Abstract
Currently, the antioxidant properties of amino acids and their role in the physicochemical processes accompanying oxidative stress in cancer remain unclear. Cancer cells are known to extensively uptake amino acids, which are used as an energy source, antioxidant precursors that reduce oxidative stress in cancer, and as regulators of inhibiting or inducing tumor cell-associated gene expression. This review examines nine amino acids (Cys, His, Phe, Met, Trp, Tyr, Pro, Arg, Lys), which play a key role in the non-enzymatic oxidative process in various cancers. Conventionally, these amino acids can be divided into two groups, in one of which the activity increases (Cys, Phe, Met, Pro, Arg, Lys) in cancer, and in the other, it decreases (His, Trp, Tyr). The review examines changes in the metabolism of nine amino acids in eleven types of oncology. We have identified the main nonspecific mechanisms of changes in the metabolic activity of amino acids, and described direct and indirect effects on the redox homeostasis of cells. In the future, this will help to understand better the nature of life of a cancer cell and identify therapeutic targets more effectively.
1. Introduction
Recent research in the field of cancer cell metabolism is significantly changing our understanding of metabolic processes such as the redox mechanism, carbon metabolism, epigenetic regulation, and immune response associated with tumorigenesis and metastasis [1,2]. This includes reconsidering the role of amino acids and changes in their composition during oncological processes. It is known that cancer cells intensively absorb amino acids and use them as a source of energy, precursors of antioxidants that reduce the level of oxidative stress in cancer, and also as regulators of inhibition or induction of gene expression associated with the regulation of tumor cell activity [3].
The pathophysiological process begins with a change in the oxidative capacity of proteins, through modification of the structure and activity of protein binding sites with other molecules, which are elements in the cascade reaction of antioxidant activity [4,5,6]. The choice of the oxidative process path and its activity is determined the features of the reactive type like its the activity and specificity. It is deteremined by the cell type, state of cell differentiation, state of the extracellular environment, level of antioxidants and antioxidant enzymes. The activity of the redox process will depend on the specific type of modified protein, and this, in turn, depends on the amino acid composition of the protein. In addition to the above, there is the concept of protein network formation, which explains the coordinated interaction between proteins and the activation of the redox pathway [7,8].
A number of studies on model systems have established that amino acids exhibit the ability to reduce damaging oxidative effects of various natures at different levels of biological organization [9]. These effects probably happen because of the physicochemical properties of individual amino acids, which are connected to their ability to react with reactive oxygen species. For example, the ability of citrulline and Arg to eliminate the superoxide anion radical has been shown, and leads to normalization of the functioning of the heart muscle when exposed to oxidizing factors [10,11]. It has been established that proline is an effective scavenger of singlet oxygen and prevents cell death under oxidative stress [12]. The ability of His to intercept peroxyl radicals and prevent the carboxylation of proteins and the formation of protein cross-links has been revealed [13]. It was discovered that a number of amino acids prevent the formation of 8-oxoguanine in DNA by protecting guanine from one-electron oxidation to the guanine radical cation [14]. It is still unclear to what extent the change in amino acid composition is tissue-specific, as well as how certain types of cancer are metabolically dependent on a specific amino acid composition [15].
It is known that free amino acids can influence the production of hydrogen peroxide and hydroxyl radicals in aqueous solutions when exposed to X-ray radiation [9]. According to the literature, the α-carbon atom is an essential part of all amino acids; it is capable of reacting with a hydroxyl radical, which results in the abstraction of a hydrogen atom at the α-carbon atom to form an amino acid radical [16]. In addition, side amino acid residues are also capable of reacting with the hydroxyl radical; therefore, individual amino acids are observed to have different effects on their formation. In this regard, six amino acids that react most actively with hydroxyl radicals and significantly reduce their production can be identified: Cys, His, Phe, Met, Trp, Tyr [9]. The other most common sites of amino acid oxidation in proteins include Pro, Arg, and Lys [16]. It can be assumed that the oxidation of these amino acids occurs partially along a different path, which is not associated with the action of hydroxyl radicals, for example, through superoxide anion radicals and peroxides. Indeed, Arg and Lys are characterized by their interaction with superoxide anion radicals, and Pro very efficiently forms peroxides [17].
We have specifically chosen these amino acids for a more detailed consideration of their role in the non-enzymatic oxidative process that occurs in various types of cancer because of their features. We have identified nine basic amino acids that play a key role in redox processes. These amino acids include Cys, His, Phe, Met, Trp, Tyr, Pro, Arg, and Lys.
2. Basic Aspects of Amino Acid Metabolism during Redox Stress in Oncology
In oncological diseases, an imbalance of redox homeostasis occurs [1]. At local levels, cancer cells use amino acids as elements of antioxidant protection against the body’s aggressive response to their growth and development. At the systemic level, oxidative stress increases due to the body’s reaction and the activation of free radicals, which are reactive oxygen species that aim to destroy cancer cells. In addition, the cancer cell itself reprograms the metabolism of amino acids, which leads to the synthesis of by-products, thereby increasing oxidative stress. This allows tumor cells to freely invade adjacent tissues and organs and cause unhindered metastasis to distant organs [1,2,3].
Conventionally, amino acids are commonly thought of as the building blocks of protein synthesis [5]. However, according to the latest data, their role is much broader. They take part in the regulation of protein metabolism and signal transmission, maintain redox homeostasis, act as an energy source along with glucose, take part in the transport of nitrogen and carbon throughout the organs and tissues of the body, and act as neurotransmitters [2,3,4,5,6,7]. The uniqueness of amino acids is due to the presence of a free carboxyl group and a free amino group on the a-carbon atom [3,4,5,6]. Differences in the properties of amino acids are due to differences in side chains. The biosynthesis of amino acids includes several metabolic pathways through which they are synthesized from other precursors [5,6,7,8,9,10,11]. A significant difference between the biosynthesis of amino acids and the biosynthesis of lipids and carbohydrates is the active use of nitrogen [5].
A specific relationship between amino acid biosynthesis and cancer cell metabolism has not been identified yet. This can be explained by the fact that, firstly, not enough data have been collected to allow us to draw conclusions about the specific relationships between amino acid content and cancer metabolism. Secondly, amino acids are molecules that are basic elements in ensuring the normal functioning of the cell. It must be remembered that any pathology leading to an imbalance of homeostasis will affect the uneven metabolism of amino acids. The main issue here is that the aggressiveness of the pathology directly affects the basic elements of cell life, including the amino acid composition. An important reason for studying amino acid metabolism is that there are still not enough data regarding (1) how a cancer cell maintains its energy balance other than the active consumption of glucose; (2) through which mechanisms metastasis occurs, and how they can be regulated; (3) due to which specific abilities the cancer cell bypasses the attack from the immune system; and (4) how exactly the imbalance in the redox system occurs. A deep understanding of what type of cancer causes reprogramming of the amino acid metabolic pathway and what positive effect this brings to the cancer cell will primarily help to identify effective therapeutic targets, and to better understand the nature of the cancer cell.
