

Molecular Mechanism of Cuscuta Haustorium Specialization Inferences from Transcriptome and Metabolome Analysis

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. LC-MS Analysis
2.3. Total RNA Extraction and Transcriptomic Analysis
3. Results
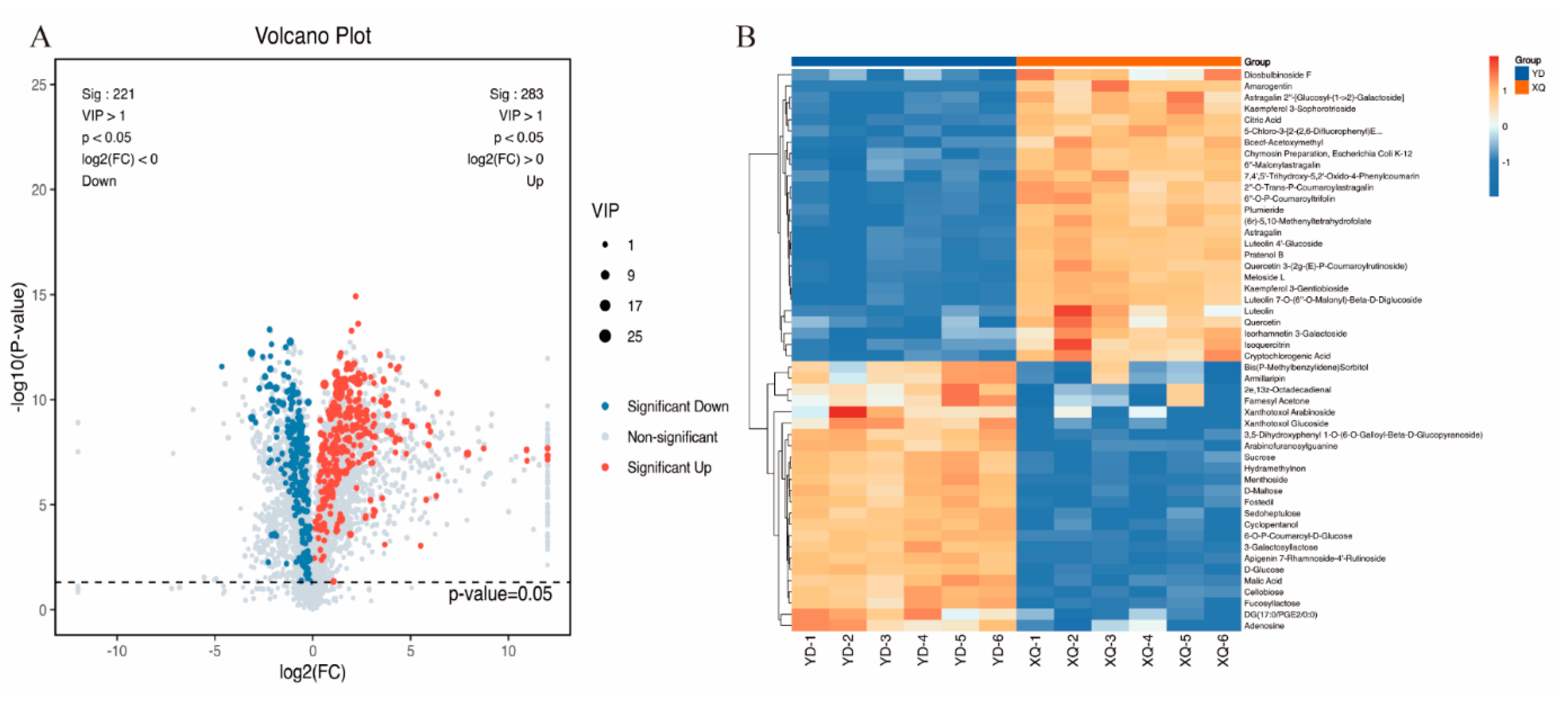
3.1. LC-MS Metabolome Analysis of C. australis Aspirator and Distal Diameter Segments
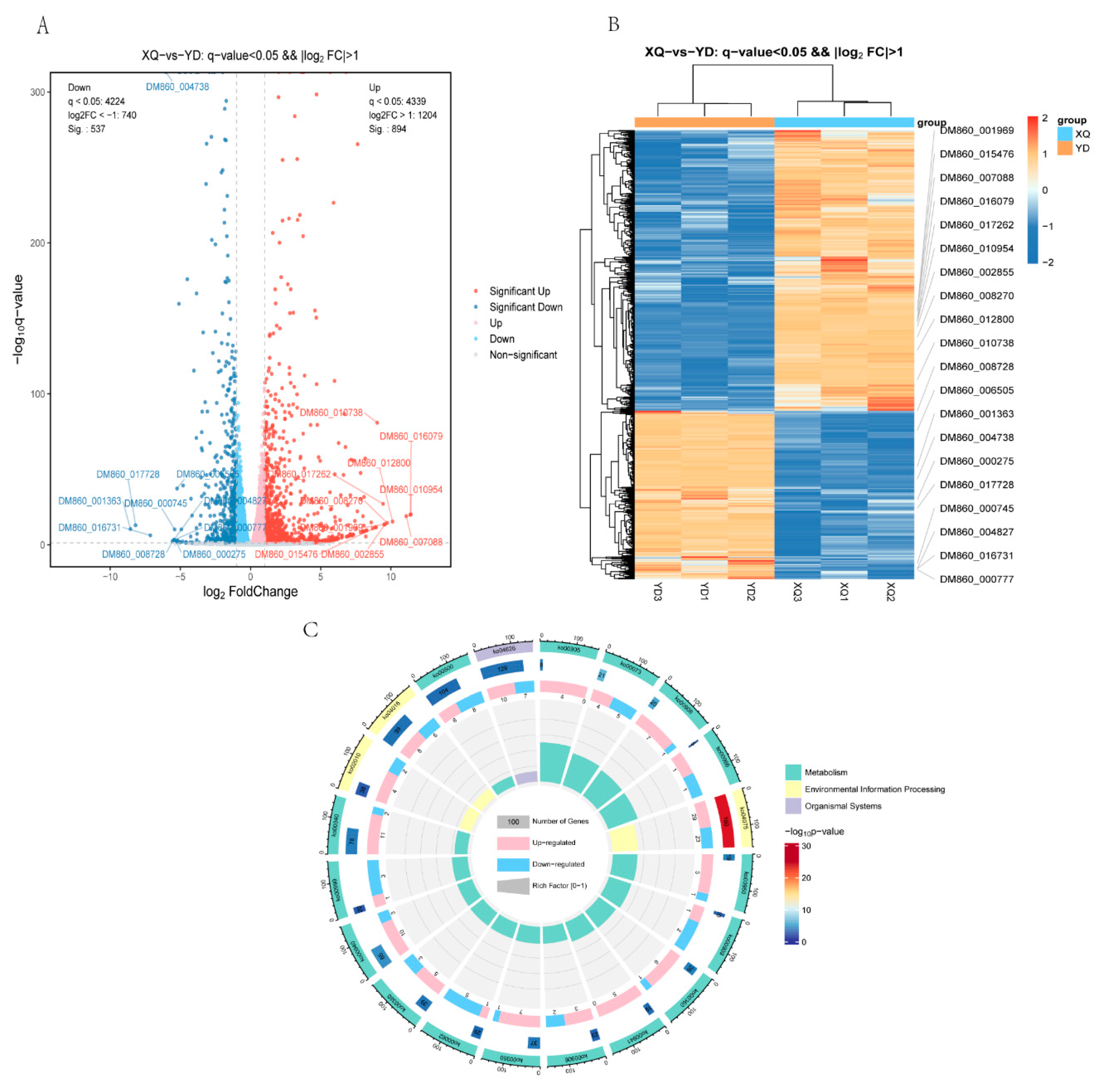
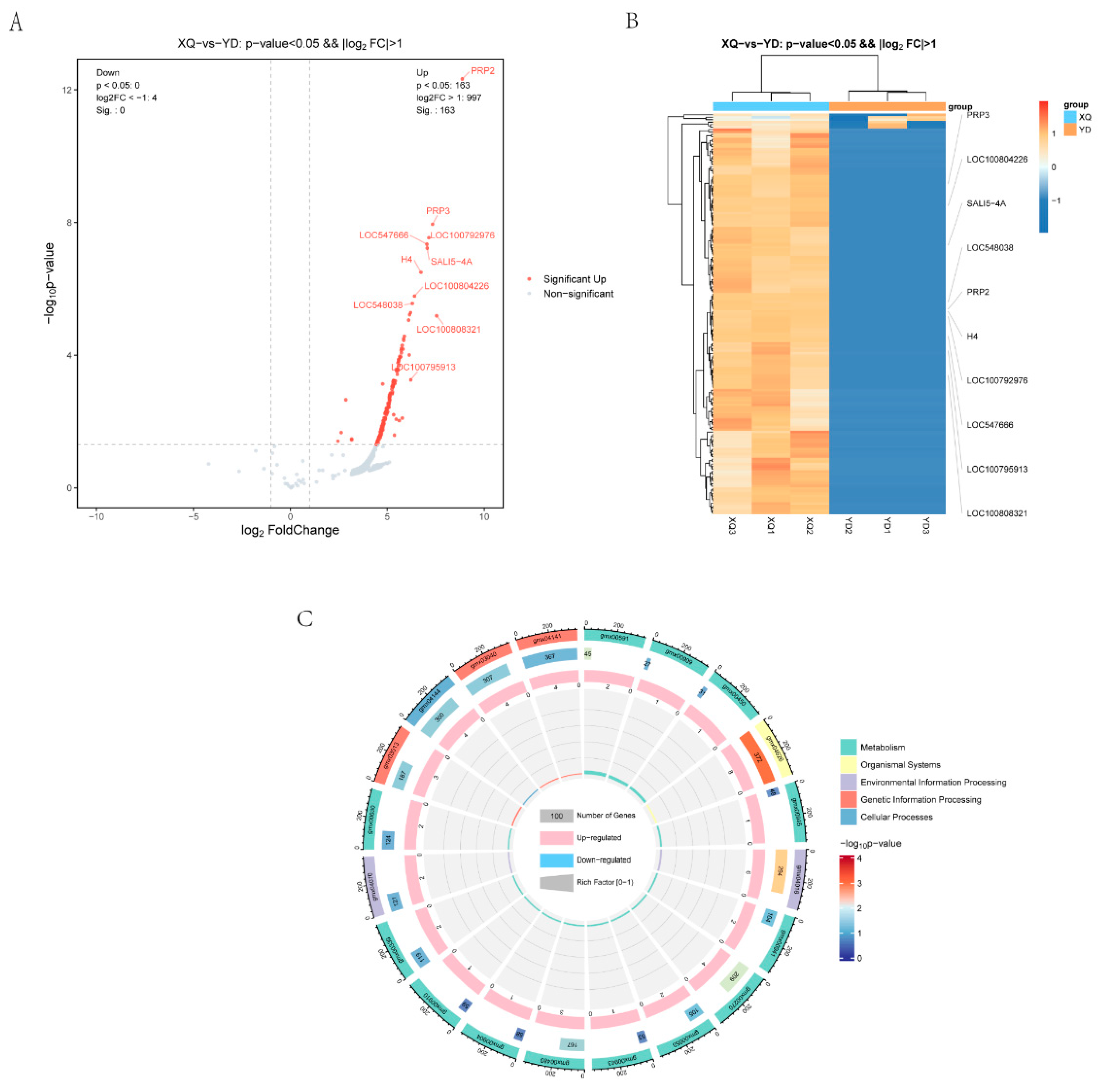
3.2. Analysis of Differentially Expressed Genes in C. australis and Diameter
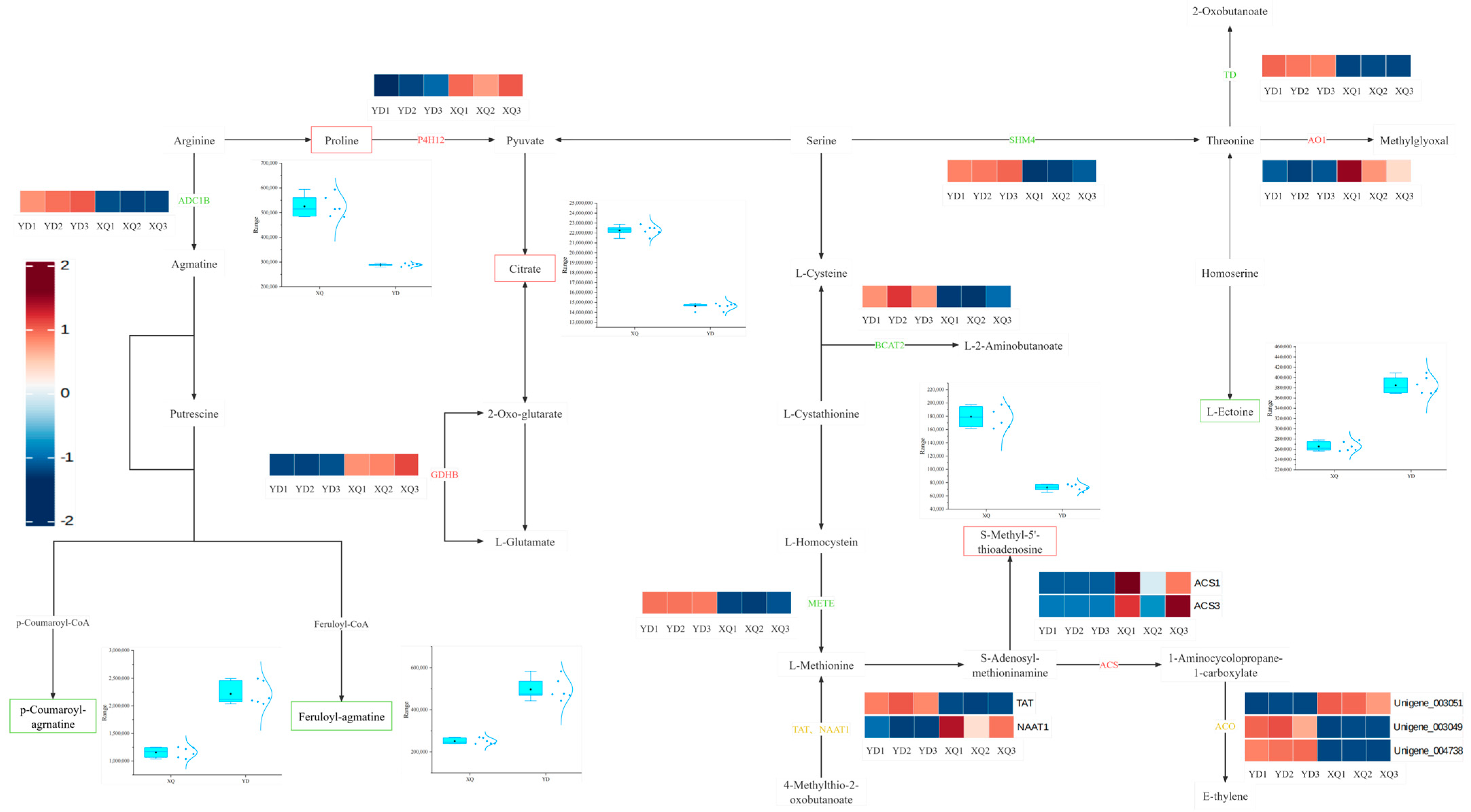
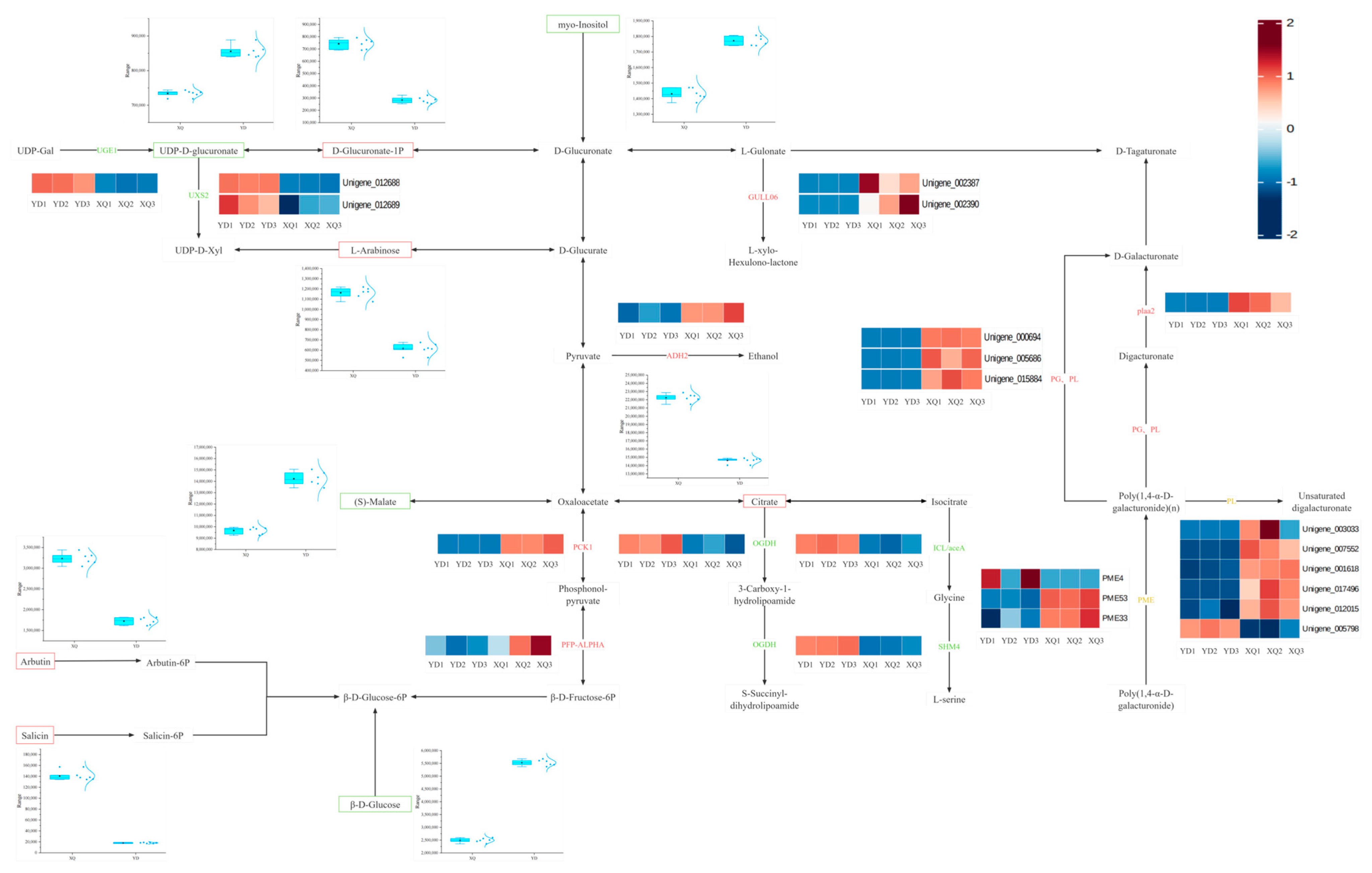
3.3. Differences in Metabolite and Gene Co-Expression Networks Between the Pipette and the Diameter End
3.4. C. australis Seeds Communicate Through Genes with the Host
3.5. Differences in the Co-Expression Network of Genes and Metabolites in G. max Sucked by the C. australis Dodder Between the Sucker and the Distal Diameter Segment
4. Discussion
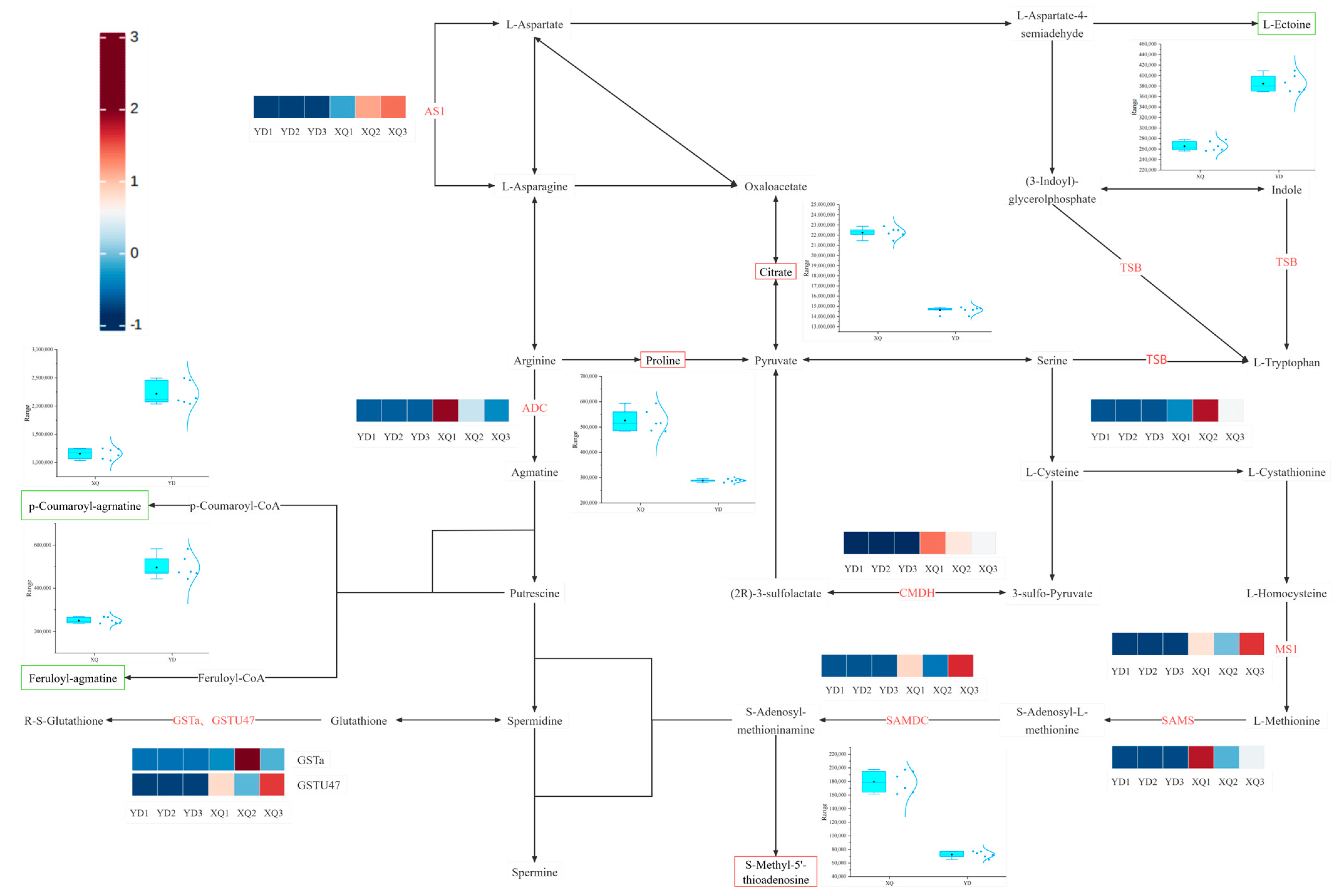
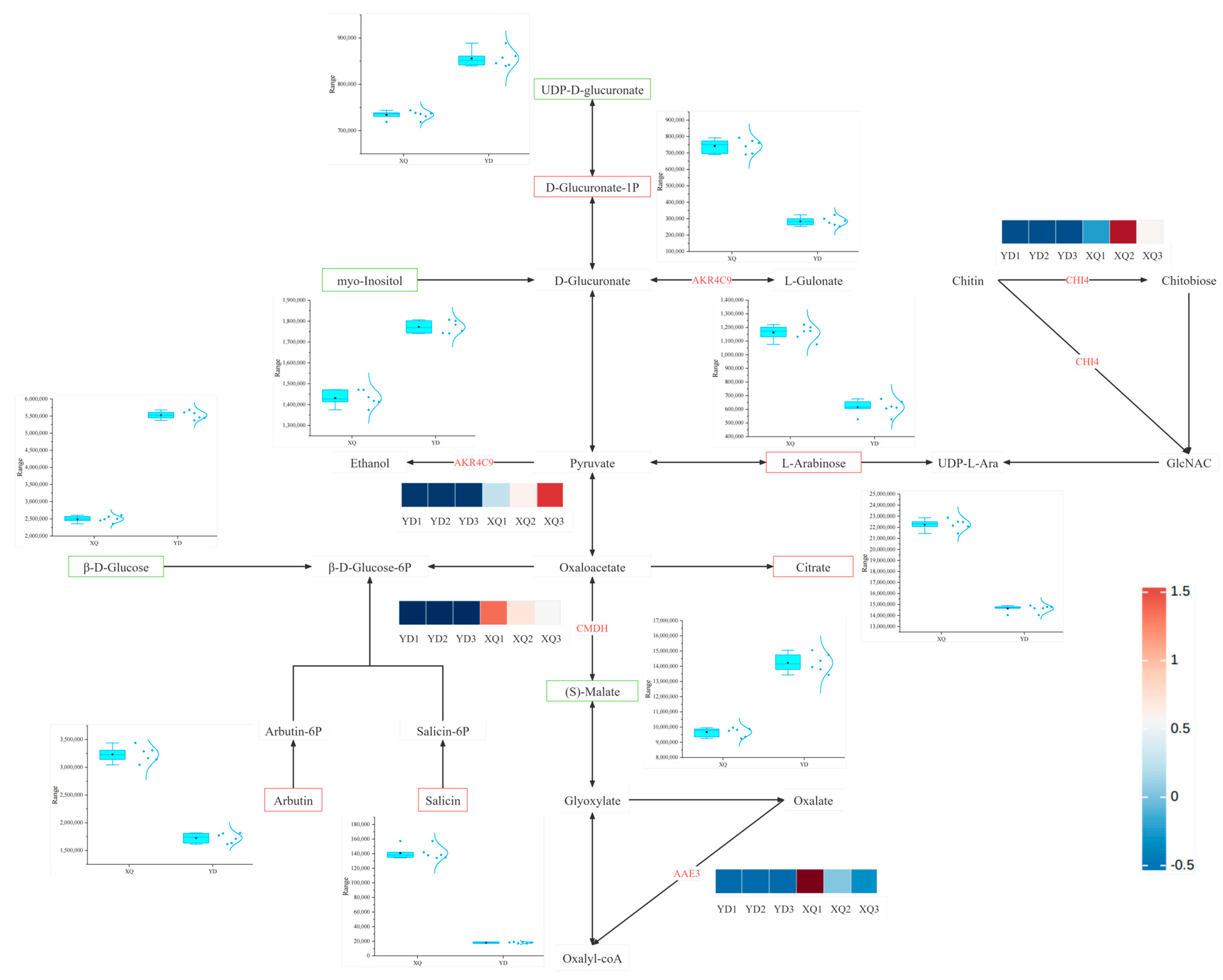
4.1. Differential Analysis of Gene and Metabolite Expression in the Aspirator and Distal Stem of C. australis
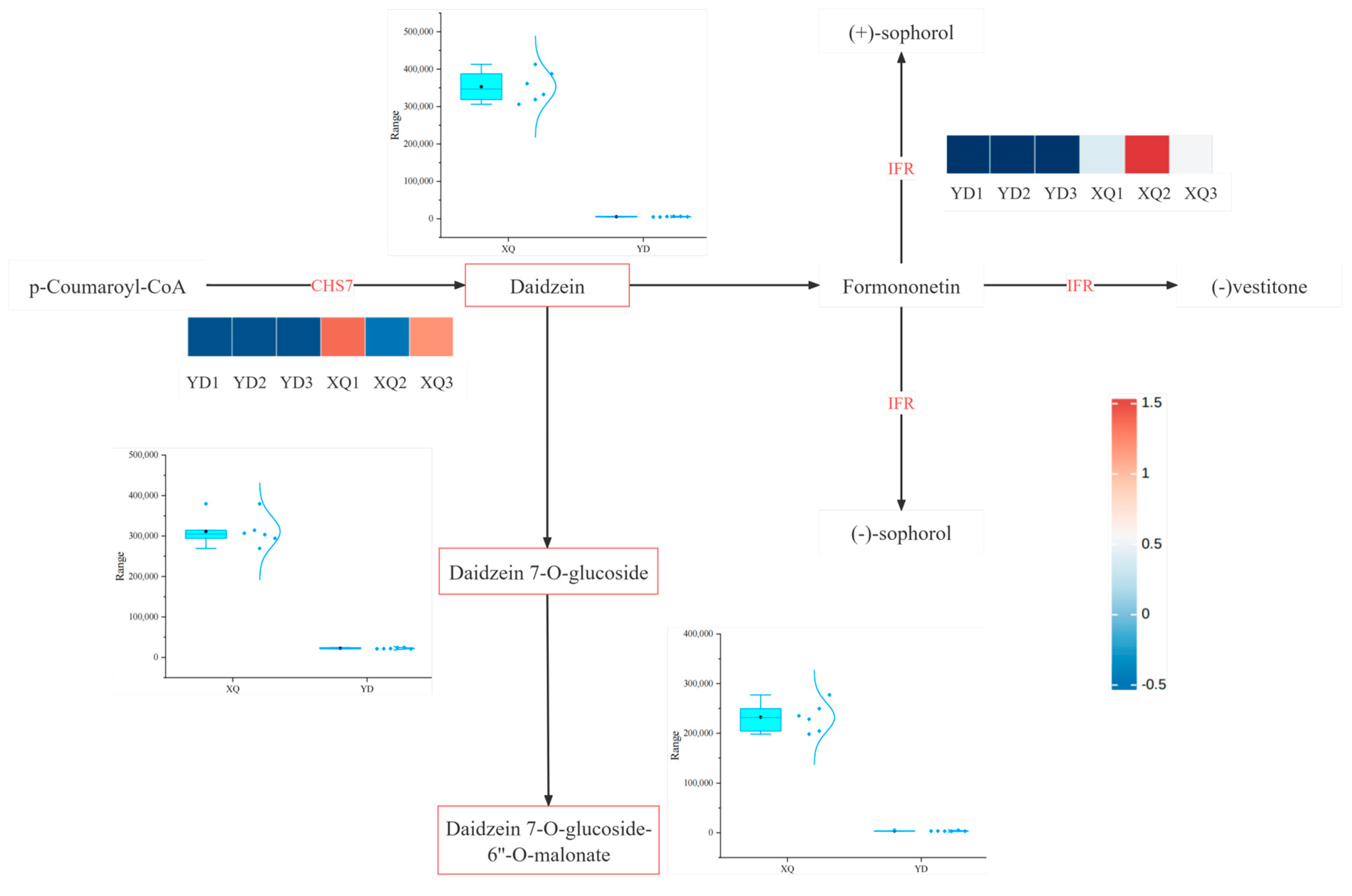
4.2. Differential Genes Taken by C. australis Chinensis from G. max Hosts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Birschwilks, M.; Haupt, S.; Hofius, D.; Neumann, S. Transfer of phloem-mobile substances from the host plants to the holoparasite Cuscuta sp. J. Exp. Bot. 2006, 57, 911–921. [Google Scholar] [CrossRef]
