
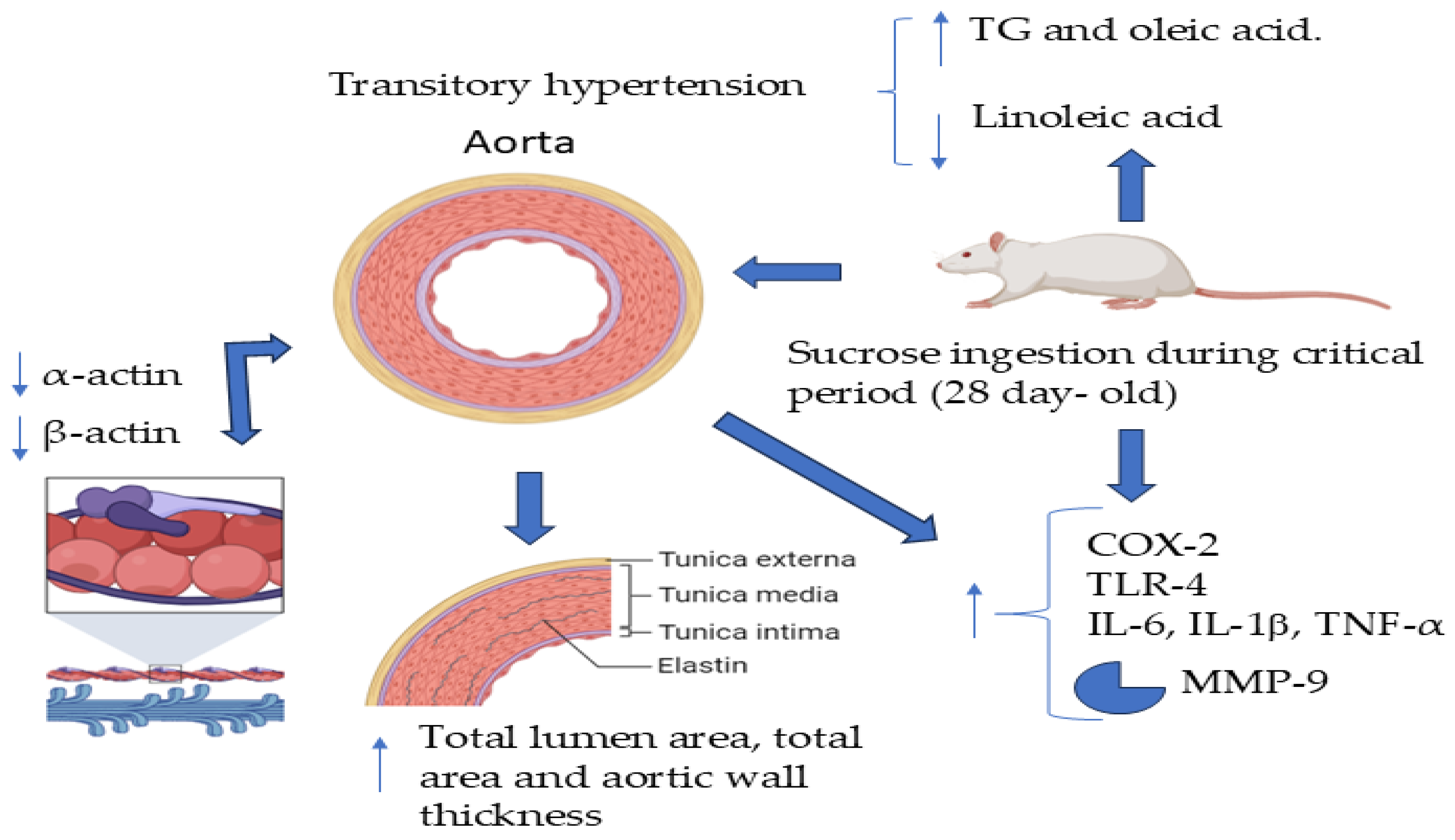
Rats Exposed to Excess Sucrose During a Critical Period Develop Inflammation and Express a Secretory Phenotype of Vascular Smooth Muscle Cells


 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
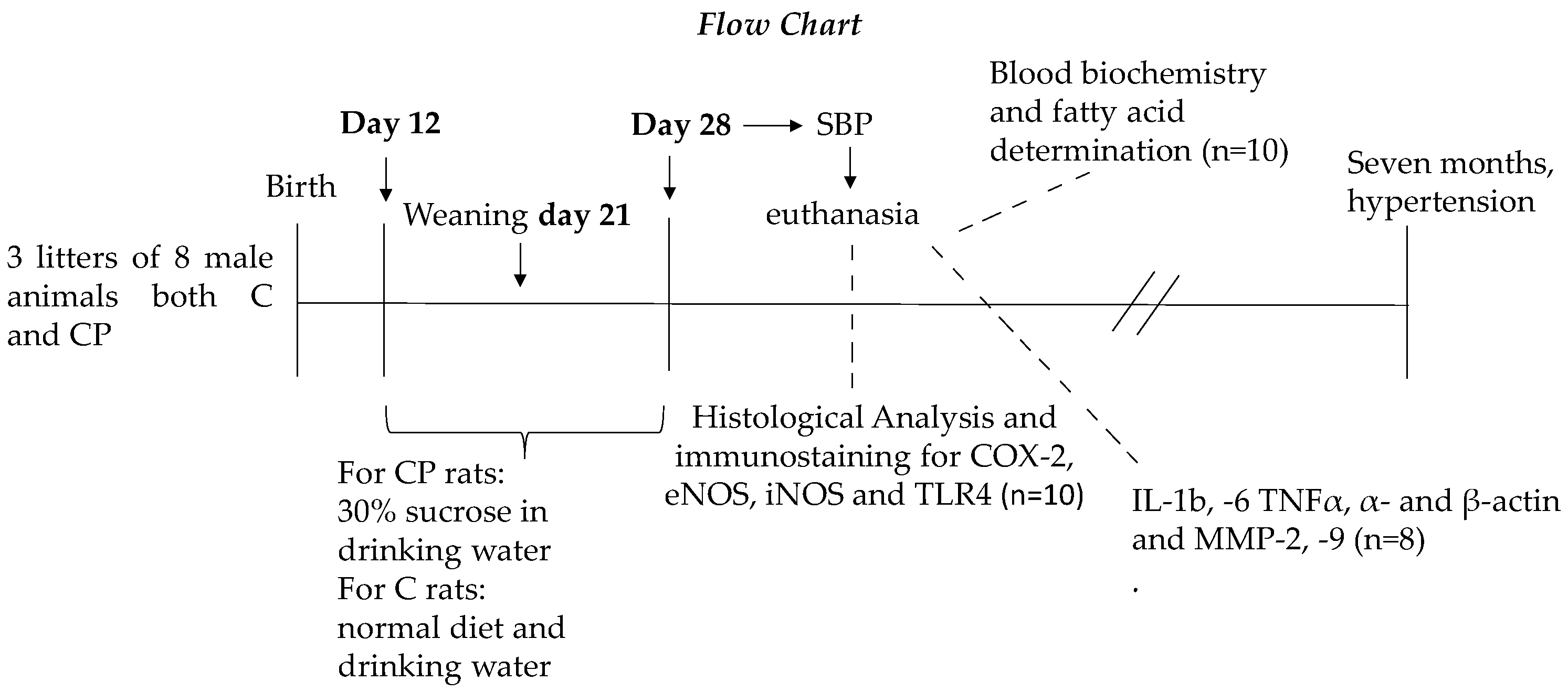
2.1. Animals and Experimental Groups
2.2. Blood Pressure Measurement and Sacrifice
2.3. Biochemical and Physiological Determinations
2.4. Histological Analysis
2.5. Western Blott Analysis (MMP-2, MMP-9, SMA, β-Actin)
2.6. Total Fatty Acid (TFA) Determination
2.7. Determination of Inflammatory Mediators
2.8. Statistical Analysis
3. Results
3.1. Body Variables and Blood Pressure
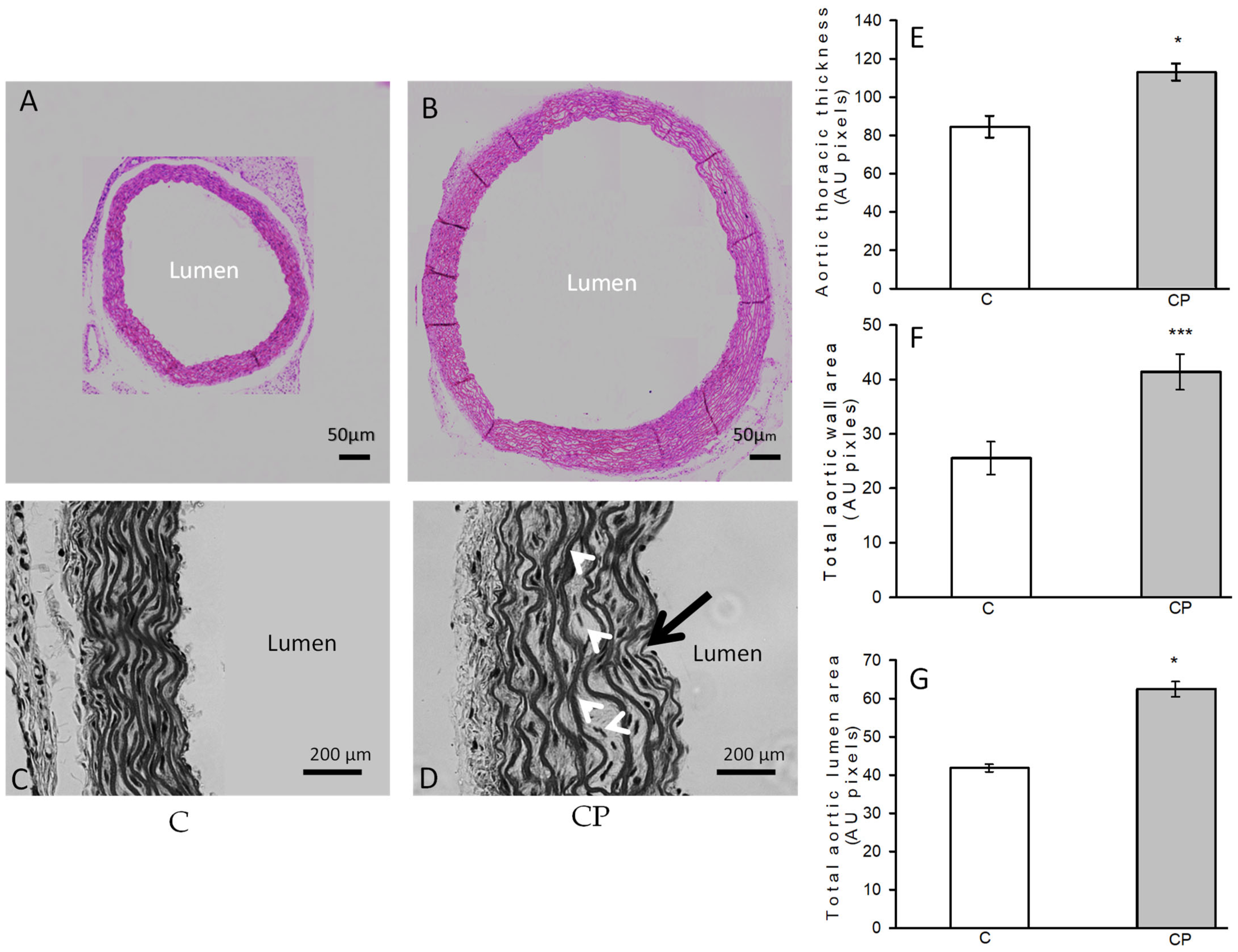
3.2. Changes in the Histology of the Aortic Rings from C and CP Rats
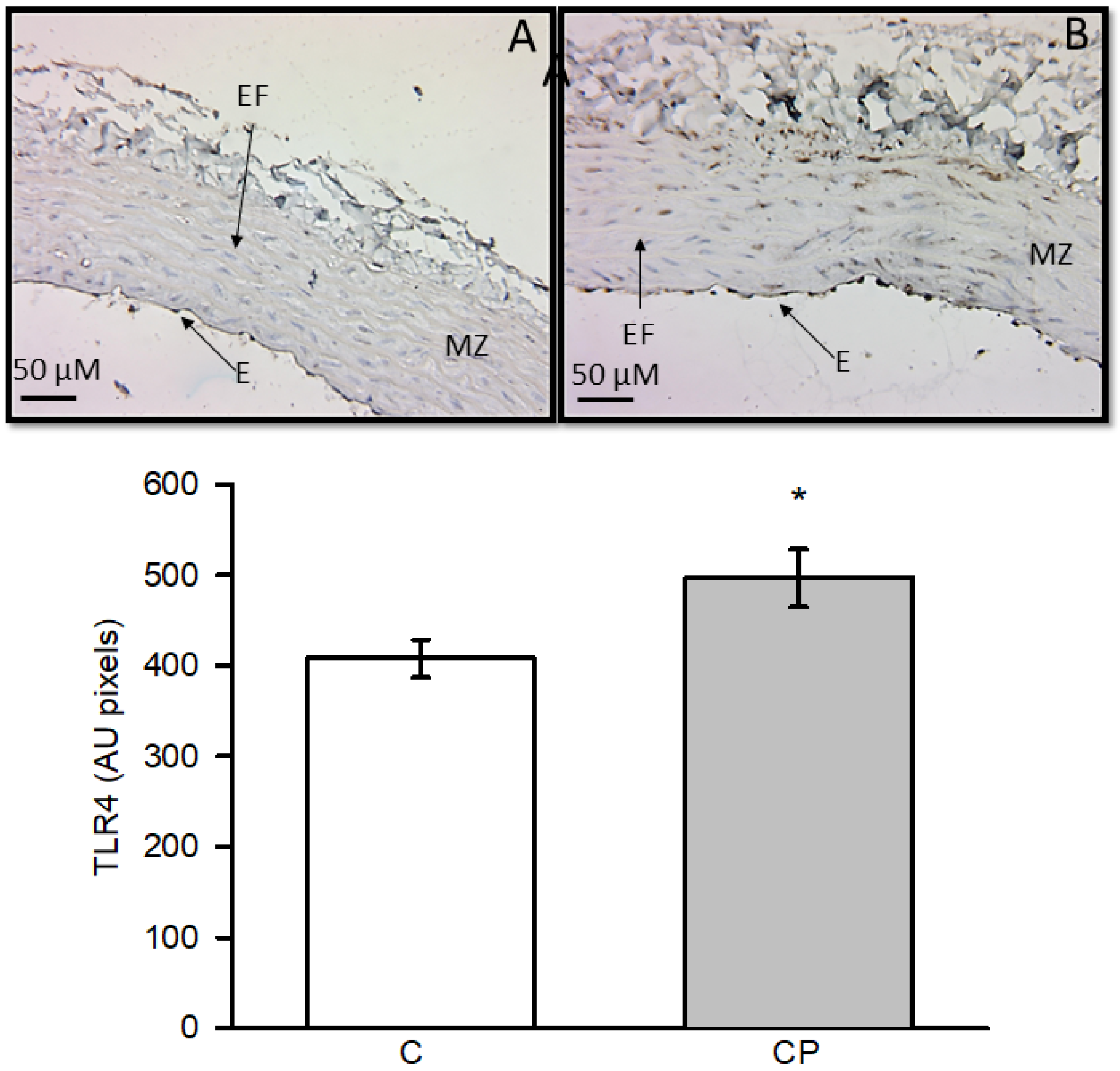
3.3. Expression of COX-2, eNOS, iNOS and TLR4
3.4. Inflammatory Cytokines IL-1β, IL-6, and TNF-α
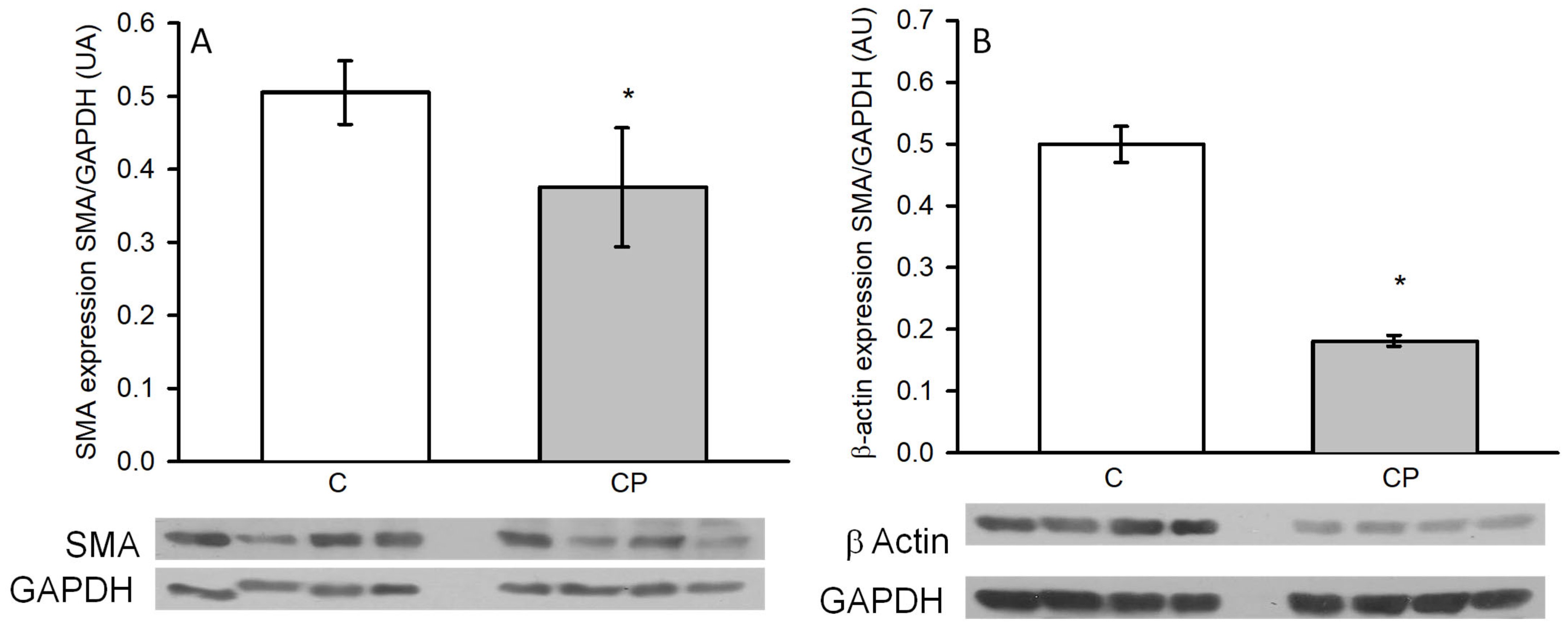
3.5. VSMC Phenotype and Expression of Metalloproteinases and Immune Mediators
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Torres, I.; Hafidi, M.E.; Zamora-González, J.; Infante, O.; Chavira, R.; Baños, G. Modulation of aortic vascular reactivity by sex hormones in a male rat model of metabolic syndrome. Life Sci. 2007, 80, 2170–2180. [Google Scholar] [CrossRef]
- Guarner-Lans, V.; Ramírez-Higuera, A.; Rubio-Ruiz, M.E.; Castrejón-Téllez, V.; Soto, M.E.; Pérez-Torres, I. Early Programming of Adult Systemic Essential Hypertension. Int. J. Mol. Sci. 2020, 21, 1203. [Google Scholar] [CrossRef]
- Berillis, P. The role of collagen in the Aorta’s structure. Open Circ. Vasc. J. 2013, 6, 1–8. [Google Scholar] [CrossRef]
