
Blood Metabolic Biomarkers of Diabetes Mellitus Type 2 in Aged Adults Determined by a UPLC-MS Metabolomic Approach

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Population Characteristics and Recruitment
2.3. Metabolomic Experiments
2.3.1. Sample Harvesting and Processing
2.3.2. UPLC-MS Measurements
2.3.3. UPLC-MS Data Analysis
2.3.4. Classical Statistical Analysis
3. Results
3.1. Evaluation of Cohort Characteristics
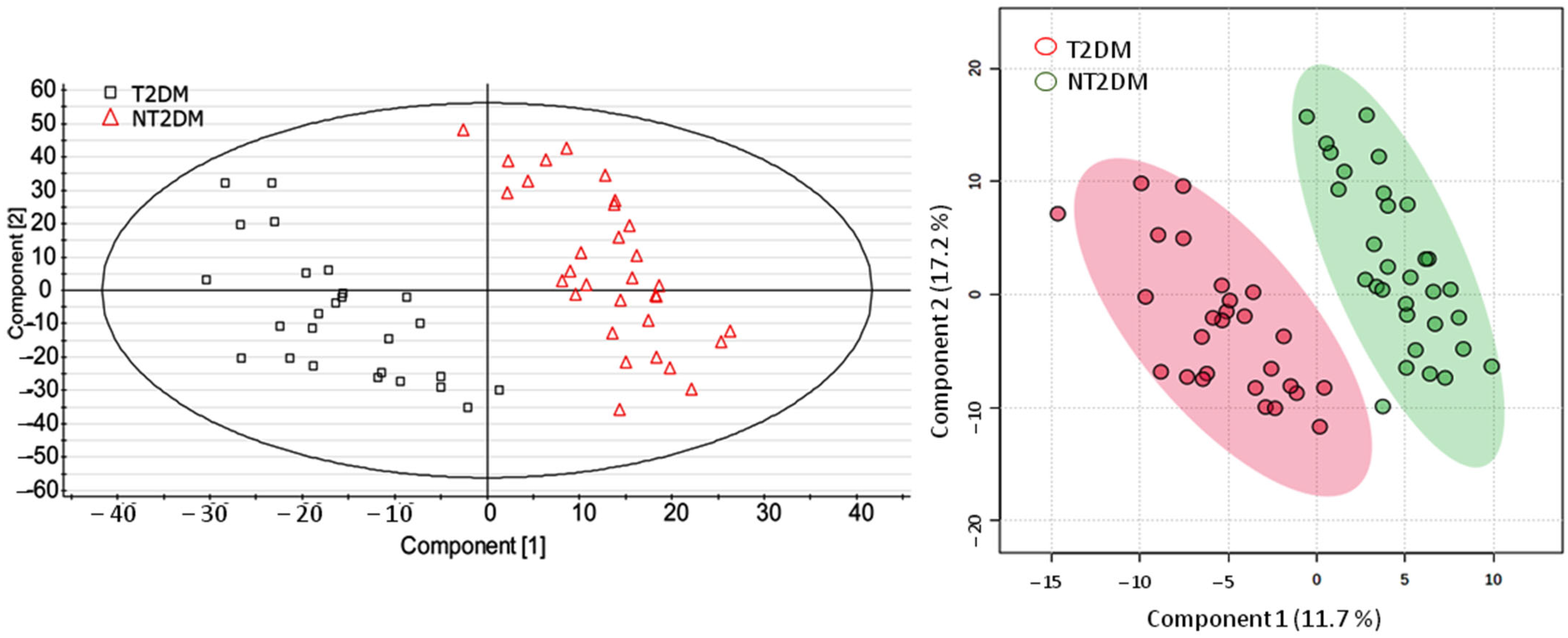
3.2. UPLC-MS Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| UPLC | ultrahigh-pressure liquid chromatography |
| MS | mass spectrometry |
| QToF | quadrupole time of flight |
| XS | extended statistics |
| MEDAS-14 | 14-item Mediterranean Diet Adherence Screener |
| LPC | lysophosphatidylcholine |
| T2DM | type 2 diabetes mellitus |
| GSH | glutathione |
| BCAAs | branched-chain amino acids |
| AAAs | aromatic amino acids |
| IR | insulin resistance |
| NT2DM | non-diabetic metabolic syndrome |
| PC | phosphatidylcholine |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| TLR4 | Toll-like receptor 4 |
| CE | cholesteryl ester |
| OR | odds ratio |
| CI | confidence interval |
References
- Magliano, D.; Boyko, E.J. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; ISBN 978-2-930229-98-0. [Google Scholar]
- Lu, X.; Xie, Q.; Pan, X.; Zhang, R.; Zhang, X.; Peng, G.; Zhang, Y.; Shen, S.; Tong, N. Type 2 Diabetes Mellitus in Adults: Pathogenesis, Prevention and Therapy. Sig. Transduct. Target. Ther. 2024, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Fernández De Alba, I.; Orlando, V.; Monetti, V.M.; Mucherino, S.; Gimeno-Miguel, A.; Vaccaro, O.; Forjaz, M.J.; Poblador Plou, B.; Prados-Torres, A.; Riccardi, G.; et al. Comorbidity in an Older Population with Type-2 Diabetes Mellitus: Identification of the Characteristics and Healthcare Utilization of High-Cost Patients. Front. Pharmacol. 2020, 11, 586187. [Google Scholar] [CrossRef]
- Johannsen, D.L.; Conley, K.E.; Bajpeyi, S.; Punyanitya, M.; Gallagher, D.; Zhang, Z.; Covington, J.; Smith, S.R.; Ravussin, E. Ectopic Lipid Accumulation and Reduced Glucose Tolerance in Elderly Adults Are Accompanied by Altered Skeletal Muscle Mitochondrial Activity. J. Clin. Endocrinol. Metab. 2012, 97, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Arneth, B.; Arneth, R.; Shams, M. Metabolomics of Type 1 and Type 2 Diabetes. Int. J. Mol. Sci. 2019, 20, 2467. [Google Scholar] [CrossRef]
- Arroyo, M.N.; Green, J.A.; Cnop, M.; Igoillo-Esteve, M. tRNA Biology in the Pathogenesis of Diabetes: Role of Genetic and Environmental Factors. Int. J. Mol. Sci. 2021, 22, 496. [Google Scholar] [CrossRef]
