

Nitrogen Deprivation Drives Red Motile Cell Formation in Haematococcus pluvialis: Physiological and Transcriptomic Insights

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Algal Growth and Processing
2.2. Morphological Observation of Algal Cells
2.3. Determination of Astaxanthin Content
2.4. Chlorophyll Fluorescence Analysis
2.5. Analysis of Total Sugar Content
2.6. Quantification of Protein Content
2.7. Quantification of Starch Content
2.8. Quantification of Lipid Content
2.9. Quantification of Free Fatty Acids Composition
2.10. Qualitative and Quantitative Analysis of Reactive Oxygen Species
2.11. Transcriptome Analysis
2.12. Validation of DEGs by qRT-PCR
2.13. Statistical Analysis
3. Results
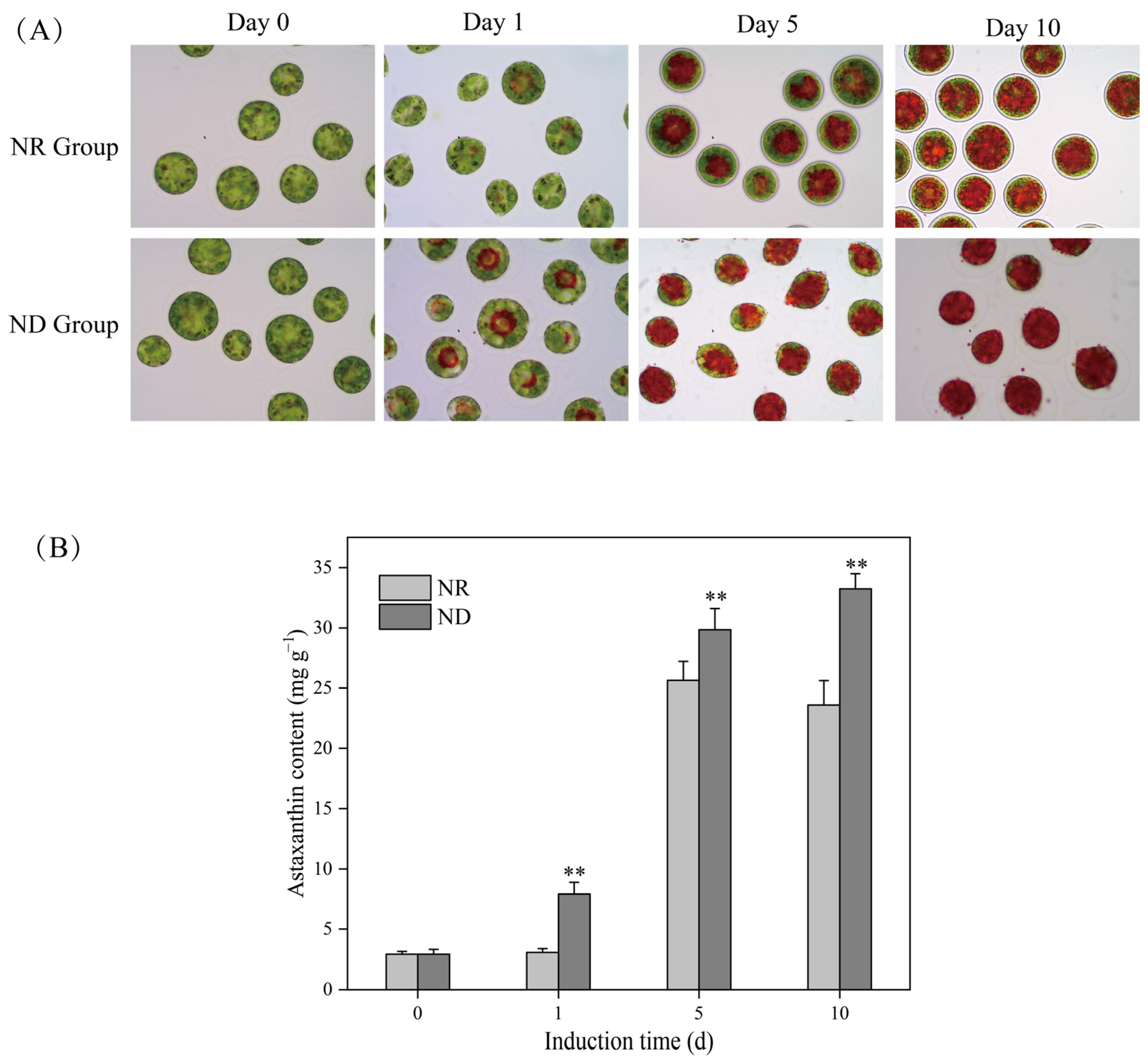
3.1. Effects of Nitrogen Deprivation on Morphology
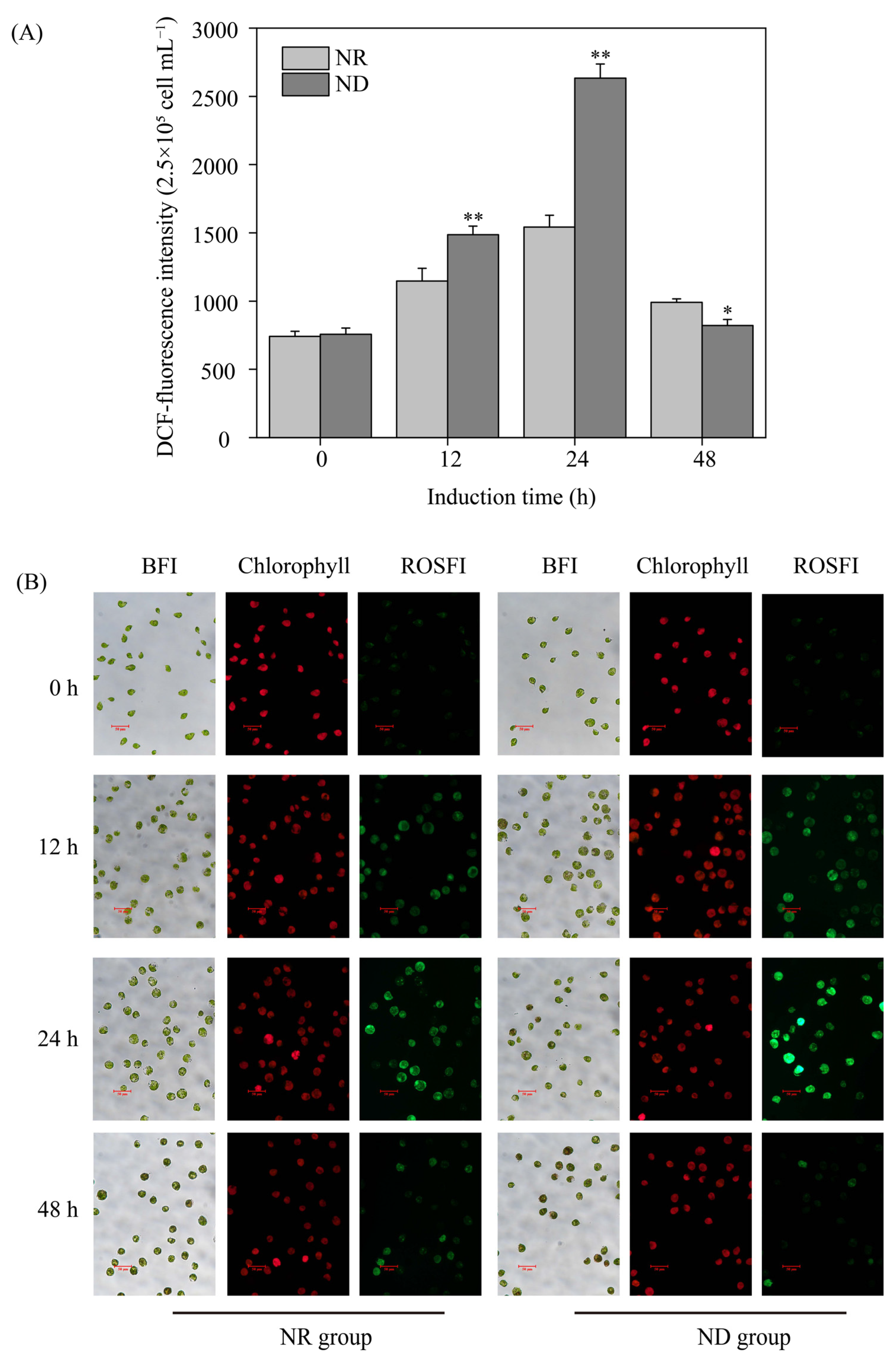
3.2. Effects of Nitrogen Deprivation on ROS Levels
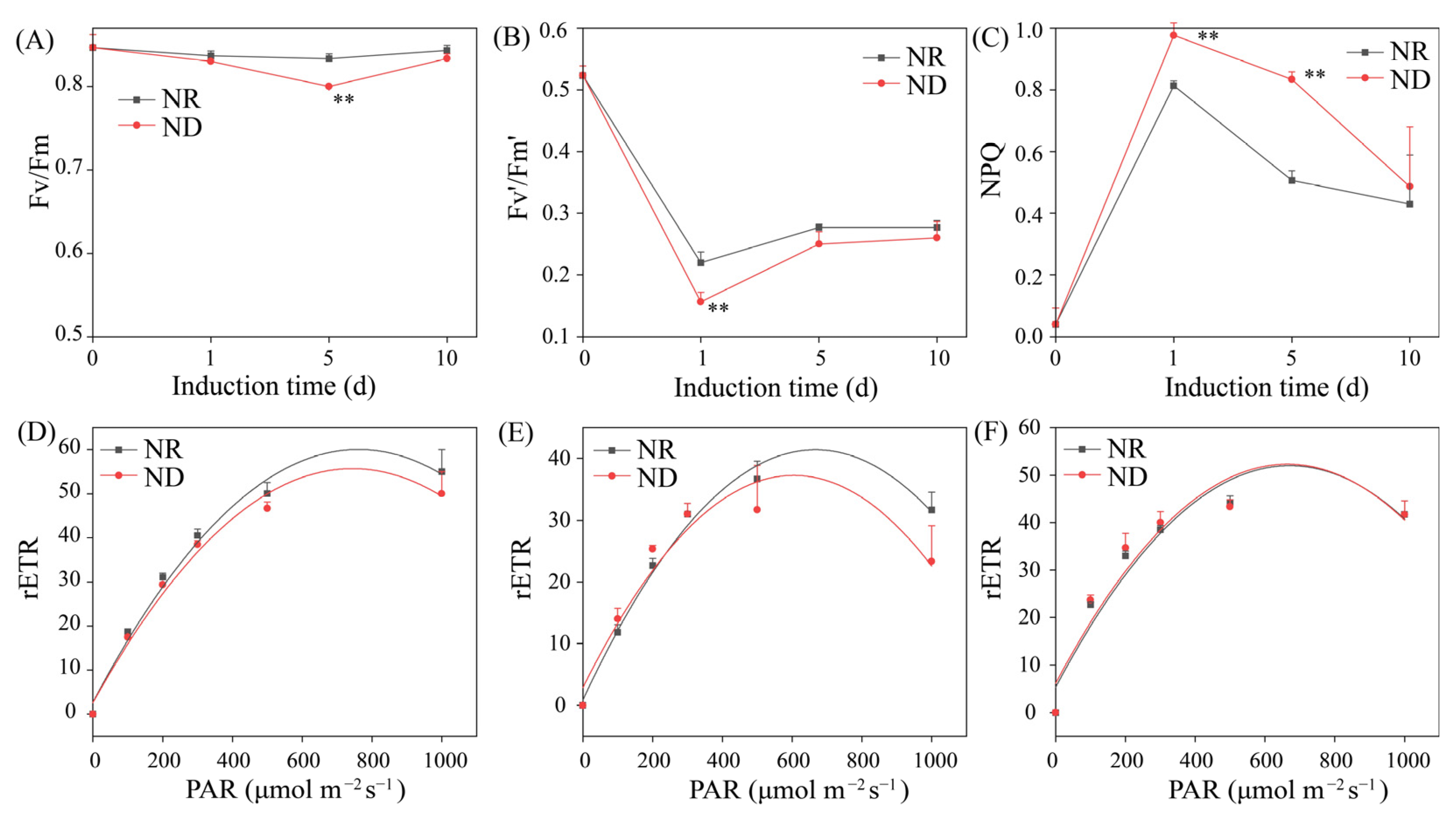
3.3. Effects of Nitrogen Deprivation on Photosynthetic Parameters
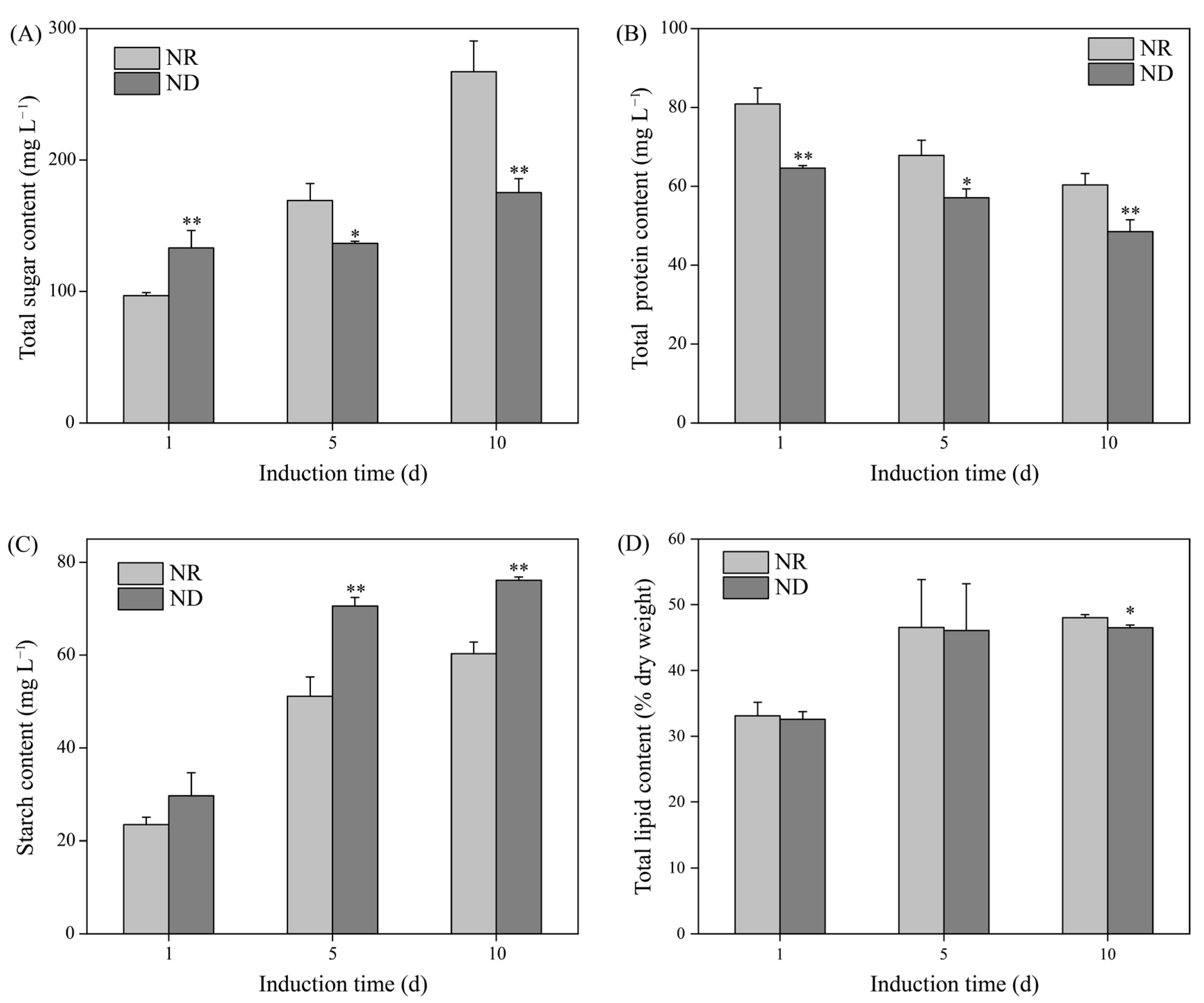
3.4. Effects of Nitrogen Deprivation on Contents of Total Sugar, Protein, Starch, and Lipid
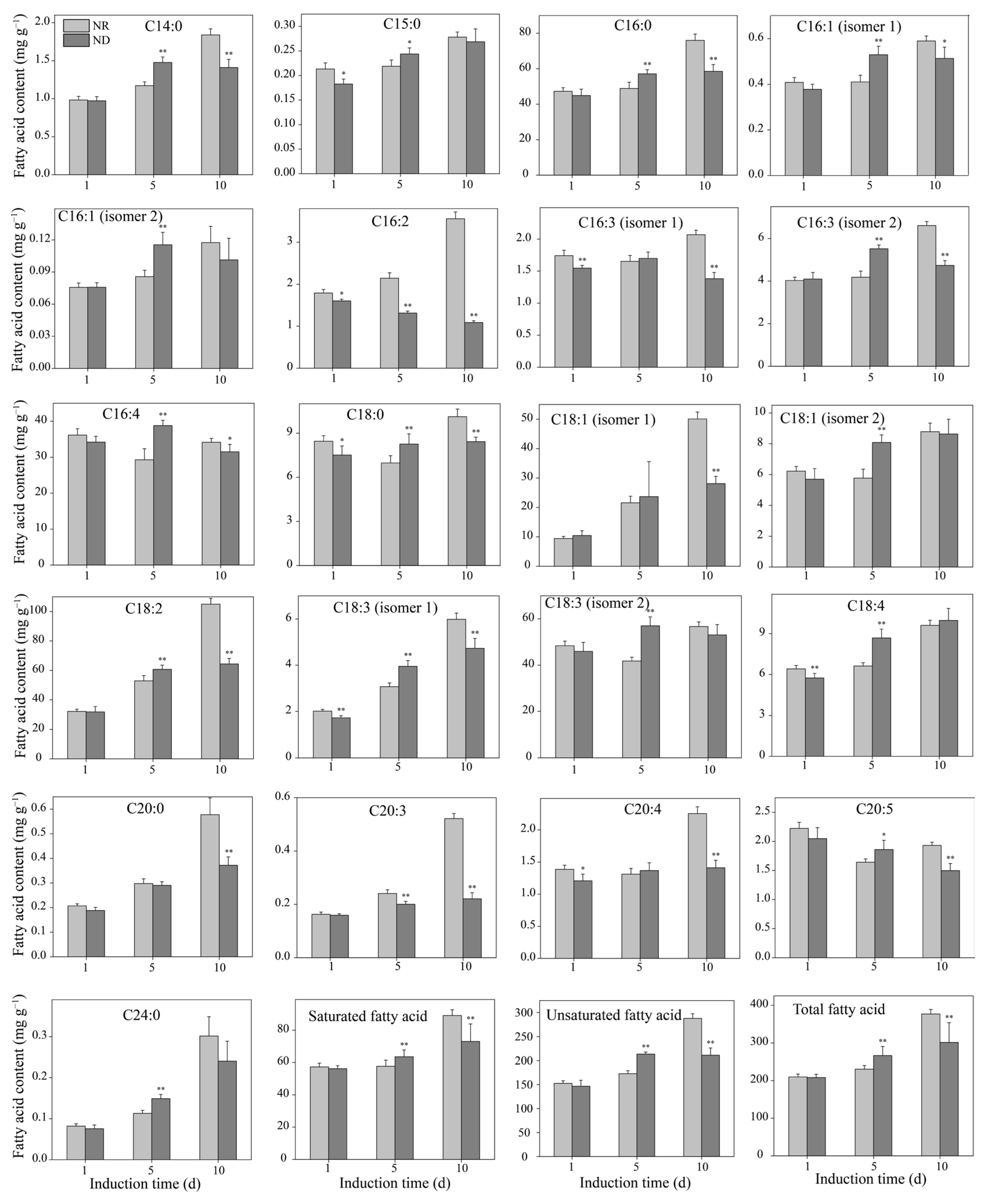
3.5. Effect of Nitrogen-Deprivation on Free Fatty Acid Contents
3.6. Comparative Transcriptome Analysis
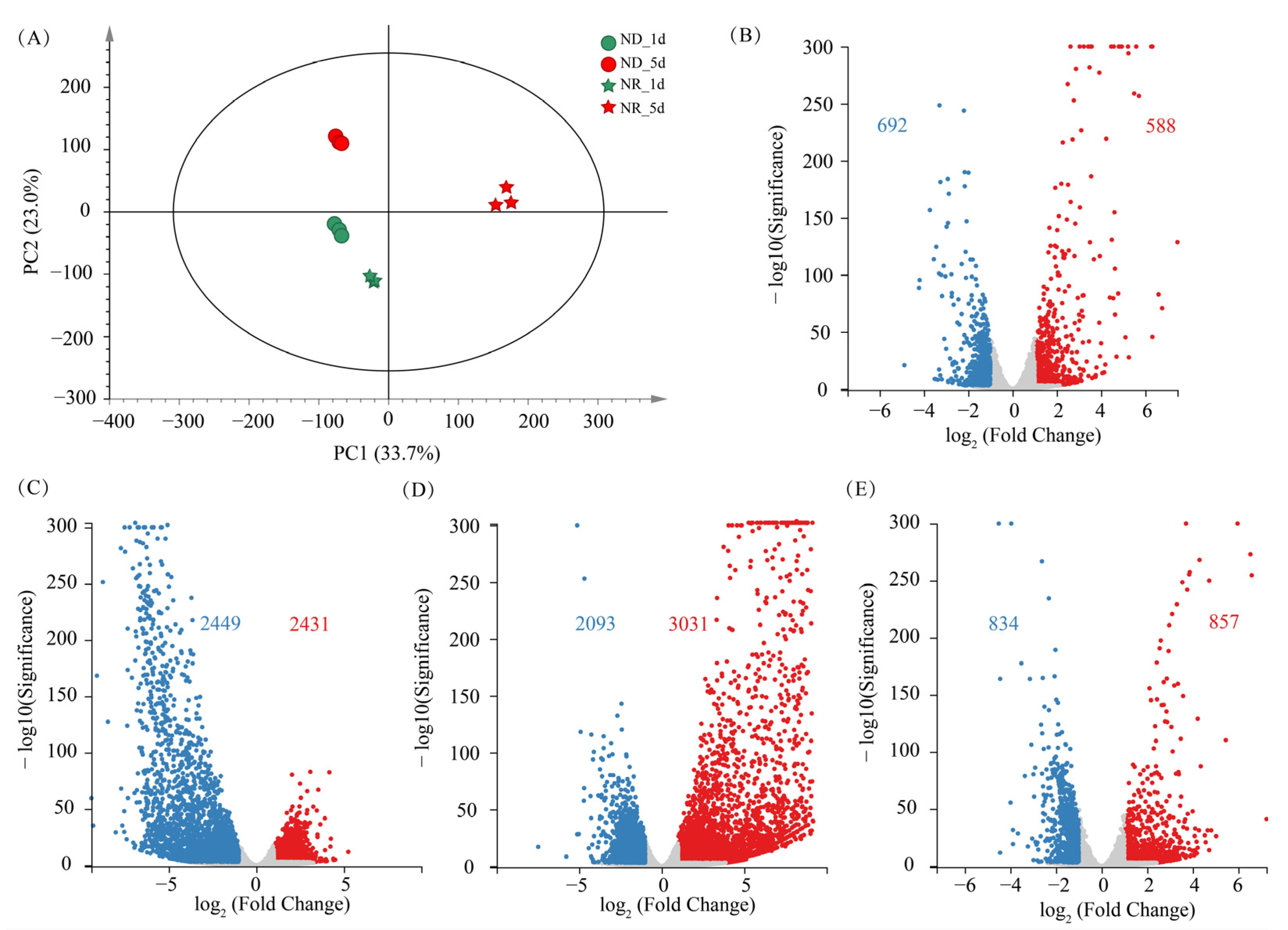
3.6.1. Overview of the Transcriptomic Difference
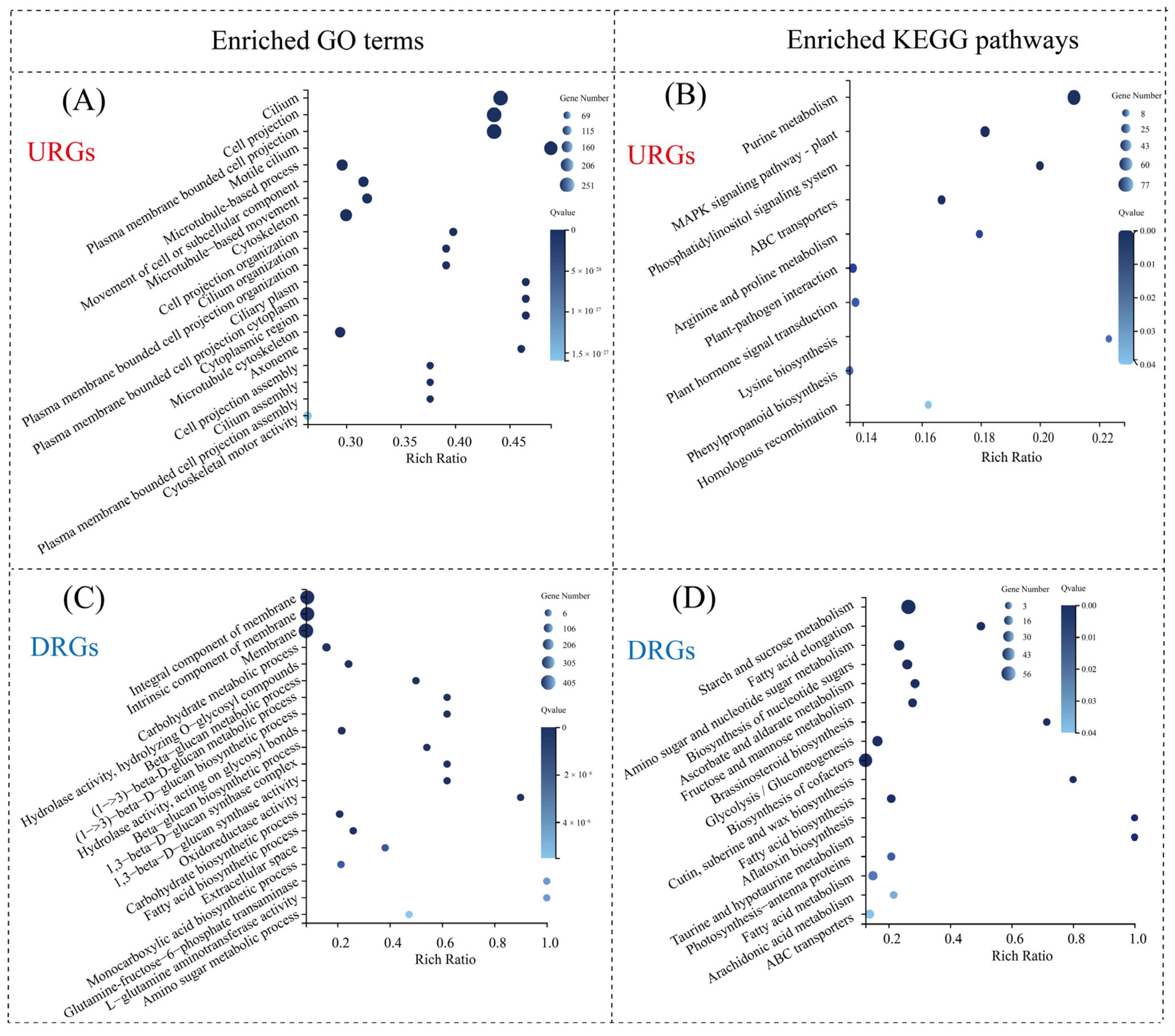
3.6.2. GO and KEGG Pathway Enrichment Analysis
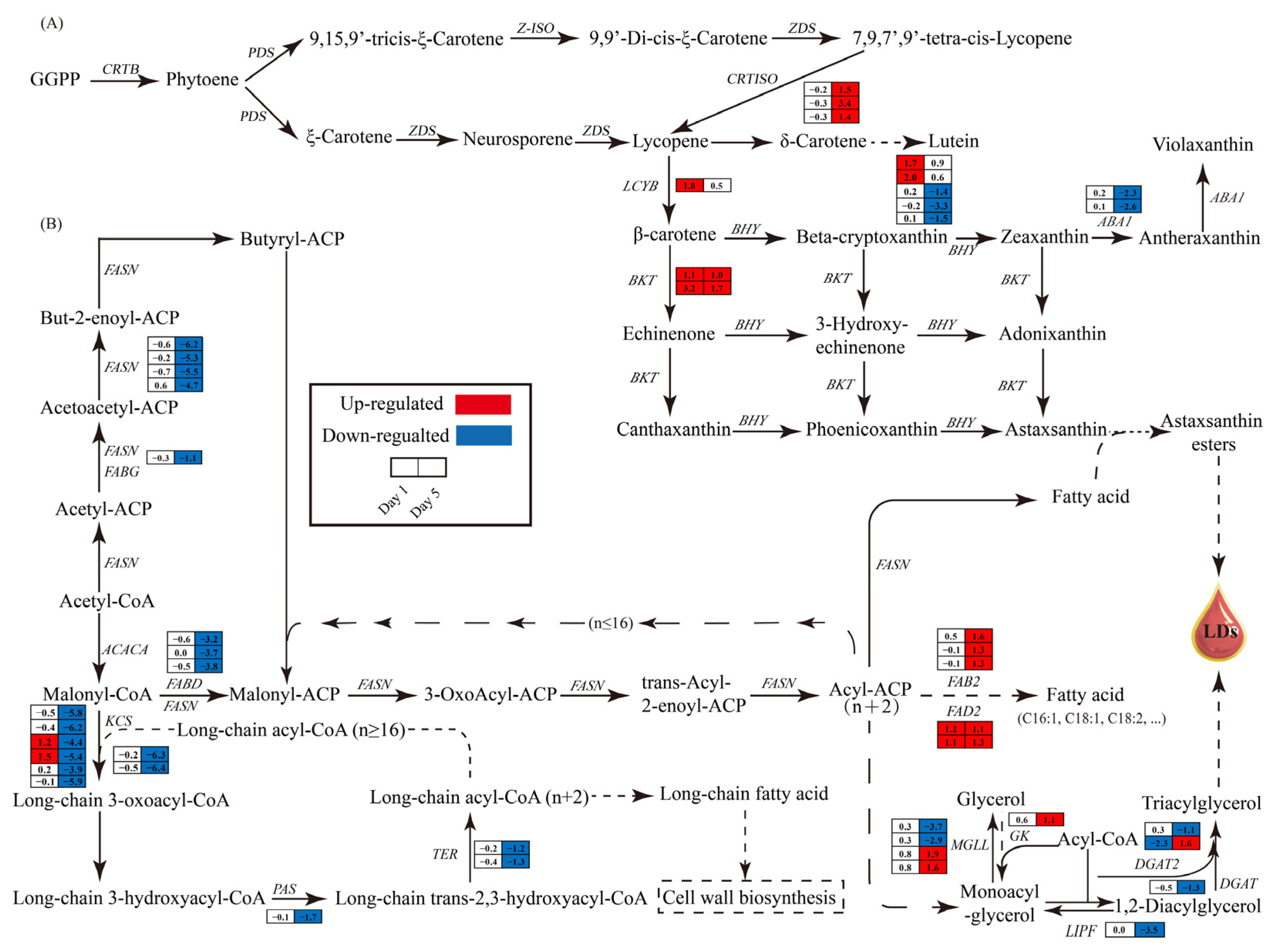
3.6.3. Impact of Nitrogen Deprivation on Astaxanthin Synthesis Pathway in H. pluvialis
3.6.4. Impact of Nitrogen Deprivation on Lipid Synthesis Pathway
3.6.5. Impact of Nitrogen Deprivation on Expression of Photosynthesis-Related Genes
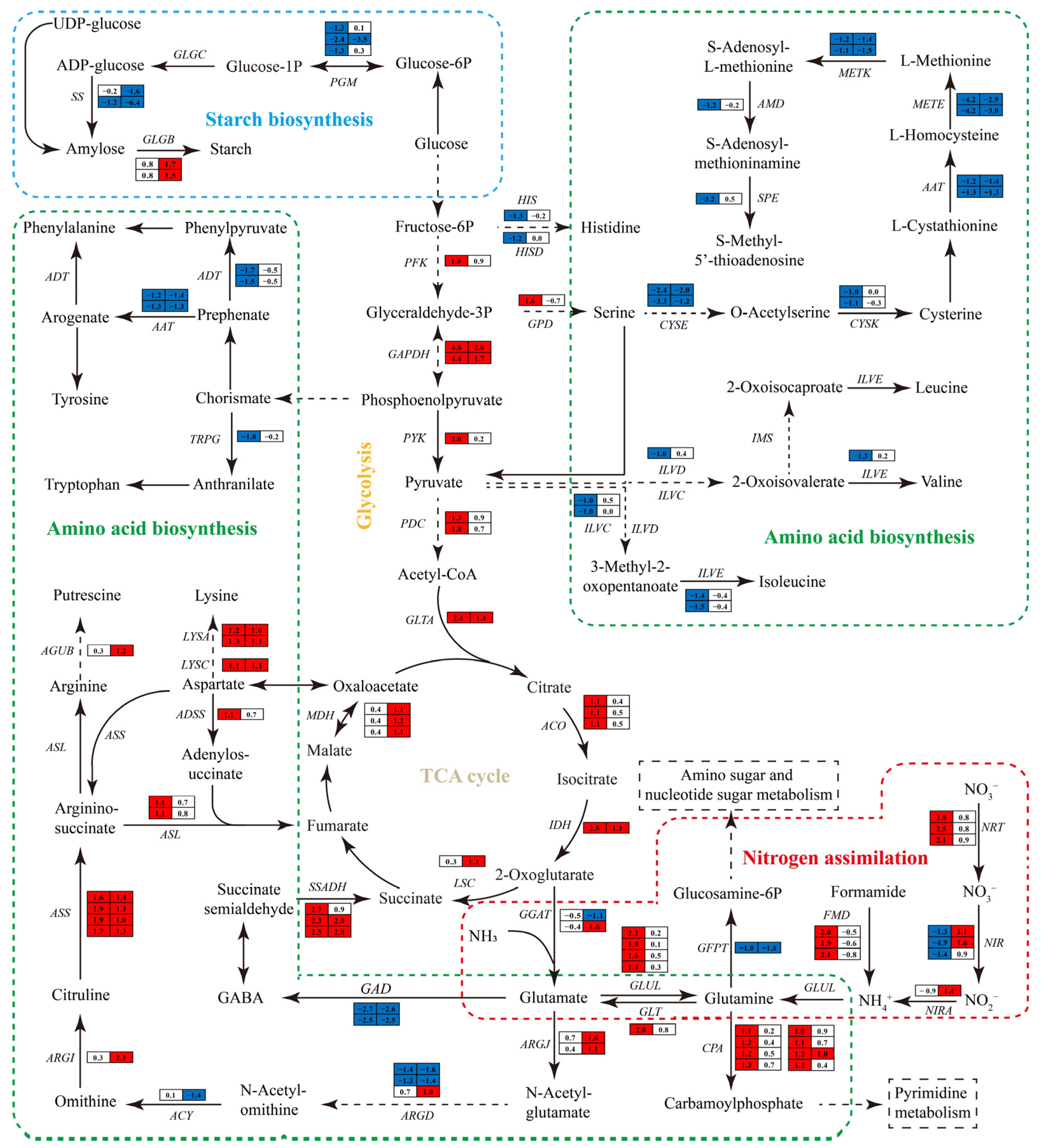
3.6.6. Impact of Nitrogen Deprivation on Sugar Metabolism Pathway
3.6.7. Impact of Nitrogen Deprivation on Nitrogen Metabolism Pathways
3.6.8. Impact of Nitrogen Deprivation on Ribosome Biogenesis and Ubiquitin–Proteasome Degradation Pathway
3.6.9. Impact of Nitrogen Deprivation on Cell Wall Biosynthesis-Related Genes
3.6.10. Impact of Nitrogen Deprivation on Movement-Related Genes in H. pluvialis
3.6.11. Impact of Nitrogen Deprivation on ROS Scavenging-Related Genes in H. pluvialis
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ariyadasa, T.U.; Thevarajah, B.; Anthonio, R.A.D.P.; Nimarshana, P.H.V.; Wasath, W.A.J. From Present to Prosperity: Assessing the Current Status and Envisioning Opportunities in the Industrial-Scale Cultivation of Haematococcus pluvialis for Astaxanthin Production. Phytochem. Rev. 2024, 23, 749–779. [Google Scholar] [CrossRef]