Our review identified several possible amino acid derivatives that arise during metabolic reprogramming. When the Phe content is imbalanced, phenyllactic and phenylacetic acids accumulate [18]. With an increase in Met content, homocysteine accumulates [19]. Disorders of Trp metabolism lead to the accumulation of kynurenine, kynurenic acid, 3-hydroxykynurenine, anthranilic acid, 3-hydroxyanthranilic acid, picolinic acid and quinolinic acid [20]. An increase in Arg either directly leads to the activation of genes such as epidermal growth factor receptor (EGFR) and tuberous sclerosis complex 2 (TSC2) [21], or excessive accumulation of nitric oxide causes the expression of the vascular endothelial growth factor (VEGF) gene [1]. A decrease in the activity of hydrogen peroxide [1] and a decrease in the content of NAD(P)+/NAD(P)H also allow for the growth of cancer cells [2]. In addition, the formation of interesting compounds such as nitrogen–oxygen–sulfur bridges in proteins, coinciding with an increase in Lys content, also favors the growth and activity of cancer cells [4].
In oncology, the state of redox stress increases, and amino acid metabolism plays a significant role in this. Some of them (Pro, His) directly affect the production of reactive oxygen species, allowing for the action of hydrogen peroxide; others (Met, Cys) increase the production of glutathione, as one of the strong antioxidants that minimizes the amount of reactive oxygen species and neutralizes free radicals in the body. Arginine increases the production of nitric oxide. High Trp content leads to an increase in de novo NAD+ synthesis, which enhances antioxidant activity. It is important to note that the role of amino acids in antioxidant defense is twofold. In the normal state of the body, all of the listed antioxidant agents work as protectors, but in oncology, some of the listed molecules can have a damaging effect. At what stage and under what exact circumstances this change occurs remain to be investigated.
3. The Role of Individual Amino Acids in Redox Processes
3.1. Arginine
Particular attention is drawn to Arg and its ambiguous role in the tumor cell. On the one hand, an increase in Arg content leads to an increase in the formation of nitric oxide (NO). Nitric oxide is known to be a strong antioxidant [22]. In this case, Arg can be indirectly attributed to antioxidant potential. By acting as an antioxidant, Arg can scavenge O2− and thereby prevent eNOS-mediated production of O2− in the unbound state. In addition to its indirect antioxidant effects [23,24], NO prevents oxidative stress in tissues by interrupting the lipid peroxidation chain reaction through the formation of non-radical new nitrogen-containing lipid adducts [25] and inhibiting the catalytic activity of CYP2E1 [26,27]. Secondly, NO can trigger the expression of antioxidant enzymes and new genes for resistance to nitrosative stress [28]. Finally, NO enhances the antioxidant activity of glutathione (GSH) [29] by producing S-nitrosoglutathione, which is approximately 100 times more potent than GSH [30]. GSH is not only a major antioxidant, but also activates glutathione peroxidase activity, which protects against oxidation and nitration reactions [31].
There are the additional ways in which Arg can modulate cellular signaling pathways [23]. As already mentioned, Arg is a precursor of NO through the action of NO synthase (NOS) [32]. The dual functions of NO as a signaling messenger in cancer have been noted [33]. NO has been considered as an antitumor reagent due to its antioxidant and radical scavenging properties [34]. However, new data show a diverse role of NO in tumorigenesis, including angiogenesis, metastasis, anti-apoptosis, and anti-host immune response [35]. Increased levels of NO and NOS expression are observed in cancer patients, which strongly correlates with VEGF expression, angiogenesis and metastasis [1,2] (Figure 1). Induction of NO and NOS leads to inactivation of tumor suppressors such as p53 and pRb [36,37]. Induction of NOS and cyclooxygenase (COX-2) has also been shown to increase NO and prostaglandin levels, leading to angiogenesis in cancer [38,39,40]. According to scientific research in recent years, a significant concentration of Arg in a cancer cell can also lead to the activation of another metabolic pathway, namely the activation of mTOR1, which contributes to aggressive and invasive tumor growth. This occurs through S-nitrosylation of key molecules involved in cancer induction, such as EGFR and TSC2, which affects the mammalian target of rapamycin (mTOR) pathway [21,41].
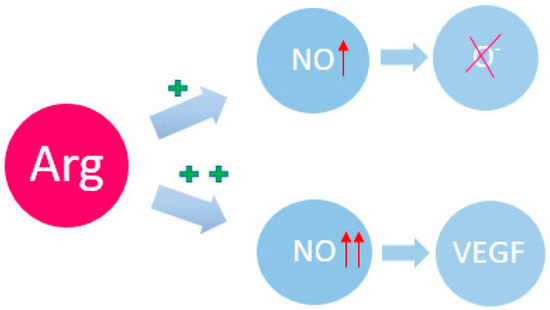
Figure 1.
Arg and its participation in redox regulation. NO—nitric oxide; VEGF—vascular endothelial growth factor. The plus sign indicates the induction force; the arrow indicates the magnification force.
Another interesting discovery was made about the proportional change in the Arg level and its decrease and increase in the content of amino acids such as Trp, Cys and histamine, which led to the suppression of oxidative stress. This process may be due to two reasons. The first includes the ability of a tumor cell to inactivate the suppressor protein p53, which contains the maximum amount of Arg. Another is that His, Trp and histamine reduce the level of oxidative stress in the tumor microenvironment, thereby ensuring high survival of the cancer cell [42,43]. Apparently, high concentrations of Arg have contradictory effects on the body, from antioxidant activity to tumorigenesis-stimulating activity, which is associated with the internal ratio of the concentration of Arg itself to other biological products. This is what may determine further determination of the metabolic pathway of this amino acid. This assumption seems reasonable, since the launch of the cascade reaction of the metabolic process is controlled quantitatively, i.e., in the concentration of biological molecules, and qualitatively, i.e., through activation of specific molecules. In the future, it will be necessary to study the dependence of Arg concentration on the type of pathology and other products, which will ultimately determine the metabolic pathway of Arg, and, consequently, its biological effect.
3.2. Tryptophan
In recent decades, the role of Trp in pathological processes, ranging from chronic systemic inflammation to cancer, has been actively studied [44,45,46]. Thus, an inverse relationship between Trp content and the risk of developing colorectal cancer has been shown, i.e., in patients with a high Trp content, the risk of developing this pathology is significantly lower. This is due to a number of reasons. Trp is actively metabolized by the microbiota of the large intestine, resulting in the formation of end products in the form of indole and indole-3-propionylic acid. These metabolites cover the surface of enterocytes, exerting a protective effect against direct mechanical irritation and the chemical and biological effects of nutrients and microorganisms, and reduce the permeability and leakiness of enterocytes, which may cause the development of colorectal cancer [47,48,49,50,51]. Additionally, Trp is the third source of de novo synthesis of NAD+ (NAD+ is also synthesized from nicotinic acid and nicotinamide mononucleotide), which, in turn, changes via the pentose phosphate pathway [52,53] into NADPH-, which is actively involved in antioxidant protection and regulation intercellular signaling through the NADH oxidase (NOX) system (Figure 2) [54,55].
