
Unraveling Metabolic Changes following Stroke: Insights from a Urinary Metabolomics Analysis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Stroke Patient Information and Urine Sample Collection
2.2. Clinical Measures
2.3. NMR Sample Preparation, Data Collection, and Post-Processing
2.4. Statistical Analysis
3. Results
3.1. Stroke Patient Characteristics
3.2. Metabolomic Analysis of Urine Samples
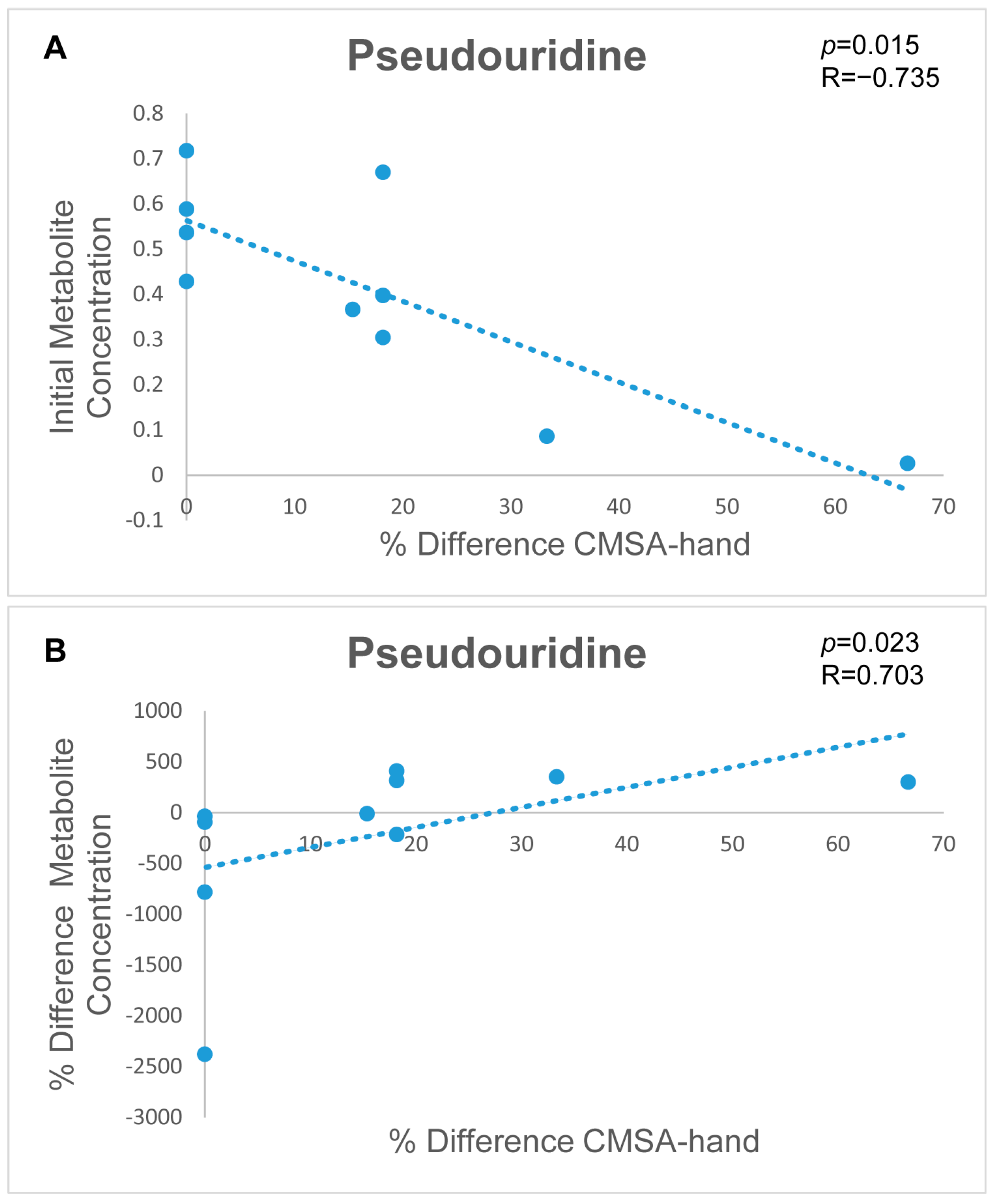
3.3. Correlation of Metabolomic Signatures Linked to Motor Recovery
4. Discussion
4.1. Clinical Translation and Classification of Stroke Metabolites
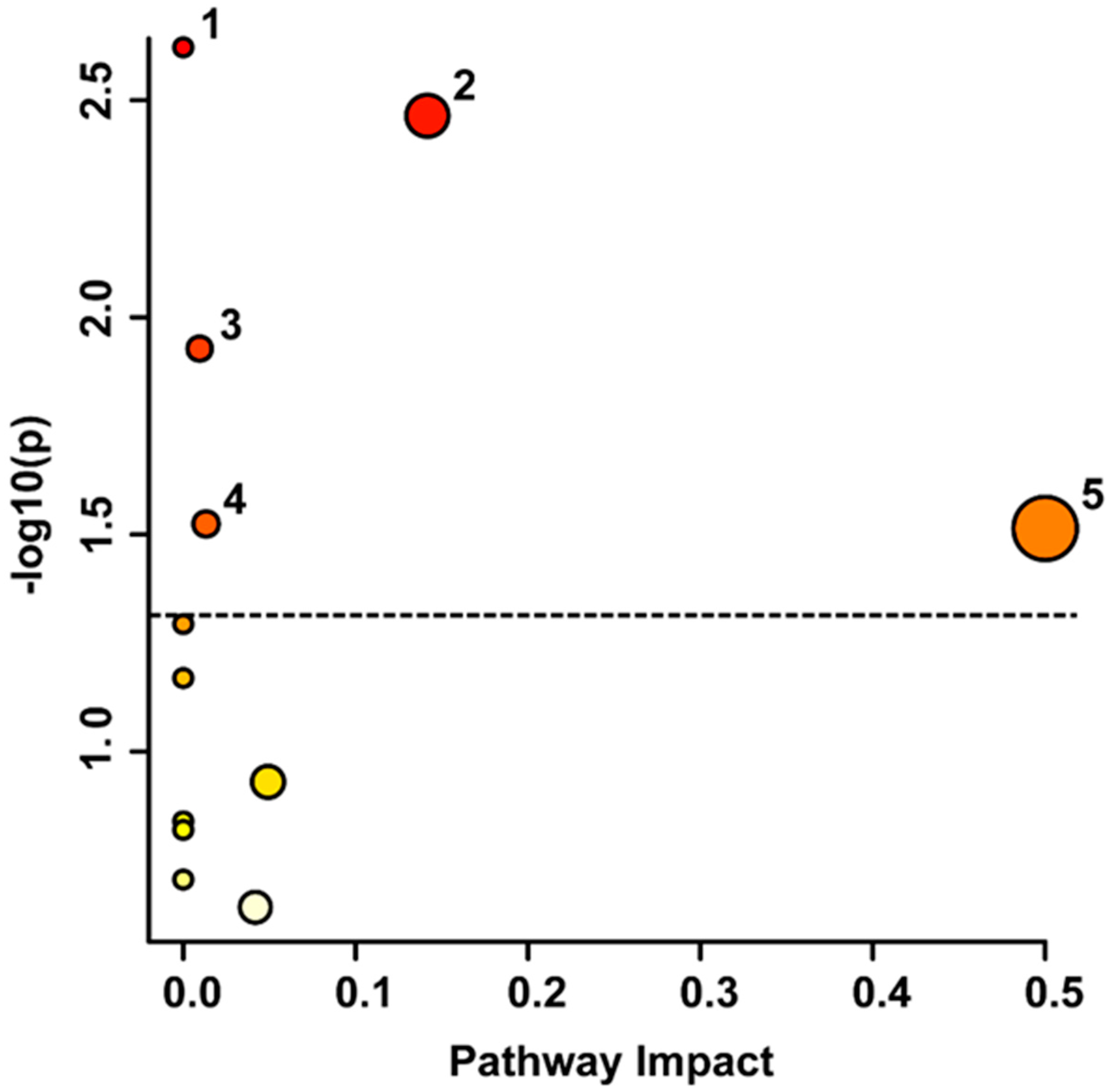
4.2. Metabolic Pathways Involved in the Stroke Recovery Process
4.2.1. Phenylalanine Pathways
4.2.2. Tyrosine Metabolism
4.2.3. Purine Metabolism
4.2.4. Glycerophospholipid Metabolism
4.3. The Relationship of Metabolic Biomarkers to Clinical Parameters
4.3.1. Metabolic Signatures to Predict Stroke Patient Outcomes
4.3.2. Metabolic Signatures to Monitor Stroke Patient Recovery
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global burden of cardiovascular diseases and risk factors, 1990–2019 update from the GBD 2019 study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Sacco, R.L.; Kasner, S.E.; Broderick, J.P.; Caplan, L.R.; Connors, J.J.; Culebras, A.; Elkind, M.S.V.; George, M.G.; Hamdan, A.D.; Higashida, R.T.; et al. An updated definition of stroke for the 21st century: A statement for healthcare professionals from the American heart Association/American stroke association. Stroke 2013, 44, 2064–2089. [Google Scholar] [CrossRef] [PubMed]
- Timpone, V.M.; Jensen, A.; Poisson, S.N.; Trivedi, P.S. Compliance with imaging guidelines for workup of transient ischemic attack evidence from the nationwide emergency department sample. Stroke 2020, 51, 2563–2567. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.C.V.; Khatri, P. Stroke. Lancet 2020, 396, 129. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.I.; Vorkas, P.A.; Coupland, A.P.; Jenkins, I.H.; Holmes, E.; Davies, A.H. Lessons from metabonomics on the neurobiology of stroke. SAGE Pub. 2017, 23, 374–382. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef]
- Emwas, A.M. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods. Mol. Biol. 2015, 1277, 161. [Google Scholar]
- Wishart, D.S. Quantitative metabolomics using NMR. Trends Analyt. Chem. 2008, 27, 228–237. [Google Scholar] [CrossRef]
- Ambeskovic, M.; Hopkins, G.; Hoover, T.; Joseph, J.T.; Montina, T.; Metz, G.A.S. Metabolomic signatures of Alzheimer’s disease indicate brain region-specific neurodegenerative progression. Int. J. Mol. Sci. 2023, 24, 14769. [Google Scholar] [CrossRef] [PubMed]
- Bykowski, E.A.; Petersson, J.N.; Dukelow, S.; Ho, C.; Debert, C.T.; Montina, T.; Metz, G.A.S. Urinary biomarkers indicative of recovery from spinal cord injury: A pilot study. IBRO Neurosci. Rep. 2021, 10, 178–185. [Google Scholar] [CrossRef]
- Bykowski, E.A.; Petersson, J.N.; Dukelow, S.; Ho, C.; Debert, C.T.; Montina, T.; Metz, G.A.S. Identification of serum metabolites as prognostic biomarkers following spinal cord injury: A pilot study. Metabolites 2023, 13, 605. [Google Scholar] [CrossRef]
