Am. J. Clin. Nutr. 2003, 77, 1133–1139. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yagi, N.; Asakawa, H.; Tokunaga, K.; Watanabe, H.; Matsuo, N.; Tokimitsu, I. Long-Term Ingestion of Dietary Diacylglycerol Lowers Serum Triacylglycerol in Type II Diabetic Patients with Hypertriglyceridemia. J. Nutr. 2001, 131, 3204–3207. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zhang, Q.; Dong, Z.; Yan, Y.; Fu, Y.; Liu, X.; Zhao, B.; Duan, X. Phosphatidylcholine Ameliorates LPS-Induced Systemic Inflammation and Cognitive Impairments via Mediating the Gut–Brain Axis Balance. J. Agric. Food Chem. 2020, 68, 14884–14895. [Google Scholar] [CrossRef] [PubMed]
- Ly, R.; MacIntyre, B.C.; Philips, S.M.; McGlory, C.; Mutch, D.M.; Britz-McKibbin, P. Lipidomic studies reveal two specific circulating phosphatidylcholines as surrogate biomarkers of the omega-3 index. J. Lipid Res. 2023, 64, 100445. [Google Scholar] [CrossRef] [PubMed]
- Rockenfeller, P.; Koska, M.; Pietrocola, F.; Minois, N.; Knittelfelder, O.; Sica, V.; Franz, J.; Carmona-Gutierrez, D.; Kroemer, G.; Madeo, F. Phosphatidylethanolamine positively regulates autophagy and longevity. Cell Death Differ. 2015, 22, 499–508. [Google Scholar] [CrossRef]
- Park, S.; Kim, B.-K.; Park, S.-K. Supplementation with phosphatidylethanolamine confers anti-oxidant and anti-aging effects via hormesis and reduced insulin/IGF-1-like signaling in C. elegans. Mech. Ageing Dev. 2021, 197, 111498. [Google Scholar] [CrossRef]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine Metabolism in Health and Disease. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 321, pp. 29–88. [Google Scholar] [CrossRef]
- Tsoupras, A.; Adamantidi, T.; Finos, M.A.; Philippopoulos, A.; Detopoulou, P.; Tsopoki, I.; Kynatidou, M.; Demopoulos, C.A. Re-Assessing the Role of Platelet Activating Factor and Its Inflammatory Signaling and Inhibitors in Cancer and Anti-Cancer Strategies. Front. Biosci.-Landmark 2024, 29, 345. [Google Scholar] [CrossRef]
- Toni, L.S.; Garcia, A.M.; Jeffrey, D.A.; Jiang, X.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Optimization of phenol-chloroform RNA extraction. MethodsX 2018, 5, 599–608. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Yield for Kiwi Extracts | Max | Min | Median |
|---|---|---|---|
| TAC% (g/100 mL of juice) | 0.032 | 0.030 | 0.030 |
| TLC% (g/100 mL of juice) | 0.066 | 0.045 | 0.066 |
| TL% (g/100 mL of juice) | 0.098 | 0.074 | 0.096 |
| TAC% (g/100 g of by-product) | 0.263 | 0.249 | 0.250 |
| TLC% (g/100 g of by-product) | 0.322 | 0.107 | 0.166 |
| TL%(g/100 g of by-product) | 0.584 | 0.356 | 0.416 |
| Juice | By-Product | ||||
|---|---|---|---|---|---|
| TCC of TAC | TCC of TLC | TCC of TL | TCC of TAC | TCC of TLC | TCC of TL |
| Median | Median | Median | Median | Median | Median |
| 1.96 | 2.82 | 5.37 | 1.54 | 1.17 | 2.77 |
| Max | Max | Max | Max | Max | Max |
| 6.77 | 3.41 | 9.59 | 1.60 | 2.34 | 3.07 |
