

Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Serum and CSF Collection
2.3. Sample Preparation for LC-MS Analysis of Thyroid Hormone Metabolites from CSF and Serum
2.4. Chromatographic Conditions for Polar Metabolite Detection
2.5. Chromatographic Conditions for T3/T4 Detection with Standards and Biofluids
2.6. MS Data Acquisition Conditions for Targeted Analysis of Polar Metabolites and Thyroid Hormones
2.7. Targeted Metabolomics Data Analysis
2.8. Data Analysis for TH Levels
3. Results
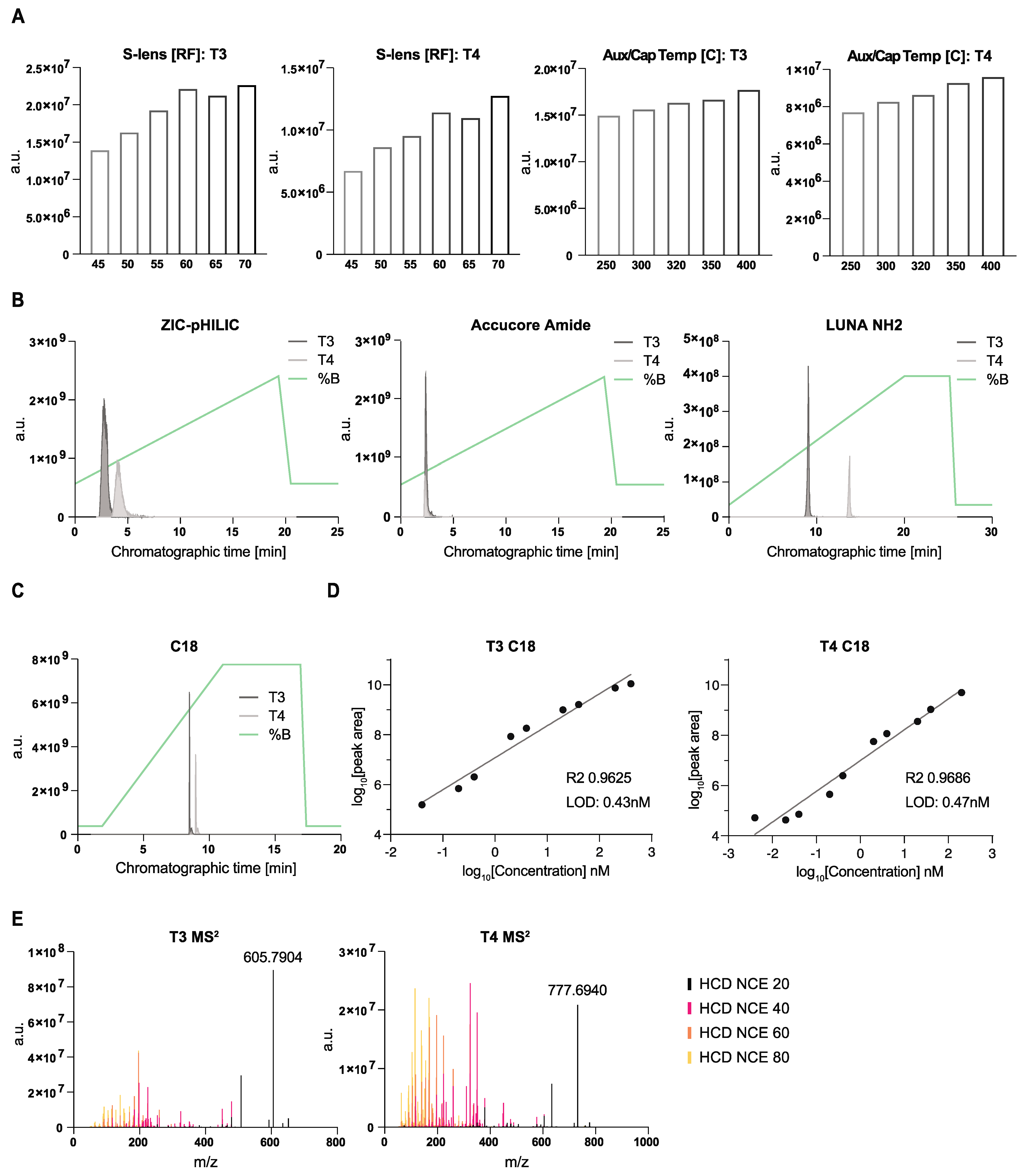
3.1. Optimization of LC-MS Conditions for the Detection of T3 and T4 Hormones in Mouse CSF
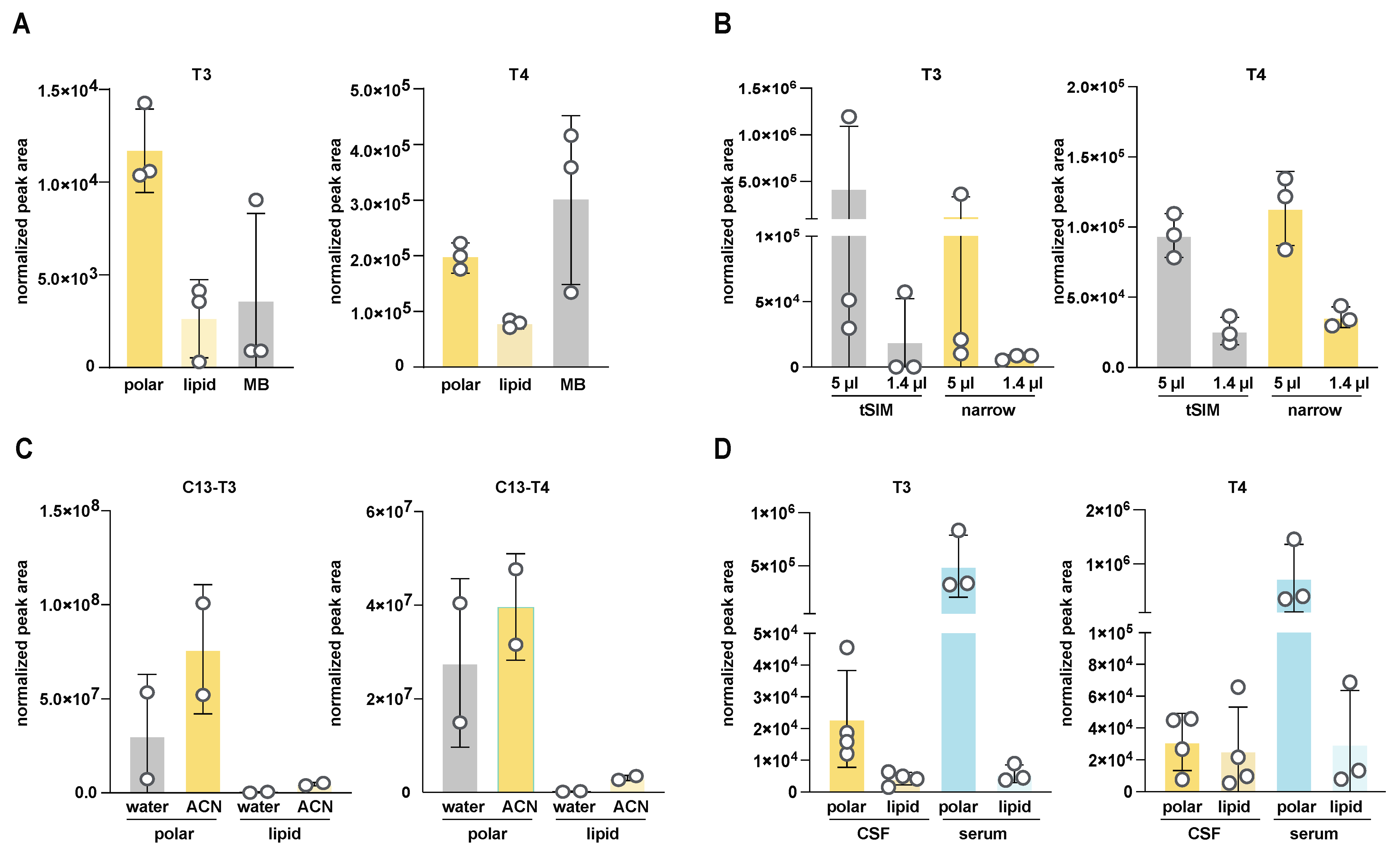
3.2. Optimization of CSF Sample Preparation for T3 and T4 Detection by LC-MS
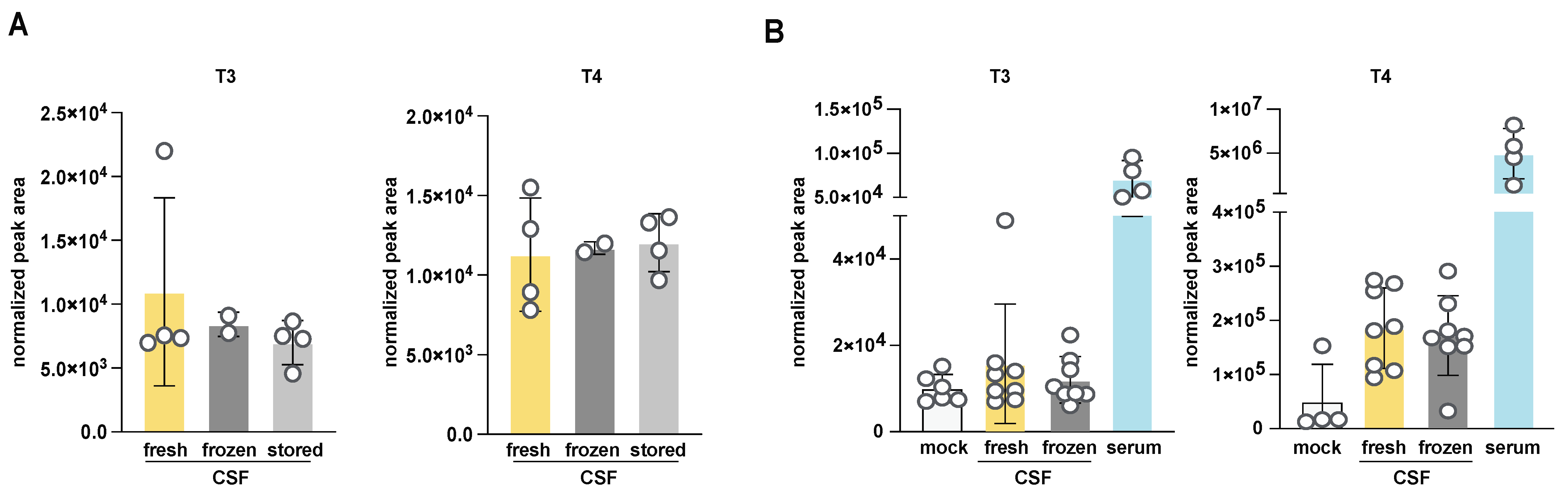
3.3. Investigation of the Effect of Handling and Storage Conditions on T3 and T4 Quantitation from Mouse CSF
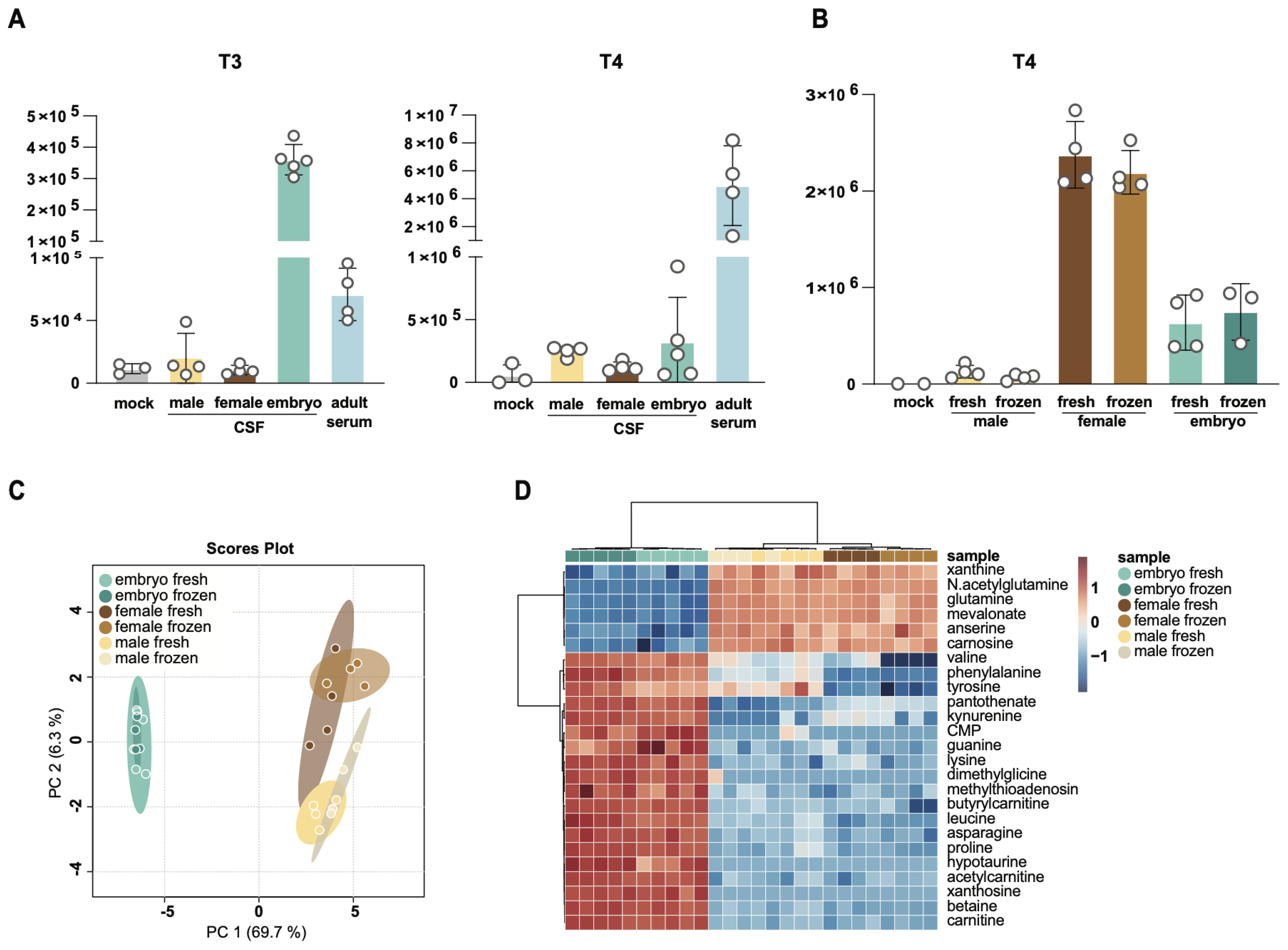
3.4. Comparison of T3 and T4 Levels in Adult and Embryonic CSF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kodding, R.; Fuhrmann, H.; Muhlen, A.V.Z. Investigations on Iodothyronine Deiodinase Activity in the Maturing Rat Brain. Endocrinology 1986, 118, 1347–1352. [Google Scholar] [CrossRef]
- Bernal, J.; Guadaño-Ferraz, A.; Morte, B. Thyroid hormone transporters—Functions and clinical implications. Nat. Rev. Endocrinol. 2015, 11, 406–417. [Google Scholar] [CrossRef]
