
Fungal-Mediated Biotransformation of the Plant Growth Regulator Forchlorfenuron by Cunninghamella elegans


 ,
, 
Abstract
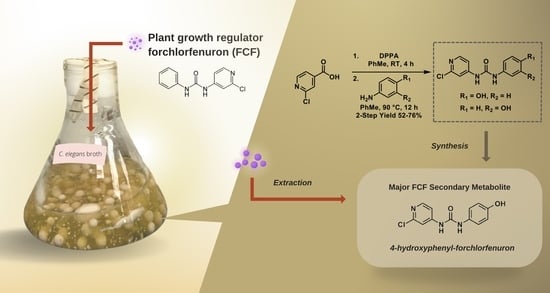
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Microorganisms and Culture Medium
2.3. Culture and Biotransformation Conditions
2.3.1. Standard Conditions
2.3.2. Optimized Conditions
2.4. Extraction and Isolation
2.5. Identification of Metabolites
2.6. Synthesis of Metabolites
2.6.1. General Procedure for Preparation of Reference Standards
2.6.2. Characterization Data


3. Results and Discussion
3.1. Rapid Screening Results from Fungal Panel under Standard Conditions
3.2. Optimization of Biotransformation Conditions
3.2.1. Solid Support System
3.2.2. Media Screen
3.2.3. Inoculation with Solid Pre-Formed Fungal Mass of C. elegans
3.2.4. Optimization Analysis and Percentage Change
3.3. Identification and Structural Elucidation of the Isolated Metabolite
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kunhorm, P.; Chueaphromsri, P.; Chaicharoenaudomrung, N.; Noisa, P. Enhancement of cordycepin production from Cordyceps militaris culture by epigenetic modification. Biotechnol. Lett. 2022, 44, 581–593. [Google Scholar] [CrossRef]
- Shan, T.; Zhang, X.; Guo, C.; Guo, S.; Zhao, X.; Yuan, Y.; Yue, T. Identity, Synthesis, and Cytotoxicity of Forchlorfenuron Metabolites in Kiwifruit. J. Agric. Food Chem. 2021, 69, 9529–9535. [Google Scholar] [CrossRef]
- Aremu, A.; Fawole, O.; Makunga, N.; Masondo, N.; Moyo, M.; Buthelezi, N.; Amoo, S.; Spíchal, L.; Doležal, K. Applications of Cytokinins in Horticultural Fruit Crops: Trends and Future Prospects. Biomolecules 2020, 10, 1222. [Google Scholar] [CrossRef]
- Fathy, M.; Saad Eldin, S.M.; Naseem, M.; Dandekar, T.; Othman, E.M. Cytokinins: WideSpread Signaling Hormones from Plants to Humans with High Medical Potential. Nutrients 2022, 14, 1495. [Google Scholar] [CrossRef]
- Henzi, T.; Diep, K.-L.; Oberson, A.; Salicio, V.; Bochet, C.G.; Schwaller, B. Forchlorfenuron and Novel Analogs Cause Cytotoxic Effects in Untreated and CisplatinResistant Malignant Mesothelioma-Derived Cells. Int. J. Mol. Sci. 2022, 23, 3963. [Google Scholar] [CrossRef]
- Lappas, C.M. The Plant Hormone Zeatin Riboside Inhibits Tlymphocyte Activity via Adenosine A2A Receptor Activation. Cell. Mol. Immunol. 2014, 12, 107–112. [Google Scholar] [CrossRef][Green Version]
- Gonzalez, G.; Grúz, J.; D’Acunto, C.W.; Kanovský, P.; Strnad, M. Cytokinin Plant Hormones Have Neuroprotective Activity in In Vitro Models of Parkinson’s Disease. Molecules 2021, 26, 361. [Google Scholar] [CrossRef]
- Valverde, A.; Aguilera, A.; Ferrer, C.; Camacho, F.; Cammarano, A. Analysis of forchlorfenuron in vegetables by LC/TOF-MS after extraction with the buffered QuEChERS method. J. Agric. Food Chem. 2010, 58, 2818–2823. [Google Scholar] [CrossRef]
- Sosnowski, J.; Truba, M.; Vasileva, V. The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops. Agriculture 2023, 13, 724. [Google Scholar] [CrossRef]
- US Environmental Protection Agency, Office of Prevention, Pesticides and Toxic Substances. Pesticide Fact Sheet [EB/OL]. U.S.EPA. 2004. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-128819_01-Sep-04.pdf (accessed on 14 June 2023).
