
S100B Secretion in Astrocytes, Unlike C6 Glioma Cells, Is Downregulated by Lactate

 ,
,  ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Acute Hippocampal Slices
2.4. Primary Astrocytes Culture
2.5. C6 Glioma Cell Culture
2.6. Treatments
2.7. Cell Viability and Integrity
2.7.1. MTT Reduction Assay
2.7.2. Lactate Dehydrogenase Assay
2.8. Glucose Uptake Assay
2.9. Extracellular Lactate Assay
2.10. Glutamate Uptake
2.11. Glutamine Synthetase Activity
2.12. Glutathione Levels
2.13. ELISA for S100B
2.14. Western Blotting
2.15. Immunofluorescence
2.16. Protein Determination
2.17. Statistical Analysis
3. Results
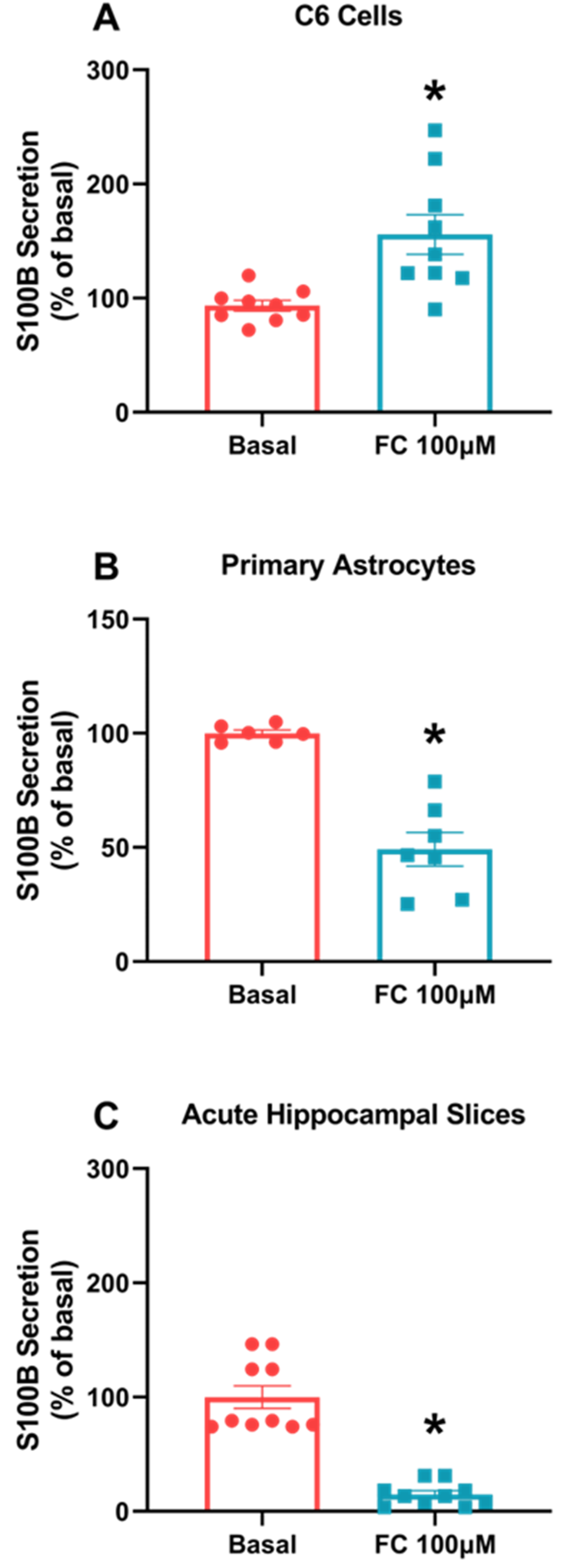
3.1. S100B Secretion Differs in C6 Glioma Cells and Astrocytes
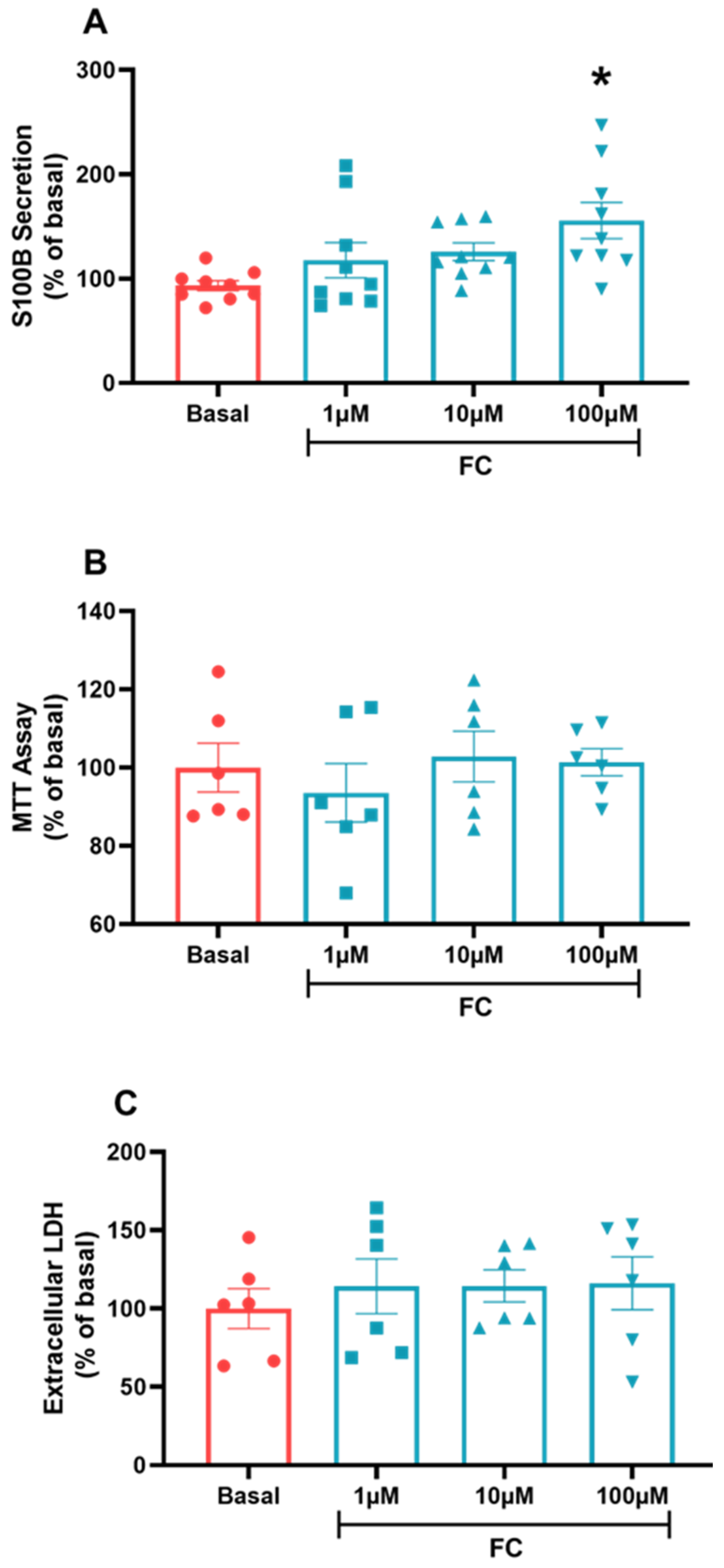
3.2. FC Concentrations Do Not Alter the Viability and Integrity of C6 Glioma Cells
3.3. Some Astrocytic Parameters in C6 Glioma Cells Change in a Similar Manner to Astrocytes in Cultures When Exposed to FC
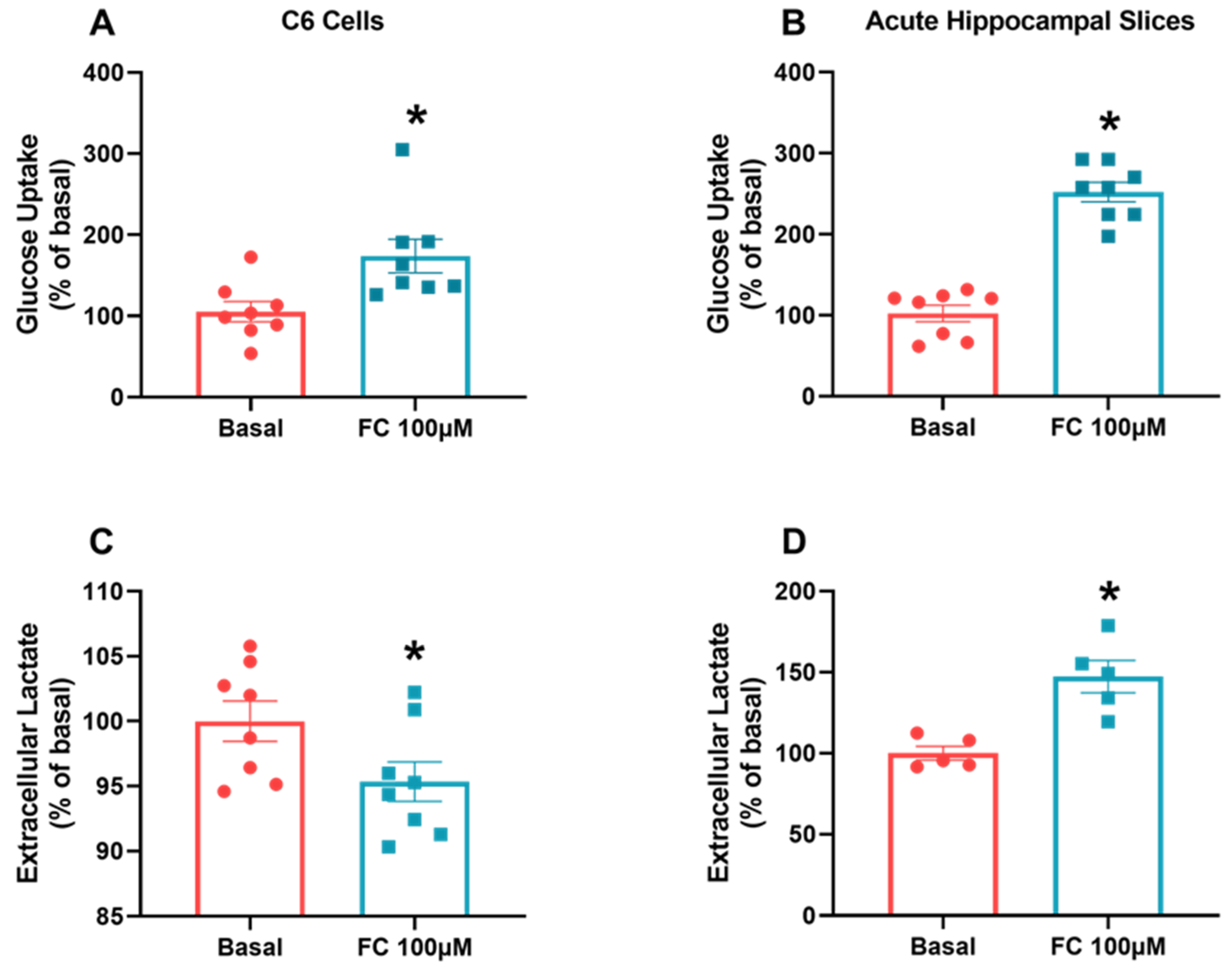
3.4. FC Alters Glucose Uptake but Responds Differently to Lactate Metabolism in C6 Glioma Cells and Hippocampal Slices
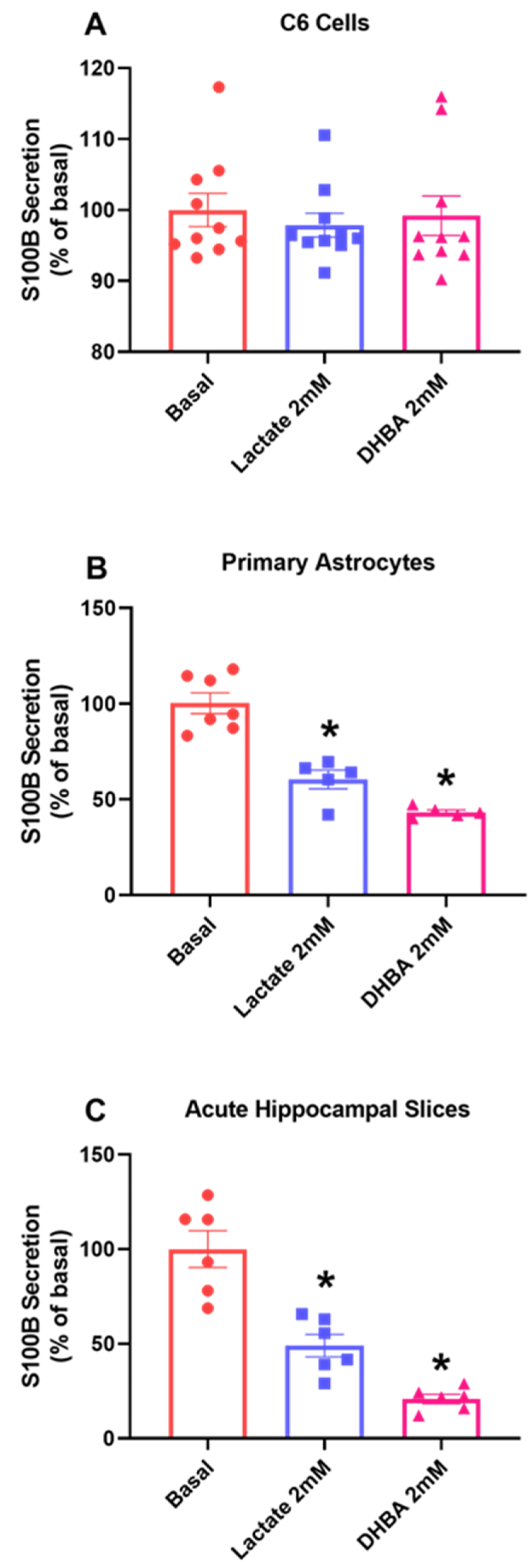
3.5. HCAR1 Could Be Involved in S100B Secretion in Astrocytes but Not in C6 Glioma Cells
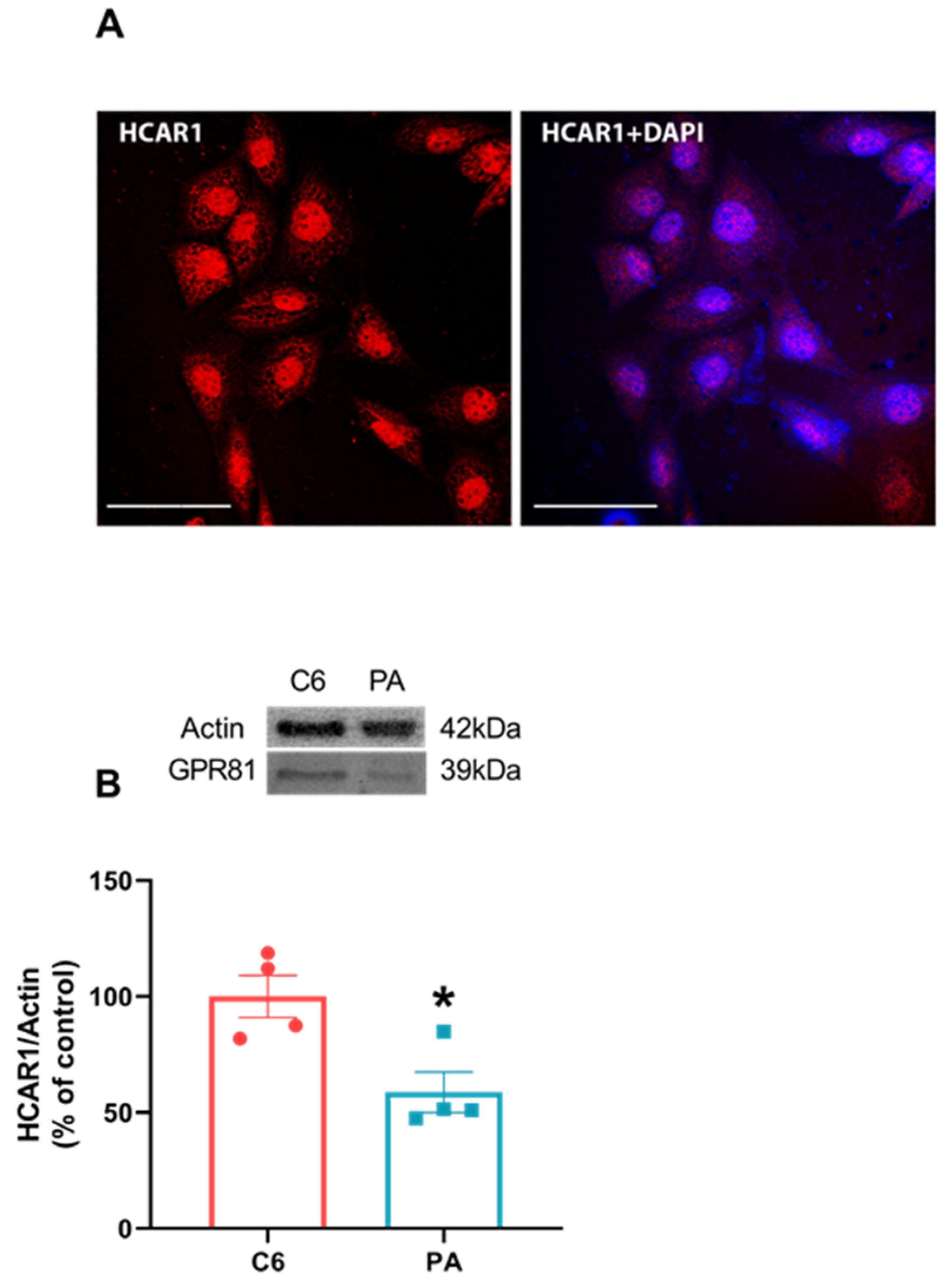
3.6. C6 Glioma Cells Normally Expressed HCAR1
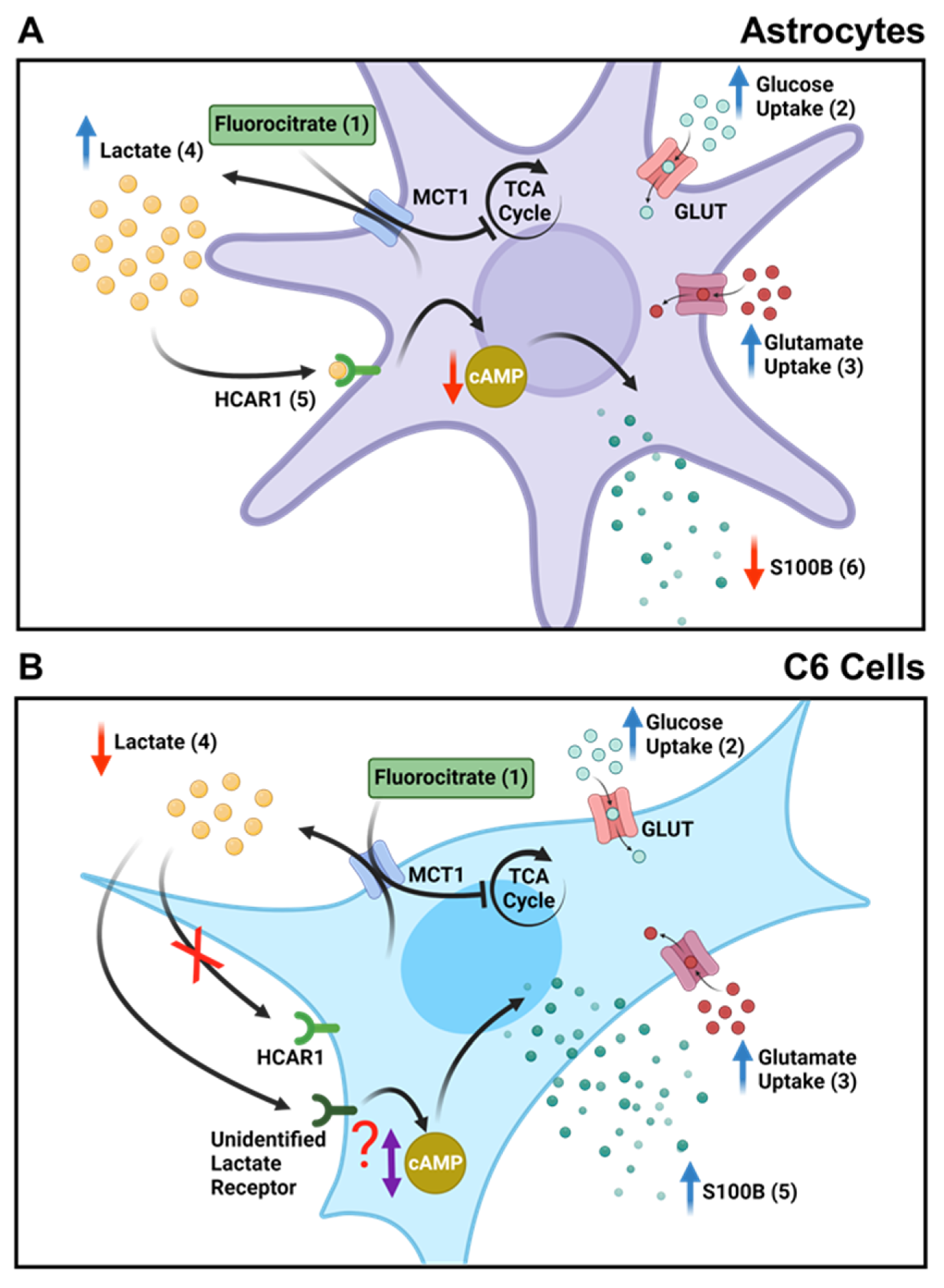
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galland, F.; Seady, M.; Taday, J.; Smaili, S.S.; Gonçalves, C.A.; Leite, M.C. Astrocyte Culture Models: Molecular and Function Characterization of Primary Culture, Immortalized Astrocytes and C6 Glioma Cells. Neurochem. Int. 2019, 131. [Google Scholar] [CrossRef] [PubMed]
- Quincozes-Santos, A.; Bobermin, L.D.; de Assis, A.M.; Gonçalves, C.-A.; Souza, D.O. Fluctuations in Glucose Levels Induce Glial Toxicity with Glutamatergic, Oxidative and Inflammatory Implications. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Sahajpal, N.S.; Singh, H.; Vanita, V.; Roy, P.; Paul, S.; Singh, S.K.; Kaur, I.; Jain, S.K. Pre-Clinical and Cellular Toxicity Evaluation of 7-Methylxanthine: An Investigational Drug for the Treatment of Myopia. Drug Chem. Toxicol. 