

Thyroid Hormone Metabolites Quantified in Pup and Adult Rat Cerebellum, Cortex and Whole-Brain Samples Using an Automated Online SPE-LC-MS/MS Method

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Analytes
2.1.2. Standard Solutions
2.1.3. Brain Samples
2.2. Methods
2.2.1. Sample Preparation
2.2.2. Solid-Phase Extraction (SPE) and Liquid Chromatography (LC)
2.2.3. Instrumentation
2.2.4. Statistical Analysis
3. Results
3.1. Method Validation
3.2. Sample Preparation Is Robust for Brain Matrix
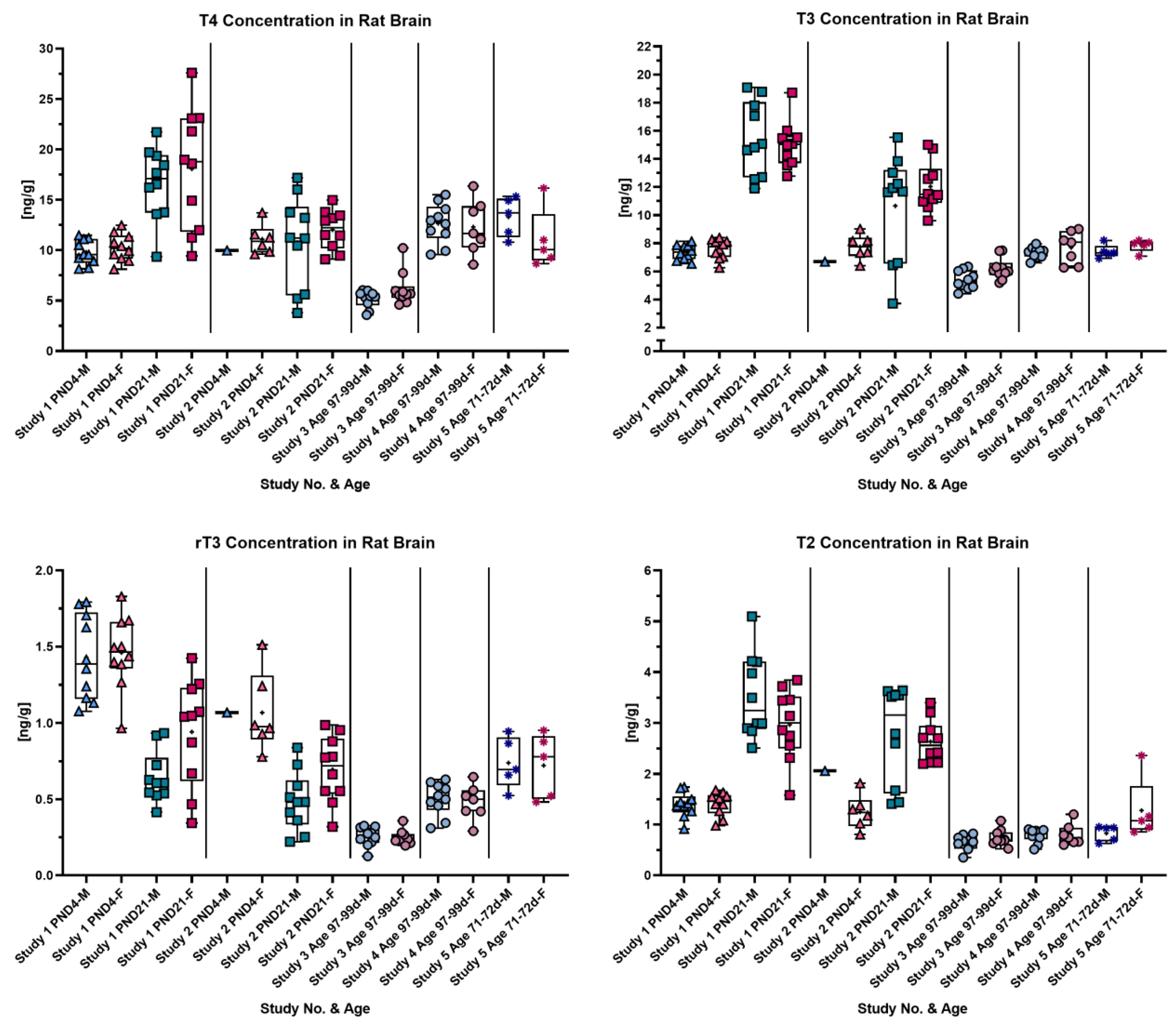
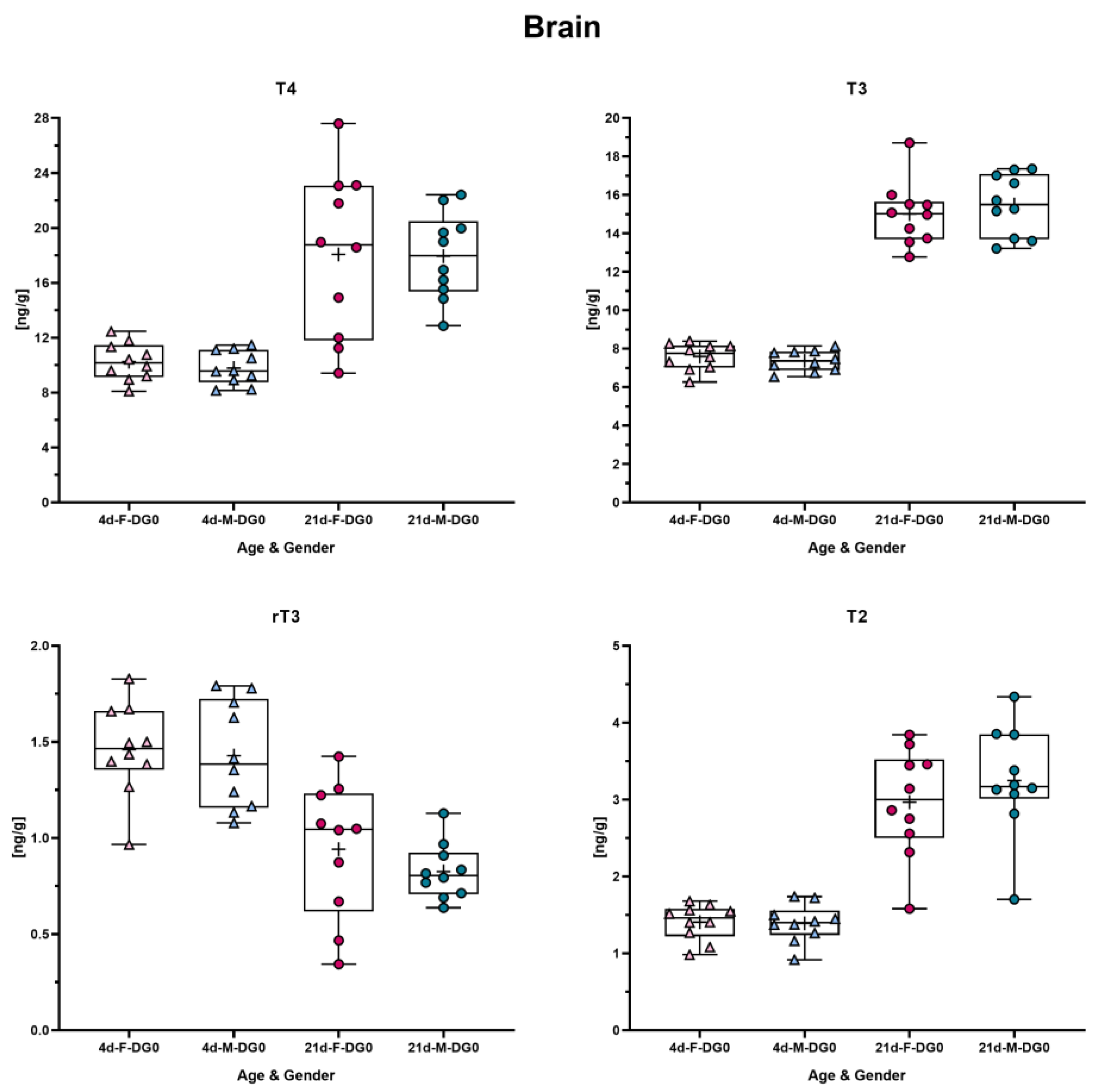
3.3. TH Concentrations in Rat Brain at Different Ages
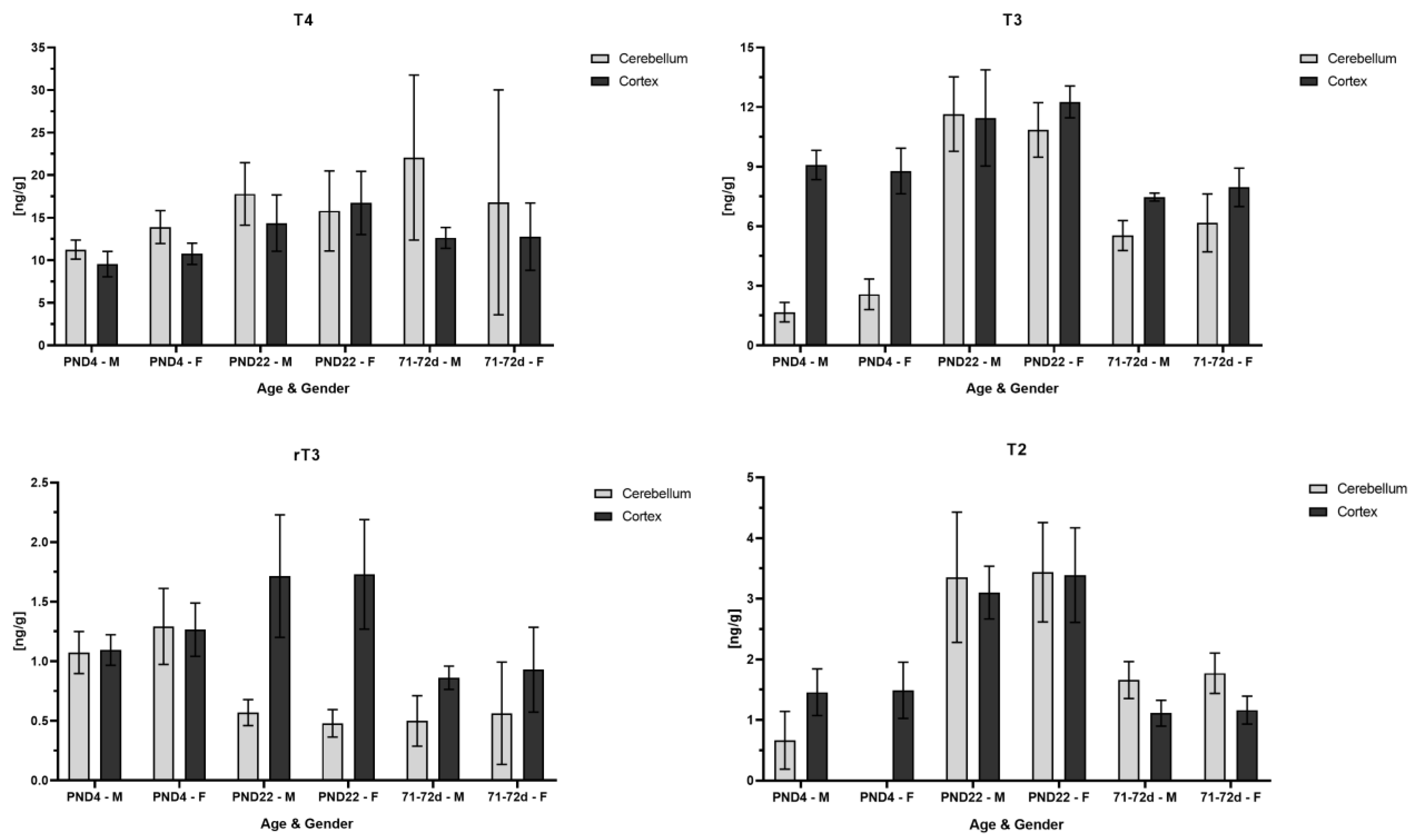
3.4. TH Concentrations in Rat Cerebellum and Cortex at Different Ages
3.5. TH Concentrations in the Brain Do Not Differ between Male and Female Pups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schroeder, A.C.; Privalsky, M.L. Thyroid hormones, T3 and T4, in the brain. Front. Endocrinol. 2014, 5, 40. [Google Scholar] [CrossRef]
- Chatonnet, F.; Picou, F.; Fauquier, T.; Flamant, F. Thyroid Hormone Action in Cerebellum and Cerebral Cortex Development. J. Thyroid. Res. 2011, 145762. [Google Scholar] [CrossRef] [PubMed]