- Bykowski, E.A.; Petersson, J.N.; Dukelow, S.; Ho, C.; Debert, C.T.; Montina, T.; Metz, G.A.S. Urinary metabolomic signatures as indicators of injury severity following traumatic brain injury: A pilot study. IBRO Neurosci. Rep. 2021, 11, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Paxman, E.J.; Boora, N.S.; Kiss, D.; Laplante, D.P.; King, S.; Montina, T.; Metz, G.A.S. Prenatal maternal stress from a natural disaster alters urinary metabolomic profiles in project ice storm participants. Sci. Rep. 2018, 8, 12932. [Google Scholar] [CrossRef]
- Wanner, Z.R.; Southam, C.G.; Sanghavi, P.; Boora, N.S.; Paxman, E.J.; Dukelow, S.P.; Benson, B.W.; Montina, T.; Metz, G.A.S.; Debert, C.T. Alterations in urine metabolomics following sport-related concussion: A 1H NMR-based analysis. Front. Neurol. 2021, 12, 645829. [Google Scholar] [CrossRef]
- Mollenhauer, B. Status of current biofluid biomarkers in parkinson’s disease. Mov. Disord. Clin. Pract. 2023, 10, S18–S20. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.J.; Colburn, W.A.; DeGruttola, V.G.; De Mets, D.L.; Downing, G.J.; Hoth, D.F.; Oates, J.A.; Peck, C.C.; Schooley, R.T.; Spilker, B.A.; et al. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Laborde, C.M.; Mourino-Alvarez, L.; Akerstrom, F.; Padial, L.R.; Vivanco, F.; Gil-Dones, F.; Barderas, M.G. Potential blood biomarkers for stroke. Expert Rev. Proteomics 2012, 9, 437–449. [Google Scholar] [CrossRef]
- Emwas, A.M.; Salek, R.M.; Griffin, J.L.; Merzaban, J. NMR-based metabolomics in human disease diagnosis: Applications, limitations, and recommendations. Metabolomics 2013, 9, 1048–1072. [Google Scholar] [CrossRef]
- Koen, N.; Du Preez, I.; Loots, D.T. Metabolomics and personalized medicine. Adv. Protein Chem. Struct. Biol. 2016, 102, 53–78. [Google Scholar] [CrossRef]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, H.; Kang, D.; Kim, N.S.; Cha, M.H.; Bang, O.; Ryu, D.H.; Hwang, G. 1H-NMR-based metabolomics study of cerebral infarction. Stroke 2011, 42, 1282. [Google Scholar] [CrossRef]
- Bonifačić, D.; Aralica, M.; Sotošek Tokmadžić, V.; Rački, V.; Tuškan-Mohar, L.; Kučić, N. Values of vanillylmandelic acid and homovanillic acid in the urine as potential prognostic biomarkers in ischaemic stroke patients. Biomarkers 2017, 22, 790–797. [Google Scholar] [CrossRef]
- Sidorov, E.; Bejar, C.; Xu, C.; Ray, B.; Reddivari, L.; Chainakul, J.; Vanamala, J.K.P.; Sanghera, D.K. Potential metabolite biomarkers for acute versus chronic stage of ischemic stroke: A pilot study. J. Stroke Cerebrovasc. Dis. 2020, 29, 104618. [Google Scholar] [CrossRef]
- Brott, T.; Adams, J.H.P.; Olinger, C.P.; Marler, J.R.; Barsan, W.G.; Biller, J.; Spilker, J.; Holleran, R.; Eberle, R.; Hertzberg, V.; et al. Measurements of acute cerebral infarction: A clinical examination scale. Stroke 1989, 20, 864–870. [Google Scholar] [CrossRef]