- Manzano-Pech, L.; Guarner-Lans, V.; Soto, M.E.; Díaz-Díaz, E.; Pérez-Torres, I. Alteration of the aortic vascular reactivity associated to excessive consumption of Hibiscus sabdariffa Linnaeus: Preliminary findings. Heliyon 2023, 9, e20020. [Google Scholar] [CrossRef]
- Rzucidlo, E.M.; Martin, K.A.; Powell, R.J. Regulation of vascular smooth muscle cell differentiation. J. Vasc. Surg. 2007, 45, A25–A32. [Google Scholar] [CrossRef]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef]
- Lacolley, P.; Regnault, V.; Segers, P.; Laurent, S. Vascular smooth muscle cells and arterial stiffening: Relevance in development, aging, and disease. Physiol. Rev. 2017, 97, 1555–1617. [Google Scholar] [CrossRef]
- Nian, M.; Lee, P.; Khaper, N.; Liu, P. Inflammatory cytokines and postmyocardial infarction remodeling. Circ. Res. 2004, 94, 1543–1553. [Google Scholar] [CrossRef]
- Schreuder, M.F.; van Wijk, J.A.E.; Delemarre-van de Waal, H.A. Intrauterine growth restriction increases blood pressure and central pulse pressure measured with telemetry in aging rats. J. Hypertens. 2006, 24, 1337–1343. [Google Scholar] [CrossRef]
- Porter, J.P.; King, S.H.; Honeycut, A.D. Prenatal high-salt diet in the Sprague-Dawley rat programs blood pressure and heart rate hyperresponsiveness to stress in adult female offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R334–R342. [Google Scholar] [CrossRef]
- Villegas-Romero, M.; Castrejón-Téllez, V.; Pérez-Torres, I.; Rubio-Ruiz, M.E.; Carreón-Torres, E.; Díaz-Díaz, E.; del Val-le-Mondragón, L.; Guarner-Lans, V. Short-term exposure to high sucrose levels near weaning has a similar long-lasting effect on hypertension as a long-term exposure in rats. Nutrients 2018, 10, 728. [Google Scholar] [CrossRef]