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The Haustorium, a Specialized Invasive Organ in Parasitic Plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef]
- Kirschner, G.K.; Xiao, T.T.; Jamil, M.; Al-Babili, S.; Lube, V.; Blilou, I. A roadmap of haustorium morphogenesis in parasitic plants. J. Exp. Bot. 2023, 74, 7034–7044. [Google Scholar] [CrossRef]
- Kokla, A.; Leso, M.; Zhang, X.; Simura, J.; Serivichyaswat, P.T.; Cui, S.; Ljung, K.; Yoshida, S.; Melnyk, C.W. Nitrogen represses haustoria formation through abscisic acid in the parasitic plant Phtheirospermum japonicum. Nat. Commun. 2022, 13, 2976. [Google Scholar] [CrossRef]
- Leso, M.; Kokla, A.; Feng, M.; Melnyk, C.W. Pectin modifications promote haustoria development in the parasitic plant Phtheirospermum japonicum. Plant Physiol. 2023, 194, 229–242. [Google Scholar] [CrossRef]
- Ogawa, S.; Wakatake, T.; Spallek, T.; Ishida, J.K.; Sano, R.; Kurata, T.; Demura, T.; Yoshida, S.; Ichihashi, Y.; Schaller, A.; et al. Subtilase activity in intrusive cells mediates haustorium maturation in parasitic plants. Plant Physiol. 2020, 185, 1381–1394. [Google Scholar] [CrossRef]
- Aoki, N.; Cui, S.; Yoshida, S. Cytokinins Induce Prehaustoria Coordinately with Quinone Signals in the Parasitic Plant Striga hermonthica. Plant Cell Physiol. 2022, 63, 1446–1456. [Google Scholar] [CrossRef]