2021, 44, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Schroeter, M.L.; Schiltz, K.; Bernstein, H.G.; Müller, U.J.; Richter-Landsberg, C.; Müller, W.E.; Walter, M.; Gos, T.; Bogerts, B.; et al. Haloperidol and Clozapine Decrease S100B Release from Glial Cells. Neuroscience 2010, 167, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s Double Life: Intracellular Regulator and Extracellular Signal. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2009, 1793, 1008–1022. [Google Scholar] [CrossRef]
- Frizzo, J.K.; Tramontina, F.; Bortoli, E.; Gottfried, C.; Leal, R.B.; Lengyel, I.; Donato, R.; Dunkley, P.R.; Gonçalves, C.-A. S100B-Mediated Inhibition of the Phosphorylation of GFAP Is Prevented by TRTK-12. Neurochem. Res. 2004, 29, 735–740. [Google Scholar] [CrossRef]
- Balança, B.; Desmurs, L.; Grelier, J.; Perret-Liaudet, A.; Lukaszewicz, A.-C. DAMPs and RAGE Pathophysiology at the Acute Phase of Brain Injury: An Overview. Int. J. Mol. Sci. 2021, 22, 2439. [Google Scholar] [CrossRef]
- Loureiro, S.O.; Heimfarth, L.; de Lima, B.O.; Leite, M.C.; Guerra, M.C.; Gonçalves, C.A.; Pessoa-Pureur, R. Dual Action of Chronic Ethanol Treatment on LPS-Induced Response in C6 Glioma Cells. J. Neuroimmunol. 2012, 249, 8–15. [Google Scholar] [CrossRef]
- Quincozes-Santos, A.; Bobermin, L.D.; Souza, D.G.; Bellaver, B.; Gonçalves, C.-A.; Souza, D.O. Guanosine Protects C6 Astroglial Cells against Azide-Induced Oxidative Damage: A Putative Role of Heme Oxygenase 1. J. Neurochem. 2014, 130, 61–74. [Google Scholar] [CrossRef]
- de Souza, D.F.; Wartchow, K.; Hansen, F.; Lunardi, P.; Guerra, M.C.; Nardin, P.; Gonçalves, C.-A. Interleukin-6-Induced S100B Secretion Is Inhibited by Haloperidol and Risperidone. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 43, 14–22. [Google Scholar] [CrossRef]