- Aguayo-Mazzucato, C.; Sanchez-Soto, C.; Godinez-Puig, V.; Gutierrez-Ospina, G.; Hiriart, M. Restructuring of pancreatica islets and insulin secretion in a postnatal vritical window. PLoS ONE 2006, 1, e35. [Google Scholar] [CrossRef]
- Rubio-Ruiz, M.E.; Vargas-Gonzalez, A.; Monter-Garrido, M.; Guarner-Lans, V. Aortic vaso-reactivity during a postnatal critical window of the pancreas in rats. Heart Vessel. 2010, 25, 248–253. [Google Scholar] [CrossRef]
- Lee, M.Y.; Griendling, K.K. Redox signaling, vascular function, and hypertension. Antioxid. Redox. Signal. 2008, 10, 1045–1059. [Google Scholar] [CrossRef]
- Nedachi, T.; Kadotani, A.; Ariga, M.; Katagiri, H.; Kanzaki, M. Ambient glucose levels qualify the potency of insulin myogenic actions by regulating SIRT1 and FoxO3a in C2C12 myocytes. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E668–E678. [Google Scholar] [CrossRef]
- Chen, Q.; Jin, M.; Yang, F.; Zhu, J.; Xiao, Q.; Zhang, L. Matrix metalloproteinases: Inflammatory regulators of cell behaviors in vascular formation and remodeling. Mediat. Inflamm. 2013, 2013, 928315. [Google Scholar] [CrossRef]
- Humphrey, J.D. Mechanisms of Vascular Remodeling in Hypertension. Am. J. Hypertens. 2021, 34, 432–441. [Google Scholar] [CrossRef]
- Lamb, F.S.; Choi, H.; Miller, M.R.; Stark, R.J. TNFα and reactive oxygen signaling in vascular smooth muscle cells in hypertension and atherosclerosis. Am. J. Hypertens. 2020, 33, 902–913. [Google Scholar] [CrossRef]
- Orr, A.W.; Hastings, N.E.; Blackman, B.R.; Wamhoff, B.R. Complex regulation and function of the inflammatory smooth muscle cell phenotype in atherosclerosis. J. Vasc. Res. 2010, 47, 168–180. [Google Scholar] [CrossRef]
- Hellerstein, M.K. Carbohydrate-induced hypertriglyceridemia: Modifying factors and implications for cardiovascular risk. Curr. Opin. Lipidol. 2002, 13, 33–40. [Google Scholar] [CrossRef]
