
Pesticides: Unintended Impact on the Hidden World of Gut Microbiota



Abstract
1. Introduction
2. Gut Microbiome
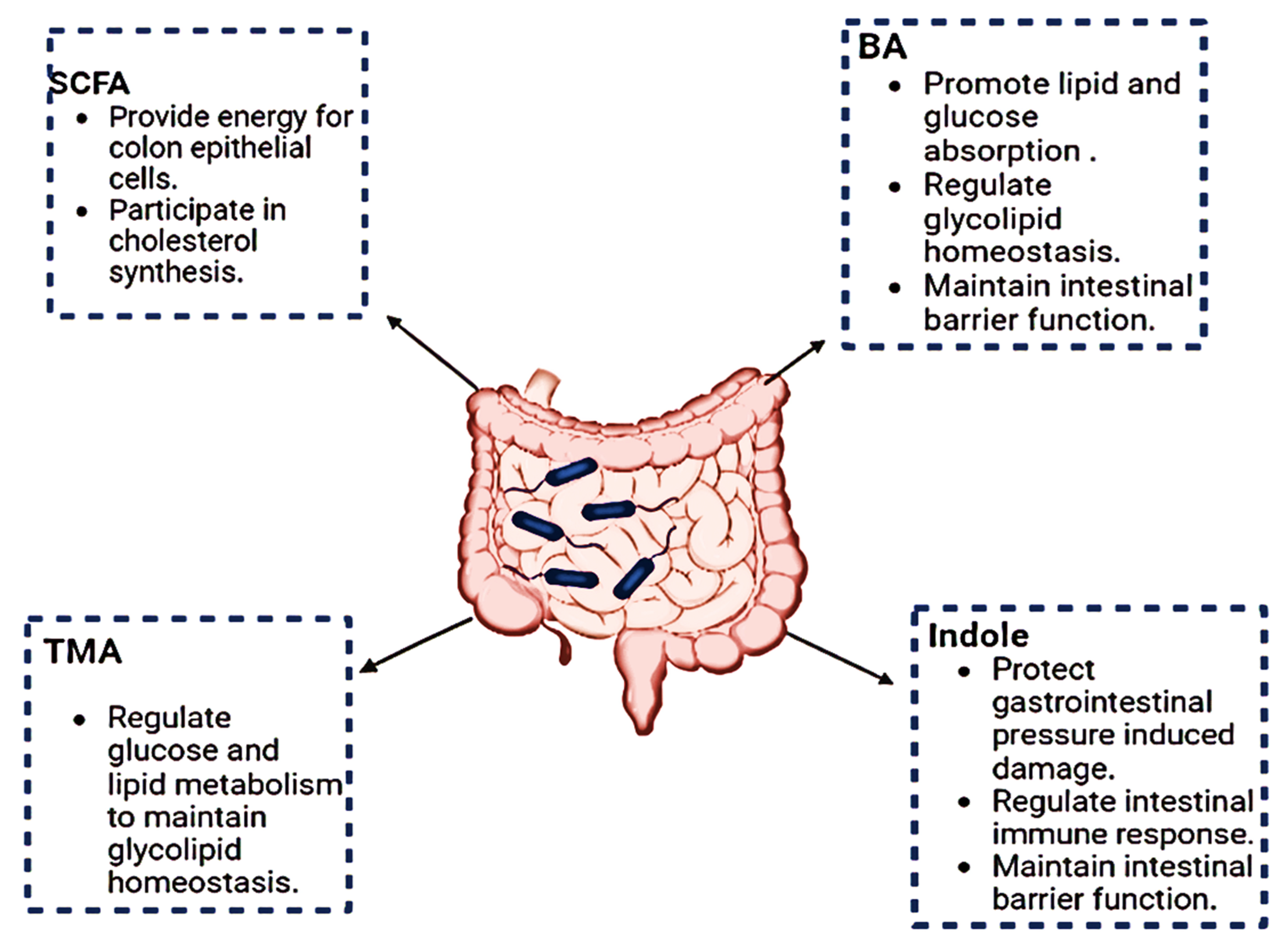
2.1. Function of Gut Microbe Metabolites
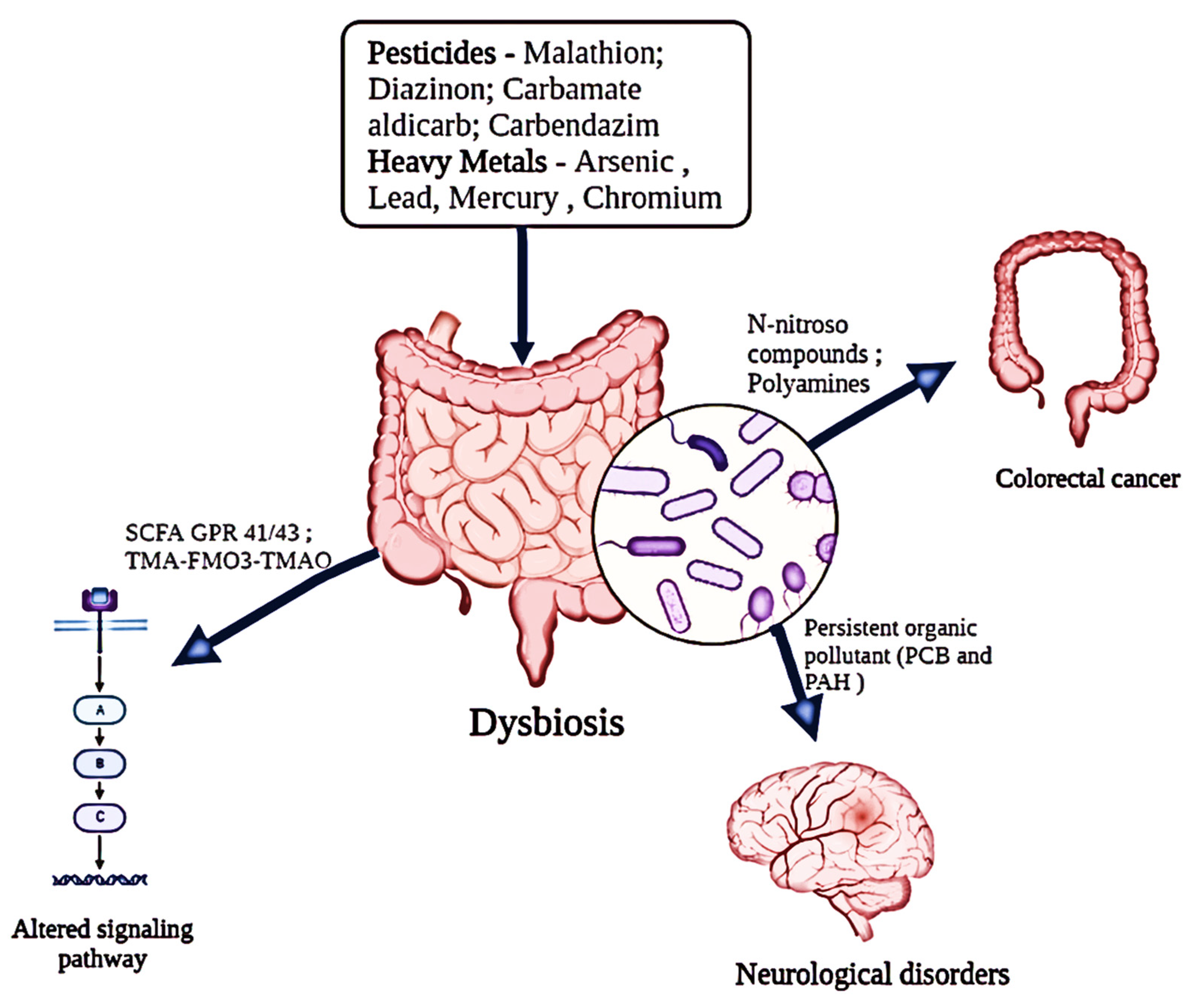
2.2. Dysbiosis
3. Effect of Insecticides on Gut Microbiota
3.1. Organophosphate Pesticides (OPPs)
3.2. Organochlorine Pesticides (OCPs)
3.3. Permethrin (PEM)
4. Effects of Fungicides on the Gut Microbiota
4.1. Carbendazim (CBZ)
4.2. Imazalil (IMZ)
4.3. Propamocarb (PM)
4.4. Epoxiconazole
5. Effects of Herbicides on the Gut Microbiota
5.1. Glyphosate (GLY)
5.2. Pentachlorophenol (PCP)
6. Effects of Pesticides on Gut Microbiota-Mediated Host Immune System
7. Other Potential Environmental Contaminants
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| AAA | Aromatic amino acid |
| AhR | Aryl hydrocarbon receptor |
| CBZ | Carbendazim (methyl 2-benzimidazole carbamate) |
| CPF | Chlorpyrifos |
| DDT | Dichlorodiphenyltrichloroethane |
| FMO3 | Flavin mono-oxygenase 3 |
| FXR | Farnesoid X receptor |
| GBA | Gut–brain axis |
| GI | Gastrointestinal |
| GLY | Glyphosate |
| GM | Gut microbiota |
| HCH | hexachlorocyclohexane |
| IL | Interleukin |
| IMZ | Imazalil |
| LPS | Lipopolysaccharide |
| NAFLD | Non–alcoholic fatty liver disease |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| NOS | Synthase |
| OCPs | Organochlorine pesticides |
| OPPs | Organophosphate pesticides |
| p,p’-DDE | p,p’-dichlorodiphenyldichloroethylene |
| PCP | Pentachlorophenol |
| PEM | Permethrin |
| PM | Propamocarb |
| SCFAs | Short-chain fatty acids |
| TMA | Trimethylamine |
| TMO | Trimethylamine N-oxide |
References
- Jin, Y.; Wu, S.; Zeng, Z.; Fu, Z. Effects of Environmental Pollutants on Gut Microbiota. Environ. Pollut. 2017, 222, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-Induced Pathogen Susceptibility Is Mitigated by Lactobacillus Plantarum Immune Stimulation in a Drosophila Melanogaster Model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Briguglio, G.; Catanoso, R.; Giambò, F.; Polito, I.; Teodoro, M.; Fenga, C. New Perspectives on Cytokine Pathways Modulation by Pesticide Exposure. Curr. Opin. Toxicol. 2020, 19, 99–104. [Google Scholar] [CrossRef]
- Kittle, R.P.; McDermid, K.J.; Muehlstein, L.; Balazs, G.H. Effects of Glyphosate Herbicide on the Gastrointestinal Microflora of Hawaiian Green Turtles (Chelonia mydas) Linnaeus. Mar. Pollut. Bull. 2018, 127, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Defois, C.; Ratel, J.; Garrait, G.; Denis, S.; Le Goff, O.; Talvas, J.; Mosoni, P.; Engel, E.; Peyret, P. Food Chemicals Disrupt Human Gut Microbiota Activity and Impact Intestinal Homeostasis as Revealed by In Vitro Systems. Sci. Rep. 2018, 8, 11006. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, Z.; Gibson, D.L.; Hekmatdoost, A. Nonalcoholic Fatty Liver Disease, the Gut Microbiome, and Diet. Adv. Nutr. 2017, 8, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Singer-Englar, T.; Barlow, G.; Mathur, R. Obesity, Diabetes, and the Gut Microbiome: An Updated Review. Expert. Rev. Gastroenterol. Hepatol. 2019, 13, 3–15. [Google Scholar] [CrossRef]
- Salazar, J.; Durán, P.; Díaz, M.P.; Chacín, M.; Santeliz, R.; Mengual, E.; Gutiérrez, E.; León, X.; Díaz, A.; Bernal, M.; et al. Exploring the Relationship between the Gut Microbiota and Ageing: A Possible Age Modulator. Int. J. Environ. Res. Public Health 2023, 20, 5845. [Google Scholar] [CrossRef]
- Barlow, G.M.; Yu, A.; Mathur, R. Role of the Gut Microbiome in Obesity and Diabetes Mellitus. Nutr. Clin. Pract. 2015, 30, 787–797. [Google Scholar] [CrossRef]
- Tokuhara, D. Role of the Gut Microbiota in Regulating Non-Alcoholic Fatty Liver Disease in Children and Adolescents. Front. Nutr. 2021, 8, 700058. [Google Scholar] [CrossRef]
- Fujita, K.; Matsushita, M.; Banno, E.; De Velasco, M.A.; Hatano, K.; Nonomura, N.; Uemura, H. Gut Microbiome and Prostate Cancer. Int. J. Urol. 2022, 29, 793–798. [Google Scholar] [CrossRef]
- Quesada-Vázquez, S.; Aragonès, G.; Del Bas, J.M.; Escoté, X. Diet, Gut Microbiota and Non-Alcoholic Fatty Liver Disease: Three Parts of the Same Axis. Cells 2020, 9, 176. [Google Scholar] [CrossRef]
- Bakhshimoghaddam, F.; Alizadeh, M. Contribution of Gut Microbiota to Nonalcoholic Fatty Liver Disease: Pathways of Mechanisms. Clin. Nutr. ESPEN 2021, 44, 61–68. [Google Scholar] [CrossRef]
- Vallianou, N.; Christodoulatos, G.S.; Karampela, I.; Tsilingiris, D.; Magkos, F.; Stratigou, T.; Kounatidis, D.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Non-Alcoholic Fatty Liver Disease: Current Evidence and Perspectives. Biomolecules 2022, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tripathi, P. Gut Microbiome and Type 2 Diabetes: Where We Are and Where to Go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, K.; Zhou, P.; Yong, W. Gut Microbiota in Obesity and Nonalcoholic Fatty Liver Disease. Surg. Pract. Sci. 2021, 5, 100030. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional Interactions between the Gut Microbiota and Host Metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Schippa, S.; Conte, M.P. Dysbiotic Events in Gut Microbiota: Impact on Human Health. Nutrients 2014, 6, 5786–5805. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy Regulates Lipid Metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic T Reg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile Acids and the Gut Microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef]