- Wartchow, K.M.; Tramontina, A.C.; de Souza, D.F.; Biasibetti, R.; Bobermin, L.D.; Gonçalves, C.-A. Insulin Stimulates S100B Secretion and These Proteins Antagonistically Modulate Brain Glucose Metabolism. Neurochem. Res. 2016, 41, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Nardin, P.; Tramontina, F.; Leite, M.C.; Tramontina, A.C.; Quincozes-Santos, A.; de Almeida, L.M.V.; Battastini, A.M.; Gottfried, C.; Gonçalves, C.-A. S100B Content and Secretion Decrease in Astrocytes Cultured in High-Glucose Medium. Neurochem. Int. 2007, 50, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Bobermin, L.D.; Roppa, R.H.A.; Gonçalves, C.-A.; Quincozes-Santos, A. Ammonia-Induced Glial-Inflammaging. Mol. Neurobiol. 2020, 57, 3552–3567. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; McCandless, D.W. Effect of 6-Aminonicotinamide on Metabolism of Astrocytes and C6-Glioma Cells. Metab. Brain Dis. 1997, 12, 29–45. [Google Scholar] [CrossRef]
- Vizuete, A.F.K.; Hansen, F.; Ré, C.D.; Leal, M.B.; Galland, F.; Leite, M.C.; Gonçalves, C.-A. GABAA Modulation of S100B Secretion in Acute Hippocampal Slices and Astrocyte Cultures. Neurochem. Res. 2019, 44, 301–311. [Google Scholar] [CrossRef]
- Hassel, B.; Paulsen, R.E.; Johnsen, A.; Fonnum, F. Selective Inhibition of Glial Cell Metabolism in Vivo by Fluorocitrate. Brain Res. 1992, 576, 120–124. [Google Scholar] [CrossRef]
- Rae, C.; Fekete, A.D.; Kashem, M.A.; Nasrallah, F.A.; Bröer, S. Metabolism, Compartmentation, Transport and Production of Acetate in the Cortical Brain Tissue Slice. Neurochem. Res. 2012, 37, 2541–2553. [Google Scholar] [CrossRef]
- Tang, F.; Lane, S.; Korsak, A.; Paton, J.F.R.; Gourine, A.V.; Kasparov, S.; Teschemacher, A.G. Lactate-Mediated Glia-Neuronal Signalling in the Mammalian Brain. Nat. Commun. 2014, 5, 3284. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the Brain: From Metabolic End-Product to Signalling Molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Rodrigues, L.; Wartchow, K.M. In Vitro and in Vivo Toxicity of Streptozotocin on Hippocampal S100B and BDNF Proteins. Neurotoxicology 2017, 1603–1611. [Google Scholar] [CrossRef]
- Nardin, P.; Tortorelli, L.; Quincozes-Santos, A.; de Almeida, L.M.V.; Leite, M.C.; Thomazi, A.P.; Gottfried, C.; Wofchuk, S.T.; Donato, R.; Gonçalves, C.-A. S100B Secretion in Acute Brain Slices: Modulation by Extracellular Levels of Ca2+ and K+. Neurochem. Res. 2009, 34, 1603–1611. [Google Scholar] [CrossRef]