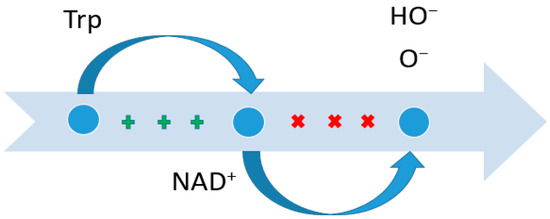
Figure 2.
Suppression of the content of certain radicals due to Trp. NAD—nicotinamide adenine dinucleotide. The plus sign indicates the induction force; crosses indicate suppression.
The indirect role of Trp in the redox process was mentioned in a study of Trp metabolism through the kynurenine pathway and its role in the immunobiology of breast cancer [56]. Trp has three possible metabolic pathways: indole, serotonin, and kynurenine. Particular attention was paid to the kynurenine pathway, especially its metabolites: kynurenine, kynurenic acid, 3-hydroxykynurenine, anthranilic acid, 3-hydroxyanthranilic acid, picolinic acid, and quinolinic acid [57]. Under certain intracellular conditions or in a certain microenvironment, the listed products of this pathway are capable of generating reactive oxygen species such as hydrogen peroxide and hydroxyl radicals. This is due to the emergence and progressive growth of the tumor, leading to lipid peroxidation [57], DNA damage or mutation [20,58], and cell proliferation [59,60,61,62,63,64,65,66,67,68,69].
3.3. Histidine
Interesting data on the redox properties of His were obtained by a group of scientists from the University of Nebraska Medical Center, USA. In their work, they demonstrated a connection between pancreatic cancer and His levels. It was found that in pancreatic cancer, the level of His decreases, which is a protective mechanism of the cancer cell, since a significant amount of His increases oxidative stress in the cell due to increased production of hydrogen peroxide [70]. Increased expression of the HAL gene was observed, indicating active His metabolism. In their work, they also referred to previous studies in which changes in the metabolism of cancer cells were observed, but in non-small cell lung cancer. In this case, the patients were initially resistant to antitumor treatment, but after adjusting the chemotherapy with the addition of hydrogen peroxide, His increased its cytotoxic effect (Figure 3) [71].
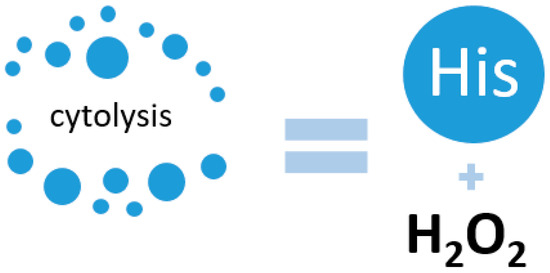
Figure 3.
Activation of cell cytolysis due to the interaction of His and hydrogen peroxide.
Another group of scientists studied the effect of His on DNA damage and assessed the cytotoxicity caused by hydrogen peroxide in cultured mammalian cells. They showed that His markedly enhanced the growth and DNA synthesis-inhibiting effects of hydrogen peroxide in cultured Chinese hamster ovary cells. DNA single-strand breaks were also higher in the presence of the amino acid, and in addition, these breaks had a slower rate of repair compared to breaks caused by the oxidant alone. In the presence of His, hydrogen peroxide also caused a double-stranded DNA break, a damage that was undetectable in cells treated with even extremely high concentrations of a single oxidant. The data presented here suggest that His-mediated enhancement of the cytotoxic response of cultured Chinese hamster ovary cells to hydrogen peroxide may be at least partially dependent on the formation of DNA double-strand breaks [71].
3.4. Tyrosine
Tyr, like any other amino acid, is a building material for proteins on the one hand, and acts as an alternative source of energy for cell life on the other hand. The main site of Tyr metabolism is the liver. In it, the formation of precursors for gluconeogenesis and ketogenesis occurs, as well as the decomposition of amino acids into secondary metabolites with the participation of five key enzymes. Disturbances in Tyr metabolism lead to the occurrence of pathologies such as Huntington’s disease [72] or esophageal cancer [73,74]. Nguyen et al. conducted a comprehensive cross-platform data study to search for potential therapeutic and preventive biomarkers for hepatocellular carcinoma (HCC) [75]. They found a pathologically reduced level of expression of the genes tyrosine aminotransferase (TAT), heme-dependent peroxin (HDP), homogentisate-1,2-dioxygenase (HTG), Glutathione-S-transferase zeta 1 (GSTZ1), and fumarylacetoacetase (FAH), and a consequent decrease in the translation of these enzymes [76]. A positive correlation has been established between the levels of enzymes such as TAT, HTG, and GSTZ1 and the survival of patients with an established diagnosis of HCC. An explanation for the positive prognosis associated with TAT is given in a study by Fu et al. [77]. The proapoptotic effect of tyrosine aminotransferase was experimentally demonstrated by comparing two cell cultures, one of which was transfected with TAT; the other was transfected with an empty vector. The apoptosis index was assessed before and after treatment with straurosporine. The apoptosis index was higher in cell culture with the TAT vector. The proapoptotic effect was due to changes in mitochondrial membrane potential. The mechanism of apoptosis was triggered as follows: the mitochondrial permeability of the cell increased, which led to an increase in the release of Cytochrome-C into the cytoplasm. Cytochrome-C changed the conformation of Apaf-1, initiating apoptosis through activation of caspase-9, which activated caspase-3 and Poly ADP Ribose Polymerase (PARP) [78]. High expression of GSTZ1 enhanced oxidative phosphorylation and respiratory electron transport, which had a negative effect on the development of HCC. Santacatterina et al. found that limited oxidative phosphorylation activity in hepatocytes is associated with decreased apoptotic cell death and increased cancer development (Figure 4) [79].
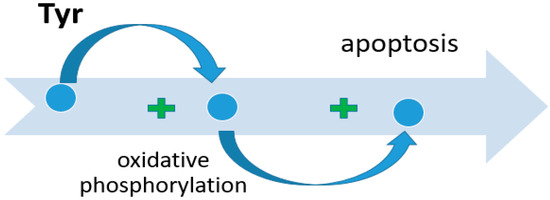
Figure 4.
The role of Tyr in the activation of cell apoptosis.
