
Comprehensive Analysis of Small RNA Modifications in Arabidopsis thaliana and Their Dynamics During Seed Germination


 ,
,  ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
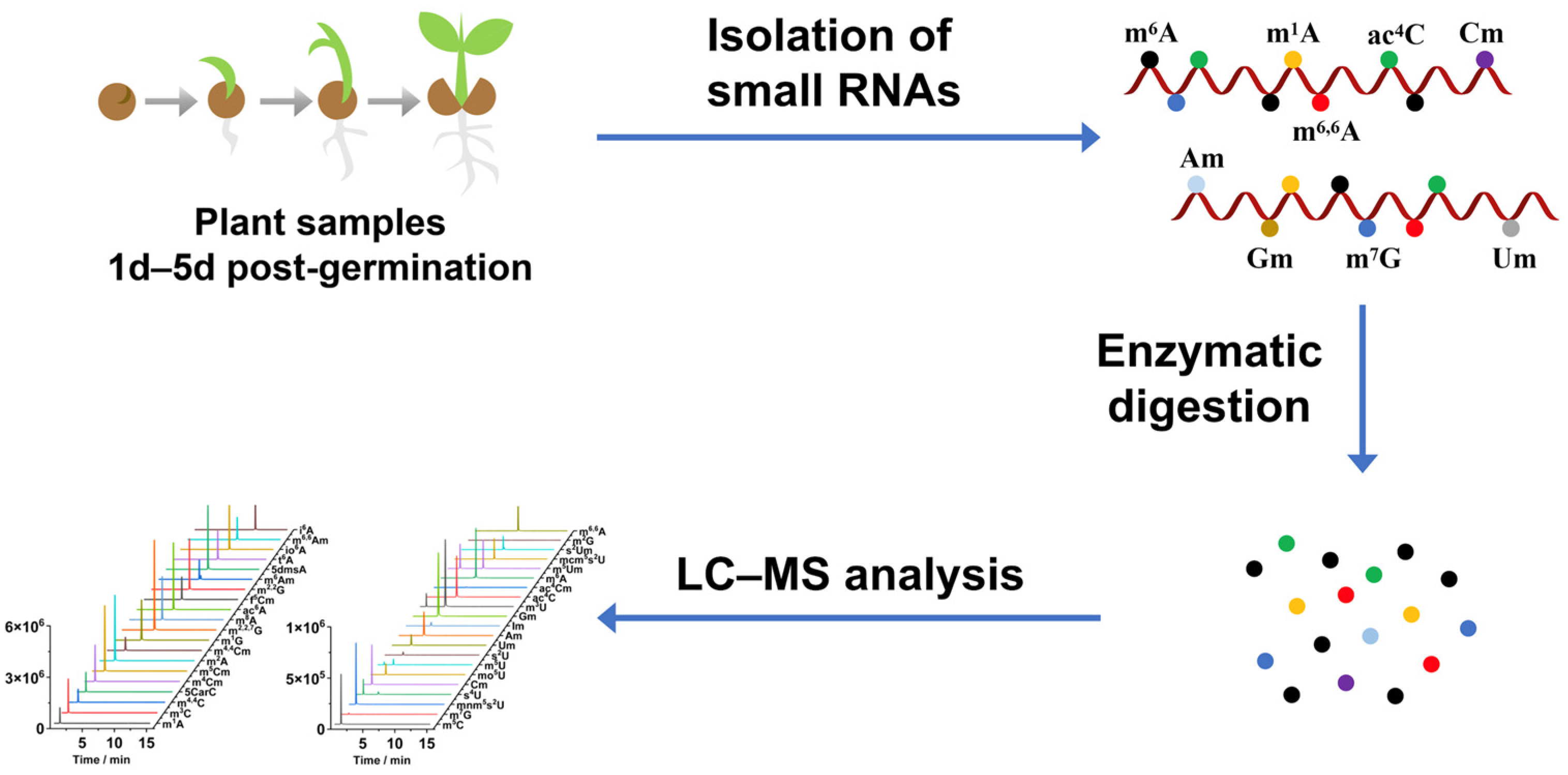
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Materials
2.3. Purification of Small RNAs
2.4. Enzymatic Digestion of Small RNAs
2.5. LC-MS/MS Analysis
2.6. High-Resolution Mass Spectrometry Analysis
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results and Discussion
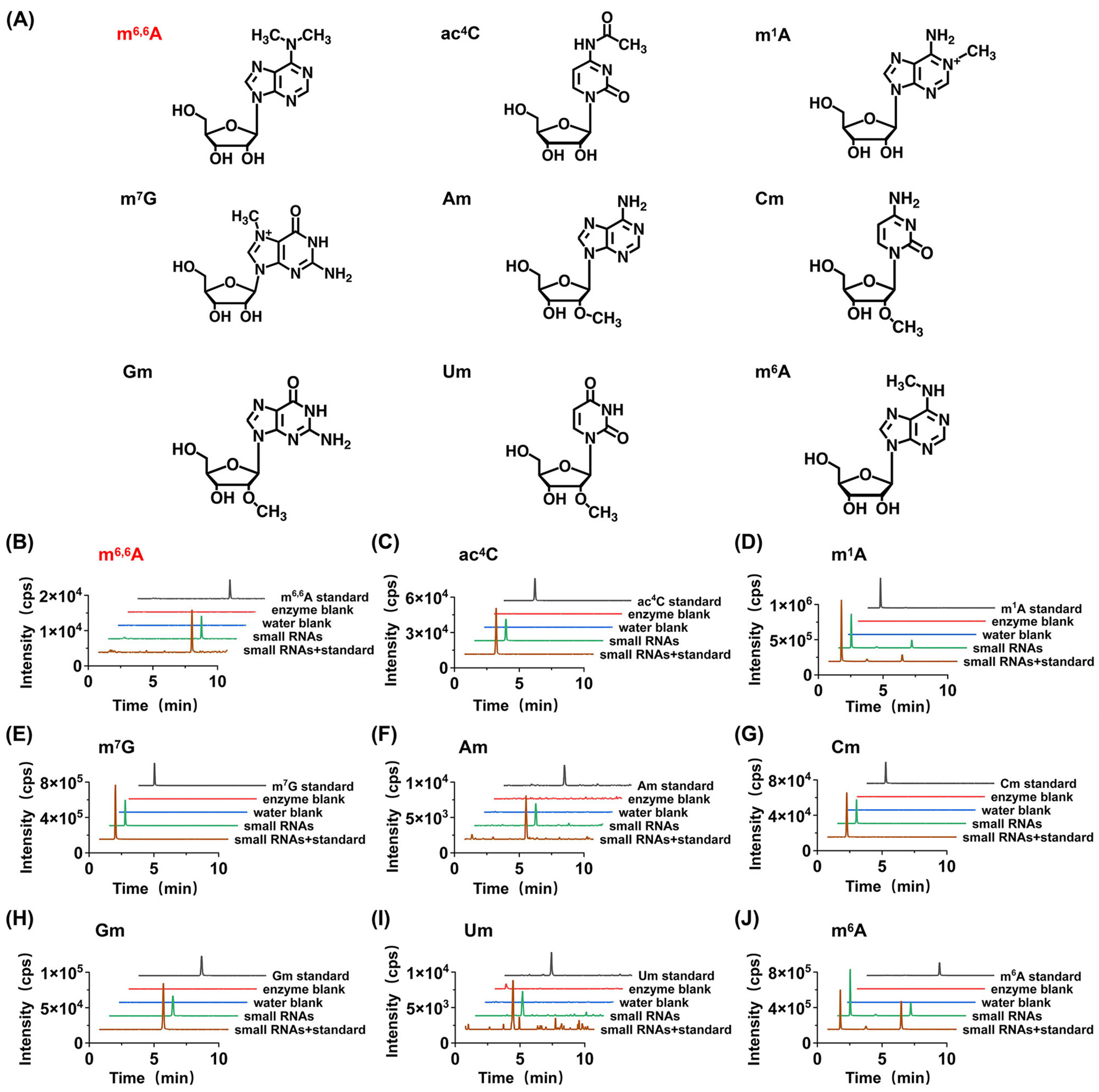
3.1. Establishment of LC-MS/MS Analytical Method
3.2. Qualitative Analysis of Modifications in Small RNA
3.3. Quantitative Analysis of Modifications in Small RNA of A. thaliana at Various Germination Stages
3.4. The Contents of 2′-O-Methylation Modification (Nm) Showed a Decreasing Trend During the Germination Process
3.5. The Contents of Methylation Modifications (m1A, m6A, and m6,6A) Also Exhibited a Decreasing Trend During the Germination Process
3.6. The Content of ac4C Gradually Decreases During Germination
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marquardt, S.; Petrillo, E.; AManavella, P. Cotranscriptional RNA processing and modification in plants. Plant Cell 2022, 35, 1654–1670. [Google Scholar] [CrossRef]
- Wen, J.; Zhu, Q.; Liu, Y.; Gou, L.-T. RNA modifications: Emerging players in the regulation of reproduction and development. Acta Biochim. Biophys. Sin. 2024, 57, 33–58. [Google Scholar] [PubMed]
- Chen, P.; Jäger, G.; Zheng, B. Transfer RNA modifications and genes for modifying enzymes in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 201. [Google Scholar] [CrossRef]
- Li, X.; Peng, J.; Yi, C. The epitranscriptome of small non-coding RNAs. Noncoding RNA Res. 2021, 6, 167–173. [Google Scholar] [CrossRef]
- Shen, L.; Liang, Z.; Wong, C.E.; Yu, H. Messenger RNA Modifications in Plants. Trends Plant Sci. 2019, 24, 328–341. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Wang, J.; Li, X.; Li, J.; Ye, K. Profiling of RNA ribose methylation in Arabidopsis thaliana. Nucleic Acids Res. 2021, 49, 4104. [Google Scholar] [CrossRef]
- Boo, S.H.; Kim, Y.K. The emerging role of RNA modifications in the regulation of mRNA stability. Exp. Mol. Med. 2020, 52, 400–408. [Google Scholar] [CrossRef]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA modification landscape in human disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef]