- Gottfried, C.; Tramontina, F.; Gonçalves, D.; Gonçalves, C.A.; Moriguchi, E.; Dias, R.D.; Wofchuk, S.T.; Souza, D.O. Glutamate Uptake in Cultured Astrocytes Depends on Age: A Study about the Effect of Guanosine and the Sensitivity to Oxidative Stress Induced by H2O2. Mech. Ageing Dev. 2002, 123, 1333–1340. [Google Scholar] [CrossRef]
- Bobermin, L.D.; Quincozes-Santos, A.; Guerra, M.C.; Leite, M.C.; Souza, D.O.; Gonçalves, C.-A.; Gottfried, C. Resveratrol Prevents Ammonia Toxicity in Astroglial Cells. PLoS ONE 2012, 7, e0052164. [Google Scholar] [CrossRef]
- Tramontina, A.C.; Nardin, P.; Quincozes-Santos, A.; Tortorelli, L.; Wartchow, K.M.; Andreazza, A.C.; Braganhol, E.; de Souza, D.O.G.; Gonçalves, C.-A. High-Glucose and S100B Stimulate Glutamate Uptake in C6 Glioma Cells. Neurochem. Res. 2012, 37, 1399–1408. [Google Scholar] [CrossRef]
- Hassel, B.; Westergaard, N.; Schousboe, A.; Fonnum, F. Metabolic Differences between Primary Cultures of Astrocytes and Neurons from Cerebellum and Cerebral Cortex. Effects of Fluorocitrate. Neurochem. Res. 1995, 20, 413–420. [Google Scholar] [CrossRef]
- Hassel, B.; Sonnewald, U.; Unsgård, G.; Fonnum, F. NMR Spectroscopy of Cultured Astrocytes: Effects of Glutamine and the Gliotoxin Fluorocitrate. J. Neurochem. 1994, 62, 2187–2194. [Google Scholar] [CrossRef]
- D’Adamo, P.; Horvat, A.; Gurgone, A.; Mignogna, M.L.; Bianchi, V.; Masetti, M.; Ripamonti, M.; Taverna, S.; Velebit, J.; Malnar, M.; et al. Inhibiting Glycolysis Rescues Memory Impairment in an Intellectual Disability Gdi1-Null Mouse. Metab.-Clin. Exp. 2021, 116, 154463. [Google Scholar] [CrossRef]
- Liu, C.; Kuei, C.; Zhu, J.; Yu, J.; Zhang, L.; Shih, A.; Mirzadegan, T.; Shelton, J.; Sutton, S.; Connelly, M.A.; et al. 3,5-Dihydroxybenzoic Acid, a Specific Agonist for Hydroxycarboxylic Acid 1, Inhibits Lipolysis in Adipocytes. J. Pharmacol. Exp. Ther. 2012, 341, 794–801. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate Uptake into Astrocytes Stimulates Aerobic Glycolysis: A Mechanism Coupling Neuronal Activity to Glucose Utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Feoli, A.M.; Siqueira, I.; Almeida, L.M.V.; Tramontina, A.C.; Battu, C.; Wofchuk, S.T.; Gottfried, C.; Perry, M.L.; Gonçalves, C.-A. Brain Glutathione Content and Glutamate Uptake Are Reduced in Rats Exposed to Pre- and Postnatal Protein Malnutrition. J. Nutr. 2006, 136, 2357–2361. [Google Scholar] [CrossRef]