Changes in the metabolism of Tyr (and other serum amino acids) are observed in many types of cancer pathology. Much research work has been carried out on serum metabolic profiling using a combination of LC-MS and 1H-NMR, for example, by Zhang et al. [80]. Their study was conducted with several groups: a control group, and patients with esophageal adenocarcinoma, Barrett’s esophagus, and high-grade esophageal dysplasia, respectively. The results of this study showed differences between healthy people at risk and cancer patients. There was a gradual depletion of free amino acids: Val, Leu/Ile, Met, Trp, 5-hydroxytryptophan, Tyr, fatty acids with parallel accumulation of glucose, lactate, carnitine, glutamate, and Lys [81,82,83,84,85].
3.5. Methionine
Met plays a significant role in antioxidant processes in oncology [86,87,88,89,90]. According to its physicochemical properties, Met participates in redox reactions, reducing the level of oxidative stress both by stimulating the synthesis of glutathione as the main antioxidant agent and through the formation of methyl sulfoxide (MetO) (Figure 5) [91,92,93,94,95]. The modified form of Met binds reactive oxygen species, thereby suppressing oxidative stress. This effect is most likely to be observed in a generally healthy human body. Moreover, with more significant pathological imbalances, Met can act as one of the reasons for increased oxidative stress [96,97,98].
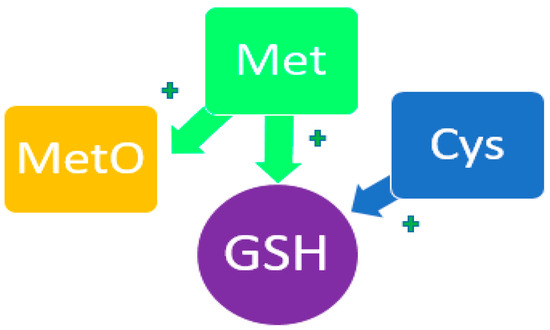
Figure 5.
Participation of Met and His in the redox process. MetO—methyl sulfoxide; GSH—glutathione. The plus symbol indicates potentiation of the synthesis of glutathione and methylsulfoxide.
With an abundant intake of Met from food, the substrate flux through the trans methylation pathway decreases, and that through the trans sulfuration pathway increases. The occurrence of various defects in this pathway can lead to fasting homocysteine levels. Abundant accumulation of this amino acid increases oxidative stress, reduces the availability of nitric oxide, and enhances inflammatory reactions [19,99]. In experimental studies, restricted Met intake has been demonstrated to cause a reduction in oxidative stress. Apparently, during oncological processes, a serious biochemical imbalance and modification of metabolic processes occurs, leading to the depletion of the Met enzymatic system and the accumulation of a large number of intermediate products that have an altering effect [100,101,102,103,104,105,106].
3.6. Cysteine
In various histological types of cancer, a nonspecific increase in total Cys content occurs. This amino acid is involved in the generation of glutathione (a powerful antioxidant molecule) and sulfur-containing products such as taurine, biotin, coenzyme A, and hydrogen sulfide through various metabolic pathways [107,108,109,110]. Cys is found mainly in the extracellular space in the form of cystine and in small amounts in the form of Cys, which subsequently takes part in non-enzymatic antioxidant mechanisms. To better adapt to oxidative stress, tumor cells interact with their microenvironment through the lymphatic network, fibroblasts, and immune cells, thus stimulating the transport of extracellular cystine into the intracellular space and converting cystine to Cys, which in turn supports the synthesis and secretion of glutathione [111] in the tumor microenvironment [112,113,114,115,116,117]. Recent studies have reported that Cys potentiates the reprogramming of carbon and sulfur metabolism, which underlies the adaptation of ovarian cancer cells to a hypoxic microenvironment through the production of hydrogen sulfide, which enhances the uptake of glucose by cancer cells and facilitates the production of reactive oxygen species [118,119,120,121,122].
An increase in Cys content has been demonstrated in several recent studies in different types of cancer: epithelial ovarian cancer [123], colorectal cancer, prostate cancer, papillary thyroid cancer, breast cancer, and chronic myelogenous leukemia [124,125]. The increase in Cys content in such different types of cancer once again emphasizes the nonspecificity of the process of metabolic reprogramming, and more so characterizes the general fundamental processes underlying the life of a cancer cell, which are still far from being fully understood.
3.7. Proline
Pro is a non-essential amino acid. It makes a significant contribution to the formation of protein structures and the determination of protein functions, and also maintains cellular redox homeostasis [126]. Intracellular catabolism of Pro can generate both reactive oxygen species and adenosine triphosphate (ATP). Recent studies have demonstrated that Pro biosynthesis and catabolism are important processes in pathological conditions. This is due not only to the fact that this amino acid is responsible for protein synthesis, but also to the fact that it regulates the redox balance and is a source of energy. Pro is synthesized by the cells of the body and is also supplied with food. The main site of Pro metabolism is the small intestine. About 60% of free hydroxyproline and Pro are released into the bloodstream [3]. Recent studies have demonstrated that there is a significant increase in hydroxyproline concentrations in liver fibrosis and bone tumors [127]. In cachexia [128], carcinogenesis, and diabetes, Pro increases [129]. Increased Pro content in the cerebrospinal fluid and vascular bed of the brain can be used as markers of pathological processes, including oncological ones. Some amino acids, including Pro, due to their large molecular weight, are not able to overcome the blood–brain barrier and enter the brain and spinal cord [130]. Only during pathological processes, when there is a violation of the permeability of the vascular wall, can molecules that previously could not penetrate the brain area in a normal physiological state accumulate in this area, thus affecting the metabolism of the central nervous system [131,132,133].
The enzyme prolineoxidase (PRODH), which is involved in Pro metabolism, can be used as a marker of the oncological process. Elia et al. found an increase in the content of this enzyme in breast cancer. During metastasis, collagen degradation, Pro release, and activation of prolineoxidase occur [134]. Interestingly, prolineoxidase, in turn, promotes tumor progression in non-small cell lung cancer. In this case, the enzyme acts as a chromatin remodeler and lymphoid-specific helicase [135].
Separately, it is worth highlighting the role of Pro in redox homeostasis (Figure 6) [136]. In chronic diseases and oncological conditions [137], a local increase in Pro content occurs [138]. Such pathological conditions are characterized by tissue hypoxia. During inflammation, cytokines are released and stimulate the activation of myofibroblasts and other fibroblast subtypes [139], which are specialized in the deposition of extracellular matrix, including collagen. Collagen consists of 30% prolyl residues. The state of fibrosis during inflammation and cancer requires increased consumption of Pro; therefore, there is an increase in the consumption of exogenous forms of this amino acid and activation of the synthesis of endogenous forms. This can be described as an indirect effect on the redox balance. It is also known that Pro can directly influence redox homeostasis through the removal of reactive oxygen species [140].