- Gowland, C.; Stratford, P.; Ward, M.; Moreland, J.; Torresin, W.; Vanhullenaar, S.; Sanford, J.; Barreca, S.; Vanspall, B.; Plews, N. Measuring physical impairment and disability with the chedoke-mcmaster stroke assessment. Stroke 1993, 24, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Keith, R.A.; Granger, C.V.; Hamilton, B.B.; Sherwin, F.S. The functional independence measure: A new tool for rehabilitation. Adv. Clin. Rehabil. 1987, 1, 6–18. [Google Scholar] [PubMed]
- Gil, R.B.; Lehmann, R.; Schmitt-Kopplin, P.; Heinzmann, S.S. 1H NMR-based metabolite profiling workflow to reduce inter-sample chemical shift variations in urine samples for improved biomarker discovery. Anal. Bioanal. 2016, 408, 4683–4691. [Google Scholar] [CrossRef]
- Smelter, A.; Rouchka, E.C.; Moseley, H.N.B. Detecting and accounting for multiple sources of positional variance in peak list registration analysis and spin system grouping. J. Biomol. NMR 2017, 68, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Vignoli, A.; Ghini, V.; Meoni, G.; Licari, C.; Takis, P.G.; Tenori, L.; Turano, P.; Luchinat, C. High-Throughput metabolomics by 1D NMR. Angew. Chem. Int. Ed. Engl. 2019, 58, 968–994. [Google Scholar] [CrossRef]
- Veselkov, K.A.; Lindon, J.C.; Ebbels, T.M.D.; Crockford, D.; Volynkin, V.V.; Holmes, E.; Davies, D.B.; Nicholson, J.K. Recursive segment-wise peak alignment of biological 1H NMR spectra for improved metabolic biomarker recovery. Anal. Chem. 2009, 81, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.E.; Mahle, D.A.; Doom, T.E.; Reo, N.V.; DelRaso, N.J.; Raymer, M.L. Dynamic adaptive binning: An improved quantification technique for NMR spectroscopic data. Metabolomics 2011, 7, 179–190. [Google Scholar] [CrossRef]
- Craig, A.; Cloarec, O.; Holmes, E.; Nicholson, J.K.; Lindon, J.C. Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal. Chem. 2006, 78, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, S.; Johansson, E.; Sjöström, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef]
- van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Box, G.E.P.; Cox, D.R. An analysis of transformations. J. R. Stat. Soc. 1964, 26, 211–252. [Google Scholar] [CrossRef]
- Dinga, R.; Schmaal, L.; Penninx, B.W.; Veltman, D.J.; Marquand, A.F. Controlling for effects of confounding variables on machine learning predictions. BioRxiv 2020. [Google Scholar] [CrossRef]
- Debik, J.; Sangermani, M.; Wang, F.; Madssen, T.S.; Giskeødegård, G.F. Multivariate analysis of NMR-based metabolomic data. NMR Biomed. 2022, 35, e4638. [Google Scholar] [CrossRef]
- Wanichthanarak, K.; Jeamsripong, S.; Pornputtapong, N.; Khoomrung, S. Accounting for biological variation with linear mixed-effects modelling improves the quality of clinical metabolomics data. Comput. Struct. Biotechnol. J. 2019, 17, 611–618. [Google Scholar] [CrossRef]