- Simon, M.C.; Bilan, S.; Nowotny, B.; Dickhaus, T.; Burkart, V.; Schloot, N.C. Fatty acids modulate cytokine and chemokine secretion of stimulated human whole blood cultures in diabetes. Clin. Exp. Immunol. 2013, 172, 383–393. [Google Scholar] [CrossRef]
- Chen, W.; Zhong, Y.; Feng, N.; Guo, Z.; Wang, S.; Xing, D. New horizons in the roles and associations of COX-2 and novel natural inhibitors in cardiovascular diseases. Mol. Med. 2021, 27, 123. [Google Scholar] [CrossRef]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Pérez-Sala, D.; Lamas, S. Regulation of cyclooxygenase-2 expression by nitric oxide in cells. Antioxid. Redox. Signal. 2001, 3, 231–248. [Google Scholar] [CrossRef]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef]
- Shramko, V.S.; Polonskaya, Y.V.; Kashtanova, E.V.; Stakhneva, E.M.; Ragino, Y.I. The Short overview on the relevance of fatty acids for human cardiovascular disorders. Biomolecules 2020, 10, 1127. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Castrejón-Téllez, V.; Rubio-Ruiz, M.E.; Cano-Martínez, A.; Pérez-Torres, I.; del Valle-Mondragón, L.; Carreón-Torres, E.; Guarner-Lans, V. High sucrose ingestion during a critical period of vessel development promotes the synthetic phenotype of vascular smooth muscle cells and modifies vascular contractility leading to hypertension in adult rats. Int. J. Hypertens. 2022, 2022, 2298329. [Google Scholar] [CrossRef]
- Castrejón-Téllez, V.; Villegas-Romero, M.; Pérez-Torres, I.; Zarco, G.; Rubio-Ruiz, M.E.; Carreón-Torres, E.; Díaz-Díaz, E.; Grimaldo, O.E.; Guarner-Lans, V. Effect of sucrose ingestion at the end of a critical window that increases hypertension susceptibility on peripheral mechanisms regulating blood pressure in rats. role of sirtuins 1 and 3. Nutrients 2019, 11, 309. [Google Scholar] [CrossRef]