- Mayerl, S.; Müller, J.; Bauer, R.; Richert, S.; Kassmann, C.M.; Darras, V.M.; Buder, K.; Boelen, A.; Visser, T.J.; Heuer, H. Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis. J. Clin. Investig. 2014, 124, 1987–1999. [Google Scholar] [CrossRef]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the Thyroid Hormone Transporters Monocarboxylate Transporter-8 (SLC16A2) and Organic Ion Transporter-14 (SLCO1C1) at the Blood-Brain Barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J.; Michals, E.A. Transport of thyroxine across the blood-brain barrier is directed primarily from brain to blood in the mouse. Life Sci. 1985, 37, 2407–2414. [Google Scholar] [CrossRef]
- Cserr, H.F.; Berman, B.J. Iodide and thiocyanate efflux from brain following injection into rat caudate nucleus. Am. J. Physiol.-Ren. Physiol. 1978, 235, F331–F337. [Google Scholar] [CrossRef]
- Davson, H.; Hollingsworth, J.R. Active transport of 131I across the blood—Brain barrier. J. Physiol. 1973, 233, 327–347. [Google Scholar] [CrossRef]
- Bárez-López, S.; Guadaño-Ferraz, A. Thyroid Hormone Availability and Action during Brain Development in Rodents. Front. Cell. Neurosci. 2017, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, D.; Wilsch-Bräuninger, M.; Wong, F.K.; Heuer, H.; Huttner, W.B. Integrin αvβ3 and thyroid hormones promote expansion of progenitors in embryonic neocortex. Development 2014, 141, 795–806. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Z.; Zhong, M.; Zhang, Y.; Li, M.; Zhang, L.; Qu, M.; Yang, J.; Wang, Y.; Yu, Z. Thyroid Hormone Promotes Neuronal Differentiation of Embryonic Neural Stem Cells by Inhibiting STAT3 Signaling Through TRα1. Stem Cells Dev. 2012, 21, 2667–2681. [Google Scholar] [CrossRef]