- Deng, W.; Babu, I.R.; Su, D.; Yin, S.; Begley, T.J.; Dedon, P.C. Trm9-Catalyzed tRNA Modifications Regulate Global Protein Expression by Codon-Biased Translation. PLoS Genet. 2015, 11, 12. [Google Scholar] [CrossRef]
- Nedialkova Danny, D.; Leidel Sebastian, A. Optimization of Codon Translation Rates via tRNA Modifications Maintains Proteome Integrity. Cell 2015, 161, 1606–1608. [Google Scholar] [CrossRef]
- Liang, Z.; Riaz, A.; Chachar, S.; Ding, Y.; Du, H.; Gu, X. Epigenetic Modifications of mRNA and DNA in Plants. Mol. Plant 2019, 13, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, R.; Coimbra, S.; Correia, S.; Canhoto, J. RNA methyltransferases in plants: Breakthroughs in function and evolution. Plant Physiol. Biochem. 2022, 194, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Wang, B.; Gao, Y.; Zhao, L.; Bi, Y.; Zhang, J.; Wang, N.; Kang, H.; Pang, J.; Liu, Y.; et al. Detailed resume of RNA m6A demethylases. Acta Pharm. Sin. B 2022, 12, 2193–2205. [Google Scholar] [CrossRef]
- Xu, T.; Wu, X.; Wong, C.E.; Fan, S.; Zhang, Y.; Zhang, S.; Liang, Z.; Yu, H.; Shen, L. FIONA1-Mediated m6A Modification Regulates the Floral Transition in Arabidopsis. Adv. Sci. 2022, 9, 2103628. [Google Scholar] [CrossRef]
- Cui, X.; Liang, Z.; Shen, L.; Zhang, Q.; Bao, S.; Geng, Y.; Zhang, B.; Leo, V.; Vardy, L.A.; Lu, T.; et al. 5-Methylcytosine RNA Methylation in Arabidopsis thaliana. Mol. Plant 2017, 10, 1387–1399. [Google Scholar] [CrossRef]
- Kumari, P.; Bhattacharjee, S.; Raman, K.V.; Tilgam, J.; Paul, K.; Senthil, K.; Baaniya, M.; Prashat, G.R.; Sreevathsa, R.; Pattanayak, D. Identification of methyltransferase and demethylase genes and their expression profiling under biotic and abiotic stress in pigeon pea (Cajanus cajan [L.] Millspaugh). Front. Plant Sci. 2025, 15, 1521758. [Google Scholar] [CrossRef]
- Susheel, S.B.; Dawid, B.; Tomasz, G.; Bodi, Z.; Yu, X.; Anderson, S.J.; Szewc, L.; Bajczyk, M.; Dolata, J.; Grzelak, N.; et al. mRNA adenosine methylase (MTA) deposits m6A on pri-miRNAs to modulate miRNA biogenesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2020, 117, 21785–21795. [Google Scholar]
- Wang, Y.; Pang, C.; Li, X.; Hu, Z.; Lv, Z.; Zheng, B.; Chen, P. Identification of tRNA nucleoside modification genes critical for stress response and development in rice and Arabidopsis. BMC Plant Biol. 2017, 17, 261. [Google Scholar] [CrossRef]