- Mu, J.J.; Liu, Z.Q.; Yang, J.; Ren, J.; Liu, W.M.; Xu, X.L.; Xiong, S.E. Association between higher blood pressure level in children and adult blood pressure: 17 years follow-up results. Zhonghua Xin Xue Guan Bing Za Zhi 2008, 36, 229–231. [Google Scholar]
- Osmond, C.; Barker, D.J. Fetal, infant, and childhood growth are predictors of coronary heart disease, diabetes, and hypertension in adult men and women. Environ. Health Perspect. 2000, 108, 545–553. [Google Scholar]
- Berenson, G.S.; Srinivasan, S.R.; Hunter, S.M.; Nicklas, T.A.; Freedman, D.S.; Shear, C.L.; Webber, L.S. Risk factors in early life as predictors of adult heart disease: The Bogalusa Heart Study. Am. J. Med. Sci. 1989, 298, 141–151. [Google Scholar] [CrossRef]
- Grobbee, D.E.; van Hooft, I.M.; de Man, S.A. Determinants of blood pressure in the first decades of life. J. Cardiovasc. Pharmacol. 1990, 16, S71–S74. [Google Scholar] [CrossRef]
- Wang, Y.I.; Schulze, J.; Raymond, N.; Tomita, T.; Tam, K.; Simon, S.I.; Passerini, A.G. Endothelial inflammation correlates with subject triglycerides and waist size after a high-fat meal. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H784–H791. [Google Scholar] [CrossRef]
- Lehman, W.; Morgan, K.G. Structure and dynamics of the actin based smooth muscle contractile and cytoskeletal apparatus. J. Muscle Res. Cell Motil. 2012, 33, 461–469. [Google Scholar] [CrossRef]
- Touyz, R.M. Intracellular mechanisms involved in vascular remodeling of resistance arteries in hypertension: Role of angiotensin II. Exp. Physiol. 2005, 90, 449–555. [Google Scholar] [CrossRef]