- Crantz, F.R.; Silva, J.E.; Larsen, P.R. An Analysis of the Sources and Quantity of 3,5,3′-Triiodothyronine Specifically Bound to Nuclear Receptors in Rat Cerebral Cortex and Cerebellum. Endocrinology 1982, 110, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Korevaar, T.I.; Muetzel, R.; Medici, M.; Chaker, L.; Jaddoe, V.W.; de Rijke, Y.B.; Steegers, E.A.; Visser, T.J.; White, T.; Tiemeier, H.; et al. Association of maternal thyroid function during early pregnancy with offspring IQ and brain morphology in childhood: A population-based prospective cohort study. Lancet Diabetes Endocrinol. 2016, 4, 35–43. [Google Scholar] [CrossRef]
- Noyes, P.D.; Friedman, K.P.; Browne, P.; Haselman, J.T.; Gilbert, M.E.; Hornung, M.W.; Barone, S., Jr.; Crofton, K.M.; Laws, S.C.; Stoker, T.E.; et al. Evaluating Chemicals for Thyroid Disruption: Opportunities and Challenges with In Vitro Testing and Adverse Outcome Pathway Approaches. Environ. Health Perspect. 2019, 127, 95001. [Google Scholar] [CrossRef]
- Marty, S.; Beekhuijzen, M.; Charlton, A.; Hallmark, N.; Hannas, B.R.; Jacobi, S.; Melching-Kollmuss, S.; Sauer, U.G.; Sheets, L.P.; Strauss, V.; et al. Towards a science-based testing strategy to identify maternal thyroid hormone imbalance and neurodevelopmental effects in the progeny—Part II: How can key events of relevant adverse outcome pathways be addressed in toxicological assessments? Crit. Rev. Toxicol. 2021, 51, 328–358. [Google Scholar] [CrossRef]