- Cui, S.; Kubota, T.; Nishiyama, T.; Ishida, J.K.; Shigenobu, S.; Shibata, T.F.; Toyoda, A.; Hasebe, M.; Shirasu, K.; Yoshida, S. Ethylene signaling mediates host invasion by parasitic plants. Sci. Adv. 2020, 6, eabc2385. [Google Scholar] [CrossRef]
- Jhu, M.-Y.; Sinha, N.R. Cuscuta species: Model organisms for haustorium development in stem holoparasitic plants. Frontiers 2022, 13, 1086384. [Google Scholar] [CrossRef]
- Spallek, T.; Melnyk, C.W.; Wakatake, T.; Zhang, J.; Sakamoto, Y.; Kiba, T.; Yoshida, S.; Matsunaga, S.; Sakakibara, H.; Shirasu, K. Interspecies hormonal control of host root morphology by parasitic plants. Proc. Natl. Acad. Sci. USA 2017, 114, 5283–5288. [Google Scholar] [CrossRef]
- Clarke, C.R.; Timko, M.P.; Yoder, J.I.; Axtell, M.J.; Westwood, J.H. Molecular Dialog Between Parasitic Plants and Their Hosts. Annu. Rev. Phytopathol. 2019, 57, 279–299. [Google Scholar] [CrossRef]
- Wu, Y.; Luo, D.; Fang, L.; Zhou, Q.; Liu, W.; Liu, Z. Bidirectional lncRNA Transfer between Cuscuta Parasites and Their Host Plant. Int. J. Mol. Sci. 2022, 23, 561. [Google Scholar] [CrossRef]
- Hettenhausen, C.; Li, J.; Zhuang, H.; Sun, H.; Xu, Y.; Qi, J.; Zhang, J.; Lei, Y.; Qin, Y.; Sun, G.; et al. Stem parasitic plant Cuscuta australis (dodder) transfers herbivory-induced signals among plants. Proc. Natl. Acad. Sci. USA 2017, 114, E6703–E6709. [Google Scholar] [CrossRef]
- Shen, G.; Liu, N.; Zhang, J.; Xu, Y.; Baldwin, I.T.; Wu, J. Cuscuta australis (dodder) parasite eavesdrops on the host plants’ FT signals to flower. Proc. Natl. Acad. Sci. USA 2020, 117, 23125–23130. [Google Scholar] [CrossRef]