- Mouzaki, M.; Loomba, R. Insights into the Evolving Role of the Gut Microbiome in Nonalcoholic Fatty Liver Disease: Rationale and Prospects for Therapeutic Intervention. Therap. Adv. Gastroenterol. 2019, 12, 1756284819858470. [Google Scholar] [CrossRef]
- Xie, C.; Halegoua-Demarzio, D. Role of Probiotics in Non-Alcoholic Fatty Liver Disease: Does Gut Microbiota Matter? Nutrients 2019, 11, 2837. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The Gut Microbiota and Host Health: A New Clinical Frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef]
- Illiano, P.; Brambilla, R.; Parolini, C. The Mutual Interplay of Gut Microbiota, Diet and Human Disease. FEBS J. 2020, 287, 833–855. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Degruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.; Nigam, P.S. Antibiotic-Therapy-Induced Gut Dysbiosis Affecting Gut Microbiota—Brain Axis and Cognition: Restoration by Intake of Probiotics and Synbiotics. Int. J. Mol. Sci. 2023, 24, 3074. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.A.; Konnikova, L.; Gerber, J.S. Impact of Antibiotics on Necrotizing Enterocolitis and Antibiotic-Associated Diarrhea. Gastroenterol. Clin. N. Am. 2017, 46, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Sadrekarimi, H.; Gardanova, Z.R.; Bakhshesh, M.; Ebrahimzadeh, F.; Yaseri, A.F.; Thangavelu, L.; Hasanpoor, Z.; Zadeh, F.A.; Kahrizi, M.S. Emerging Role of Human Microbiome in Cancer Development and Response to Therapy: Special Focus on Intestinal Microflora. J. Transl. Med. 2022, 20, 301. [Google Scholar] [CrossRef]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid Pesticide Residues in the Global Environment: An Overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.V.; Lopez-Santamarina, A.; del Carmen Mondragón, A.; Regal, P.; Lamas, A.; Rodriguez, J.A.; Miranda, J.M.; Franco, C.M. Effects of Pesticides Carried by Foods on Human Gut Microbiota. Lett. Funct. Foods 2023, 1, e160523216967. [Google Scholar] [CrossRef]
- Matsuzaki, R.; Gunnigle, E.; Geissen, V.; Clarke, G.; Nagpal, J.; Cryan, J.F. Pesticide Exposure and the Microbiota-Gut-Brain Axis. ISME J. 2023, 17, 1153–1166. [Google Scholar] [CrossRef]
- Sharma, T.; Sirpu Natesh, N.; Pothuraju, R.; Batra, S.K.; Rachagani, S. Gut Microbiota: A Non-Target Victim of Pesticide-Induced Toxicity. Gut Microbes 2023, 15, 2187578. [Google Scholar] [CrossRef]
- Velmurugan, G.; Ramprasath, T.; Gilles, M.; Swaminathan, K.; Ramasamy, S. Gut Microbiota, Endocrine-Disrupting Chemicals, and the Diabetes Epidemic. Trends Endocrinol. Metab. 2017, 28, 612–625. [Google Scholar] [CrossRef]