- Yu, B.; Yang, Z.Y.; Li, J.J.; Minakhina, S.; Yang, M.; Padgett, R.W.; Steward, R.; Chen, X. Methylation as a Crucial Step in Plant microRNA Biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J.; Song, P.; Zhang, W.; Lu, Q.; Yu, Q.; Jia, G. FIONA1 is an RNA N6-methyladenosine methyltransferase affecting Arabidopsis photomorphogenesis and flowering. Genome Biol. 2022, 23, 40. [Google Scholar] [CrossRef]
- Yu, B.; Bi, L.; Zhai, J.; Agarwal, M.; Li, S.; Wu, Q.; Ding, S.-W.; Meyers, B.C.; Vaucheret, H.; Chen, X. siRNAs compete with miRNAs for methylation by HEN1 in Arabidopsis. Nucleic Acids Res. 2010, 38, 5844–5850. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, S.; Yu, L.; Xiao, Y.; Zhang, S.; Wang, X.; Xu, Y.; Yu, H.; Li, Y.; Yang, J.; et al. RNA demethylation increases the yield and biomass of rice and potato plants in field trials. Nat. Biotechnol. 2021, 39, 1581–1588. [Google Scholar] [CrossRef]
- Shi, K.; Hong, X.; Xu, D.; Pan, T.; Wang, H.; Feng, H.; Guo, C.; Pan, Y. Analysis of RNA modifications in peripheral white blood cells from breast cancer patients by mass spectrometry. Chin. Chem. Lett. 2024, 36, 110079. [Google Scholar] [CrossRef]
- Tang, X.M.; Ye, T.T.; You, X.J.; Yin, X.-M.; Ding, J.-H.; Shao, W.-X.; Chen, M.-Y.; Yuan, B.-F.; Feng, Y.-Q. Mass spectrometry profiling analysis enables the identification of new modifications in ribosomal RNA. Chin. Chem. Lett. 2022, 34, 107531. [Google Scholar] [CrossRef]
- Xiong, J.; Wu, J.; Liu, Y.; Feng, Y.-J.; Yuan, B.-F. Quantification and mapping of RNA modifications. Trends Anal. Chem. 2024, 172, 117606. [Google Scholar] [CrossRef]
- Chen, M.Y.; Zhu, G.; Ke-Ke, C.; Jiang-Hui, D.; Jin-Gang, H.; Jun, X.; Jia-Le, L.; Jie, W.; Bi-Feng, Y.; Yu-Qi, F. Adolescent alcohol exposure alters DNA and RNA modifications in peripheral blood by liquid chromatography-tandem mass spectrometry analysis. Chin. Chem. Lett. 2021, 33, 2086–2090. [Google Scholar] [CrossRef]