- Ferdous, S.-E.; Ferrell, J.M. Pathophysiological Relationship between Type 2 Diabetes Mellitus and Metabolic Dysfunction-Associated Steatotic Liver Disease: Novel Therapeutic Approaches. Int. J. Mol. Sci. 2024, 25, 8731. [Google Scholar] [CrossRef]
- Pallares-Méndez, R.; Aguilar-Salinas, C.A.; Cruz-Bautista, I.; Del Bosque-Plata, L. Metabolomics in Diabetes, a Review. Ann. Med. 2016, 48, 89–102. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Xu, F.; Tavintharan, S.; Sum, C.F.; Woon, K.; Lim, S.C.; Ong, C.N. Metabolic Signature Shift in Type 2 Diabetes Mellitus Revealed by Mass Spectrometry-Based Metabolomics. J. Clin. Endocrinol. Metab. 2013, 98, E1060–E1065. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Koulman, A.; Griffin, J.L. Towards Metabolic Biomarkers of Insulin Resistance and Type 2 Diabetes: Progress from the Metabolome. Lancet Diabetes Endocrinol. 2014, 2, 65–75. [Google Scholar] [CrossRef]
- Feng, L.; He, B.; Xia, J.; Wang, Z. Untargeted and Targeted Lipidomics Unveil Dynamic Lipid Metabolism Alterations in Type 2 Diabetes. Metabolites 2024, 14, 610. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-Chain Amino Acids in Metabolic Signalling and Insulin Resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-S.; Ahn, E.; Park, E.S.; Huh, T.; Choi, S.; Kwon, Y.; Choi, B.H.; Lee, J.; Choi, Y.H.; Jeong, Y.L.; et al. Impaired BCAA Catabolism in Adipose Tissues Promotes Age-Associated Metabolic Derangement. Nat. Aging 2023, 3, 982–1000. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Hyun, S.; Koo, S.-H. The Role of BCAA Metabolism in Metabolic Health and Disease. Exp. Mol. Med. 2024, 56, 1552–1559. [Google Scholar] [CrossRef]
- Shahisavandi, M.; Wang, K.; Ghanbari, M.; Ahmadizar, F. Exploring Metabolomic Patterns in Type 2 Diabetes Mellitus and Response to Glucose-Lowering Medications—Review. Genes 2023, 14, 1464. [Google Scholar] [CrossRef]
- Nguyen, D.; Samson, S.L.; Reddy, V.T.; Gonzalez, E.V.; Sekhar, R.V. Impaired Mitochondrial Fatty Acid Oxidation and Insulin Resistance in Aging: Novel Protective Role of Glutathione. Aging Cell 2013, 12, 415–425. [Google Scholar] [CrossRef]
- Gu, X.; Al Dubayee, M.; Alshahrani, A.; Masood, A.; Benabdelkamel, H.; Zahra, M.; Li, L.; Abdel Rahman, A.M.; Aljada, A. Distinctive Metabolomics Patterns Associated With Insulin Resistance and Type 2 Diabetes Mellitus. Front. Mol. Biosci. 2020, 7, 609806. [Google Scholar] [CrossRef]
- Cortés Tormo, M.; Marcos Tomás, J.V.; Giner Galvañ, V.; Redón I Mas, J. Metabolomics as a tool towards personalized medicine in type 2 diabetes mellitus. Rev. Med. Lab. 2021, 2, 30–40. [Google Scholar] [CrossRef]