- Goodpaster, A.M.; Romick-Rosendale, L.E.; Kennedy, M.A. Statistical significance analysis of nuclear magnetic resonance-based metabonomics data. Anal. Biochem. 2010, 401, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Liang, F.; Deng, B.; Lai, G.; Vicente Gonçalves, C.M.; Lu, H.; Yan, J.; Huang, X.; Yi, L.; Liang, Y. Informative metabolites identification by variable importance analysis based on random variable combination. Metabolomics 2015, 11, 1539–1551. [Google Scholar] [CrossRef]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Szymanska, E.; Saccenti, E.; Smilde, A.K.; Westerhuis, J.A. Double-check: Validation of diagnostic statistics for PLS-DA models in metabolomics studies. Metabolomics 2012, 8, 3–16. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an optimized workflow for global metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef]
- Qi, B.; Zhang, Y.; Xu, B.; Zhang, Y.; Fei, G.; Lin, L.; Li, Q. Metabolomic characterization of acute ischemic stroke facilitates metabolomic biomarker discovery. Appl. Biochem. Biotechnol. 2022, 194, 5443–5455. [Google Scholar] [CrossRef]
- Chen, J.; Lv, Y.; Li, X.; Xiong, J.; Liang, H.; Xie, L.; Wan, C.; Chen, Y.; Wang, H.; Liu, P.; et al. Urinary metabolite signatures for predicting elderly stroke survivors with depression. Neuropsychiatr. Dis. Treat. 2021, 17, 925–933. [Google Scholar] [CrossRef]
- Liu, H.; Pu, J.; Zhou, Q.; Yang, L.; Bai, D. Peripheral blood and urine metabolites and biological functions in post-stroke depression. Metab. Brain Dis. 2022, 37, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.G.; Leipnitz, G.; Seminotti, B.; Amaral, A.U.; Zanatta, Â.; Vargas, C.R.; Dutra Filho, C.S.; Wajner, M. Experimental evidence that phenylalanine provokes oxidative stress in hippocampus and cerebral cortex of developing rats. Cell. Mol. Neurobiol. 2010, 30, 317–326. [Google Scholar] [CrossRef]
- Sanayama, Y.; Nagasaka, H.; Takayanagi, M.; Ohura, T.; Sakamoto, O.; Ito, T.; Ishige-Wada, M.; Usui, H.; Yoshino, M.; Ohtake, A.; et al. Experimental evidence that phenylalanine is strongly associated to oxidative stress in adolescents and adults with phenylketonuria. Mol. Genet. Metab. 2011, 103, 220–225. [Google Scholar] [CrossRef]
- Sitta, A.; Manfredini, V.; Biasi, L.; Treméa, R.; Schwartz, I.V.D.; Wajner, M.; Vargas, C.R. Evidence that DNA damage is associated to phenylalanine blood levels in leukocytes from phenylketonuric patients. Mutat. Res. 2009, 679, 13–16. [Google Scholar] [CrossRef]
- Kong, Y.; Feng, Y.; Lu, Y.; Feng, S.; Huang, Z.; Wang, Q.; Huang, H.; Ling, X.; Su, Z.; Guo, Y. Predictive serum biomarkers of patients with cerebral infarction. Neurol. Res. 2022, 44, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Molinoff, P.B.; Axelrod, J. Biochemistry of Catecholamines. Annu. Rev. Biochem. 1971, 40, 465–500. [Google Scholar] [CrossRef]
- Ormstad, H.; Verkerk, R.; Aass, H.C.D.; Amthor, K.; Sandvik, L. Inflammation-induced catabolism of tryptophan and tyrosine in acute ischemic stroke. J. Mol. Neurosci. 2013, 51, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Palkovits, M.; Kobayashi, R.; Kizer, J.; Jacobowitz, D.; Kopin, I. Effects of stress on catecholamines and tyrosine-hydroxylase activity of individual hypothalamic nuclei. Neuroendocrinology 1975, 18, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Eisenhofer, G.; Kopin, I.; Goldstein, D. Catecholamine metabolism: A contemporary view with implications for physiology and medicine. Pharmacol. Rev. 2004, 56, 331–349. [Google Scholar] [CrossRef]