- Villaró, S.; Ciardi, M.; Morillas-España, A.; Sánchez-Zurano, A.; Acién-Fernández, G.; Lafarga, T. Microalgae Derived Astaxanthin: Research and Consumer Trends and Industrial Use as Food. Foods 2021, 10, 2303. [Google Scholar] [CrossRef] [PubMed]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic Astaxanthin Is Significantly Inferior to Algal-Based Astaxanthin as an Antioxidant and May Not Be Suitable as a Human Nutraceutical Supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- Debnath, T.; Bandyopadhyay, T.K.; Vanitha, K.; Bobby, M.N.; Nath Tiwari, O.; Bhunia, B.; Muthuraj, M. Astaxanthin from Microalgae: A Review on Structure, Biosynthesis, Production Strategies and Application. Food Res. Int. 2024, 176, 113841. [Google Scholar] [CrossRef]
- Sun, J.; Yan, J.; Dong, H.; Gao, K.; Yu, K.; He, C.; Mao, X. Astaxanthin with Different Configurations: Sources, Activity, Post Modification, and Application in Foods. Curr. Opin. Food Sci. 2023, 49, 100955. [Google Scholar] [CrossRef]
- Nishida, Y.; Berg, P.C.; Shakersain, B.; Hecht, K.; Takikawa, A.; Tao, R.; Kakuta, Y.; Uragami, C.; Hashimoto, H.; Misawa, N.; et al. Astaxanthin: Past, Present, and Future. Mar. Drugs 2023, 21, 514. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A Review on Haematococcus pluvialis Bioprocess Optimization of Green and Red Stage Culture Conditions for the Production of Natural Astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef]
- Hu, C.; Cui, D.; Sun, X.; Shi, J.; Song, L.; Li, Y.; Xu, N. Transcriptomic Analysis Unveils Survival Strategies of Autotrophic Haematococcus pluvialis against High Light Stress. Aquaculture 2019, 513, 734430. [Google Scholar] [CrossRef]
- Cui, H.; Zhu, X.; Yu, X.; Li, S.; Wang, K.; Wei, L.; Li, R.; Qin, S. Advancements of Astaxanthin Production in Haematococcus pluvialis: Update Insight and Way Forward. Biotechnol. Adv. 2025, 79, 108519. [Google Scholar] [CrossRef]
- Du, F.; Hu, C.; Sun, X.; Xu, N. Transcriptome Analysis Reveals Pathways Responsible for the Promoting Effect of Sucrose on Astaxanthin Accumulation in Haematococcus pluvialis under High Light Condition. Aquaculture 2021, 530, 735757. [Google Scholar] [CrossRef]
- Du, F.; Hu, C.; Sun, X.; Zhang, L.; Xu, N. Transcriptome Analysis Reveals the Promoting Effect of Trisodium Citrate on Astaxanthin Accumulation in Haematococcus pluvialis under High Light Condition. Aquaculture 2021, 543, 736978. [Google Scholar] [CrossRef]