- Albillos, S.M.; Montero, O.; Calvo, S.; Solano-Vila, B.; Trejo, J.M.; Cubo, E. Plasma Acyl-Carnitines, Bilirubin, Tyramine and Tetrahydro-21-Deoxycortisol in Parkinson’s Disease and Essential Tremor. A Case Control Biomarker Study. Park. Relat. Disord. 2021, 91, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; François, R.; Henry, L.; Müller, K. Vaughan D. Dplyr: A Grammar of Data Manipulation. R Package Version 1.1.4. Available online: https://dplyr.tidyverse.org (accessed on 12 July 2024).
- Sjoberg, D.D.; Whiting, K.; Curry, M.; Lavery, J.A.; Larmarange, J. Reproducible Summary Tables with the Gtsummary Package. R J. 2021, 13, 570. [Google Scholar] [CrossRef]
- Ahola-Olli, A.V.; Mustelin, L.; Kalimeri, M.; Kettunen, J.; Jokelainen, J.; Auvinen, J.; Puukka, K.; Havulinna, A.S.; Lehtimäki, T.; Kähönen, M.; et al. Circulating Metabolites and the Risk of Type 2 Diabetes: A Prospective Study of 11,896 Young Adults from Four Finnish Cohorts. Diabetologia 2019, 62, 2298–2309. [Google Scholar] [CrossRef]
- Kimura, K.; Nakamura, Y.; Inaba, Y.; Matsumoto, M.; Kido, Y.; Asahara, S.; Matsuda, T.; Watanabe, H.; Maeda, A.; Inagaki, F.; et al. Histidine Augments the Suppression of Hepatic Glucose Production by Central Insulin Action. Diabetes 2013, 62, 2266–2277. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; McCarty, M.F.; OKeefe, J.H. Role of Dietary Histidine in the Prevention of Obesity and Metabolic Syndrome. Open Heart 2018, 5, e000676. [Google Scholar] [CrossRef]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.-P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. α-Hydroxybutyrate Is an Early Biomarker of Insulin Resistance and Glucose Intolerance in a Nondiabetic Population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.-G.; Fritsche, A.; Häring, H.-U.; Hrabě De Angelis, M.; Peters, A.; et al. Identification of Serum Metabolites Associated With Risk of Type 2 Diabetes Using a Targeted Metabolomic Approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.-P.; Milburn, M.V.; Kastenmüller, G.; Adamski, J.; Tuomi, T.; et al. Early Metabolic Markers of the Development of Dysglycemia and Type 2 Diabetes and Their Physiological Significance. Diabetes 2013, 62, 1730–1737. [Google Scholar] [CrossRef]
- Wittenbecher, C.; Mühlenbruch, K.; Kröger, J.; Jacobs, S.; Kuxhaus, O.; Floegel, A.; Fritsche, A.; Pischon, T.; Prehn, C.; Adamski, J.; et al. Amino Acids, Lipid Metabolites, and Ferritin as Potential Mediators Linking Red Meat Consumption to Type 2 Diabetes. Am. J. Clin. Nutr. 2015, 101, 1241–1250. [Google Scholar] [CrossRef]