- Xie, J.; Han, Y.; Hong, Y.; Li, W.; Pei, Q.; Zhou, X.; Zhang, B.; Wang, Y. Identification of potential metabolite markers for middle-aged patients with post-stroke depression using urine metabolomics. Neuropsychiatr. Dis. Treat. 2020, 16, 2017–2024. [Google Scholar] [CrossRef]
- Ke, C.; Pan, C.; Zhang, Y.; Zhu, X.; Zhang, Y. Metabolomics facilitates the discovery of metabolic biomarkers and pathways for ischemic stroke: A systematic review. Metabolomics 2019, 15, 152. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, J.; Zhong, D.; Li, G. Potential serum biomarkers and metabonomic profiling of serum in ischemic stroke patients using UPLC/Q-TOF MS/MS. PLoS ONE 2017, 12, e0189009. [Google Scholar] [CrossRef]
- Yu, F.; Li, X.; Feng, X.; Wei, M.; Luo, Y.; Zhao, T.; Xiao, B.; Xia, J. Phenylacetylglutamine, a novel biomarker in acute ischemic stroke. Front. Cardiovasc. Med. 2021, 8, 798765. [Google Scholar] [CrossRef]
- Dale, N.; Tian, F.; Sagoo, R.; Phillips, N.; Imray, C.; Roffe, C. Point-of-care measurements reveal release of purines into venous blood of stroke patients. Purinergic Signal. 2019, 15, 237–246. [Google Scholar] [CrossRef]
- Abbracchio, M.P.; Burnstock, G. Purinergic signalling: Pathophysiological roles. Jpn. J. Pharmacol. 1998, 78, 113–145. [Google Scholar] [CrossRef]
- Bell, M.; Kochanek, P.; Carcillo, J.; Mi, Z.; Schiding, J.; Wisniewski, S.R.; Clark, R.S.; Dixon, C.; Marion, D.; Jackson, E. Interstitial adenosine, inosine, and hypoxanthine are increased after experimental traumatic brain injury in the rat. J. Neurotrauma 1998, 15, 163–170. [Google Scholar] [CrossRef]
- Barsotti, C.; Ipata, P.L. Metabolic regulation of ATP breakdown and of adenosine production in rat brain extracts. Int. J. Biochem. Cell. Biol. 2004, 36, 2214–2225. [Google Scholar] [CrossRef]
- Burnstock, G. An introduction to the roles of purinergic signalling in neurodegeneration, neuroprotection and neuroregeneration. Neuropharmacology 2016, 104, 4–17. [Google Scholar] [CrossRef]
- Fredholm, B.B. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Johansson, S.; Wang, Y. Adenosine and the regulation of metabolism and body temperature. Adv. Pharmacol. 2011, 61, 77–94. [Google Scholar] [CrossRef]
- Mayne, M.; Fotheringham, J.; Yan, H.; Power, C.; Del Bigio, M.R.; Peeling, J.; Geiger, J.D. Adenosine A2A receptor activation reduces proinflammatory events and decreases cell death following intracerebral hemorrhage. Ann. Neurol. 2001, 49, 727–735. [Google Scholar] [CrossRef]
- Williams-Karnesky, R.; Stenzel-Poore, M. Adenosine and stroke: Maximizing the therapeutic potential of adenosine as a prophylactic and acute neuroprotectant. Curr. Neuropharmacol. 2009, 7, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Haskó, G.; Sitkovsky, M.V.; Szabó, C. Immunomodulatory and neuroprotective effects of inosine. Trends Pharmacol. Sci. 2004, 25, 152–157. [Google Scholar] [CrossRef]
- Benowitz, L.I.; Goldberg, D.E.; Madsen, J.R.; Soni, D.; Irwin, N. Inosine stimulates extensive axon collateral growth in the rat corticospinal tract after injury. Proc. Natl. Acad. Sci. USA 1999, 96, 13486–13490. [Google Scholar] [CrossRef]