- Li, Y.; Yin, M.; Wang, J.; Zhao, X.; Xu, J.; Wang, W.; Fu, B. Epitranscriptome profiles reveal participation of the RNA methyltransferase gene OsMTA1 in rice seed germination and salt stress response. BMC Plant Biol. 2025, 25, 115. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Liu, B.; Xu, Z.-Y. Dynamic regulation of DNA methylation and histone modifications in response to abiotic stresses in plants. J. Integr. Plant Biol. 2022, 64, 2252–2274. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Ye, T.; Zhao, S.; Liu, Z.; Feng, Y.Q.; Wu, Y. Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression. Plant J. 2011, 68, 249–261. [Google Scholar] [CrossRef]
- Yin, X.; Bai, Y.-L.; Ye, T.; Yu, M.; Wu, Y.; Feng, Y.-Q. Cinnamoyl coA: NADP oxidoreductase-like 1 regulates abscisic acid response by modulating phaseic acid homeostasis in Arabidopsis thaliana. J. Exp. Bot. 2021, 73, 860–872. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Dou, Y.; Chen, L.; Wang, J.; Jiang, N.; Guo, C.; Yao, Q.; Wang, C.; Liu, L.; et al. Degradation of unmethylated miRNA/miRNA*s by a DEDDy-type 3′ to 5′ exoribonuclease Atrimmer 2 in Arabidopsis [Plant Biology]. Proc. Natl. Acad. Sci. USA 2018, 115, 6659–6667. [Google Scholar]
- Ren, G.; Chen, X.; Yu, B. Uridylation of miRNAs by HEN1 SUPPRESSOR1 in Arabidopsis. Curr. Biol. 2012, 22, 695–700. [Google Scholar] [CrossRef]
- Yang, W.; Choi, M.-H.; Noh, B.; Noh, Y.-S. De Novo Shoot Regeneration Controlled by HEN1 and TCP3/4 in Arabidopsis. Plant Cell Physiol. 2020, 61, 1600–1613. [Google Scholar] [CrossRef]
- Lange, H.; Zuber, H.; Sement, F.M.; Chicher, J.; Kuhn, L.; Hammann, P.; Brunaud, V.; Bérard, C.; Bouteiller, N.; Balzergue, S.; et al. The RNA Helicases AtMTR4 and HEN2 Target Specific Subsets of Nuclear Transcripts for Degradation by the Nuclear Exosome in Arabidopsis thaliana. PLoS Genet. 2014, 10, e1004564. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Asgari, M.; Mermet, S.; Mishra, P.; Kindgren, P. The nuclear exosome subunit HEN2 acts independently of the core exosome to assist transcription in Arabidopsis. Plant Physiol. 2024, 196, 2625–2637. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. microRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 824240. [Google Scholar]
- Reichel, M.; Köster, T.; Staiger, D. Marking RNA: m6A writers, readers, and functions in Arabidopsis. J. Mol. Cell Biol. 2019, 11, 899–910. [Google Scholar] [CrossRef]
- Jin, H.; Huo, C.; Zhou, T.; Xie, S. m1A RNA Modification in Gene Expression Regulation. Genes 2022, 13, 910. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Jia, P.; Xin, P.; Chu, J.; Shi, D.-Q.; Yang, W.-C. The Arabidopsis TRM61/TRM6 complex is a bona fide tRNA N1-methyladenosine methyltransferase. J. Exp. Bot. 2020, 71, 3024–3036. [Google Scholar] [CrossRef]