- You, J.; Wu, J.; Zhang, Q.; Ye, Y.; Wang, S.; Huang, J.; Liu, H.; Wang, X.; Zhang, W.; Bu, L.; et al. Differential cardiac hypertrophy and signaling pathways in pressure versus volume overload. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H552–H562. [Google Scholar] [CrossRef]
- Yang, J.W.; Cho, K.I.; Kim, J.H.; Kim, S.Y.; Kim, C.S.; You, G.I.; Lee, J.Y.; Choi, S.Y.; Lee, S.W.; Kim, H.S.; et al. Wall shear stress in hypertensive patients is associated with carotid vascular deformation assessed by speckle tracking strain imaging. Clin. Hypertens. 2014, 20, 10. [Google Scholar] [CrossRef]
- Staarmann, B.; Smith, M.; Prestigiacomo, C.J. Shear stress and aneurysms: A review. Neurosurg. Focus 2019, 47, E2. [Google Scholar] [CrossRef]
- Spitzer, J.J.; Bagby, G.J.; Mészáros, K.; Lang, C.H. Alterations in lipid and carbohydrate metabolism in sepsis. JPEN J. Parenter. Enter. Nutr. 1988, 12, 53S–58S. [Google Scholar] [CrossRef]
- Adelizzi, R.A. COX-1 and COX-2 in health and disease. J. Am. Osteopath. Assoc. 1999, 99, S7–S12. [Google Scholar] [CrossRef]
- Qu, D.; Wang, L.; Huo, M.; Song, W.; Lau, C.W.; Xu, J.; Xu, A.; Yao, X.; Chiu, J.J.; Tian, X.Y.; et al. Focal TLR4 activation mediates disturbed flow-induced endothelial inflammation. Cardiovasc. Res. 2020, 116, 226–236. [Google Scholar] [CrossRef]
- Rocha, D.M.; Caldas, A.P.; Oliveira, L.L.; Bressan, J.; Hermsdorff, H.H. Saturated fatty acids trigger TLR4-mediated inflammatory response. Atherosclerosis 2016, 244, 211–215. [Google Scholar] [CrossRef]
- Hyo-Jin, K.; Hyemin, K.; Jeong-Hyung, L.; Cheol, H. Toll-like receptor 4 (TLR4): New insight immune and aging. Immun. Ageing 2023, 20, 67. [Google Scholar]