- Mohan, V.; Sinha, R.A.; Pathak, A.; Rastogi, L.; Kumar, P.; Pal, A.; Godbole, M.M. Maternal thyroid hormone deficiency affects the fetal neocorticogenesis by reducing the proliferating pool, rate of neurogenesis and indirect neurogenesis. Exp. Neurol. 2012, 237, 477–488. [Google Scholar] [CrossRef]
- Chantoux, F.; Francon, J. Thyroid hormone regulates the expression of NeuroD/BHF1 during the development of rat cerebellum. Mol. Cell. Endocrinol. 2002, 194, 157–163. [Google Scholar] [CrossRef]
- Espíndola, D.L.; Morales-Bastos, C.; Grijota-Martinez, C.; Liao, X.-H.; Lev, D.; Sugo, E.; Verge, C.F.; Refetoff, S.; Bernal, J.; Guadaño-Ferraz, A. Mutations of the Thyroid Hormone Transporter MCT8 Cause Prenatal Brain Damage and Persistent Hypomyelination. J. Clin. Endocrinol. Metab. 2014, 99, E2799–E2804. [Google Scholar] [CrossRef]
- Krieger, T.G.; Moran, C.M.; Frangini, A.; Visser, W.E.; Schoenmakers, E.; Muntoni, F.; Clark, C.A.; Gadian, D.; Chong, W.K.; Kuczynski, A.; et al. Mutations in thyroid hormone receptor α1 cause premature neurogenesis and progenitor cell depletion in human cortical development. Proc. Natl. Acad. Sci. USA 2019, 116, 22754–22763. [Google Scholar] [CrossRef]
- Martínez-Galán, J.R.; Pedraza, P.; Santacana, M.; del Ray, F.E.; de Escobar, G.M.; Ruiz-Marcos, A. Early effects of iodine deficiency on radial glial cells of the hippocampus of the rat fetus. A model of neurological cretinism. J. Clin. Investig. 1997, 99, 2701–2709. [Google Scholar] [CrossRef]
- Schreiber, G.; Aldred, A.R.; Jaworowski, A.; Nilsson, C.; Achen, M.G.; Segal, M.B. Thyroxine transport from blood to brain via transthyretin synthesis in choroid plexus. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1990, 258, R338–R345. [Google Scholar] [CrossRef]