- Zhuang, H.; Li, J.; Song, J.; Hettenhausen, C.; Schuman, M.; Sun, G.; Zhang, C.; Li, J.; Song, D.; Wu, J. Aphid (Myzus persicae) feeding on the parasitic plant dodder (Cuscuta australis) activates defense responses in both the parasite and soybean host. New Phytol. 2018, 218, 1586–1596. [Google Scholar] [CrossRef]
- Kumar, K.; Amir, R. The Effect of a Host on the Primary Metabolic Profiling of Cuscuta Campestris’ Main Organs, Haustoria, Stem and Flower. Plants 2021, 10, 2098. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Mutuku, J.M.; Yoshida, S.; Shirasu, K. Transcriptomics exposes the uniqueness of parasitic plants. Brief. Funct. Genom. 2015, 14, 275–282. [Google Scholar] [CrossRef]
- Zhou, L.; Zawaira, A.; Lu, Q.; Yang, B.; Li, J. Transcriptome analysis reveals defense-related genes and pathways during dodder (Cuscuta australis) parasitism on white clover (Trifolium repens). Front. Genet. 2023, 14, 1106936. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Kusano, M.; Kobayashi, M.; Suetsugu, K.; Yoshida, S.; Wakatake, T.; Kumaishi, K.; Shibata, A.; Saito, K.; Shirasu, K. Transcriptomic and Metabolomic Reprogramming from Roots to Haustoria in the Parasitic Plant, Thesium chinense. Plant Cell Physiol. 2018, 59, 729–738. [Google Scholar] [CrossRef]
- Lehtonen, P.; Helander, M.; Wink, M.; Sporer, F.; Saikkonen, K.J.E.L. Transfer of endophyte-origin defensive alkaloids from a grass to a hemiparasitic plant. Ecol. Lett. 2005, 8, 1256–1263. [Google Scholar] [CrossRef]