- Minet, R.; Villie, F.; Marcollet, M.; Meynial-Denis, D.; Cynober, L. Measurement of Glutamine Synthetase Activity in Rat Muscle by a Colorimetric Assay. Clin. Chim. Acta 1997, 268, 121–132. [Google Scholar] [CrossRef]
- Browne, R.W.; Armstrong, D. Reduced Glutathione and Glutathione Disulfide. Methods Mol. Biol. 1998, 108, 347–352. [Google Scholar] [CrossRef]
- Leite, M.C.; Galland, F.; Brolese, G.; Guerra, M.C.; Bortolotto, J.W.; Freitas, R.; de Almeida, L.M.V.; Gottfried, C.; Gonçalves, C.A. A Simple, Sensitive and Widely Applicable ELISA for S100B: Methodological Features of the Measurement of This Glial Protein. J. Neurosci. Methods 2008, 169, 93–99. [Google Scholar] [CrossRef]
- Peterson, G.L. A Simplification of the Protein Assay Method of Lowry et al. Which Is More Generally Applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- Biswas, J.; Gupta, S.; Verma, D.K.; Singh, S. Streptozotocin Alters Glucose Transport, Connexin Expression and Endoplasmic Reticulum Functions in Neurons and Astrocytes. Neuroscience 2017, 356, 151–166. [Google Scholar] [CrossRef]
- Hayakawa, K.; Nakano, T.; Irie, K.; Higuchi, S.; Fujioka, M.; Orito, K.; Iwasaki, K.; Jin, G.; Lo, E.H.; Mishima, K.; et al. Inhibition of Reactive Astrocytes with Fluorocitrate Retards Neurovascular Remodeling and Recovery after Focal Cerebral Ischemia in Mice. J. Cereb. Blood Flow Metab. 2010, 30, 871–882. [Google Scholar] [CrossRef]
- Largo, C.; Ibarz, J.M.; Herreras, O. Effects of the Gliotoxin Fluorocitrate on Spreading Depression and Glial Membrane Potential in Rat Brain in Situ. J. Neurophysiol. 1997, 78, 295–307. [Google Scholar] [CrossRef]
- Rezagholizadeh, A.; Karimi, S.A.; Hosseinmardi, N.; Janahmadi, M.; Sayyah, M. The Effects of Glial Cells Inhibition on Spatial Reference, Reversal and Working Memory Deficits in a Rat Model of Traumatic Brain Injury (TBI). Int. J. Neurosci. 2022, 132, 226–236. [Google Scholar] [CrossRef]