- Picó, Y.; El-Sheikh, M.A.; Alfarhan, A.H.; Barceló, D. Target vs Non-Target Analysis to Determine Pesticide Residues in Fruits from Saudi Arabia and Influence in Potential Risk Associated with Exposure. Food Chem. Toxicol. 2018, 111, 53–63. [Google Scholar] [CrossRef]
- Yuan, X.; Pan, Z.; Jin, C.; Ni, Y.; Fu, Z.; Jin, Y. Gut Microbiota: An Underestimated and Unintended Recipient for Pesticide-Induced Toxicity. Chemosphere 2019, 227, 425–434. [Google Scholar] [CrossRef]
- Réquilé, M.; Gonzàlez Alvarez, D.O.; Delanaud, S.; Rhazi, L.; Bach, V.; Depeint, F.; Khorsi-Cauet, H. Use of a Combination of in Vitro Models to Investigate the Impact of Chlorpyrifos and Inulin on the Intestinal Microbiota and the Permeability of the Intestinal Mucosa. Environ. Sci. Pollut. Res. 2018, 25, 22529–22540. [Google Scholar] [CrossRef] [PubMed]
- Giambò, F.; Teodoro, M.; Costa, C.; Fenga, C. Toxicology and Microbiota: How Do Pesticides Influence Gut Microbiota? A Review. Int. J. Environ. Res. Public. Health 2021, 18, 5510. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Y.; Wang, G.; Han, R.; Xie, X. Effects of Chlorpyrifos on the Gut Microbiome and Urine Metabolome in Mouse (Mus musculus). Chemosphere 2016, 153, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, M.; Zhou, J.; Jin, Y. Chlorpyrifos Disturbs Hepatic Metabolism Associated with Oxidative Stress and Gut Microbiota Dysbiosis in Adult Zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 216, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Deng, X.; Tuli, A.; Goh, K.S. Diazinon—Chemistry and Environmental Fate: A California Perspective. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 223, pp. 107–140. [Google Scholar]
- Gao, B.; Bian, X.; Chi, L.; Tu, P.; Ru, H.; Lu, K. Organophosphate Diazinon Altered Quorum Sensing, Cell Motility, Stress Response, and Carbohydrate Metabolism of Gut Microbiome. Toxicol. Sci. 2017, 157, 354–364. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, L.; Yan, S.; Sun, W.; Jia, M.; Tian, S.; Huang, S.; Zhou, Z.; Zhu, W. Gut Microbiota: A Key Factor in the Host Health Effects Induced by Pesticide Exposure? J. Agric. Food Chem. 2020, 68, 10517–10531. [Google Scholar] [CrossRef]
- Maisano, M.; Cappello, T.; Oliva, S.; Natalotto, A.; Giannetto, A.; Parrino, V.; Battaglia, P.; Romeo, T.; Salvo, A.; Spanò, N.; et al. PCB and OCP Accumulation and Evidence of Hepatic Alteration in the Atlantic Bluefin Tuna, T. thynnus, from the Mediterranean Sea. Mar. Environ. Res. 2016, 121, 40–48. [Google Scholar] [CrossRef]
- Zhan, J.; Liang, Y.; Liu, D.; Ma, X.; Li, P.; Zhai, W.; Zhou, Z.; Wang, P. Pectin Reduces Environmental Pollutant-Induced Obesity in Mice through Regulating Gut Microbiota: A Case Study of p,p’-DDE. Environ. Int. 2019, 130, 104861. [Google Scholar] [CrossRef]