| Min | Min | Min | Min | Min | Min |
| 1.91 | 1.87 | 3.78 | 0.73 | 1.08 | 2.62 |
| Juice | By-Product | ||||
|---|---|---|---|---|---|
| TPC of TAC | TPC of TLC | TPC of TL | TPC of TAC | TPC of TLC | TPC of TL |
| Median | Median | Median | Median | Median | Median |
| 29.26 | 15.24 | 44.50 | 20.51 | 17.91 | 40.88 |
| Max | Max | Max | Max | Max | Max |
| 48.69 | 16.59 | 55.75 | 25.61 | 22.45 | 43.51 |
| Min | Min | Min | Min | Min | Min |
| 6.74 | 7.06 | 23.33 | 18.42 | 11.39 | 31.89 |
| ABTS Values | |||
|---|---|---|---|
| Juice | Median | Max | Min |
| TAC | 4.54 | 13.64 | 4.48 |
| TLC | 1.81 | 2.56 | 1.08 |
| TL | 6.29 | 16.20 | 5.62 |
| By-product | Median | Max | Min |
| TAC | 2.88 | 2.93 | 2.17 |
| TLC | 5.97 | 16.37 | 1.58 |
| TL | 8.90 | 19.25 | 3.75 |
| TEAC Values | |||
|---|---|---|---|
| Juice | Median | Max | Min |
| TAC | 0.0015 | 0.0015 | 0.0014 |
| TLC | 0.0002 | 0.0008 | 0.0002 |
| TL | 0.0017 | 0.0022 | 0.0002 |
| By-product | Median | Max | Min |
| TAC | 0.0010 | 0.0013 | 0.0007 |
| TLC | 0.0015 | 0.0017 | 0.0003 |
| TL | 0.0015 | 0.0030 | 0.0010 |
| Compounds | β-Carotene | Catechin | Gallic Acid | Polar Lipid | Quercetin |
|---|---|---|---|---|---|
| Peak (cm−1) | 1644 | 3600–3100 | 3400–2800 | 3550–3100 | 3600–3050 |
| Functional Groups | C=C alkene | O-H alcohol | COOH carboxylic acid | O-H alcohol | O-H alcohol |
| Peak (cm−1) | 1630–1600 | 1610 | 2860 | 1670 | |
| Functional Groups | C=C aromatic, C=C alkene | C=C aromatic | C H alkane | C=C alkane, C=O ketone | |
| Peak (cm−1) | 1450 | 1635 | 1450 | ||
| Functional Groups | C=C aromatic | C=C alkene, C=O ketone | C=C aromatic | ||
| Peak (cm−1) | 814 (Fingerprint region) | 1455 | 950, 814,640 (Fingerprint region) | ||
| Functional Groups | C=C aromatic | ||||
| Peak(cm−1) | 822 (Fingerprint region) |
| FreeFattyAcids (FFA) Obtained Without Saponification of TAC Extracts | FattyAcids of PL, Obtained After Saponification of TAC Extracts | ||||||
|---|---|---|---|---|---|---|---|
| Emperical Name | Name for FYP Tables | Juice | By-Products | Emperical Name | Name for FYP Tables | Juice | By-Products |
| Caprylic | C8:0 | ND | ND | Caprylic | C8:0 | ND | 0.06 ± 0.004 |
| Pelargonic | C9:0 | 0.39 ± 0.05 | 0.34 ± 0.01 | Pelargonic | C9:0 | 0.04 ± 0.00 | 0.15 ± 0.013 |
| Capric | C10:0 | ND | ND | ||||
| Lauric | C12:0 | 0.38 ± 0.01 | 0.00 ± 0.00 | Lauric | C12:0 | 0.06 ± 0.01 | ND |
| Tridecylic | C13:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | Tridecylic | C13:0 | ND | 0.02 ± 0.002 |
| Myristic | C14:0 | 0.96 ± 0.06 | 0.85 ± 0.06 | Myristic | C14:0 | 0.33 ± 0.03 | 0.40 ± 0.041 |
| Pentadecylic | C15:0 | 0.00 ± 0.00 | 0.58 ± 0.03 | Pentadecylic | C15:0 | 0.11 ± 0.02 | ND |
| Palmitic | C16:0 | 24.63 ± 1.34 | 45.83 ± 0.69 | Palmitic | C16:0 | 18.12 ± 0.14 | 23.00 ± 2.097 |
| Palmitoleic | C16:1 c9 (n7 MUFA) | 0.56 ± 0.01 | 0.54 ± 0.03 | Palmitoleic | C16:1 c9 (n7 MUFA) | 1.46 ± 0.03 | 0.64 ± 0.075 |