- Hu, C.; Cui, D.; Sun, X.; Shi, J.; Xu, N. Primary Metabolism Is Associated with the Astaxanthin Biosynthesis in the Green Algae Haematococcus pluvialis under Light Stress. Algal Res. 2020, 46, 101768. [Google Scholar] [CrossRef]
- Hagen, C.; Siegmund, S.; Braune, W. Ultrastructural and Chemical Changes in the Cell Wall of Haematococcus pluvialis (Volvocales, Chlorophyta) during Aplanospore Formation. Eur. J. Phycol. 2002, 37, 217–226. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a Green Alga Red: Engineering Astaxanthin Biosynthesis by Intragenic Pseudogene Revival in Chlamydomonas reinhardtii. Plant Biotechnol. J. 2020, 18, 2053–2067. [Google Scholar] [CrossRef] [PubMed]
- Praveenkumar, R.; Lee, K.; Lee, J.; Oh, Y.-K. Breaking Dormancy: An Energy-Efficient Means of Recovering Astaxanthin from Microalgae. Green Chem. 2015, 17, 1226–1234. [Google Scholar] [CrossRef]
- Choubert, G.; Mendes-Pinto, M.M.; Morais, R. Pigmenting Efficacy of Astaxanthin Fed to Rainbow Trout Oncorhynchus mykiss: Effect of Dietary Astaxanthin and Lipid Sources. Aquaculture 2006, 257, 429–436. [Google Scholar] [CrossRef]
- Hart, B.; Colombo, S.M. Effects of a Novel Weakened Whole-cell form of Haematococcus pluvialis on Flesh Pigmentation of Rainbow Trout (Oncorhynchus mykiss) When Compared to Synthetic Astaxanthin. Aquac. Res. 2022, 53, 2408–2419. [Google Scholar] [CrossRef]
- Chen, F.; Xu, N.; Liu, K.; Lv, R.; Shi, J.; Liu, J.; Sun, X.; Hu, C. Increasing Production and Bio-Accessibility of Natural Astaxanthin in Haematococcus pluvialis by Screening and Culturing Red Motile Cells under High Light Condition. Bioresour. Technol. 2022, 364, 128067. [Google Scholar] [CrossRef]
- Xing, H.; Sun, X.; Xu, N.; Su, X.; Qin, Y.; Zhang, L.; Liu, K.; Li, M.; Hu, C. The Combination of Uridine and Nitrogen-Deprivation Promotes the Efficient Formation of Astaxanthin-Rich Motile Cells in Haematococcus pluvialis. Bioresour. Technol. 2023, 393, 130150. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, T.; Yao, S.; Hu, C.; Xing, H.; Liu, K.; Sun, X.; Xu, N. Enhancement of Astaxanthin Production, Recovery, and Bio-Accessibility in Haematococcus pluvialis through Taurine-Mediated Inhibition of Secondary Cell Wall Formation under High Light Conditions. Bioresour. Technol. 2023, 389, 129802. [Google Scholar] [CrossRef]
- Jiao, X.; Lyu, Y.; Wu, X.; Li, H.; Cheng, L.; Zhang, C.; Yuan, L.; Jiang, R.; Jiang, B.; Rengel, Z.; et al. Grain Production versus Resource and Environmental Costs: Towards Increasing Sustainability of Nutrient Use in China. J. Exp. Bot. 2016, 67, 4935–4949. [Google Scholar] [CrossRef] [PubMed]