- Dai, J.; Wang, X.; Meng, X.; Zhang, X.; Zhou, Q.; Zhang, Z.; Zhang, X.; Yi, Y.; Liu, L.; Shen, T. UPLC–MS/MS and Gene Expression Research to Distinguish the Colour Differences of Rhododendron liliiflorum H. Lév. Hortic 2023, 9, 1351. [Google Scholar] [CrossRef]
- Supral, A.; Asha, M.; Lydia, P.; Hyeyoung, L.; Vivian, B.G.; Hope, G.; James, H.W.; So-Yon, P. Agrobacterium-mediated Cuscuta campestris transformation as a tool for understanding plant–plant interactions. New Phytol. 2025, 245, 1774–1786. [Google Scholar] [CrossRef]
- Shen, G.; Zhang, J.; Lei, Y.; Xu, Y.; Wu, J. Between-Plant Signaling. Annu. Rev. Plant Biol. 2023, 74, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Balios, V.A.; Fischer, K.; Bawin, T.; Krause, K. One organ to infect them all: The Cuscuta haustorium. Ann. Bot. 2024, mcae208. [Google Scholar] [CrossRef]
- Furuta, K.M.; Xiang, L.; Cui, S.; Yoshida, S. Molecular dissection of haustorium development in Orobanchaceae parasitic plants. Plant Physiol. 2021, 186, 1424–1434. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, M.; Wang, X.; Dai, J.; Zhang, X.; Zhang, Z.; Zhang, X.; Tang, M.; Tang, J.; Gong, J.; et al. Transcriptome Analysis of Rhododendron liliiflorum H. Lév. Flower Colour Differences. Hortic 2023, 9, 82. [Google Scholar] [CrossRef]