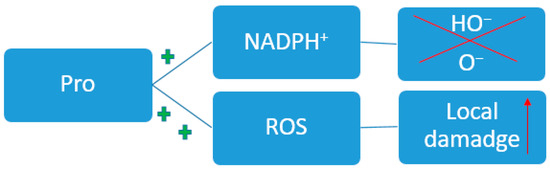
Figure 6.
Participation of Pro in the redox process. ROS—Reactive oxygen species; NADPH—nicotinamide adenine dinucleotide phosphate; the plus symbol indicates an increase in the effect.
The availability of Pro at the systemic and local level affects the balance of proteins [141], nucleotides and cellular NAD(P)+/NAD(P)H [142,143]. When the listed components of homeostasis are disrupted at the level of one cell or group of cells, the local availability of Pro is disrupted, thereby affecting nearby cells. An altered pathological microenvironment is formed.
3.8. Lysine
Lys and its derivatives can be used as biological markers of oxidative stress [144]. Lys, upon oxidation, forms allysine (adipic semialdehyde), a protein carbonyl [145,146]. Additionally, von Pappenheim et al. [147] demonstrated the role of Lys and Cys as a redox switch in nitrogen–oxygen–sulfur (NOS) covalent bridge proteins [148]. In most proteins, the NOS switch contains Lys, which is involved in enzymatic catalysis or binding of substrates, DNA, or effectors, thus linking the chemical properties of Lys and the biological functions of the redox process. This NOS bridge functions as an allosteric switch. By influencing the allosteric center of a protein through NOS, one can change its structure and the structure of its active center, and regulate enzymatic activity (Figure 7) [4].
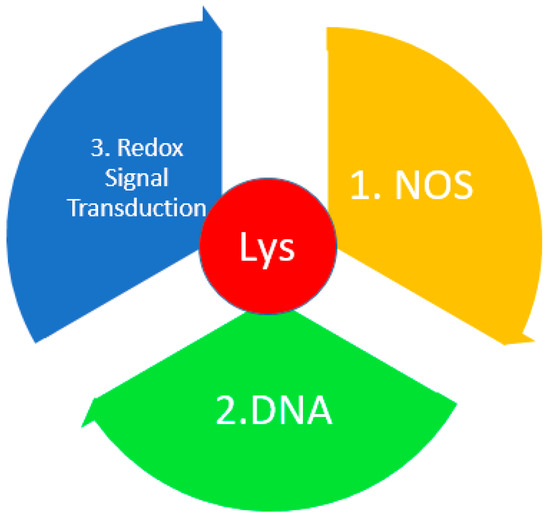
Figure 7.
The role of Lys in redox homeostasis. DNA—deoxyribonucleic acid; NADPH—nicotinamide adenine dinucleotide phosphate; NOS—nitrogen–oxygen–sulfur bridge.
The main discovery of this study is that the family of proteins that are responsible for the response to oxidative stress are involved in the transmission of redox signals; maintain homeostatic balance; are involved in DNA repair, transcription, and translation of ribosomal proteins and degradation proteins [5,6]; and are also involved in signal transduction and biosynthesis of redox-sensitive cofactors contain the amino acids Cys and Lys with great catalytic potential and NOS bridges. With an increase in the content of reactive oxygen species, NOS bridges are formed, which bind to cysteine or lysine, causing their mutations in the allosteric center, thereby changing the structure and function of proteins [149]. These bridges and switches have been found in proteins in diseases such as Alzheimer’s disease, Parkinson’s disease, obesity, autoimmune diseases, cancer, etc. [144].
3.9. Phenylalanine
Schulpis et al. have shown that high levels of Phe damage the structure of DNA, proteins, lipids, and also lead to a decrease in antioxidant protection in the body [150,151]. Using the example of patients with phenylketonuria, damage to proteins and lipids was described by recording an increase in products such as carbonyl, oxidized sulfhydryl, the content of active forms of thiobarbituric acid, and malondialdehyde [18]. It was found that Phe metabolites, phenyllactic and phenylacetic acids have a significant effect on antioxidant enzymes (Figure 8). An increase in the content of superoxide dismutase in tissues was observed [152,153].

Figure 8.
The effect of excess accumulation of Phe on the redox balance.
Recently, more and more information has become available on the concentration of Phe in a particular type of cancer using mass spectrometry. Thus, in their study, Cao et al. demonstrated a high content of the amino acids Phe and Tyr in viable tumor areas of non-small cell lung cancer [154]. Another study found an increase in serum Phe and Tyr in hepatocellular cancer (HCC) [155]. It is noteworthy that there are practically no data on the biochemical processes and reasons for the increase in this amino acid in oncological conditions, and on its role in redox homeostasis.
4. Comparative Analysis of the Role of Amino Acids in Non-Enzymatic Oxidative Mechanisms
Based on the analyzed information, we came to the conclusion that in serious pathological conditions, including cancer, a nonspecific change in amino acid metabolism occurs. The data obtained cannot be used as specific markers for a particular type of cancer. At the same time, changes in the activity of amino acids and their role in redox homeostasis can help us to better understand the pathophysiology of the oncological process, and to identify more effective therapeutic targets.
Table 1 summarizes the changes in the activity of the nine most studied amino acids in relation to redox imbalance in cancer. It has been shown that amino acids can be divided conditionally into two groups: those that decrease in concentration in oncology (Tyr, Trp, His), and those that increase (Arg, Phe, Met, Cys, Pro, Lys).

Table 1.
Amino acid concentrations in different types of cancer.
At the same time, for none of the amino acids considered, multidirectional changes in concentration were shown in different types of cancer, which suggests a similar mechanism of involvement in redox processes regardless of the type of cancer (Table 1).
Table 2 provides a summary of the major amino acid metabolic changes in cancer. Each group is characterized by certain mechanisms of alteration and protection in oncology.
Table 2.
Characteristics of amino acid metabolism in redox imbalance and oncology.
Thus, in the group of amino acids whose activity decreases in oncology, the main mechanisms of protection, in our opinion, lie in the formation of intermediate metabolic products and activation of enzymes that are responsible for maintaining the homeostatic properties of a healthy cell (Table 2). Another option for protection is to potentiate the cytotoxic effect on cancer cells by interacting with other molecules [5].
Among the group of amino acids whose activity and content increases, two mechanisms of action can also be distinguished. The first is the reprogramming of amino acid metabolism, which results in the accumulation of intermediate metabolic products that disrupt the body’s antioxidant defenses. The second is a direct effect on the activity of genes and allosteric centers of proteins, which changes the activity of the enzymatic system.
There is not enough information about changes in metabolism when the redox balance is disturbed both in chronic diseases and in oncology for all amino acids discussed in this review. It seems clear that the role of Phe in maintaining redox homeostasis is significant, but has not been adequately studied. A significant increase in this amino acid leads to the accumulation of intermediate products in the form of acids, which significantly affect redox processes. To date, amino acids and their changes are considered exclusively as a descriptive aspect of the disease, as one of many signs of pathology, and no attention is paid to the basic issues of the nature of this change. The answer lies in a targeted study of the physicochemical and biochemical properties of Phe and its metabolic network with other agents in pathology.