- Poitout, A.; Crabos, A.; Petřík, I.; Novák, O.; Krouk, G.; Lacombe, B.; Ruffel, S. Responses to Systemic Nitrogen Signaling in Arabidopsis Roots Involve Trans-Zeatin in Shoots. Plant Cell 2018, 30, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Tohge, T.; Ishizaki, K.; Leaver, C.J.; Fernie, A.R. Protein Degradation—An Alternative Respiratory Substrate for Stressed Plants. Trends Plant Sci. 2011, 16, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Asad, M.A.U.; Guan, X.; Zhou, L.; Qian, Z.; Yan, Z.; Cheng, F. Involvement of Plant Signaling Network and Cell Metabolic Homeostasis in Nitrogen Deficiency-Induced Early Leaf Senescence. Plant Sci. 2023, 336, 111855. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Chen, Y. The Physiological Response of Photosynthesis to Nitrogen Deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef]
- Fernandes, J.C.; García-Angulo, P.; Goulao, L.F.; Acebes, J.L.; Amâncio, S. Mineral Stress Affects the Cell Wall Composition of Grapevine (Vitis vinifera L.) Callus. Plant Sci. 2013, 205–206, 111–120. [Google Scholar] [CrossRef]
- Rivai, R.R.; Miyamoto, T.; Awano, T.; Takada, R.; Tobimatsu, Y.; Umezawa, T.; Kobayashi, M. Nitrogen Deficiency Results in Changes to Cell Wall Composition of Sorghum Seedlings. Sci. Rep. 2021, 11, 23309. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Han, D.; Sommerfeld, M.; Lu, Y.; Chen, F.; Hu, Q. Molecular Mechanisms of the Coordination between Astaxanthin and Fatty Acid Biosynthesis in Haematococcus pluvialis (Chlorophyceae). Plant J. 2015, 81, 95–107. [Google Scholar] [CrossRef]
- Liang, C.; Zhai, Y.; Xu, D.; Ye, N.; Zhang, X.; Wang, Y.; Zhang, W.; Yu, J. Correlation between Lipid and Carotenoid Synthesis and Photosynthetic Capacity in Haematococcus pluvialis Grown under High Light and Nitrogen Deprivation Stress. Grasas Aceites 2015, 66, e077. [Google Scholar] [CrossRef]
- Zarei, Z.; Zamani, H. Biorefinery Potential of Microalga Haematococcus pluvialis to Produce Astaxanthin and Biodiesel under Nitrogen Deprivation. BioEnergy Res. 2023, 16, 1777–1788. [Google Scholar] [CrossRef]
- Samhat, K.; Kazbar, A.; Takache, H.; Ismail, A.; Pruvost, J. Influence of Light Absorption Rate on the Astaxanthin Production by the Microalga Haematococcus pluvialis during Nitrogen Starvation. Bioresour. Bioprocess 2023, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hou, Y.; Chai, W.; Liu, Z.; Wang, X.; He, C.; Hu, Z.; Chen, S.; Wang, W.; Chen, F. Transcriptome Analysis of Haematococcus pluvialis of Multiple Defensive Systems against Nitrogen Starvation. Enzyme Microb. Technol. 2020, 134, 109487. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.; McDougall, G.; Campbell, R.; Stanley, M.; Day, J. Media Screening for Obtaining Haematococcus pluvialis Red Motile Macrozooids Rich in Astaxanthin and Fatty Acids. Biology 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cai, M.; Wu, Y.; Lian, Q.; Qian, Z.; Luo, J.; Zhang, Y.; Zhang, N.; Li, C.; Huang, X. Effects of Nitrogen and Light Intensity on the Astaxanthin Accumulation in Motile Cells of Haematococcus pluvialis. Front. Mar. Sci. 2022, 9, 909237. [Google Scholar] [CrossRef]
- Li, Y.; Miao, F.; Geng, Y.; Lu, D.; Zhang, C.; Zeng, M. Accurate Quantification of Astaxanthin from Haematococcus Crude Extract Spectrophotometrically. Chin. J. Oceanol. Limnol. 2012, 30, 627–637. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, M.; Lin, J.; Zhou, S.; Sun, T.; Xu, N. Darkness and Low Nighttime Temperature Modulate the Growth and Photosynthetic Performance of Ulva prolifera under Lower Salinity. Mar. Pollut. Bull. 2019, 146, 85–91. [Google Scholar] [CrossRef]
- Liu, K.; Lv, R.; Xing, H.; Sun, X.; Xu, N.; Hu, C.; Zhang, L. Xanthosine-Mediated Modulation of Haematococcus pluvialis Growth and Stress Response under High Light: Insights from Physiological and Transcriptome Analysis. Aquaculture 2025, 599, 742156. [Google Scholar] [CrossRef]
- Cui, D.; Hu, C.; Zou, Z.; Sun, X.; Shi, J.; Xu, N. Comparative Transcriptome Analysis Unveils Mechanisms Underlying the Promoting Effect of Potassium Iodide on Astaxanthin Accumulation in Haematococcus pluvialis under High Light Stress. Aquaculture 2020, 525, 735279. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive Oxygen Species: Response of Algal Cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Chen, L.; Xu, M.; Cheng, Z.; Yang, L.-T. Effects of Nitrogen Deficiency on the Photosynthesis, Chlorophyll a Fluorescence, Antioxidant System, and Sulfur Compounds in Oryza sativa. Int. J. Mol. Sci. 2024, 25, 10409. [Google Scholar] [CrossRef]
- Chekanov, K.; Schastnaya, E.; Solovchenko, A.; Lobakova, E. Effects of CO2 Enrichment on Primary Photochemistry, Growth and Astaxanthin Accumulation in the Chlorophyte Haematococcus pluvialis. J. Photochem. Photobiol. B-Biol. 2017, 171, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Christa, G.; Cruz, S.; Jahns, P.; De Vries, J.; Cartaxana, P.; Esteves, A.C.; Serôdio, J.; Gould, S.B. Photoprotection in a Monophyletic Branch of Chlorophyte Algae Is Independent of Energy-dependent Quenching (qE). New Phytol. 2017, 214, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Scibilia, L. Photosynthetic Response to Nitrogen Starvation and High Light in Haematococcus pluvialis. Algal Res. 2015, 12, 170–181. [Google Scholar] [CrossRef]