- Liang, Y.; Zhan, J.; Liu, D.; Luo, M.; Han, J.; Liu, X.; Liu, C.; Cheng, Z.; Zhou, Z.; Wang, P. Organophosphorus Pesticide Chlorpyrifos Intake Promotes Obesity and Insulin Resistance through Impacting Gut and Gut Microbiota. Microbiome 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Shao, W.; Zhang, C.; Xu, C.; Wang, Q.; Liu, H.; Sun, H.; Jiang, Z.; Gu, A. Organochloride Pesticides Modulated Gut Microbiota and Influenced Bile Acid Metabolism in Mice. Environ. Pollut. 2017, 226, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, J.; Hayashi, M.; Hirayama, M.; Nishiwaki, H.; Ito, M.; Saito, I.; Tsuboi, Y.; Isobe, T.; Ohno, K. Effects of Pesticide Intake on Gut Microbiota and Metabolites in Healthy Adults. Int. J. Environ. Res. Public. Health 2023, 20, 213. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, T.; Yang, T.; Zhu, J.; Zhou, Y.; Wang, M.; Wang, Q. Gut Microbiota Dysbiosis Involves in Host Non-Alcoholic Fatty Liver Disease upon Pyrethroid Pesticide Exposure. Environ. Sci. Ecotechnol. 2022, 11, 100185. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Peng, H.; Li, Y.; Meng, B.; Wang, S.; Bi, S.; Zhao, X. Pyrethroids Exposure Alters the Community and Function of the Internal Microbiota in Aedes Albopictus. Ecotoxicol. Environ. Saf. 2023, 252, 114579. [Google Scholar] [CrossRef]
- Jin, Y.; Zeng, Z.; Wu, Y.; Zhang, S.; Fu, Z. Oral Exposure of Mice to Carbendazim Induces Hepatic Lipid Metabolism Disorder and Gut Microbiota Dysbiosis. Toxicol. Sci. 2015, 147, 116–126. [Google Scholar] [CrossRef]
- Seth, R.K.; Kimono, D.; Alhasson, F.; Sarkar, S.; Albadrani, M.; Lasley, S.K.; Horner, R.; Janulewicz, P.; Nagarkatti, M.; Nagarkatti, P.; et al. Increased Butyrate Priming in the Gut Stalls Microbiome Associated-Gastrointestinal Inflammation and Hepatic Metabolic Reprogramming in a Mouse Model of Gulf War Illness. Toxicol. Appl. Pharmacol. 2018, 350, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Nasuti, C.; Coman, M.M.; Olek, R.A.; Fiorini, D.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Fedeli, D.; Gabbianelli, R. Changes on Fecal Microbiota in Rats Exposed to Permethrin during Postnatal Development. Environ. Sci. Pollut. Res. 2016, 23, 10930–10937. [Google Scholar] [CrossRef]
- Veerappan, M.; Hwang, I.; Pandurangan, M. Effect of Cypermethrin, Carbendazim and Their Combination on Male Albino Rat Serum. Int. J. Exp. Pathol. 2012, 93, 361–369. [Google Scholar] [CrossRef]
- Yu, G.; Liu, Y.; Xie, L.; Wang, X. Involvement of Sertoli Cells in Spermatogenic Failure Induced by Carbendazim. Environ. Toxicol. Pharmacol. 2009, 27, 287–292. [Google Scholar] [CrossRef]
- Gao, B.; Chi, L.; Tu, P.; Gao, N.; Lu, K. The Carbamate Aldicarb Altered the Gut Microbiome, Metabolome, and Lipidome of C57BL/6J Mice. Chem. Res. Toxicol. 2019, 32, 67–79. [Google Scholar] [CrossRef]