- Hamsanathan, S.; Gurkar, A.U. Lipids as regulators of cellular senescence. Front. Physiol. 2022, 13, 796850. [Google Scholar] [CrossRef]
- Peng, Z.; Shu, B.; Zhang, Y.; Wang, M. Endothelial response to pathophysiological stress. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e233–e243. [Google Scholar] [CrossRef]
- Brozovich, F.V.; Nicholson, C.J.; Degen, C.V.; Gao, Y.Z.; Aggarwal, M.; Morgan, K.G. Mechanisms of vascular smooth muscle contraction and the basis for pharmacologic treatment of smooth muscle disorders. Pharmacol. Rev. 2016, 68, 476–532. [Google Scholar] [CrossRef]
- Prado, A.F.; Batista, R.I.M.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix metalloproteinases and arterial hypertension: Role of oxidative stress and nitric oxide in vascular functional and structural alterations. Biomolecules 2021, 11, 585. [Google Scholar] [CrossRef]
- Raffetto, J.D.; Khalil, R.A. Matrix metalloproteinases and their inhibitors in vascular remodeling and vascular disease. Biochem. Pharmacol. 2008, 75, 346–359. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Lopes, R.A.; Rios, F.; Touyz, R.M. Vascular Fibrosis in Aging and Hypertension: Molecular Mechanisms and Clinical Implications. Can. J. Cardiol. 2016, 32, 659–668. [Google Scholar] [CrossRef]
- Wang, X.; Khalil, R.A. Matrix metalloproteinases, vascular remodeling, and vascular disease. Adv. Pharmacol. 2018, 81, 241–330. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | CP | |
|---|---|---|
| Body Weight (g) | 128.3 ± 1.5 | 105.2 ± 2.2 * |
| VAT (mg) | 453.8 ± 0.05 | 460.7 ± 0.07 |
| Glucose (mg/dL) | 60.5 ± 3.9 | 63.0 ± 3.3 |
| Insulin (µU/mL) | 0.93 ± 0. 04 | 0.85 ± 0. 07 |
| HOMA-IR | 0.149 ± 0.047 | 0.102 ± 0.06 |
| Triglycerides (mg/dL) | 74.99 ± 3.58 | 120.93 ± 10.38 * |
| Oleic Acid (%) | 17.88 ± 2.69 | 22.93 ± 0.77 * |