- Holtin, K.; Kuehnle, M.; Rehbein, J.; Schuler, P.; Nicholson, G.; Albert, K. Determination of Astaxanthin and Astaxanthin Esters in the Microalgae Haematococcus pluvialis by LC-(APCI)MS and Characterization of Predominant Carotenoid Isomers by NMR Spectroscopy. Anal. Bioanal. Chem. 2009, 395, 1613–1622. [Google Scholar] [CrossRef]
- Yuan, J.-P.; Chen, F. Purification of Trans-Astaxanthin from a High-Yielding Astaxanthin Ester-Producing Strain of the Microalga Haematococcus pluvialis. Food Chem. 2000, 68, 443–448. [Google Scholar] [CrossRef]
- Huang, L.; Gao, B.; Wu, M.; Wang, F.; Zhang, C. Comparative Transcriptome Analysis of a Long-Time Span Two-Step Culture Process Reveals a Potential Mechanism for Astaxanthin and Biomass Hyper-Accumulation in Haematococcus pluvialis JNU35. Biotechnol. Biofuels 2019, 12, 18. [Google Scholar] [CrossRef]
- Pecoraro, A.; Pagano, M.; Russo, G.; Russo, A. Ribosome Biogenesis and Cancer: Overview on Ribosomal Proteins. Int. J. Mol. Sci. 2021, 22, 5496. [Google Scholar] [CrossRef]
- Finley, D. Recognition and Processing of Ubiquitin-Protein Conjugates by the Proteasome. Annu. Rev. Biochem. 2009, 78, 477–513. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The Ubiquitin-Proteasome Proteolytic Pathway: Destruction for the Sake of Construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Morreale, F.E.; Walden, H. Types of Ubiquitin Ligases. Cell 2016, 165, 248.e1. [Google Scholar] [CrossRef]
- Guo, C.; Mei, R.; Anwar, M.; Zhao, D.; Lan, C.; Jiang, Y.; Zhuang, J.; Wang, C.; Hu, Z. The Functionally Characterization of Putative Genes Involved in the Formation of Mannose in the Aplanospore Cell Wall of Haematococcus pluvialis (Volvocales, Chlorophyta). Metabolites 2021, 11, 725. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Chen, Y.; Yan, H.; Nahidian, B.; Hu, Q.; Han, D. High-Throughput Fluorescence-Activated Cell Sorting for Cell Wall-Deficient Microalgal Mutants Screening. Algal Res. 2020, 50, 102011. [Google Scholar] [CrossRef]
- Cuvillier, A.; Redon, F.; Antoine, J.-C.; Chardin, P.; DeVos, T.; Merlin, G. LdARL-3A, a Leishmania Promastigote-Specific ADP-Ribosylation Factor-like Protein, Is Essential for Flagellum Integrity. J. Cell Sci. 2000, 113, 2065–2074. [Google Scholar] [CrossRef]
- Jurczyk, A.; Gromley, A.; Redick, S.; Agustin, J.S.; Witman, G.; Pazour, G.J.; Peters, D.J.M.; Doxsey, S. Pericentrin Forms a Complex with Intraflagellar Transport Proteins and Polycystin-2 and Is Required for Primary Cilia Assembly. J. Cell Biol. 2004, 166, 637–643. [Google Scholar] [CrossRef]