- Pinna, G.; Brodel, O.; Visser, T.; Jeitner, A.; Grau, H.; Eravci, M.; Meinhold, H.; Baumgartner, A. Concentrations of seven iodothyronine metabolites in brain regions and the liver of the adult rat. Endocrinology 2002, 143, 1789–1800. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kunisue, T.; Fisher, J.W.; Kannan, K. Determination of Six Thyroid Hormones in the Brain and Thyroid Gland Using Isotope-Dilution Liquid Chromatography/Tandem Mass Spectrometry. Anal. Chem. 2011, 83, 417–424. [Google Scholar] [CrossRef]
- Ford, J.; Riutta, C.; Kosian, P.A.; O’Shaughnessy, K.; Gilbert, M. Reducing uncertainties in quantitative adverse outcome pathways by analysis of thyroid hormone in the neonatal rat brain. Toxicol. Sci. 2023, 193, 192–203. [Google Scholar] [CrossRef]
- Gilbert, M.E.; Hedge, J.M.; Valentín-Blasini, L.; Blount, B.C.; Kannan, K.; Tietge, J.; Zoeller, R.T.; Crofton, K.M.; Jarrett, J.M.; Fisher, J.W. An Animal Model of Marginal Iodine Deficiency During Development: The Thyroid Axis and Neurodevelopmental Outcome. Toxicol. Sci. 2013, 132, 177–195. [Google Scholar] [CrossRef]
- Bastian, T.W.; Prohaska, J.R.; Georgieff, M.K.; Anderson, G.W. Perinatal Iron and Copper Deficiencies Alter Neonatal Rat Circulating and Brain Thyroid Hormone Concentrations. Endocrinology 2010, 151, 4055–4065. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; O’Shaughnessy, K.L.; Thomas, S.E.; Riutta, C.; Wood, C.R.; Smith, A.; Oshiro, W.O.; Ford, R.L.; Hotchkiss, M.G.; Hassan, I.; et al. Thyroid Disruptors: Extrathyroidal Sites of Chemical Action and Neurodevelopmental Outcome—An Examination Using Triclosan and Perfluorohexane Sulfonate. Toxicol. Sci. 2021, 183, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; Hassan, I.; Wood, C.; O’Shaughnessy, K.L.; Spring, S.; Thomas, S.; Ford, J. Gestational Exposure to Perchlorate in the Rat: Thyroid Hormones in Fetal Thyroid Gland, Serum, and Brain. Toxicol. Sci. 2022, 188, 117–130. [Google Scholar] [CrossRef] [PubMed]
- O’Shaughnessy, K.L.; Thomas, S.E.; Spring, S.R.; Ford, J.L.; Ford, R.L.; Gilbert, M.E. A transient window of hypothyroidism alters neural progenitor cells and results in abnormal brain development. Sci. Rep. 2019, 9, 4662. [Google Scholar] [CrossRef]
- Hindrichs, C.; Walk, T.; Melching-Kollmuss, S.; Landsiedel, R.; Kamp, H.; Funk-Weyer, D. A Novel and Fast Online-SPE-LC-MS/MS Method to Quantify Thyroid Hormone Metabolites in Rat Plasma. Chem. Res. Toxicol. 2023, 37, 33–41. [Google Scholar] [CrossRef]
- Sante/2020/12830; Guidance Document on Pesticide Analytical Methods for Risk Assessment and Post-approval Control and Monitoring Purposes. LAUS: Warrington, UK, 2021.