- Chen, P.; Goldberg, D.E.; Kolb, B.; Lanser, M.; Benowitz, L.I. Inosine induces axonal rewiring and improves behavioral outcome after stroke. Proc. Natl. Acad. Sci. USA 2002, 99, 9031–9036. [Google Scholar] [CrossRef]
- Zai, L.; Ferrari, C.; Subbaiah, S.; Havton, L.A.; Coppola, G.; Strittmatter, S.; Irwin, N.; Geschwind, D.; Benowitz, L.I. Inosine alters gene expression and axonal projections in neurons contralateral to a cortical infarct and improves skilled use of the impaired limb. J. Neurosci. 2009, 29, 8187–8197. [Google Scholar] [CrossRef] [PubMed]
- Potter, P.; White, T. Release of adenosine 5′-triphosphate from synaptosomes from different regions of rat-brain. Neuroscience 1980, 5, 1351–1356. [Google Scholar] [CrossRef]
- Poelchen, W.; Sieler, D.; Wirkner, K.; Illes, P. Co-transmitter function of ATP in central catecholaminergic neurons of the rat. Neuroscience 2001, 102, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhou, T.; Yuan, J.; Xiong, X.; Liu, X.; Qiu, Z.; Hu, L.; Lu, H.; He, Q.; Liu, C.; et al. Metabolomics profiling to characterize cerebral ischemia-reperfusion injury in mice. Front. Pharmacol. 2023, 14, 1091616. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Glycerophospholipids in Brain: Their Metabolism, Incorporation into Membranes, Functions, and Involvement in Neurological Disorders. Chem. Phys. Lipids 2000, 106, 1–29. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, J.; Zhao, J.; Zhang, L.; Wang, Q.; Wang, G. Serum metabolomic patterns in young patients with ischemic stroke: A case study. Metabolomics 2021, 17, 24. [Google Scholar] [CrossRef]
- Jiang, W.; Chen, J.; Gong, L.; Liu, F.; Zhao, H.; Mu, J. Alteration of glycerophospholipid metabolism in hippocampus of post-stroke depression rats. Neurochem. Res. 2022, 47, 2052–2063. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Modulation of inflammation in brain: A matter of fat. J. Neurochem. 2007, 101, 577–599. [Google Scholar] [CrossRef]
- Haidar, N.; Carrara, M.; Andriamampandry, C.; Kanfer, J.; Freysz, L.; Dreyfus, H.; Massarelli, R. Incorporation of [h-3] ethanolamine into acetylcholine by a human cholinergic neuroblastoma clone. Neurochem. Res. 1994, 19, 9–13. [Google Scholar] [CrossRef]
- Smart, S.; Fox, G.; Allen, K.; Swanson, A.; Newman, M.; Swayne, G.; Clark, J.; Sales, K.; Williams, S. Identification of ethanolamine in rat and gerbil brain-tissue extracts by NMR-spectroscopy. NMR Biomed. 1994, 7, 356–365. [Google Scholar] [CrossRef]
- Meltzer, S.; Bagley, J.A.; Perez, G.L.; O’Brien, C.E.; DeVault, L.; Guo, Y.; Jan, L.Y.; Jan, Y. Phospholipid homeostasis regulates dendrite morphogenesis in drosophila sensory neurons. Cell Rep. 2017, 21, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Shen, F.; He, Y.; Degos, V.; Camus, M.; Maze, M.; Young, W.L.; Su, H. Activation of alpha-7 nicotinic acetylcholine receptor reduces ischemic stroke injury through reduction of pro-inflammatory macrophages and oxidative stress. PLoS ONE 2014, 9, e105711. [Google Scholar] [CrossRef]