- Silverman, M.A.; Leroux, M.R. Intraflagellar Transport and the Generation of Dynamic, Structurally and Functionally Diverse Cilia. Trends Cell Biol. 2009, 19, 306–316. [Google Scholar] [CrossRef]
- Pan, J.; Wang, Q.; Snell, W.J. An Aurora Kinase Is Essential for Flagellar Disassembly in Chlamydomonas. Dev. Cell 2004, 6, 445–451. [Google Scholar] [CrossRef]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The Evolution of Reactive Oxygen Species Metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.; Zhang, Y.; Gao, H.; Zhao, Y.; Yu, X. Chemical Inducers Regulate ROS Signalling to Stimulate Astaxanthin Production in Haematococcus pluvialis under Environmental Stresses: A Review. Trends Food Sci. Technol. 2023, 136, 181–193. [Google Scholar] [CrossRef]
- Vidhyavathi, R.; Venkatachalam, L.; Sarada, R.; Ravishankar, G.A. Regulation of Carotenoid Biosynthetic Genes Expression and Carotenoid Accumulation in the Green Alga Haematococcus pluvialis under Nutrient Stress Conditions. J. Exp. Bot. 2008, 59, 1409–1418. [Google Scholar] [CrossRef]
- Ye, S.; Huang, Y.; Ma, T.; Ma, X.; Li, R.; Shen, J.; Wen, J. BnaABF3 and BnaMYB44 Regulate the Transcription of Zeaxanthin Epoxidase Genes in Carotenoid and Abscisic Acid Biosynthesis. Plant Physiol. 2024, 195, 2372–2388. [Google Scholar] [CrossRef]
- Nguyen, T.; Xu, Y.; Abdel-Hameed, M.; Sorensen, J.L.; Singer, S.D.; Chen, G. Characterization of a Type-2 Diacylglycerol Acyltransferase from Haematococcus pluvialis Reveals Possible Allostery of the Recombinant Enzyme. Lipids 2020, 55, 425–433. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-Z.; Jiang, S.; Zhang, L.; Yu, Z.-B. Mitochondrial Electron Transport Chain, ROS Generation and Uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Nunes-Nesi, A.; Nikoloski, Z.; Sweetlove, L.J.; Fernie, A.R. Metabolic Control and Regulation of the Tricarboxylic Acid Cycle in Photosynthetic and Heterotrophic Plant Tissues. Plant Cell Environ. 2012, 35, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.C.; Kwon, H.D.; Jo, H.-S.; Choi, Y.Y.; Seok, J.-I.; Kang, Y.; Lee, D.Y.; Kim, D.-S. Explorative Study of Serum Biomarkers of Liver Failure after Liver Resection. Sci. Rep. 2020, 10, 9960. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, L.; Zhao, J.; Liu, J. Enhancement of Astaxanthin Accumulation in Haematococcus pluvialis by Exogenous Oxaloacetate Combined with Nitrogen Deficiency. Bioresour. Technol. 2022, 345, 126484. [Google Scholar] [CrossRef]