- Shang, X.-L.; Wang, Q.-B.; Liu, X.-P.; Yao, X.-Q.; Cao, F.-Y.; Wang, Q.; Zhang, J.-Y.; Wang, J.-Z.; Liu, G.-P. Fluorocitrate Induced the Alterations of Memory-Related Proteins and Tau Hyperphosphorylation in SD Rats. Neurosci. Lett. 2015, 584, 230–235. [Google Scholar] [CrossRef]
- Vizuete, A.F.K.; Fróes, F.; Seady, M.; Zanotto, C.; Bobermin, L.D.; Roginski, A.C.; Wajner, M.; Quincozes-Santos, A.; Gonçalves, C.A. Early Effects of LPS-Induced Neuroinflammation on the Rat Hippocampal Glycolytic Pathway. J. Neuroinflammation 2022, 19, 255. [Google Scholar] [CrossRef]
- Wang, T.; Xu, G.; Zhang, X.; Ren, Y.; Yang, T.; Xiao, C.; Zhou, C. Malfunction of Astrocyte and Cholinergic Input Is Involved in Postoperative Impairment of Hippocampal Synaptic Plasticity and Cognitive Function. Neuropharmacology 2022, 217, 109191. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, X.; Dong, J.; Liu, W.C.; Song, M.; Sun, Y.; Shu, H.; Towse, C.L.; Liu, W.; Liu, C.F.; et al. Inhibition of Reactive Astrocytes with Fluorocitrate Ameliorates Learning and Memory Impairment Through Upregulating CRTC1 and Synaptophysin in Ischemic Stroke Rats. Cell. Mol. Neurobiol. 2019, 39, 1151–1163. [Google Scholar] [CrossRef]
- Hosoi, R.; Matsuyama, Y.; Hirose, S.; Koyama, Y.; Matsuda, T.; Gee, A.; Inoue, O. Characterization of (14)C-Acetate Uptake in Cultured Rat Astrocytes. Brain Res. 2009, 1253, 69–73. [Google Scholar] [CrossRef]
- Mac, M.; Nałecz, K.A. Expression of Monocarboxylic Acid Transporters (MCT) in Brain Cells. Implication for Branched Chain Alpha-Ketoacids Transport in Neurons. Neurochem. Int. 2003, 43, 305–309. [Google Scholar] [CrossRef]
- Leite, M.C.; Galland, F.; Guerra, M.C.; Rodrigues, L.; Taday, J.; Monteforte, P.T.; Hirata, H.; Gottfried, C.; Donato, R.; Smaili, S.; et al. Astroglial S100B Secretion Is Mediated by Ca2+ Mobilization from Endoplasmic Reticulum: A Study Using Forskolin and DMSO as Secretagogues. Int. J. Mol. Sci. 2023, 24, 16576. [Google Scholar] [CrossRef]