- Hozumi, A.; Bera, S.; Fujiwara, D.; Obayashi, T.; Yokoyama, R.; Nishitani, K.; Aoki, K. Arabinogalactan Proteins Accumulate in the Cell Walls of Searching Hyphae of the Stem Parasitic Plants, Cuscuta campestris and Cuscuta japonica. Plant Cell Physiol. 2017, 58, 1868–1877. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Li, R.; Peng, G.; Chen, W.; Rautengarten, C.; Liu, M.; Zhu, L.; Xiao, Y.; Song, F.; et al. BOTRYOID POLLEN 1 regulates ROS-triggered PCD and pollen wall development by controlling UDP-sugar homeostasis in rice. Plant Cell 2023, 35, 3522–3543. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, W.; Feng, Z.; Zhang, S.; Zheng, X.; Xu, Y.; Shen, G.; Zhao, M.; Cao, G.; et al. Large-scale interplant exchange of macromolecules between soybean and dodder under nutrient stresses. Plant Divers. 2024, 46, 116–125. [Google Scholar] [CrossRef]
- Jhu, M.Y.; Ichihashi, Y.; Farhi, M.; Wong, C.; Sinha, N.R. LATERAL ORGAN BOUNDARIES DOMAIN 25 functions as a key regulator of haustorium development in dodders. Plant Physiol. 2021, 186, 2093–2110. [Google Scholar] [CrossRef]
- Mutlu, B.; Sharabi, K.; Sohn, J.H.; Yuan, B.; Latorre-Muro, P.; Qin, X.; Yook, J.-S.; Lin, H.; Yu, D.; Camporez, J.P.G.; et al. Small molecules targeting selective PCK1 and PGC-1α lysine acetylation cause anti-diabetic action through increased lactate oxidation. Cell Chem. Biol. 2024, 31, 1772–1786.e1775. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Ichihashi, Y.; Farhi, M.; Zumstein, K.; Townsley, B.; David-Schwartz, R.; Sinha, N.R. De Novo Assembly and Characterization of the Transcriptome of the Parasitic Weed Dodder Identifies Genes Associated with Plant Parasitism. Plant Physiol. 