- Drabkova, P.; Sanderova, J.; Kovarik, J.; Kandar, R. An Assay of Selected Serum Amino Acids in Patients with Type 2 Diabetes Mellitus. Adv. Clin. Exp. Med. 2015, 24, 447–451. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine Metabolism in Animals and Humans: Implications for Nutrition and Health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.Y.; Kim, J.Y.; Paik, J.K.; Kim, O.Y.; Paik, Y.; Lee, E.J.; Lee, J.H. The Association of Specific Metabolites of Lipid Metabolism with Markers of Oxidative Stress, Inflammation and Arterial Stiffness in Men with Newly Diagnosed Type 2 Diabetes. Clin. Endocrinol. 2012, 76, 674–682. [Google Scholar] [CrossRef]
- Lewis, G.F.; Carpentier, A.; Adeli, K.; Giacca, A. Disordered Fat Storage and Mobilization in the Pathogenesis of Insulin Resistance and Type 2 Diabetes. Endocr. Rev. 2002, 23, 201–229. [Google Scholar] [CrossRef] [PubMed]
- Wang-Sattler, R.; Yu, Z.; Herder, C.; Messias, A.C.; Floegel, A.; He, Y.; Heim, K.; Campillos, M.; Holzapfel, C.; Thorand, B.; et al. Novel Biomarkers for Pre-diabetes Identified by Metabolomics. Mol. Syst. Biol. 2012, 8, 615. [Google Scholar] [CrossRef]
- Mirza, A.Z.; Althagafi, I.I.; Shamshad, H. Role of PPAR Receptor in Different Diseases and Their Ligands: Physiological Importance and Clinical Implications. Eur. J. Med. Chem. 2019, 166, 502–513. [Google Scholar] [CrossRef]
- Donath, M.Y.; Shoelson, S.E. Type 2 Diabetes as an Inflammatory Disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism Pathways of Arachidonic Acids: Mechanisms and Potential Therapeutic Targets. Sig. Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Samouda, H. Clinical and Biological Risk Factors Associated with Inflammation in Patients with Type 2 Diabetes Mellitus. BMC Endocr. Disord. 2022, 22, 16. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, J.; Zhang, W.; Guo, Z.; Ma, Q. Arachidonic Acid Metabolism in Health and Disease. MedComm 2023, 4, e363. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Zhang, W.; Cai, Y.; Shan, P.; Wu, D.; Zhang, B.; Liu, H.; Khan, Z.A.; Liang, G. Arachidonic Acid Inhibits Inflammatory Responses by Binding to Myeloid Differentiation Factor-2 (MD2) and Preventing MD2/Toll-like Receptor 4 Signaling Activation. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2020, 1866, 165683. [Google Scholar] [CrossRef]
- Liu, M.; Subramanian, V.S.; Subbaiah, P.V. Modulation of the Positional Specificity of Lecithin−Cholesterol Acyltransferase by the Acyl Group Composition of Its Phosphatidylcholine Substrate: Role of the Sn -1-Acyl Group. Biochemistry 1998, 37, 13626–13633. [Google Scholar] [CrossRef] [PubMed]