- Zhang, Z.; Guo, K.; Bai, Y.; Dong, J.; Gao, Z.; Yuan, Y.; Wang, Y.; Liu, L.; Yue, T. Identification, Synthesis, and Safety Assessment of Forchlorfenuron (1-(2-Chloro-4-pyridyl)-3-phenylurea) and Its Metabolites in Kiwifruits. J. Agric. Food Chem. 2015, 63, 3059–3066. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Zurowski, C.; Looney, N.E. Phenylureas CPPU and thidiazuron affect yield components, fruit composition, and storage potential of four seedless grape selections. J. Am. Soc. Hortic. Sci. 1992, 117, 85–89. [Google Scholar] [CrossRef]
- Blank, R.H.; Richardson, A.C.; Oshima, K.; Hampton, R.E.; Olson, M.H.; Dawson, T.E. Effect of a forchlorfenuron dip on kiwifruit fruit size. N. Z. J. Crop Hortic. Sci. 1992, 20, 73–78. [Google Scholar] [CrossRef]
- Shi, H.; Liao, J.; Cui, S.; Luo, Z.; Ma, X. Effects of Forchlorfenuron on the Morphology, Metabolite Accumulation, and Transcriptional Responses of Siraitia grosvenorii Fruit. Molecules 2019, 24, 4076. [Google Scholar] [CrossRef]
- Matsumoto, K.; Fujita, T.; Sato, S. Exogenous applications of plant growth regulators improve quality of ‘Fuji’ apple. Int. J. Agric. Biol. 2018, 20, 2083–2090. [Google Scholar]
- Bu, Q.; Wang, X.; Xie, H.; Zhong, K.; Wu, Y.; Zhang, J.; Wang, Z.; Gao, H.; Huang, Y. 180 Day repeated-dose toxicity study on forchlorfenuron in sprague-dawley rats and its effects on the production of steroid hormones. J. Agric. Food Chem. 2019, 67, 10207–10213. [Google Scholar] [CrossRef]
- Gong, G.; Kam, H.; Tse, Y.; Lee, S.M. Cardiotoxicity of forchlorfenuron (CPPU) in zebrafish (Danio rerio) and H9c2 cardiomyocytes. Chemosphere 2019, 235, 153–162. [Google Scholar] [CrossRef]
- Vardi-Oknin, D.; Golan, M.; Mabjeesh, N. Forchlorfenuron disrupts SEPT9_i1 filaments and inhibits HIF-1. PLoS ONE 2013, 8, e73179. [Google Scholar] [CrossRef]
- Guiyi, G.; Hiotong, K.; Yu-Chung, T.; Giesy, J.; Sai-Wang, S.; Simon, M. Forchlorfenuron (CPPU) causes disorganization of the cytoskeleton and dysfunction of human umbilical vein endothelial cells, and abnormal vascular development in zebrafish embryos. Environ. Pollut. 2021, 271, 115791. [Google Scholar] [CrossRef]
- Ping, L.; Xu, B.; Zhou, Q.; Hong, Y.; Sun, Q.; Wang, J.; Zhu, D. Comparative Pharmacokinetic Study of Forchlorfenuron in Adult and Juvenile Rats. Molecules 2021, 26, 4276. [Google Scholar] [CrossRef]
- Yan, B.; Guo, W. A novel identification method for CPPU—Treated kiwifruits based on images. J. Sci. Food Agric. 2019, 99, 6234–6240. [Google Scholar] [CrossRef]
- Shi, X.M.; Jin, F.; Huang, Y.T.; Du, X.W.; Li, C.M.; Wang, M.; Shao, H.; Jin, M.J.; Wang, J. Simultaneous determination of five plant growth regulators in fruits by modified quick, easy, cheap, effective, rugged, and safe (QuEChERS) extraction and liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2012, 60, 60–65. [Google Scholar] [CrossRef]
- Yavuz, K.A.; Yakar, B. Genotoxicity of a synthetic plant growth regulator, Forchlorfenuron (CPPU), on human lymphocytes using chromosome aberration assay. Caryologia 2023, 76, 47–56. [Google Scholar] [CrossRef]
- Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; Marques, D.C.; et al. EFSA Peer review of the pesticide risk assessment of the active substance forchlorfenuron. EFSA J. 2017, 15, 4874. [Google Scholar] [CrossRef]
- Trevor, H. Public Release Summary on Evaluation of the New Active Forchlorfenuron in the Product Sitofex 10EC Plant Growth Regulator; Australian Pesticides and Veterinary Medicines Authority: Symonston, Australia, 2005. Available online: https://apvma.gov.au/sites/default/files/publication/13791-prs-forchlorfenuron.pdf (accessed on 27 December 2022).