- Adermark, L.; Lagström, O.; Loftén, A.; Licheri, V.; Havenäng, A.; Loi, E.A.; Stomberg, R.; Söderpalm, B.; Domi, A.; Ericson, M. Astrocytes Modulate Extracellular Neurotransmitter Levels and Excitatory Neurotransmission in Dorsolateral Striatum via Dopamine D2 Receptor Signaling. Neuropsychopharmacology 2022, 47, 1493–1502. [Google Scholar] [CrossRef]
- Morioka, N.; Sugimoto, T.; Tokuhara, M.; Nakamura, Y.; Abe, H.; Hisaoka, K.; Dohi, T.; Nakata, Y. Spinal Astrocytes Contribute to the Circadian Oscillation of Glutamine Synthase, Cyclooxygenase-1 and Clock Genes in the Lumbar Spinal Cord of Mice. Neurochem. Int. 2012, 60, 817–826. [Google Scholar] [CrossRef]
- Bridges, R.J.; Natale, N.R.; Patel, S.A. System Xc- Cystine/Glutamate Antiporter: An Update on Molecular Pharmacology and Roles within the CNS. Br. J. Pharmacol. 2012, 165, 20–34. [Google Scholar] [CrossRef]
- Pallardó, F.V.; Markovic, J.; García, J.L.; Viña, J. Role of Nuclear Glutathione as a Key Regulator of Cell Proliferation. Mol. Aspects Med. 2009, 30, 77–85. [Google Scholar] [CrossRef]
- Grillon, E.; Farion, R.; Fablet, K.; Waard, M.D.; Tse, C.M.; Donowitz, M.; Rémy, C.; Coles, J.A. The Spatial Organization of Proton and Lactate Transport in a Rat Brain Tumor. PLoS ONE 2011, 6, e0017416. [Google Scholar] [CrossRef]
- Gonçalves, C.-A.; Rodrigues, L.; Bobermin, L.D.; Zanotto, C.; Vizuete, A.; Quincozes-Santos, A.; Souza, D.O.; Leite, M.C. Glycolysis-Derived Compounds From Astrocytes That Modulate Synaptic Communication. Front. Neurosci. 2019, 12, 1035. [Google Scholar] [CrossRef] [PubMed]
- Mosienko, V.; Teschemacher, A.G.; Kasparov, S. Is L-Lactate a Novel Signaling Molecule in the Brain? J. Cereb. Blood Flow Metab. 2015, 35, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, D.; Karl, J.; Leite, M.; Rotta, L.; Salbego, C.; Rocha, E.; Wofchuk, S.; Gonçalves, C.-A. High Glutamate Decreases S100B Secretion Stimulated by Serum Deprivation in Astrocytes. Neuroreport 2002, 13, 1533–1535. [Google Scholar] [CrossRef]
- Pinto, S.S.; Gottfried, C.; Mendez, A.; Gonçalves, D.; Karl, J.; Gonçalves, C.A.; Wofchuk, S.; Rodnight, R. Immunocontent and Secretion of S100B in Astrocyte Cultures from Different Brain Regions in Relation to Morphology. FEBS Lett. 2000, 486, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Büyükuysal, R.L. Protein S100B Release from Rat Brain Slices during and after Ischemia: Comparison with Lactate Dehydrogenase Leakage. Neurochem. Int. 2005, 47, 580–588. [Google Scholar] [CrossRef]