| Margaric | C17:0 | 2.00 ± 0.49 | 1.43 ± 0.09 | Margaric | C17:0 | 0.45 ± 0.02 | 0.86 ± 0.136 |
| Stearic | C18:0 | 64.54 ± 1.06 | 38.87 ± 0.51 | Stearic | C18:0 | 10.20 ± 0.14 | 30.21 ± 2.065 |
| Oleic | C18:1 c9 (n9 MUFA) | 5.06 ± 0.09 | 9.23 ± 0.08 | Oleic | C18:1 c9 (n9 MUFA) | 30.00 ± 0.03 | 19.44 ± 0.525 |
| Linoleic | C18:2 c9,12 (n6 PUFA) | 0.62 ± 0.05 | 1.04 ± 0.02 | Linoleic | C18:2 c9,12 (n6 PUFA) | 9.13 ± 0.03 | 7.41 ± 0.586 |
| Linolenic | C18:3 c9,12,15 (n3 PUFA) | 0.87 ± 0.06 | 1.28 ± 0.04 | Linolenic | C18:3 c9,12,15 (n3 PUFA) | 28.24 ± 0.04 | 16.48 ± 4.696 |
| Stearidonic | C18:4 c6,9,12,15 (n3 PUFA) | ND | ND | Stearidonic | C18:4 c6,9,12,15 (n3 PUFA) | 0.65 ± 0.06 | 0.30 ± 0.005 |
| Nonadecylic | C19:0 | ND | ND | Nonadecylic | C19:0 | ND | ND |
| Arachidic | C20:0 | ND | ND | ||||
| Gadoleic | C20:1 c9 (n11 MUFA) | ND | ND | Gadoleic | C20:1 c9 (n11 MUFA) | 0.49 ± 0.01 | 0.62 ± 0.042 |
| DihomoLinoleic | C18:2 c10,12 (n6 PUFA) | ND | ND | DihomoLinoleic | C18:2 c10,12 (n6 PUFA) | 0.27 ± 0.01 | 0.27 ± 0.022 |
| Dihomolinolenic | C20:3 c8,11,14 (n6 PUFA) | ND | ND | Dihomolinolenic | C20:3 c8,11,14 (n6 PUFA) | 0.43 ± 0.01 | 0.16 ± 0.005 |
| Arachidonic | C20:4 c5,8,11,14 (n6 PUFA) | ND | ND | Arachidonic | C20:4 c5,8,11,14 (n6 PUFA) | ND | ND |
| EPA | C20:5 c5,8,11,14,17 (n3 PUFA) | ND | ND | EPA | C20:5 c5,8,11,14,17 (n3 PUFA) | ND | ND |
| Docosadienoic | C22:2 c13,16 (n6 PUFA) | ND | ND | Docosadienoic | C22:2 c13,16 (n6 PUFA) | ND | ND |
| Eranthic | C22:3 c5,13,16 (n6 PUFA) | ND | ND | Eranthic | C22:3 c5,13,16 (n6 PUFA) | ND | ND |
| Adrenic | C22:4 c7,10,13,16 (n6 PUFA) | ND | ND | Adrenic | C22:4 c7,10,13,16 (n6 PUFA) | ND | ND |
| DPA | C22:5 c7,10,13,16,19 (n3 PUFA) | ND | ND | DPA | C22:5 c7,10,13,16,19 (n3 PUFA) | ND | ND |
| DHA | C22:6 c4,7,10,13,16,19 (n3 PUFA) | ND | ND | DHA | C22:6 c4,7,10,13,16,19 (n3 PUFA) | 0.02 ± 0.00 | ND |
| SFA | 92.89 ± 0.06 | 87.91 ± 0.08 | SFA | 29.31 ± 0.09 * | 54.68 ± 4.231 * | ||
| UFA | 7.11 ± 0.06 | 12.09 ± 0.08 | UFA | 70.69 ± 0.09 | 45.32 ± 4.231 | ||
| MUFA | 5.62 ± 0.10 | 9.78 ± 0.05 | MUFA | 31.94 ± 0.05 | 20.70 ± 0.520 | ||
| PUFA | 1.49 ± 0.05 | 2.32 ± 0.03 | PUFA | 38.75 ± 0.05 | 24.62 ± 4.111 | ||
| n3PUFA | 0.87 ± 0.06 | 1.28 ± 0.04 | n3PUFA | 28.91 ± 0.07 | 16.78 ± 4.692 | ||
| n6PUFA | 0.62 ± 0.05 | 1.04 ± 0.02 | n6PUFA | 9.83 ± 0.05 | 7.84 ± 0.606 | ||
| n6/n3 | 0.71 ± 0.09 | 0.82 ± 0.04 | n6/n3 | 0.34 ± 0.00 | 0.51 ± 0.209 | ||
| TAC Extracts from Kiwi Juice | TAC Extracts from Kiwi By-Products | |||||||
|---|---|---|---|---|---|---|---|---|
| Mainclasses of PL | Elutiontime (min) | Mr | Representative Molecular Species | Proposed Structures | Elutiontime (min) | Mr | Representative Molecular Species | Proposed Structures |
| DGs | 12–12.7 | 679.435 | DG 38:2;O2 | [i.e., DG 20:1/18:1] | 2.629 | 637.3073 | DG 36:1;O | [i.e., DG 18:0/18:1 or DG 16:0/20:1] |
| 12–12.7 | 585.3298 | DG 34:5 | [i.e., DG 16:1/18:4] | 2.629 | 637.3073 | DG O-36:2;O2 | [i.e., DG 18:1/18:1 or DG 16:1/20:1] | |