- Richardson, S.J.; Wijayagunaratne, R.C.; D’Souza, D.G.; Darras, V.M.; Herck, S.L.J.V. Transport of thyroid hormones via the choroid plexus into the brain: The roles of transthyretin and thyroid hormone transmembrane transporters. Front. Neurosci. 2015, 9, 66. [Google Scholar] [CrossRef]
- Dratman, M.B.; Crutchfield, F.L.; Schoenhoff, M.B. Transport of iodothyronines from bloodstream to brain: Contributions by blood:brain and choroid plexus:cerebrospinal fluid barriers. Brain Res. 1991, 554, 229–236. [Google Scholar] [CrossRef]
- Chen, R.L.; Kassem, N.A.; Preston, J.E. Dose-dependent transthyretin inhibition of T4 uptake from cerebrospinal fluid in sheep. Neurosci. Lett. 2006, 396, 7–11. [Google Scholar] [CrossRef]
- Russell, W.; Harrison, R.F.; Smith, N.; Darzy, K.; Shalet, S.; Weetman, A.P.; Ross, R.J. Free Triiodothyronine Has a Distinct Circadian Rhythm That Is Delayed but Parallels Thyrotropin Levels. J. Clin. Endocrinol. Metab. 2008, 93, 2300–2306. [Google Scholar] [CrossRef]
- Fame, R.M.; Kalugin, P.N.; Petrova, B.; Xu, H.; Soden, P.A.; Shipley, F.B.; Dani, N.; Grant, B.; Pragana, A.; Head, J.P.; et al. Defining diurnal fluctuations in mouse choroid plexus and CSF at high molecular, spatial, and temporal resolution. Nat. Commun. 2023, 14, 3720. [Google Scholar] [CrossRef]
- Sabatino, L.; Vassalle, C.; Seppia, C.D.; Iervasi, G. Deiodinases and the Three Types of Thyroid Hormone Deiodination Reactions. J. Korean Endocr. Soc. 2021, 36, 952–964. [Google Scholar] [CrossRef]
- Bernal, J. Thyroid hormone regulated genes in cerebral cortex development. J. Endocrinol. 2017, 232, R83–R97. [Google Scholar] [CrossRef]