- Lu, Z.; Dai, J.; Zheng, L.; Teng, Z.; Zhang, Q.; Qiu, D.; Song, L. Disodium 2-Oxoglutarate Promotes Carbon Flux into Astaxanthin and Fatty Acid Biosynthesis Pathways in Haematococcus. Bioresour. Technol. 2020, 299, 122612. [Google Scholar] [CrossRef]
- Bhosale, P. Environmental and Cultural Stimulants in the Production of Carotenoids from Microorganisms. Appl. Microbiol. Biotechnol. 2004, 63, 351–361. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen Assimilation in Plants: Current Status and Future Prospects. J. Genet. Genomics 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, D.; Zhang, C.; Wu, H.; Li, Q.; Gu, M.; Sun, S.S.; Liu, Q.Q. A Connection between Lysine and Serotonin Metabolism in Rice Endosperm. Plant Physiol. 2018, 176, 1965–1980. [Google Scholar] [CrossRef]
- Xing, H.; Zhao, Y.; Li, T.; Han, B.; Zhao, P.; Yu, X. Enhancing Astaxanthin and Lipid Coproduction in Haematococcus pluvialis by the Combined Induction of Plant Growth Regulators and Multiple Stresses. Bioresour. Technol. 2022, 344, 126225. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Yan, H.; Zhao, L.; Li, Y.; Nahidian, B.; Zhu, M.; Hu, Q.; Han, D. Interaction between the Cell Walls of Microalgal Host and Fungal Carbohydrate-activate Enzymes Is Essential for the Pathogenic Parasitism Process. Environ. Microbiol. 2021, 23, 5114–5130. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, H.; Zhou, N.; Liu, K.; Yan, X.; Li, W.; Sun, X.; Zhang, L.; Liu, F.; Xu, N.; Hu, C. Nitrogen Deprivation Drives Red Motile Cell Formation in Haematococcus pluvialis: Physiological and Transcriptomic Insights. Metabolites 2025, 15, 388. https://doi.org/10.3390/metabo15060388
Xing H, Zhou N, Liu K, Yan X, Li W, Sun X, Zhang L, Liu F, Xu N, Hu C. Nitrogen Deprivation Drives Red Motile Cell Formation in Haematococcus pluvialis: Physiological and Transcriptomic Insights. Metabolites. 2025; 15(6):388. https://doi.org/10.3390/metabo15060388
Chicago/Turabian StyleXing, Hailiang, Na Zhou, Kai Liu, Xiaotian Yan, Wanxia Li, Xue Sun, Liuquan Zhang, Fengjie Liu, Nianjun Xu, and Chaoyang Hu. 2025. "Nitrogen Deprivation Drives Red Motile Cell Formation in Haematococcus pluvialis: Physiological and Transcriptomic Insights" Metabolites 15, no. 6: 388. https://doi.org/10.3390/metabo15060388
APA StyleXing, H., Zhou, N., Liu, K., Yan, X., Li, W., Sun, X., Zhang, L., Liu, F., Xu, N., & Hu, C. (2025). Nitrogen Deprivation Drives Red Motile Cell Formation in Haematococcus pluvialis: Physiological and Transcriptomic Insights. Metabolites, 15(6), 388. https://doi.org/10.3390/metabo15060388