- Li, X.; Ma, S.; Yi, C. Pseudouridine: The fifth RNA nucleotide with renewed interests. Curr. Opin. Chem. Biol. 2016, 33, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Maden, B.E. The numerous modified nucleotides in eukaryotic ribosomal RNA. Prog. Nucleic Acid Res. Mol. Biol. 1990, 39, 241. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X. A novel urinary metabolite signature for non-invasive post-stroke depression diagnosis. Cell Biochem. Biophys. 2015, 72, 661–667. [Google Scholar] [CrossRef]
- Ament, Z.; Patki, A.; Chaudhary, N.; Bhave, V.M.; Garcia Guarniz, A.; Gao, Y.; Gerszten, R.E.; Correa, A.; Judd, S.E.; Cushman, M.; et al. Nucleosides associated with incident ischemic stroke in the REGARDS and JHS cohorts. Neurology 2022, 98, e2097. [Google Scholar] [CrossRef]
- Bernhardt, J.; Hayward, K.S.; Kwakkel, G.; Ward, N.S.; Wolf, S.L.; Borschmann, K.; Krakauer, J.W.; Boyd, L.A.; Carmichael, S.T.; Corbett, D.; et al. Agreed definitions and a shared vision for new standards in stroke recovery research: The stroke recovery and rehabilitation roundtable taskforce. Neurorehabil. Neural Repair 2017, 31, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Dagonnier, M.; Donnan, G.A.; Davis, S.M.; Dewey, H.M.; Howells, D.W. Acute stroke biomarkers: Are we there yet? Front. Neurol. 2021, 12, 619721. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patient Code | Stroke Type | Vascular Territory | Affected Side | Sex | Age | Urine Collection (Days Post-Stroke) | Medications | Co-Morbidities | NIHSS | FIM | CMSA-Hand | CMSA-Arm | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | 6 Month | Initial | 6 Month | Initial | 6 Month | Initial | 6 Month | |||||||||
| ST-01 | Ischemic | MCA | Left | M | 79 | 2 | 242 | ASA, Atorvastatin, Clopidogrel, Docusate Sodium, Perindopril | A Fib, acute renal failure, motor-cycle accident 1952—Knocked out; right collar bone fracture, Left leg injury d/t combine accident as a teen | NaN | 109 | 123 | 5 | 6 | 5 | 7 |
| ST-03 | Ischemic | LACUNAR | Right | M | 37 | 5 | 221 | ASA, Synthroid, Rosuvastatin, HCTZ, Felodipine | Hypothyriodism, HTN, smoker (1/2 ppd × 5 years), EtOH | 11 | 92 | 120 | 1 | 2 | 1 | 3 |
| ST-05 | Ischemic | MCA | Left | M | 47 | 6 | 206 | ASA, Crestor | HTN, smoker (30 per day) | 1 | 116 | 125 | 5 | 7 | 7 | 7 |
| ST-06 | Ischemic | MCA | Left | M | 64 | 4 | 101 | Warfarin, Solatol, Metoprolol, Atorvastatin, Levothyroxine, Vit D, Calcium, Magnesium, Benzaclin Pump | A Fib, valve disease—dilated cardiac myopathy 1990, irregular heart rate in 2002, 2016, cardiac MRI performed, no clot found. | 3 | 105 | 115 | 5 | 5 | 6 | 5 |
| ST-08 | Ischemic | MCA/ACA | Left | M | 62 | 4 | 200 | Diabetes, smoker, increased cholesterol | 2 | 106 | 124 | 6 | 7 | 5 | 7 | |
| ST-09 | HEM | MCA | Left | F | 61 | 5 | 191 | Gravol, Gabapentin, Lovenox, Lantus, Norvasc, Pantoprazole, Restorlax, Aidactone, Vit D, Ativan, Advair | HTN, diabetes, asthma, Barrett’s esophagus, obstructive sleep apnea, chronic neck and low back pain—recurring PRP treatment | 9 | 80 | 124 | 5 | 6 | 4 | 7 |