- Croteau, W.; Davey, J.C.; Galton, V.A.; Germain, D.L.S. Cloning of the mammalian type II iodothyronine deiodinase. A selenoprotein differentially expressed and regulated in human and rat brain and other tissues. J. Clin. Investig. 1996, 98, 405–417. [Google Scholar] [CrossRef]
- Leonard, J.L.; Kaplan, M.M.; Visser, T.J.; Silva, J.E.; Larsen, P.R. Cerebral Cortex Responds Rapidly to Thyroid Hormones. Science 1981, 214, 571–573. [Google Scholar] [CrossRef]
- Bernal, J. Thyroid Hormones and Brain Development. Vitam. Horm. 2005, 71, 95–122. [Google Scholar] [CrossRef]
- Zekri, Y.; Guyot, R.; Flamant, F. An Atlas of Thyroid Hormone Receptors’ Target Genes in Mouse Tissues. Int. J. Mol. Sci. 2022, 23, 11444. [Google Scholar] [CrossRef]
- Starling, S. Thyroid hormone action during development. Nat. Rev. Endocrinol. 2021, 17, 576. [Google Scholar] [CrossRef]
- Ruuskanen, S.; Hsu, B.-Y.; Heinonen, A.; Vainio, M.; Darras, V.M.; Sarraude, T.; Rokka, A. A new method for measuring thyroid hormones using nano-LC-MS/MS. J. Chromatogr. B 2018, 1093, 24–30. [Google Scholar] [CrossRef]
- Borsò, M.; Agretti, P.; Zucchi, R.; Saba, A. Mass spectrometry in the diagnosis of thyroid disease and in the study of thyroid hormone metabolism. Mass Spectrom. Rev. 2022, 41, 443–468. [Google Scholar] [CrossRef]
- Soldin, O.P.; Soldin, S.J. Thyroid hormone testing by tandem mass spectrometry. Clin. Biochem. 2011, 44, 89–94. [Google Scholar] [CrossRef]
- Soukhova, N.; Soldin, O.P.; Soldin, S.J. Isotope dilution tandem mass spectrometric method for T4/T3. Clin. Chim. Acta 2004, 343, 185–190. [Google Scholar] [CrossRef]