2014, 166, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Wolswinkel, P.; Ammerlaan, A.; Peters, H.F.C. Phloem Unloading of Amino Acids at the Site of Attachment of Cuscuta europaea. Plant Physiol. 1984, 75, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Akram, M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- He, H.; He, L.F. Regulation of gaseous signaling molecules on proline metabolism in plants. Plant Cell Rep. 2018, 37, 387–392. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Xie, J.; Zhuang, H.; Liu, H.; Shen, G.; Wu, J. Parasite dodder enables transfer of bidirectional systemic nitrogen signals between host plants. Plant Physiol. 2021, 185, 1395–1410. [Google Scholar] [CrossRef]
- Zhao, T.; Yang, H.; Jiang, J.; Liu, G.; Zhang, H.; Xiao, D.; Chen, X.; Li, J.; Xu, X. Silencing of the SAMDC gene decreases resistance of tomato to Cladosporium fulvum. Physiol. Mol. Plant Pathol. 2018, 102, 1–7. [Google Scholar] [CrossRef]
- Ohtomo, T.; Uehara, M.; Peñalvo, J.L.; Adlercreutz, H.; Katsumata, S.-I.; Suzuki, K.; Takeda, K.; Masuyama, R.; Ishimi, Y. Comparative activities of daidzein metabolites, equol and O-desmethylangolensin, on bone mineral density and lipid metabolism in ovariectomized mice and in osteoclast cell cultures. Eur. J. Nutr. 2008, 47, 273–279. [Google Scholar] [CrossRef]
- Mameda, R.; Waki, T.; Kawai, Y.; Takahashi, S.; Nakayama, T. Involvement of chalcone reductase in the soybean isoflavone metabolon: Identification of GmCHR5, which interacts with 2-hydroxyisoflavanone synthase. Plant J. 2018, 96, 56–74. [Google Scholar] [CrossRef]