- Jin, C.; Zeng, Z.; Wang, C.; Luo, T.; Wang, S.; Zhou, J.; Ni, Y.; Fu, Z.; Jin, Y. Insights into a Possible Mechanism Underlying the Connection of Carbendazim-Induced Lipid Metabolism Disorder and Gut Microbiota Dysbiosis in Mice. Toxicol. Sci. 2018, 166, 382–393. [Google Scholar] [CrossRef]
- Jin, Y.; Zhu, Z.; Wang, Y.; Yang, E.; Feng, X.; Fu, Z. The Fungicide Imazalil Induces Developmental Abnormalities and Alters Locomotor Activity during Early Developmental Stages in Zebrafish. Chemosphere 2016, 153, 455–461. [Google Scholar] [CrossRef]
- Jin, C.; Zeng, Z.; Fu, Z.; Jin, Y. Oral Imazalil Exposure Induces Gut Microbiota Dysbiosis and Colonic Inflammation in Mice. Chemosphere 2016, 160, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Luo, T.; Zhu, Z.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z.; Jin, Y. Imazalil Exposure Induces Gut Microbiota Dysbiosis and Hepatic Metabolism Disorder in Zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 202, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Abou Diwan, M.; Lahimer, M.; Bach, V.; Gosselet, F.; Khorsi-Cauet, H.; Candela, P. Impact of Pesticide Residues on the Gut-Microbiota–Blood–Brain Barrier Axis: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 6147. [Google Scholar] [CrossRef]
- Xu, C.; Liu, Q.; Huan, F.; Qu, J.; Liu, W.; Gu, A.; Wang, Y.; Jiang, Z. Changes in Gut Microbiota May Be Early Signs of Liver Toxicity Induced by Epoxiconazole in Rats. Chemotherapy 2014, 60, 135–142. [Google Scholar] [CrossRef]
- Aitbali, Y.; Ba-M’hamed, S.; Elhidar, N.; Nafis, A.; Soraa, N.; Bennis, M. Glyphosate Based- Herbicide Exposure Affects Gut Microbiota, Anxiety and Depression-like Behaviors in Mice. Neurotoxicol. Teratol. 2018, 67, 44–49. [Google Scholar] [CrossRef]
- Kan, H.; Zhao, F.; Zhang, X.-X.; Ren, H.; Gao, S. Correlations of Gut Microbial Community Shift with Hepatic Damage and Growth Inhibition of Carassius auratus Induced by Pentachlorophenol Exposure. Environ. Sci. Technol. 2015, 49, 11894–11902. [Google Scholar] [CrossRef] [PubMed]
- Agostini, L.P.; Dettogni, R.S.; dos Reis, R.S.; Stur, E.; dos Santos, E.V.W.; Ventorim, D.P.; Garcia, F.M.; Cardoso, R.C.; Graceli, J.B.; Louro, I.D. Effects of Glyphosate Exposure on Human Health: Insights from Epidemiological and in Vitro Studies. Sci. Total Environ. 2020, 705, 135808. [Google Scholar] [CrossRef]
- Lozano, V.L.; Defarge, N.; Rocque, L.-M.; Mesnage, R.; Hennequin, D.; Cassier, R.; de Vendômois, J.S.; Panoff, J.-M.; Séralini, G.-E.; Amiel, C. Sex-Dependent Impact of Roundup on the Rat Gut Microbiome. Toxicol. Rep. 2018, 5, 96–107. [Google Scholar] [CrossRef]