- Brabandere, V.I.D.; Hou, P.; Stöckl, D.; Thienpont, L.M.; Leenheer, A.P.D. Isotope dilution-liquid chromatography/electrospray ionization-tandem mass spectrometry for the determination of serum thyroxine as a potential reference method. Rapid Commun. Mass Spectrom. 1998, 12, 1099–1103. [Google Scholar] [CrossRef]
- Tai, S.S.-C.; Sniegoski, L.T.; Welch, M.J. Candidate reference method for total thyroxine in human serum: Use of isotope-dilution liquid chromatography-mass spectrometry with electrospray ionization. Clin. Chem. 2002, 48, 637–642. [Google Scholar] [CrossRef]
- Wang, D.; Stapleton, H.M. Analysis of thyroid hormones in serum by liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 397, 1831–1839. [Google Scholar] [CrossRef]
- Welsh, K.J.; Stolze, B.R.; Yu, X.; Podsiadlo, T.R.; Kim, L.S.; Soldin, S.J. Assessment of thyroid function in intensive care unit patients by liquid chromatography tandem mass spectrometry methods. Clin. Biochem. 2017, 50, 318–322. [Google Scholar] [CrossRef]
- Hagen, G.A.; Elliott, W.J. Transport of Thyroid Hormones in Serum and Cerebrospinal Fluid 1. J. Clin. Endocrinol. Metab. 1973, 37, 415–422. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef]
- Sawicka-Gutaj, N.; Zawalna, N.; Gut, P.; Ruchała, M. Relationship between thyroid hormones and central nervous system metabolism in physiological and pathological conditions. Pharmacol. Rep. 2022, 74, 847–858. [Google Scholar] [CrossRef]
- Seifert, J.; Chen, Y.; Schöning, W.; Mai, K.; Tacke, F.; Spranger, J.; Köhrle, J.; Wirth, E.K. Hepatic Energy Metabolism under the Local Control of the Thyroid Hormone System. Int. J. Mol. Sci. 2023, 24, 4861. [Google Scholar] [CrossRef]