- Nakhjavani, M.; Morteza, A.; Karimi, R.; Banihashmi, Z.; Esteghamati, A. Diabetes Induces Gender Gap on LCAT Levels and Activity. Life Sci. 2013, 92, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Bandet, C.L.; Mahfouz, R.; Véret, J.; Sotiropoulos, A.; Poirier, M.; Giussani, P.; Campana, M.; Philippe, E.; Blachnio-Zabielska, A.; Ballaire, R.; et al. Ceramide Transporter CERT Is Involved in Muscle Insulin Signaling Defects Under Lipotoxic Conditions. Diabetes 2018, 67, 1258–1271. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, R.; Schulze, H.; Sandhoff, K. Ganglioside Metabolism in Health and Disease. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2018; Volume 156, pp. 1–62. ISBN 978-0-12-812341-6. [Google Scholar]
- Xia, Q.-S.; Lu, F.-E.; Wu, F.; Huang, Z.-Y.; Dong, H.; Xu, L.-J.; Gong, J. New Role for Ceramide in Hypoxia and Insulin Resistance. World J. Gastroenterol. 2020, 26, 2177–2186. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Variable | NT2DM Group (n = 32) | T2DM Group (n = 27) |
|---|---|---|
| Demographics and anthropometric characteristics | ||
| Age (years; p = 0.004) | 71.4 ± 9.0 | 75.9 ± 8.1 |
| Sex (female/male) | 21 (66%)/11 (34%) | 15 (56%)/12 (44%) |
| BMI (kg/m2) 1 | 28.8 ± 5.8 | 28.5 ± 6.3 |
| Underweight | 4 (13%) | 3 (11%) |
| Normal | 7 (22%) | 9 (33%) |
| Overweight | 7 (22%) | 6 (22%) |
| Obese | 14 (44%) | 9 (33%) |
| Waist circumference—female (cm) 2 | 100.6 ± 16.0 (n = 21) | 100.6 ± 14.9 (n = 15) |
| Waist circumference—male (cm) 2 | 104.8 ± 10.4 (n = 11) | 108.9 ± 11.4 (n = 12) |
| Lifestyle and dietary habits | ||
| Alcohol intake 3 (p = 0.049) | 15 (47%) | 6 (22%) |
| Smoking status | ||
| Never smoker | 22 (69%) | 20 (74%) |
| Former smoker | 6 (19%) | 5 (19%) |
| Current smoker | 4 (13%) | 2 (7.4%) |
| Physical activity (MET h/week) 4 | 77.1 ± 93.8 | 64.9 ± 67.0 |
| Vigorous intensity | 16 (50%) | 14 (52%) |
| Moderate | 10 (31%) | 7 (26%) |
| Light | 2 (6.3%) | 2 (7.4%) |
| Rest | 4 (13%) | 4 (15%) |
| Mediterranean diet score (MEDAS-14) 5 | 8.6 ± 1.5 | 7.9 ± 1.1 |
| Low adherence | 7 (22%) | 8 (30%) |
| Moderate adherence | 19 (59%) | 18 (67%) |
| Strong adherence | 6 (19%) | 1 (3.7%) |
| High sugar intake 6 (p = 0.013) | 21 (66%) | 9 (33%) |
| Sugar intake (g/day: p = 0.019) | 47.3 ± 83.7 | 26.7 ± 55.5 |
| High fat intake 7 | 13 (41%) | 8 (30%) |
| Meat intake type | ||
| No-meat diet | 5 (16%) | 6 (22%) |
| White and processed meat | 8 (25%) | 7 (26%) |
| Red and processed meat | 8 (25%) | 11 (41%) |
| White meat | 11 (34%) | 3 (11%) |
| Family history, treatments, polypharmacy, blood pressure, and biochemical parameters | ||
| Family history of cardiovascular disease | 15 (47%) | 7 (26%) |