- Nachtergaele, S.; He, C. The emerging biology of RNA post-transcriptional modifications. RNA Biol. 2017, 14, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shen, S.; Wu, P.; Li, F.; Liu, X.; Wang, C.; Gong, Q.; Wu, J.; Yao, X.; Zhang, H.; et al. Structural insights into dimethylation of 12S rRNA by TFB1M: Indispensable role in translation of mitochondrial genes and mitochondrial function. Nucleic Acids Res. 2019, 47, 7648–7665. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, H.; Wang, F.; Liu, X.; Sun, Y.; Zhao, J.; Zhu, C.; Gan, L.; Yu, J.; Witte, C.-P.; et al. N4-acetylation of cytidine in (m)RNA plays essential roles in plants. Plant Cell 2023, 35, 3739–3756. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, D.; Cai, L.; Zhou, Q.; Liu, C.; Lin, J.; Li, Y.; Zhao, X.; Li, L.; Liu, X.; et al. Transcriptome-wide profiling of RNA N4-cytidine acetylation in Arabidopsis thaliana and Oryza sativa. Mol. Plant 2023, 16, 1082–1098. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.-C.; Men, M.; Cui, X.-J.; Zeng, R.-J.; Gu, S.-Y.; Feng, T.; Zeng, C.; Ye, T.; Xiong, J.; Yuan, B.-F.; et al. Comprehensive Analysis of Small RNA Modifications in Arabidopsis thaliana and Their Dynamics During Seed Germination. Metabolites 2025, 15, 319. https://doi.org/10.3390/metabo15050319
Jiang L-C, Men M, Cui X-J, Zeng R-J, Gu S-Y, Feng T, Zeng C, Ye T, Xiong J, Yuan B-F, et al. Comprehensive Analysis of Small RNA Modifications in Arabidopsis thaliana and Their Dynamics During Seed Germination. Metabolites. 2025; 15(5):319. https://doi.org/10.3390/metabo15050319
Chicago/Turabian StyleJiang, Liu-Cheng, Meng Men, Xuan-Jun Cui, Ren-Jie Zeng, Shu-Yi Gu, Tian Feng, Chen Zeng, Tiantian Ye, Jun Xiong, Bi-Feng Yuan, and et al. 2025. "Comprehensive Analysis of Small RNA Modifications in Arabidopsis thaliana and Their Dynamics During Seed Germination" Metabolites 15, no. 5: 319. https://doi.org/10.3390/metabo15050319
APA StyleJiang, L.-C., Men, M., Cui, X.-J., Zeng, R.-J., Gu, S.-Y., Feng, T., Zeng, C., Ye, T., Xiong, J., Yuan, B.-F., & Feng, Y.-Q. (2025). Comprehensive Analysis of Small RNA Modifications in Arabidopsis thaliana and Their Dynamics During Seed Germination. Metabolites, 15(5), 319. https://doi.org/10.3390/metabo15050319