- Cioffi, F.; Giacco, A.; Goglia, F.; Silvestri, E. Bioenergetic Aspects of Mitochondrial Actions of Thyroid Hormones. Cells 2022, 11, 997. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]
- Zappaterra, M.W.; LaMantia, A.S.; Walsh, C.A.; Lehtinen, M.K. Isolation of Cerebrospinal Fluid from Rodent Embryos for use with Dissected Cerebral Cortical Explants. J. Vis. Exp. 2013, e50333. [Google Scholar] [CrossRef]
- Manier, S.K.; Meyer, M.R. Impact of the used solvent on the reconstitution efficiency of evaporated biosamples for untargeted metabolomics studies. Metabolomics 2020, 16, 34. [Google Scholar] [CrossRef]
- Lindahl, A.; Sa, S.; Lehtio, J.; Nordstro, A. Tuning Metabolome Coverage in Reversed Phase LC–MS Metabolomics of MeOH Extracted Samples Using the Reconstitution Solvent Composition. Anal. Chem. 2017, 89, 7356–7364. [Google Scholar] [CrossRef]
- Fantz, C.R.; Dagogo-Jack, S.; Ladenson, J.H.; Gronowski, A.M. Thyroid function during pregnancy. Clin. Chem. 1999, 45, 2250–2258. [Google Scholar] [CrossRef]
- Refetoff, S.; Robin, N.I.; Fang, V.S. Parameters of Thyroid Function in Serum of 16 Selected Vertebrate Species: A Study of PBI, Serum T4, Free T4, and the Pattern of T4 and T3 Binding to Serum Proteins. Endocrinology 1970, 86, 793–805. [Google Scholar] [CrossRef]
- Freeman, T.; Pearson, J.D. The use of quantitative immunoelectrophoresis to investigate thyroxine-binding human serum proteins. Clin. Chim. Acta 1969, 26, 365–368. [Google Scholar] [CrossRef]
- Refetoff, S. Thyroid Hormone Serum Transport Proteins—Endotext—NCBI Bookshelf. Available online: https://www.ncbi.nlm.nih.gov/books/NBK285566/ (accessed on 20 November 2023).