| Family history of endocrine disease (p < 0.001) | 8 (25%) | 19 (70%) |
| Treatment for dyslipidemia | 11 (34%) | 12 (44%) |
| Polypharmacy (≥3 medications, p < 0.001) | 12 (38%) | 24 (89%) |
| Systolic blood pressure, left arm (mmHg) (p = 0.038) | 134.6 ± 19.4 | 138.4 ± 15.2 |
| Diastolic blood pressure, right arm (mmHg) | 83.1 ± 9.0 | 79.1 ± 8.5 |
| HbA1c (%) | - | 6.9 ± 0.9 |
| Fasting glucose (mmol/L, p < 0.001) | 5.1 ± 0.6 | 7.2 ± 1.9 |
| Total serum cholesterol (mg/dL, p < 0.001) | 193.0 ± 28.8 | 164.2 ± 36.3 |
| HDL (mg/dL) | 61.8 ± 16.2 | 61.6 ± 32.9 |
| LDL (mg/dL, p < 0.001) | 109.0 ± 24.2 | 86.4 ± 29.5 |
| Triglycerides (mg/dL) | 102.4 ± 37.3 | 105.4 ± 43.0 |
| TG/HDL ratio | 1.8 ± 1.0 | 2.0 ± 1.1 |
| LDL/HDL cholesterol ratio | 1.7 ± 0.6 | 1.6 ± 0.7 |
| High cardiovascular risk | 13 (41%) | 11 (41%) |
| Metabolite | Formula [M + H]+ | m/z | Normalized Chromatographic Peak Areas | Retention Time (min) | FC (Log2) | Regulation | p | |
|---|---|---|---|---|---|---|---|---|
| T2DM | NT2DM | |||||||
| LPC(14:0) | C22H47NO7P | 468.3072 | 0.032 ± 0.016 | 0.053 ± 0.029 | 3.18 | −0.73 | Down | <0.001 |
| LPC(16:0) | C24H50NO7P | 496.3413 | 3.321 ± 0.982 | 4.075 ± 0.984 | 3.72 | −0.29 | Down | 0.003 |
| LPC(18:0) | C26H54NO7P | 525.3698 | 0.235 ± 0.067 | 0.332 ± 0.109 | 4.68 | −0.50 | Down | <0.001 |
| LPC(18:1) | C26H52N89P | 522.3556 | 1.253 ± 0.502 | 1.306 ± 0.494 | 3.90 | −0.06 | Down | 0.344 |
| LPC(18:2) | C26H50NO7P | 520.3401 | 1.834 ± 0.794 | 2.320 ± 1.027 | 3.45 | −0.34 | Down | 0.023 |
| LPC(20:4) | C28H50NO7P | 544.3397 | 0.323 ± 0.139 | 0.313 ± 0.159 | 3.42 | +0.05 | Up | 0.400 |
| LPC(22:6) | C30H50NO7P | 569.3391 | 0.077 ± 0.038 | 0.083 ± 0.040 | 3.36 | −0.10 | Down | 0.296 |
| PC(16:0/18:2) | C42H80NO8P | 758.5605 | 2.29 × 10−4 ± 6.55 × 10−4 | 6.22 × 10−4 ± 12.6 × 10−4 | 7.54 | −1.44 | Down | 0.081 |
| Ganglioside 1 | C75H137N3O27 | 754.9894 | 0.074 ± 0.041 | 0.096 ± 0.045 | 3.72 | −0.37 | Down | 0.032 |
| Ganglioside 2 | C75H135N3O27 | 762.9800 | 0.013 ± 0.010 | 0.019 ± 0.009 | 3.72 | −0.57 | Down | 0.009 |
| Ganglioside 3 | C78H142N2O31 | 791.4910 | 0.017 ± 0.014 | 0.024 ± 0.022 | 3.44 | −0.52 | Down | 0.080 |
| Glycine-Histidine | C8H12N4O3 | 195.0888 | 0.008 ± 0.013 | 0.021 ± 0.035 | 2.18 | −1.43 | Down | 0.040 |
| Unidentified 1 | 531.3243 | 0.100 ± 0.056 | 0.133 ± 0.064 | 3.44 | −0.41 | Down | 0.020 | |
| Gly-His | LPC(22:6) | LPC(20:4) | LPC(14:0) | Ganglioside 2 | MEDAS-14 | |
|---|---|---|---|---|---|---|
| Gly-His | - | −0.211 (0.113) | −0.057 (0.669) | 0.366 (0.005) | 0.006 (0.963) | −0.412 (0.001) |
| LPC(22:6) | −0.211 (0.113) | - | 0.513 (<0.001) | 0.250 (0.059) | 0.540 (<0.001) | 0.197 (0.137) |