Amino acids such as Pro, His, Trp, Tyr, Cys and Arg seem interesting for further study. To date, a sufficient number of studies have been carried out showing that amino acids and their metabolism are much more complex, and their influence on maintaining protein, nucleotide and energy balance is much more significant [1,2,3,4,5,6,7,8,9,10,11]. Moreover, to understand how this or that cell behaves during pathology, it is necessary to review and deepen your knowledge of the field by studying the most basic components of cell activity both in physiological and pathological conditions. We must observe where that preponderance occurs, which processes maintain homeostasis (protecting against the aggressive effects of foreign agents), and which processes have the opposite effect (on the contrary, aggravating the condition of the body). This review has demonstrated that the nature and role of amino acids is controversial. By providing beneficial effects at the systemic level and damaging cells at the local level, an imbalance can occur, and the accumulation of damage at the local level can lead to damage at the systemic level.
5. Amino Acids as Therapeutic Targets for Cancer Treatment
Scientists around the world are actively exploring the possibility of regulating amino acid metabolism in oncology [158]. In many cases, therapeutic targeting of tumor cell metabolism produces better results with fewer side effects [121]. For example, therapies targeting the regulation of essential amino acids, such as Met, have beneficial effects. This was demonstrated by scientists using dietary Met restriction in mice and rats, which resulted in an increase in the lifespan of mice and rats [159]. It has long been noted that limiting the supply of amino acids leads to a slowdown in tumor growth [159,160].
Thus, the enzyme arginase is attracting increasing interest as a therapeutic target. There are two types of isoforms of this enzyme, arginase 1 (ARG1) and arginase 2 (ARG2) [161]. Abnormal expression of these enzyme isoforms has been reported to frequently occur in cancers such as breast, gastric, colorectal, and liver cancer [162,163,164,165,166]. In addition, signaling pathways associated with arginase, such as arginase/PI3K/RAC-alpha serine/threonine-protein kinase (AKT)/mTOR, arginase/signal transducer and activator of transcription 3 (STAT3), and arginase/mitogen-activated protein kinase (MAPK), can be influenced [167].
For Lys, the enzyme lysine-specific histone demethylase 1A (KDM1A/LSD1) was discovered. It shows activity in relation to the development, metastasis, progression, and therapy resistance of cancer [168].
Pharmacological modification of Cys residues affects the biological activity of this molecule, thereby demonstrating a therapeutic effect [169]. Thus, it is possible to regulate the entry of cysteine into the cell by blocking the xCT/SLC7A11 transporter [100]. SLC7A11 has been found to be widely expressed in a variety of cancers, and has poor survival rates in patients with breast cancer, prostate cancer, and papillary thyroid carcinoma [170]. In prostate cancer patients, a positive association between xCT expression with invasion and metastasis has been widely found, through an influence on the redox status of the tumor microenvironment [171]. It is also possible to influence GSH peroxidase 4 (GPX4), triggering the mechanism of ferroptosis [172,173,174,175,176,177]. A positive effect in cancer therapy was discovered by suppressing the entry of hydrogen sulfide into cancer cells, which, through activation of GLUT, enhances the uptake of glucose and its glycolysis with the subsequent production of ATP, which is spent on maintaining the life of the cancer cell [178]. Hydrogen sulfide is produced by three enzymes, namely cystathionine β-synthase (CBS), cystathionine γ-lyase (CSE), and 3-mercaptopyruvate sulfatansferase (3MST). CBS knockdown reduced oxygen consumption and ATP production in colon and ovarian cancer cells [179,180]. Suppression of CSE expression in endothelial cells reduced their ability to induce angiogenesis in breast cancer cells [181]. Cys may also have an antitumor effect through its byproduct, taurine. Cysteine dioxygenases catalyze the oxidation of Cys to cysteine sulfinate. Cysteine sulfinic acid decarboxylase catalyzes the reaction, and carboxyl groups are removed to form hypotaurine, which subsequently generates taurine [182]. Taurine has been shown to have cytoprotective functions and maintain cell homeostasis [183]. Increased taurine content has shown an inhibitory effect on the growth of colon cancer [184], lung cancer [185], HCC [186], melanoma [187], and breast cancer [188]. There are a number of other possibilities for influencing Cys, providing an antitumor effect, for example, through protein kinases and phosphatases [189,190,191,192].
An antitumor effect was demonstrated when His was added to HCC tumor cells. It considerably reduces the release of cytokines in response to liver injury [193]. There is a decrease in the expression of tumor markers that are associated with glycolysis (GLUT1 and HK2), inflammation (pSTAT3), angiogenesis (VEGFB and VEGFC), stem cells (CD133), metastasis (snail/slug), and cell migration [194].
Pro synthesized by proline synthase PYCR1 through cGMP-PKG enhances the hallmarks of stem cell carcinoma. Knockdown of proline synthetase PYCR1 showed a significant reduction in breast cancer growth [195]. Another therapeutic target is the enzyme proline dehydrogenase (PRODH) [196]. On the one hand, increased expression of PRODH induces apoptosis [197,198] of breast cancer cells. On the other hand, it stimulates metastasis to the lung. Although there is currently no clear conclusion on the stimulation and inhibition of this enzyme, since its role in cancer metabolism is twofold, further study of the possibility of using proline dehydrogenase as a therapeutic target is very important [199]. Inhibition of Pro synthesis itself may also reduce the synthesis of collagen, which is essential for the growth and development of cancer cells, and the extracellular matrix [200]. It is necessary to find a balance between reducing and completely suppressing the synthesis of extracellular matrix components, since its complete ablation can free tumor cells from the limiting barrier [201,202,203].
To limit the availability of Trp and prevent its use along the kynurene pathway, it is possible to influence the direct transport of tryptophan into the cancer cell, suppress the activity of Trp 2,3-dioxygenase (TDO2) and indoleamine 2,3-dioxygenase 1 (IDO1), and inhibit N′-formylkynurenine formamidase (FAMID), Kyn aminotransferase 1 (KAT1) and kynureninase (KYNU), nicotinamide phosphoribosyltransferase (NMPRT), nicotinamide mononucleotide adenylyltransferase (NMNAT) aryl hydrocarbon receptor (AhR), interleukin 4-induced protein 1 (IL-4I1), poly (ADP-ribose) polymerase (PARPs), and protein acetylaters (SIRTs). All this is a potential target for depriving the cancer cell of its ability to maintain its vital activity [204].
Tyrosine kinase inhibitors have become very widely used in the treatment of various types of cancer. Medicines containing this enzyme, in addition to having a positive effect in the treatment of oncology, also have a number of serious side effects. This forces scientists and physicians to look for new ways to use this approach [205].