- Kiebooms, J.A.L.; Wauters, J.; Bussche, J.V.; Vanhaecke, L. Validated ultra high performance liquid chromatography-tandem mass spectrometry method for quantitative analysis of total and free thyroid hormones in bovine serum. J. Chromatogr. A 2014, 1345, 164–173. [Google Scholar] [CrossRef]
- Tanoue, R.; Kume, I.; Yamamoto, Y.; Takaguchi, K.; Nomiyama, K.; Tanabe, S.; Kunisue, T. Determination of free thyroid hormones in animal serum/plasma using ultrafiltration in combination with ultra-fast liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2018, 1539, 30–40. [Google Scholar] [CrossRef]
- Vulsma, T.; Gons, M.H.; de Vijlder, J.J.M. Maternal-Fetal Transfer of Thyroxine in Congenital Hypothyroidism Due to a Total Organification Defect or Thyroid Agenesis. N. Engl. J. Med. 1989, 321, 13–16. [Google Scholar] [CrossRef]
- Landers, K.; Richard, K. Traversing barriers—How thyroid hormones pass placental, blood-brain and blood-cerebrospinal fluid barriers. Mol. Cell. Endocrinol. 2017, 458, 22–28. [Google Scholar] [CrossRef]
- Friesema, E.C.H.; Ganguly, S.; Abdalla, A.; Fox, J.E.M.; Halestrap, A.P.; Visser, T.J. Identification of Monocarboxylate Transporter 8 as a Specific Thyroid Hormone Transporter. J. Biol. Chem. 2003, 278, 40128–40135. [Google Scholar] [CrossRef]
- Funkquist, A.; Bengtsson, A.; Johansson, P.; Svensson, J.; Bjellerup, P.; Blennow, K.; Wandt, B.; Sjöberg, S. Low CSF/serum ratio of free T4 is associated with decreased quality of life in mild hypothyroidism—A pilot study. J. Clin. Transl. Endocrinol. 2020, 19, 100218. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fame, R.M.; Ali, I.; Lehtinen, M.K.; Kanarek, N.; Petrova, B. Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid. Metabolites 2024, 14, 79. https://doi.org/10.3390/metabo14020079
Fame RM, Ali I, Lehtinen MK, Kanarek N, Petrova B. Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid. Metabolites. 2024; 14(2):79. https://doi.org/10.3390/metabo14020079
Chicago/Turabian StyleFame, Ryann M., Ilhan Ali, Maria K. Lehtinen, Naama Kanarek, and Boryana Petrova. 2024. "Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid" Metabolites 14, no. 2: 79. https://doi.org/10.3390/metabo14020079
APA StyleFame, R. M., Ali, I., Lehtinen, M. K., Kanarek, N., & Petrova, B. (2024). Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid. Metabolites, 14(2), 79. https://doi.org/10.3390/metabo14020079