- Asha, S.; Vidyavathi, M. Cunninghamella—A Microbial Model for Drug Metabolism Studies—A Review. Biotechnol. Adv. 2009, 27, 16–29. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Wang, W.W.; Lee, S.S.; Gao, J.M. Microbial Transformations of Two BeyeraneType Diterpenes by Cunninghamella echinulata. J. Agric. Food Chem. 2020, 68, 4624–4631. [Google Scholar] [CrossRef]
- Zhu, Y.Z.; In-Hong Jeong, M.F.; Kim, J.H.; Zhang, C.J. Metabolism of an Insecticide Fenitrothion by Cunninghamella elegans ATCC36112. J. Agric. Food Chem. 2017, 65, 10711–10718. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mobley, R.; Woodfine, S.; Drijfhout, F.; Horrocks, P.; Ren, X.D.; Li, W.W. Biotransformation of artemisinin to a novel derivative via ring rearrangement by Aspergillus niger. Appl. Microbiol. Biotechnol. 2022, 106, 2433–2444. [Google Scholar] [CrossRef]
- Sponchiado, R.; Sorrentino, J.; Cordenonsi, L.M.; Fuentefria, A.M.; Steppe, M.; Mendez, A.; Garcia, C.V. Rifampicin: Biotransformation Study Using the Fungus Cunninghamella elegans and Monitoring through UHPLC-MS. Drug Anal. Res. 2020, 4, 44–48. [Google Scholar] [CrossRef]
- Wickerham, J. A taxonomic study of Monilia albicans with special emphasis on morphology and morphological variation. J. Trop. Med. Hyg. 1939, 42, 174–179. [Google Scholar]
- Wickerham, J. Taxonomy of Yeasts; Technical Bulletin; US Department of Agriculture: Washington, DC, USA, 1951; Volume 1029, pp. 1–56.
- Parshikov, I.A.; Muraleedharan, K.M.; Avery, M.A.; Williamson, J.S. Transformation of artemisinin by Cunninghamella elegans. Appl. Microbiol. Biotechnol. 2004, 64, 782–786. [Google Scholar] [CrossRef]
- Kang, S.-I.; Kang, S.-Y.; Hur, H. Identification of Fungal Metabolites of Anticonvulsant Drug Carbamazepine. Appl. Microbiol. Biotechnol. 2008, 79, 663. [Google Scholar] [CrossRef]
- Grafinger, K.; Wilke, A.; Konig, S.; Weinmann, W. Investigating the Ability of the Microbial Model Cunninghamella elegans for the Metabolism of Synthetic Tryptamines. Drug Test Anal. 2019, 11, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Murphy, C. Nitroreduction of Flutamide by Cunninghamella elegans NADPH: Cytochrome P450 Reductase. Biochem. Biophys. Rep. 2022, 29, 101209. [Google Scholar] [CrossRef] [PubMed]
- Khaled, O.; Galal, A.; Ibrahim, A.R.; El-Feraly, F.; Khalifa, S.; El-Sohly, H. Microbial metabolism of artemisitene. Phytochemistry 1999, 51, 257–261. [Google Scholar] [CrossRef]
- Parshikov, I.A.; Moody, J.D.; Heinze, T.M.; Freeman, J.P.; Williams, A.J.; Sutherland, J.B. Transformation of cinoxacin by Beauveria bassiana. FEMS Microbiol. Lett. 2002, 214, 133–136. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Diez, D.; Sanchez, J.M.; Rodilla, J.M.; Rocha, P.M.; Mendes, R.S.; Paulino, C.; Marcos, I.S.; Basabe, P.; Urones, J.G. Microbial Hydroxylation of Sclareol by Rhizopus Stolonifer. Molecules 2005, 10, 1005–1009. [Google Scholar] [CrossRef]
- Amadio, J.; Casey, E.; Murphy, C.D. Filamentous fungi biofilm for production of human drug metabolites. Appl. Microbiol. Biotechnol. 2013, 97, 5955–5963. [Google Scholar] [CrossRef]
- Sutherland, J.B.; Freeman, J.P.; Williams, A.J.; Deck, J. Metabolism of cinnoline to N-oxidation products by Cunninghamella elegans and Aspergillus niger. J. Ind. Microbiol. Biotechnol. 1998, 21, 225–227. [Google Scholar] [CrossRef]