Phe has not yet been presented as a therapeutic target for the supervision of cancer patients. This area requires additional study and understanding of possible control points in the life of a cancer cell.
6. Conclusions
Research in recent decades has significantly expanded our understanding of the role and functions of amino acids both in health and in cancer. They make a significant contribution to the maintenance of cellular homeostasis, influencing the redox balance and participating in energy metabolism, due to the formation of ATP as a by-product during amino acid metabolism. In addition, amino acid metabolism products are involved in epigenetic regulation and modeling of the immune response, which is associated with tumorigenesis and metastasis. Transporters and transaminases especially attract attention, since they mediate the absorption and synthesis of amino acids, which in the future can be used as markers of metabolic disorders in oncology, and act as a therapeutic target. It is currently not possible to talk about specific mechanisms for increasing or decreasing the activity of certain amino acids, since the topic of amino acids and their metabolism in oncology remains not fully studied.
This area of research is quite promising, as it will allow us to deepen our understanding of the principles of the life of a cancer cell, and will allow us to develop new, more effective therapeutic targets for cancer patients, the number of whom is growing every year (and whose age is becoming younger); it will also allow us to revise and develop a new concept of understanding the properties and functions of amino acids. In the future, there are plans to study changes in metabolism and their impact more deeply, alongside studies of redox homeostasis in specific types of cancer.
Author Contributions
Conceptualization, E.I.D. and L.V.B.; methodology, L.V.B.; validation, E.I.D. and L.V.B.; formal analysis, L.V.B.; investigation, E.I.D.; resources, L.V.B.; data curation, E.I.D.; writing—original draft preparation, E.I.D.; writing—review and editing, L.V.B.; visualization, E.I.D.; supervision, L.V.B.; project administration, L.V.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 23-15-00188, https://rscf.ru/project/23-15-00188/ (accessed on 28 December 2023).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Isenberg, J.S.; Martin-Manso, G.; Maxhimer, J.B.; Roberts, D.D. Regulation of nitric oxide signalling by thrombospondin 1: Implications for anti-angiogenic therapies. Nat. Rev. Cancer 2009, 9, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, D.; Kashiwagi, S.; Jain, R.K. The role of nitric oxide in tumour progression. Nat. Rev. Cancer 2006, 6, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Wensien, M.; von Pappenheim, F.R.; Funk, L.M.; Kloskowski, P.; Curth, U.; Diederichsen, U.; Uranga, J.; Ye, J.; Fang, P.; Pan, K.T.; et al. A lysine-cysteine redox switch with an NOS bridge regulates enzyme function. Nature 2021, 593, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.S.; Stanley, B.A.; Pegg, A.E. Effect of mutations at active site residues on the activity of ornithine decarboxylase and its inhibition by active site-directed irreversible inhibitors. J. Biol. Chem. 1993, 268, 24572–24579. [Google Scholar] [CrossRef]
- Obin, M.; Shang, F.; Gong, X.; Handelman, G.; Blumberg, J.; Taylor, A. Redox regulation of ubiquitin-conjugating enzymes: Mechanistic insights using the thiol-specific oxidant diamide. FASEB J. 1998, 12, 561–569. [Google Scholar] [CrossRef]
- Wall, S.B.; Oh, J.Y.; Diers, A.R.; Landar, A. Oxidative modification of proteins: An emerging mechanism of cell signaling. Front. Physiol. 2012, 14, 369. [Google Scholar] [CrossRef]
- Vučetić, M.; Cormerais, Y.; Parks, S.K.; Pouysségur, J. The Central Role of Amino Acids in Cancer Redox Homeostasis: Vulnerability Points of the Cancer Redox Code. Front. Oncol. 2017, 21, 319. [Google Scholar] [CrossRef]
- Shtarkman, I.N.; Gudkov, S.V.; Chernikov, A.V.; Bruskov, V.I. Effect of amino acids on X-ray-induced hydrogen peroxide and hydroxyl radical formation in water and 8-oxoguanine in DNA. Biochemistry 2008, 73, 470–478. [Google Scholar] [CrossRef]
- Du, J.; Gebicki, J.M. Proteins are major initial cell targets of hydroxyl free radicals. Int. J. Biochem. Cell Biol. 2004, 36, 2334–2343. [Google Scholar] [CrossRef]
- Hayashi, T.; Juilet, P.A.R.; Matsui Hirai, H.; Miyazaki, A.; Fukatsu, A.; Funami, J.; Iguchi, A.; Ignarro, L.J. l-citrulline and l-arginine supplementation retards the progression of high-cholesterol-diet-induced atherosclerosis in rabbits. Proc. Natl. Acad. Sci. USA 2005, 102, 13681–13686. [Google Scholar] [CrossRef]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.A.; Livesey, S.A.; Zhu, S. A re-evaluation of the antioxidant activity of purified carnosine. Biochemistry 2000, 65, 901–906. [Google Scholar]
- Milligan, J.R.; Aguilera, J.A.; Ly, A.; Tran, N.Q.; Hoang, O.; Ward, J.F. Repair of oxidative DNA damage by amino acids. Nucl. Acids Res. 2003, 31, 6258–6263. [Google Scholar] [CrossRef] [PubMed]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Oxidation of free amino acids and amino acid residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, V.A.; Kornienko, I.V.; Kletsky, M.E.; Kornienko, I.E.; Lisitsyn, A.S.; Novikov, V.V. The superoxide-scavenging activity of some amino acids in water solutions. Biophysics 2005, 50, 601–605. [Google Scholar]
- Abu-Serie, M.M.; El-Gamal, B.A.; El-Kersh, M.A. Investigation into the antioxidant role of arginine in the treatment and the protection for intralipid-induced non-alcoholic steatohepatitis. Lipids Health Dis. 2015, 14, 128. [Google Scholar] [CrossRef]
- Chen, C.L.; Hsu, S.C.; Ann, D.K.; Yen, Y.; Kung, H.J. Arginine Signaling and Cancer Metabolism. Cancers 2021, 13, 3541. [Google Scholar] [CrossRef]
- Lass, A.; Suessnbacher, A.; Wolkart, G.; Mayer, B.; Brunner, F. Functional and analytical evidence for scavenging of oxygen radicals by L-arginine. Mol. Pharmacol. 2002, 61, 1081–1088. [Google Scholar] [CrossRef]