- Nielsen, L.N.; Roager, H.M.; Casas, M.E.; Frandsen, H.L.; Gosewinkel, U.; Bester, K.; Licht, T.R.; Hendriksen, N.B.; Bahl, M.I. Glyphosate Has Limited Short-Term Effects on Commensal Bacterial Community Composition in the Gut Environment Due to Sufficient Aromatic Amino Acid Levels. Environ. Pollut. 2018, 233, 364–376. [Google Scholar] [CrossRef]
- Argou-Cardozo, I.; Zeidán-Chuliá, F. Clostridium Bacteria and Autism Spectrum Conditions: A Systematic Review and Hypothetical Contribution of Environmental Glyphosate Levels. Med. Sci. 2018, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Iori, S.; Rovere, G.D.; Ezzat, L.; Smits, M.; Ferraresso, S.S.; Babbucci, M.; Marin, M.G.; Masiero, L.; Fabrello, J.; Garro, E.; et al. The Effects of Glyphosate and AMPA on the Mediterranean Mussel Mytilus Galloprovincialis and Its Microbiota. Environ. Res. 2020, 182, 108984. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yin, D.; Kong, F. The Changes of Serum Testosterone Level and Hepatic Microsome Enzyme Activity of Crucian Carp (Carassius carassius) Exposed to a Sublethal Dosage of Pentachlorophenol. Ecotoxicol. Environ. Saf. 2008, 71, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.; Falcão, M.A.P.; Rosa, J.G.S.; Disner, G.R.; Lopes-Ferreira, M. Pesticides and Their Impairing Effects on Epithelial Barrier Integrity, Dysbiosis, Disruption of the AhR Signaling Pathway and Development of Immune-Mediated Inflammatory Diseases. Int. J. Mol. Sci. 2022, 23, 12402. [Google Scholar] [CrossRef]
- Tirelli, V.; Catone, T.; Turco, L.; Di Consiglio, E.; Testai, E.; De Angelis, I. Effects of the Pesticide Clorpyrifos on an In Vitro Model of Intestinal Barrier. Toxicol. Vitr. 2007, 21, 308–313. [Google Scholar] [CrossRef]
- Linares, R.; Francés, R.; Gutiérrez, A.; Juanola, O. Bacterial Translocation as Inflammatory Driver in Crohn’s Disease. Front. Cell Dev. Biol. 2021, 9, 703310. [Google Scholar] [CrossRef]
- Zhu, L.; Qi, S.; Xue, X.; Niu, X.; Wu, L. Nitenpyram Disturbs Gut Microbiota and Influences Metabolic Homeostasis and Immunity in Honey Bee (Apis mellifera L.). Environ. Pollut. 2020, 258, 113671. [Google Scholar] [CrossRef]
- Nielsen, K.M.; Zhang, Y.; Curran, T.E.; Magnuson, J.T.; Venables, B.J.; Durrer, K.E.; Allen, M.S.; Roberts, A.P. Alterations to the Intestinal Microbiome and Metabolome of Pimephales promelas and Mus musculus following Exposure to Dietary Methylmercury. Environ. Sci. Technol. 2018, 52, 8774–8784. [Google Scholar] [CrossRef]
- Gao, B.; Chi, L.; Mahbub, R.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Multi-Omics Reveals That Lead Exposure Disturbs Gut Microbiome Development, Key Metabolites, and Metabolic Pathways. Chem. Res. Toxicol. 2017, 30, 996–1005. [Google Scholar] [CrossRef]