| ST-10 | Ischemic | Thalamus | Left | M | 72 | 6 | 189 | ASA, Diamicron, Metoprolol, Fosinopril, Gliclazide, Plavix, Metformin, Atorvastatin, Vit D, Lantus, Humulin, Drug Study (Rivanrobran vs Placebo) | HTN, diabetes, hyperlipidemia, ischemic heart disease, CABG (Aug 2015), post-CABG enrolled in COMPESS, TIA on 11 January 2017 sent to stroke prevention clinic on 13 January 2017 and then admitted to acute stroke unit | 4 | 96 | 122 | 5 | 6 | 4 | 6 |
| ST-16 | HEM | MCA | Left | M | 62 | 8 | 213 | Nicotine, Amlodipine, Baclofen, Acetominaphen | HTN, hyperlipidemia, smoker, prostate cancer, degenerative changes spine, chronic sinusitis | 10 | 70 | 115 | 5 | 5 | 4 | 5 |
| ST-17 | Ischemic | MCA + ICA | Left | M | 53 | 11 | 221 | 6 | NaN | 126 | 6 | 6 | 6 | 6 | ||
| ST-19 | HEM | PCA | Left | F | 78 | 4 | 170 | Amlodipine, Enoxaparin, Hydrochlorothiazide, Nitro patch Polythylene Glycol | HTN, hyperlipidemia, cleft Palate | 4 | 113 | 123 | 7 | 7 | 7 | 7 |
| Metabolite | Chemical Shift (ppm) | VIAVC p-Value | Paired t/Wilcoxon p-Value | Regulation |
|---|---|---|---|---|
| Pseudouridine.1 | 4.299 | 4.55 × 10−23 | 0.0020 (W) | Down |
| 4-Hydroxy-3-Methoxymandelate | 3.887 | 8.19 × 10−19 | 0.3567 | Up |
| Inosine | 8.219 | 1.59 × 10−16 | 0.1496 | Down |
| Homovanillate | 3.874 | 7.57 × 10−16 | 0.5282 | Up |
| Adenosine | 4.315 | 7.82 × 10−16 | 0.269 | Down |
| 2-Aminobutyrate | 0.982 | 1.27 × 10−15 | 0.3256 | Down |
| Ethanolamine | 3.156 | 5.08 × 10−15 | 0.7229 | Up |
| Deoxyinosine | 6.494 | 3.41 × 10−13 | 0.6182 | Up |
| Phenylacetic acid.1 | 7.328 | Not Sig. | 0.0124 | Down |
| Phenylacetic acid.2 | 7.312 | Not Sig. | 0.0132 | Down |
| Acetylcholine | 2.157 | Not Sig. | 0.021 | Down |
| L-Tyrosine | 6.907 | Not Sig. | 0.0281 | Down |
| Anserine | 7.138 | Not Sig. | 0.0362 | Down |
| Pseudouridine.2 | 4.29 | Not Sig. | 0.0137 (W) | Down |
| Alanine | 1.493 | Not Sig. | 0.0371 (W) | Up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petersson, J.N.; Bykowski, E.A.; Ekstrand, C.; Dukelow, S.P.; Ho, C.; Debert, C.T.; Montina, T.; Metz, G.A.S. Unraveling Metabolic Changes following Stroke: Insights from a Urinary Metabolomics Analysis. Metabolites 2024, 14, 145. https://doi.org/10.3390/metabo14030145
Petersson JN, Bykowski EA, Ekstrand C, Dukelow SP, Ho C, Debert CT, Montina T, Metz GAS. Unraveling Metabolic Changes following Stroke: Insights from a Urinary Metabolomics Analysis. Metabolites. 2024; 14(3):145. https://doi.org/10.3390/metabo14030145
Chicago/Turabian StylePetersson, Jamie N., Elani A. Bykowski, Chelsea Ekstrand, Sean P. Dukelow, Chester Ho, Chantel T. Debert, Tony Montina, and Gerlinde A. S. Metz. 2024. "Unraveling Metabolic Changes following Stroke: Insights from a Urinary Metabolomics Analysis" Metabolites 14, no. 3: 145. https://doi.org/10.3390/metabo14030145
APA StylePetersson, J. N., Bykowski, E. A., Ekstrand, C., Dukelow, S. P., Ho, C., Debert, C. T., Montina, T., & Metz, G. A. S. (2024). Unraveling Metabolic Changes following Stroke: Insights from a Urinary Metabolomics Analysis. Metabolites, 14(3), 145. https://doi.org/10.3390/metabo14030145