- Rani, D.; Meelaph, T.; De-Eknamkul, W.; Vimolmangkang, S. Yeast extract elicited isoflavonoid accumulation and biosynthetic gene expression in Pueraria candollei var. mirifica cell cultures. Plant Cell Tissue Organ Cult. 2020, 141, 661–667. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Lv, N.; Wang, X.; Zhou, Q.; Zhang, X.; Zhang, X.; Zhang, Z.; Liu, L.; Shen, T. Molecular Mechanism of Cuscuta Haustorium Specialization Inferences from Transcriptome and Metabolome Analysis. Metabolites 2025, 15, 172. https://doi.org/10.3390/metabo15030172
Meng X, Lv N, Wang X, Zhou Q, Zhang X, Zhang X, Zhang Z, Liu L, Shen T. Molecular Mechanism of Cuscuta Haustorium Specialization Inferences from Transcriptome and Metabolome Analysis. Metabolites. 2025; 15(3):172. https://doi.org/10.3390/metabo15030172
Chicago/Turabian StyleMeng, Xingpan, Ning Lv, Xinglin Wang, Qihang Zhou, Xu Zhang, Ximin Zhang, Zhengdong Zhang, Lunxian Liu, and Tie Shen. 2025. "Molecular Mechanism of Cuscuta Haustorium Specialization Inferences from Transcriptome and Metabolome Analysis" Metabolites 15, no. 3: 172. https://doi.org/10.3390/metabo15030172
APA StyleMeng, X., Lv, N., Wang, X., Zhou, Q., Zhang, X., Zhang, X., Zhang, Z., Liu, L., & Shen, T. (2025). Molecular Mechanism of Cuscuta Haustorium Specialization Inferences from Transcriptome and Metabolome Analysis. Metabolites, 15(3), 172. https://doi.org/10.3390/metabo15030172