- Richardson, J.B.; Dancy, B.C.R.; Horton, C.L.; Lee, Y.S.; Madejczyk, M.S.; Xu, Z.Z.; Ackermann, G.; Humphrey, G.; Palacios, G.; Knight, R.; et al. Exposure to Toxic Metals Triggers Unique Responses from the Rat Gut Microbiota. Sci. Rep. 2018, 8, 6578. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Phyla | Species | References |
|---|---|---|
| Bacteriodetes | Bacteriodes, Prevoetella and Xlanibacter | [12] |
| Firmicutes | Ruminococcus, Clostridium; Lactobacillus and Enterococcus | [13] |
| Actinobacteria | Bifidobacterium | [14] |
| Proteobacteria | Escherichia and Enterobacteriaceae | [15] |
| Verrucomicrobia | Akkermansia muciniphila | [16] |
| Compound | Impact on Gut Microbes | References |
|---|---|---|
| Chlorpyrifos | Increased Proteobacteria population, decreased Bacteroidetes; significant impact on insulin resistance and obesity in test mice | [50] |
| Organochloride pesticides (p,p’-dichlorodiphenyldichloroethylene; β-hexachlorocyclohexane) | Significant alterations in composition of gut microbiome; extended effects on bile acid metabolism, with potential effects on human health | [51] |
| Organophosphates, glyphosate, pyrethroid | Altered composition in gut microbiota, significantly among Firmicutes, Bacteroidetes, and Lactobacilli | [52] |
| Pyrethroid (bifenthrin) | Dysbiosis in gut microbiota upon exposure to pesticides, significant impact on lipid metabolism, and development of non-alcoholic fatty liver disease in Xenopus laevis | [53] |
| Pyrethroid (deltamethrin, cypermethrin, and permethrin) | Altered gut microbiome in Aedes albopticus model, with decreased impact on Butyricimonas, Prevotellaceae, Anaerococcus, Pseudorhodobacter and enriched occurrence of Wolbachia, Chryseobacterium, and Pseudomonas | [54] |
| Carbendazim | Significant decrease in richness and diversity of gut microbiota, and observable decrease in populations of Bacteroidetes and an increase in Firmicutes, Proteobacteria, and Actinobacteria in mice models exposed to carbendazim, along with an simultaneous occurrence of lipid metabolism disorders in the mice | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; AlHussaini, K.I. Pesticides: Unintended Impact on the Hidden World of Gut Microbiota. Metabolites 2024, 14, 155. https://doi.org/10.3390/metabo14030155
Ali A, AlHussaini KI. Pesticides: Unintended Impact on the Hidden World of Gut Microbiota. Metabolites. 2024; 14(3):155. https://doi.org/10.3390/metabo14030155
Chicago/Turabian StyleAli, Asghar, and Khalid I. AlHussaini. 2024. "Pesticides: Unintended Impact on the Hidden World of Gut Microbiota" Metabolites 14, no. 3: 155. https://doi.org/10.3390/metabo14030155
APA StyleAli, A., & AlHussaini, K. I. (2024). Pesticides: Unintended Impact on the Hidden World of Gut Microbiota. Metabolites, 14(3), 155. https://doi.org/10.3390/metabo14030155