| LPC(20:4) | −0.057 (0.669) | 0.513 (<0.001) | - | 0.190 (0.153) | 0.333 (0.011) | −0.042 (0.752) |
| LPC(14:0) | 0.366 (0.005) | 0.250 (0.059) | 0.190 (0.153) | - | 0.480 (<0.001) | −0.037 (0.784) |
| Ganglioside 2 | 0.006 (0.963) | 0.540 (<0.001) | 0.333 (0.011) | 0.480 (<0.001) | - | 0.106 (0.429) |
| MEDAS-14 | −0.412 (0.001) | 0.197 (0.137) | −0.042 (0.752) | −0.037 (0.784) | 0.106 (0.429) | - |
| Univariate Analysis | Multivariate Analysis | |||||||
|---|---|---|---|---|---|---|---|---|
| Characteristics | n | OR | 95% CI | p-Value | OR | 95% CI | p-Value | |
| Gender masculine | 59 | 1.527 | 0.533–4.441 | 0.430 | 1.413 | 0.281–7.305 | 0.670 | |
| Gly-Hist | Gender | 59 | 0.994 | 0.985–1.000 | 0.108 | 0.995 | 0.985–1.003 | 0.349 |
| No gender | 59 | 0.994 | 0.986–1.000 | 0.108 | 0.996 | 0.986–1.003 | 0.336 | |
| LPC(22:6) | Gender | 59 | 1.000 | 0.997–1.003 | 0.624 | 1.001 | 0.995–1.008 | 0.652 |
| No gender | 59 | 1.001 | 0.998–1.004 | 0.624 | 1.001 | 0.995–1.008 | 0.740 | |
| LPC(20:4) | Gender | 59 | 1.000 | 0.999–1.001 | 0.100 | 1.001 | 1.000–1.003 | 0.049 |
| No gender | 59 | 1.001 | 0.999–1.002 | 0.100 | 1.002 | 1.001–1.004 | 0.026 | |
| LPC(14:0) | Gender | 59 | 0.990 | 0.982–0.996 | 0.009 | 0.988 | 0.977–0.997 | 0.018 |
| No gender | 59 | 0.991 | 0.983–0.997 | 0.009 | 0.989 | 0.978–0.997 | 0.019 | |
| Ganglioside 2 | Gender | 59 | 0.986 | 0.971–0.998 | 0.044 | 0.976 | 0.950–0.996 | 0.042 |
| No gender | 59 | 0.986 | 0.971–0.999 | 0.044 | 0.977 | 0.952–0.997 | 0.045 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simón, A.; Bordonaba-Bosque, D.; Montero, O.; Solano-Castán, J.; Caro, I. Blood Metabolic Biomarkers of Diabetes Mellitus Type 2 in Aged Adults Determined by a UPLC-MS Metabolomic Approach. Metabolites 2025, 15, 395. https://doi.org/10.3390/metabo15060395
Simón A, Bordonaba-Bosque D, Montero O, Solano-Castán J, Caro I. Blood Metabolic Biomarkers of Diabetes Mellitus Type 2 in Aged Adults Determined by a UPLC-MS Metabolomic Approach. Metabolites. 2025; 15(6):395. https://doi.org/10.3390/metabo15060395
Chicago/Turabian StyleSimón, Alba, Daniel Bordonaba-Bosque, Olimpio Montero, Javier Solano-Castán, and Irma Caro. 2025. "Blood Metabolic Biomarkers of Diabetes Mellitus Type 2 in Aged Adults Determined by a UPLC-MS Metabolomic Approach" Metabolites 15, no. 6: 395. https://doi.org/10.3390/metabo15060395
APA StyleSimón, A., Bordonaba-Bosque, D., Montero, O., Solano-Castán, J., & Caro, I. (2025). Blood Metabolic Biomarkers of Diabetes Mellitus Type 2 in Aged Adults Determined by a UPLC-MS Metabolomic Approach. Metabolites, 15(6), 395. https://doi.org/10.3390/metabo15060395