- Ma, K.; Ding, X.; Song, Q.; Han, Z.; Yao, H.; Ding, J.; Hu, G. Lactate Enhances Arc/Arg3.1 Expression through Hydroxycarboxylic Acid Receptor 1-β-Arrestin2 Pathway in Astrocytes. Neuropharmacology 2020, 171, 108084. [Google Scholar] [CrossRef]
- Tramontina, F.; Leite, M.C.; Gonçalves, D.; Tramontina, A.C.; Souza, D.F.; Frizzo, J.K.; Nardin, P.; Gottfried, C.; Wofchuk, S.T.; Gonçalves, C.-A. High Glutamate Decreases S100B Secretion by a Mechanism Dependent on the Glutamate Transporter. Neurochem. Res. 2006, 31, 815–820. [Google Scholar] [CrossRef]
- Demircan, C.; Gül, Z.; Büyükuysal, R.L. High Glutamate Attenuates S100B and LDH Outputs from Rat Cortical Slices Enhanced by Either Oxygen-Glucose Deprivation or Menadione. Neurochem. Res. 2014, 39, 1232–1244. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selistre, N.G.; Rodrigues, L.; Federhen, B.C.; Gayger-Dias, V.; Taday, J.; Wartchow, K.M.; Gonçalves, C.-A. S100B Secretion in Astrocytes, Unlike C6 Glioma Cells, Is Downregulated by Lactate. Metabolites 2024, 14, 7. https://doi.org/10.3390/metabo14010007
Selistre NG, Rodrigues L, Federhen BC, Gayger-Dias V, Taday J, Wartchow KM, Gonçalves C-A. S100B Secretion in Astrocytes, Unlike C6 Glioma Cells, Is Downregulated by Lactate. Metabolites. 2024; 14(1):7. https://doi.org/10.3390/metabo14010007
Chicago/Turabian StyleSelistre, Nicholas Guerini, Leticia Rodrigues, Barbara Carolina Federhen, Vitor Gayger-Dias, Jéssica Taday, Krista Mineia Wartchow, and Carlos-Alberto Gonçalves. 2024. "S100B Secretion in Astrocytes, Unlike C6 Glioma Cells, Is Downregulated by Lactate" Metabolites 14, no. 1: 7. https://doi.org/10.3390/metabo14010007
APA StyleSelistre, N. G., Rodrigues, L., Federhen, B. C., Gayger-Dias, V., Taday, J., Wartchow, K. M., & Gonçalves, C.-A. (2024). S100B Secretion in Astrocytes, Unlike C6 Glioma Cells, Is Downregulated by Lactate. Metabolites, 14(1), 7. https://doi.org/10.3390/metabo14010007