| Linoleic Acid (%) | 15.18 ± 0.69 | 12.19 ± 0.40 * |
| AA (%) | 8.55 ± 0.70 | 7.40 ± 0.61 |
| SBP (mmHg) | 93.63 ± 0.63 | 107 ± 0.10 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guarner-Lans, V.; Soria-Castro, E.; Cano-Martínez, A.; Rubio-Ruiz, M.E.; Zarco, G.; Carreón-Torres, E.; Grimaldo, O.; Castrejón-Téllez, V.; Pérez-Torres, I. Rats Exposed to Excess Sucrose During a Critical Period Develop Inflammation and Express a Secretory Phenotype of Vascular Smooth Muscle Cells. Metabolites 2024, 14, 555. https://doi.org/10.3390/metabo14100555
Guarner-Lans V, Soria-Castro E, Cano-Martínez A, Rubio-Ruiz ME, Zarco G, Carreón-Torres E, Grimaldo O, Castrejón-Téllez V, Pérez-Torres I. Rats Exposed to Excess Sucrose During a Critical Period Develop Inflammation and Express a Secretory Phenotype of Vascular Smooth Muscle Cells. Metabolites. 2024; 14(10):555. https://doi.org/10.3390/metabo14100555
Chicago/Turabian StyleGuarner-Lans, Verónica, Elizabeth Soria-Castro, Agustina Cano-Martínez, María Esther Rubio-Ruiz, Gabriela Zarco, Elizabeth Carreón-Torres, Oscar Grimaldo, Vicente Castrejón-Téllez, and Israel Pérez-Torres. 2024. "Rats Exposed to Excess Sucrose During a Critical Period Develop Inflammation and Express a Secretory Phenotype of Vascular Smooth Muscle Cells" Metabolites 14, no. 10: 555. https://doi.org/10.3390/metabo14100555
APA StyleGuarner-Lans, V., Soria-Castro, E., Cano-Martínez, A., Rubio-Ruiz, M. E., Zarco, G., Carreón-Torres, E., Grimaldo, O., Castrejón-Téllez, V., & Pérez-Torres, I. (2024). Rats Exposed to Excess Sucrose During a Critical Period Develop Inflammation and Express a Secretory Phenotype of Vascular Smooth Muscle Cells. Metabolites, 14(10), 555. https://doi.org/10.3390/metabo14100555