- Nykodemova, J.; Sulakova, A.; Palivec, P.; Ceskova, H.; Rimpelova, S.; Sichova, K.; Leonhardt, T.; Jurasek, B.; Hajkova, K.; Palenicek, T.; et al. 2C-B-Fly-NBOMe Metabolites in Rat Urine, Human Liver Microsomes and C. elegans: Confirmation with Synthesized Analytical Standards. Metabolites 2021, 11, 775. [Google Scholar] [CrossRef]
- Amobonye, A. Biotechnological potential of Beauveria bassiana as a source of novel bio catalysts and metabolites. Crit. Rev. Biotechnol. 2020, 40, 1019–1034. [Google Scholar] [CrossRef]
- Zhang, Q.; Lin, L.; Ye, W. Techniques for extraction and isolation of natural products: A Comprehensive Review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Toyohara, J.; Gogami, A.; Hayashi, A.; Yonekura, Y.; Fujibayashi, Y. Pharmacokinetics and Metabolism of 5-125-I-Iodo-4’-Thio-2’-Deoxyuridine in Rodents. J. Nucl. Med. 2003, 44, 1671–1676. [Google Scholar]
- Boruta, T. Uncovering the repertoire of fungal secondary metabolites: From Fleming’s laboratory to the International Space Station. Bioengineered 2018, 9, 12–16. [Google Scholar] [CrossRef]
- Wang, Q.; Su, H.; Yue, N.; Li, M.; Li, C.; Wang, J.; Jin, F. Dissipation and risk assessment of forchlorfenuron and its major metabolites in oriental melon under greenhouse cultivation. Ecotoxicol. Environ. Saf. 2021, 225, 112700. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, Z.; Yuan, Y.; Wang, Y.; Yue, T. Determination of forchlorfenuron and its metabolites in kiwifruit using QuEChERS-LC-MS/MS. Mod. Food Sci. Technol. 2017, 33, 268–273. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Yeast Malt Broth (in L or g) | Potato Dextrose Broth (in L or g) |
|---|---|---|
| Distilled Water | 1 L | 1 L |
| Bacto Peptone | 5 g | --- |
| Yeast Extract | 3 g | --- |
| Malt Extract | 3 g | --- |
| Potato Extract | --- | 4 g |
| Dextrose | 10 g | 20 g |
| Compound | Formula | Retention Time (min) | [M + H] + m/z | Type of Transformation |
|---|---|---|---|---|
FCF 1 | C12H10ClN3O | 3.94 | 247.7 | — |
4-OH-FCF 2 | C12H10ClN3O2 | 2.63 | 263.7 | hydroxylation |
3-OH-FCF 3,* | C12H10ClN3O2 | 3.38 | 263.7 | hydroxylation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, C.M.; Moreno, J.N.; Valdez, M.C.; Baldwin, M.P.; Vallor, A.C.; Carvalho, P.B. Fungal-Mediated Biotransformation of the Plant Growth Regulator Forchlorfenuron by Cunninghamella elegans. Metabolites 2024, 14, 101. https://doi.org/10.3390/metabo14020101
Moreno CM, Moreno JN, Valdez MC, Baldwin MP, Vallor AC, Carvalho PB. Fungal-Mediated Biotransformation of the Plant Growth Regulator Forchlorfenuron by Cunninghamella elegans. Metabolites. 2024; 14(2):101. https://doi.org/10.3390/metabo14020101
Chicago/Turabian StyleMoreno, Charles M., Jaclyn N. Moreno, Matthew C. Valdez, Melinda P. Baldwin, Ana C. Vallor, and Paulo B. Carvalho. 2024. "Fungal-Mediated Biotransformation of the Plant Growth Regulator Forchlorfenuron by Cunninghamella elegans" Metabolites 14, no. 2: 101. https://doi.org/10.3390/metabo14020101
APA StyleMoreno, C. M., Moreno, J. N., Valdez, M. C., Baldwin, M. P., Vallor, A. C., & Carvalho, P. B. (2024). Fungal-Mediated Biotransformation of the Plant Growth Regulator Forchlorfenuron by Cunninghamella elegans. Metabolites, 14(2), 101. https://doi.org/10.3390/metabo14020101