| 12–12.7 | 585.3298 | DG O-32:0;O2 | [i.e., DG 16:0/16:0] | 10.241–10.838 | 565.3522 | DG 32:1 | [i.e., DG 16:0/16:1] | |
| 17.1–17.2 | 635.4519 | DG 36:2;O | [i.e., DG 18:1/18:1 or 18:2/18:0 ] | 10.241–10.838 | 565.3522 | DG O-32:2;O | [i.e., DG 16:1/16:1] | |
| 17.1–17.2 | 635.4519 | DG O-36:3;O2 | [i.e., DG 18:1/18:2 or 18:3/18:0 ] | 10.241–10.838 | 633.3401 | DG 36:3;O | [i.e., DG 18:2/18:1] | |
| 17.1–17.2 | 635.4519 | DG O-38:1 | [i.e., DG 20:1/18:0] | 10.241–10.838 | 633.3401 | DG O-36:4;O2 | [i.e., DG 18:3/18:1 or DG 18:2/18:2] | |
| 12.297 | 589.5187 | DG 34:3 | [i.e., DG 18:2/16:1 or DG 18:3/16:0] | |||||
| 12.297 | 589.5187 | DG O-34:4;O | [i.e., DG 18:4/16:0 or DG 18:3/16:1] | |||||
| PCs | 5.6–7.6 | 792.8564 | PC 38:5 | [i.e., PC 20:1/18:4] | 15.199–15.829 | 770.8649 | PC 36:2 | [i.e., PC 16:1/20:1 or PC 18:1/18:1] |
| 8.7–9.2 | 792.8560 | PC 38:5 | [i.e., PC 20:1/18:4] | 15.199–15.829 | 770.8649 | PC O-36:3;O | [i.e., PC 18:2/18:1] | |
| 14.1–14.6 | 792.8568 | PC 38:5 | [i.e., PC 20:1/18:4] | |||||
| 15–15.1 | 792.8593 | PC 38:5 | [i.e., PC 20:1/18:4] | |||||
| 16.2–16.5 | 792.8642 | PC 38:5 | [i.e., PC 20:1/18:4] | |||||
| PEs | 10.2–11.7 | 698.4174 | PE O-34:3 | [i.e., PE 18:3/16:0 or PE 18:2/16:1] | 15.199–15.829 | 770.8649 | PE 38:2 | [i.e., PE 18:1/20:1] |
| 10.2–11.7 | 744.4209 | PE 36:1 | [i.e., PE 18:1/18:0] | 7.604–7.72 | 770.8649 | PE O-38:3;O | [i.e., PE 18:2/20:1] | |
| 10.2–11.7 | 744.4209 | PE O-36:2;O | [i.e., PE 18:2/18:0 or PE 18:1/18:1 or PE 20:1/16:0] | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moysidou, A.M.; Cheimpeloglou, K.; Koutra, S.I.; Manousakis, V.; Ofrydopoulou, A.; Shiels, K.; Saha, S.K.; Tsoupras, A. In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of Bioactive Metabolites Extracted from Kiwi and Its By-Products. Metabolites 2025, 15, 400. https://doi.org/10.3390/metabo15060400
Moysidou AM, Cheimpeloglou K, Koutra SI, Manousakis V, Ofrydopoulou A, Shiels K, Saha SK, Tsoupras A. In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of Bioactive Metabolites Extracted from Kiwi and Its By-Products. Metabolites. 2025; 15(6):400. https://doi.org/10.3390/metabo15060400
Chicago/Turabian StyleMoysidou, Anastasia Maria, Konstantina Cheimpeloglou, Spyridoula Ioanna Koutra, Vasileios Manousakis, Anna Ofrydopoulou, Katie Shiels, Sushanta Kumar Saha, and Alexandros Tsoupras. 2025. "In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of Bioactive Metabolites Extracted from Kiwi and Its By-Products" Metabolites 15, no. 6: 400. https://doi.org/10.3390/metabo15060400
APA StyleMoysidou, A. M., Cheimpeloglou, K., Koutra, S. I., Manousakis, V., Ofrydopoulou, A., Shiels, K., Saha, S. K., & Tsoupras, A. (2025). In Vitro Antioxidant, Antithrombotic and Anti-Inflammatory Activities of Bioactive Metabolites Extracted from Kiwi and Its By-Products. Metabolites, 15(6), 400. https://doi.org/10.3390/metabo15060400