- Gilbert, M.E.; Sui, L. Developmental Exposure to Perchlorate Alters Synaptic Transmission in Hippocampus of the Adult Rat. Environ. Health Perspect. 2008, 116, 752–760. [Google Scholar] [CrossRef]
- O’Shaughnessy, K.L.; McMichael, B.D.; Sasser, A.L.; Bell, K.S.; Riutta, C.; Ford, J.L.; Stoker, T.E.; Grindstaff, R.D.; Pandiri, A.R.; Gilbert, M.E. Thyroid hormone action controls multiple components of cell junctions at the ventricular zone in the newborn rat brain. Front. Endocrinol. 2023, 14, 1090081. [Google Scholar] [CrossRef]
- Bates, J.M.; St Germain, D.L.; Galton, V.A. Expression profiles of the three iodothyronine deiodinases, D1, D2, and D3, in the developing rat. Endocrinology 1999, 140, 844–851. [Google Scholar] [CrossRef]
- Bernal, J. Thyroid Hormones and Brain Development. Vitam. Horm. 2005, 71, 95–122. [Google Scholar]
- Anderson, G.W. Thyroid hormone and cerebellar development. Cerebellum 2008, 7, 60–74. [Google Scholar] [CrossRef]
- Shimokawa, N.; Yousefi, B.; Morioka, S.; Yamaguchi, S.; Ohsawa, A.; Hayashi, H.; Azuma, A.; Mizuno, H.; Kasagi, M.; Masuda, H.; et al. Altered cerebellum development and dopamine distribution in a rat genetic model with congenital hypothyroidism. J. Neuroendocrinol. 2014, 26, 164–175. [Google Scholar] [CrossRef]
- O’Shaughnessy, K.L.; Wood, C.R.; Ford, R.L.; Kosian, P.A.; Hotchkiss, M.G.; Degitz, S.J.; Gilbert, M.E. Thyroid Hormone Disruption in the Fetal and Neonatal Rat: Predictive Hormone Measures and Bioindicators of Hormone Action in the Developing Cortex. Toxicol. Sci. 2018, 166, 163–179. [Google Scholar] [CrossRef]
- Bastian, T.W.; Prohaska, J.R.; Georgieff, M.K.; Anderson, G.W. Fetal and neonatal iron deficiency exacerbates mild thyroid hormone insufficiency effects on male thyroid hormone levels and brain thyroid hormone-responsive gene expression. Endocrinology 2014, 155, 1157–1167. [Google Scholar] [CrossRef]
- Hassan, I.; El-Masri, H.; Kosian, P.A.; Ford, J.; Degitz, S.J.; Gilbert, M.E. Neurodevelopment and Thyroid Hormone Synthesis Inhibition in the Rat: Quantitative Understanding Within the Adverse Outcome Pathway Framework. Toxicol. Sci. 2017, 160, 57–73. [Google Scholar] [CrossRef]
- Escobar, G.M.D.; Pastor, R.; Obregón, M.J.; Rey, F.E.D. Effects of Maternal Hypothyroidism on the Weight and Thyroid Hormone Content of Rat Embryonic Tissues, before and after Onset of Fetal Thyroid Function. Endocrinology 1985, 117, 1890–1900. [Google Scholar] [CrossRef]
- Lees, M.; Sloanestanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Gordon, J.T.; Crutchfield, F.L.; Jennings, A.S.; Dratman, M.B. Preparation of lipid-free tissue extracts for chromatographic determination of thyroid hormones and metabolites. Arch. Biochem. Biophys. 1982, 216, 407–415. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MS/MS | Curtain Gas (CUR) | Collision Gas (CAD) | Ion Spray Voltage (IS) | Temperature (TEM) | Ion Source Gas 1 (GS1) | Ion Source Gas 2 (GS2) |
|---|---|---|---|---|---|---|
| Parameters | 50 | 8 | 5500 | 600 | 50 | 55 |
| RSD (%)/Analyte | T4 | T3 | rT3 | T2 |
|---|---|---|---|---|
| Brain pool (adult) | 7.5 | 14.9 | 20 | 19.1 |
| Age of Rat | Brain Region | TH Concentration | Reference |
|---|---|---|---|
| PND12 males | Half brain | 4 ng/g tissue T3 | [11] |
| PND16 males | Hippocampus and cortex | 7 ng/g and 6.2 ng/g tissue T3 | [24] |
| Fetal GD20 | Whole brain | 0.6 ng/g T4 and 0.7 ng/g T3 | [23] |
| PND14 | Cortex | 3.5 ng/g T4 and 7.9 ng/g T3 | [23] |
| PND0 and 2 | Forebrain | ~0.8 ng/g T4 ~1 ng/g T3 | [9] |
| PND6 and 14 | One hemisphere | ~1.5 ng/g and~3 ng/g T4 ~2 ng/g and~4 ng/g T3 | [9] |
| PND2 | Telencephalon | ~0.5 ng/g T4 ~1.6 ng/g T3 ~0.075 ng/g rT3 | [18] |
| Adult males | Cerebellum | ~1500 fmol/g T3 ~3500 fmol/g T4 | [7] |
| Method | LOQ/MQL | Analytes | Reference |
|---|---|---|---|
| Stable isotope dilution LC-MS/MS | 0.005 ng/g | T4 and T3 | [23,25] |
| RIA | 1.09 ng/mL 6.67 ng/dL | T4 T3 | [24] |
| LC-MS/MS | 0.105 ng/g 0.04 ng/g 0.09 ng/g | T3 rT3 T4 | [9] |
| LC-MS/MS | 0.01 ng/g 0.05 ng/g | T4 and rT3 T3 | [18] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hindrichs, C.; Walk, T.; Landsiedel, R.; Kamp, H.; Schneider, S.; Melching-Kollmuss, S.; Funk-Weyer, D. Thyroid Hormone Metabolites Quantified in Pup and Adult Rat Cerebellum, Cortex and Whole-Brain Samples Using an Automated Online SPE-LC-MS/MS Method. Metabolites 2024, 14, 61. https://doi.org/10.3390/metabo14010061
Hindrichs C, Walk T, Landsiedel R, Kamp H, Schneider S, Melching-Kollmuss S, Funk-Weyer D. Thyroid Hormone Metabolites Quantified in Pup and Adult Rat Cerebellum, Cortex and Whole-Brain Samples Using an Automated Online SPE-LC-MS/MS Method. Metabolites. 2024; 14(1):61. https://doi.org/10.3390/metabo14010061
Chicago/Turabian StyleHindrichs, Christiane, Tilmann Walk, Robert Landsiedel, Hennicke Kamp, Steffen Schneider, Stephanie Melching-Kollmuss, and Dorothee Funk-Weyer. 2024. "Thyroid Hormone Metabolites Quantified in Pup and Adult Rat Cerebellum, Cortex and Whole-Brain Samples Using an Automated Online SPE-LC-MS/MS Method" Metabolites 14, no. 1: 61. https://doi.org/10.3390/metabo14010061
APA StyleHindrichs, C., Walk, T., Landsiedel, R., Kamp, H., Schneider, S., Melching-Kollmuss, S., & Funk-Weyer, D. (2024). Thyroid Hormone Metabolites Quantified in Pup and Adult Rat Cerebellum, Cortex and Whole-Brain Samples Using an Automated Online SPE-LC-MS/MS Method. Metabolites, 14(1), 61. https://doi.org/10.3390/metabo14010061