- Rubbo, H.; Radi, R.; Trujillo, M.; Telleri, R.; Kalyanaraman, B.; Barnes, S.; Kirk, M.; Freeman, B.A. Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid peroxidation. J. Biol. Chem. 1994, 269, 26066–26075. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Cederbaum, A. Nitric oxide donors prevent while the nitric oxide synthase inhibitor L-NAME increases arachidonic acid plus CYP2E1-dependent toxicity. Toxicol. Appl. Pharmacol. 2006, 216, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Jokelainen, K.; Lau, G.K.; Rahemtulla, A.; Tipoe, G.L.; Polavarapu, R.; Lalani, E.N. Arginine reverses ethanol-induced inflammatory and fibrotic changes in liver despite continued ethanol administration. J. Pharmacol. Exp. Ther. 2001, 299, 832–839. [Google Scholar] [PubMed]
- Klatt, P.; Lamas, S. Regulation of protein function by S-glutathiolation in response to oxidative and nitrosative stress. Eur. J. Biochem. 2000, 267, 4928–4944. [Google Scholar] [CrossRef] [PubMed]
- Lube, C.B.; Hayn, M.; Kitzmuller, E.; Vierhapper, H.; Lubec, G. L-arginine reduces lipid peroxidation in patients with diabetes mellitus. Free Radic. Biol. Med. 1997, 22, 355–357. [Google Scholar] [CrossRef]
- Matsubara, A.; Tamai, K.; Matsuda, Y.; Niwa, Y.; Morita, H.; Tomida, K.; Armstrong, D.; Ogura, Y. Protective effect of polyethylene glycol-superoxide dismutase on leukocyte dynamics in rat retinal microcirculation under lipid hydroperoxide-induced oxidative stress. Exp. Eye Res. 2005, 81, 193–199. [Google Scholar] [CrossRef]
- Briviba, K.; Kissner, R.; Koppenol, W.H.; Sies, H. Kinetic study of the reaction of glutathione peroxidase with peroxynitrite. Chem. Res. Toxicol. 1998, 11, 1398–1401. [Google Scholar] [CrossRef]
- Palmer, R.M.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model. Mech. 2018, 11, 11. [Google Scholar] [CrossRef]
- Wink, D.A.; Miranda, K.M.; Espey, M.G.; Pluta, R.M.; Hewett, S.J.; Colton, C.; Vitek, M.; Feelisch, M.; Grisham, M.B. Mechanisms of the antioxidant effects of nitric oxide. Antioxid. Redox Signal. 2001, 3, 203–213. [Google Scholar] [CrossRef]
- Lala, P.K.; Chakraborty, C. Role of nitric oxide in carcinogenesis and tumour progression. Lancet Oncol. 2001, 2, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Ambs, S.; Ogunfusika, M.O.; Merriam, W.G.; Bennett, W.P.; Billiar, T.R.; Harris, C.C. Up-regulation of inducible nitric oxide synthase expression in cancer-prone p53 knockout mice. Proc. Natl. Acad. Sci. USA 1998, 95, 8823–8828. [Google Scholar] [CrossRef] [PubMed]
- Radisavljevic, Z. Inactivated tumor suppressor Rb by nitric oxide promotes mitosis in human breast cancer cells. J. Cell. Biochem. 2004, 92, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.N.; Hsieh, F.J.; Cheng, Y.M.; Chang, K.J.; Lee, P.H. Expression of inducible nitric oxide synthase and cyclooxygenase-2 in angiogenesis and clinical outcome of human gastric cancer. J. Surg. Oncol. 2006, 94, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Lee, S.G.; Song, S.H.; Heo, D.S.; Park, B.J.; Lee, D.W.; Kim, K.H.; Sung, M.W. The effect of nitric oxide on cyclooxygenase-2 (COX-2) overexpression in head and neck cancer cell lines. Int. J. Cancer 2003, 107, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Dhar, D.K.; Yamaguchi, E.; Maruyama, S.; Sato, T.; Hayashi, H.; Ono, T.; Yamanoi, A.; Kohno, H.; Nagasue, N. Coexpression of inducible nitric oxide synthase and COX-2 in hepatocellular carcinoma and surrounding liver: Possible involvement of COX-2 in the angiogenesis of hepatitis C virus-positive cases. Clin. Cancer Res. 2001, 7, 1325–1332. [Google Scholar] [PubMed]
- Garrido, P.; Shalaby, A.; Walsh, E.M.; Keane, N.; Webber, M.; Keane, M.M.; Sullivan, F.J.; Kerin, M.J.; Callagy, G.; Ryan, A.E.; et al. Impact of inducible nitric oxide synthase (iNOS) expression on triple negative breast cancer outcome and activation of EGFR and ERK signaling pathways. Oncotarget 2017, 8, 80568–80588. [Google Scholar] [CrossRef]
- Lopez-Rivera, E.; Jayaraman, P.; Parikh, F.; Davies, M.A.; Ekmekcioglu, S.; Izadmehr, S.; Milton, D.R.; Chipuk, J.E.; Grimm, E.A.; Estrada, Y.; et al. Inducible nitric oxide synthase drives mTOR pathway activation and proliferation of human melanoma by reversible nitrosylation of TSC2. Cancer Res. 2014, 74, 1067–1078. [Google Scholar] [CrossRef]
- Tsuber, V.; Kadamov, Y.; Brautigam, L.; Berglund, U.W.; Helleday, T. Mutations in Cancer Cause Gain of Cysteine, Histidine, and Tryptophan at the Expense of a Net Loss of Arginine on the Proteome Level. Biomolecules 2017, 7, 49. [Google Scholar] [CrossRef]
- Poillet-Perez, L.; Xie, X.; Zhan, L.; Yang, Y.; Sharp, D.W.; Hu, Z.S.; Su, X.; Maganti, A.; Jiang, C.; Lu, W.; et al. Autophagy maintains tumour growth through circulating arginine. Nature 2018, 563, 569–573. [Google Scholar] [CrossRef]
- Musso, T.; Gusella, G.L.; Brooks, A.; Longo, D.L.; Varesio, L. Interleukin-4 inhibits indoleamine 2,3-dioxygenase expression in human monocytes. Blood 1994, 83, 1408–1411. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sorgdrager, F.J.H.; Naudé, P.J.W.; Kema, I.P.; Nollen, E.A.; Deyn, P.P. Tryptophan Metabolism in Inflammaging: From Biomarker to Therapeutic Target. Front. Immunol. 2019, 10, 2565. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Kang, M.; Martin, A. Microbiome and colorectal cancer: Unraveling host-microbiota interactions in colitis-associated colorectal cancer development. Semin. Immunol. 2017, 32, 3–13. [Google Scholar] [CrossRef]
- Itzkowitz, S.H.; Yio, X. Inflammation and cancer IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, 7–17. [Google Scholar] [CrossRef]
- Liu, L.; Su, X.; Quinn, W.J.; Hui, S.; Krukenberg, K.; Frederick, D.W.; Redpath, P.; Zhan, L.; Chellappa, K.; White, E.; et al. Quantitative Analysis of NAD Synthesis-Breakdown Fluxes. Cell Metab. 2018, 27, 1067–1080. [Google Scholar] [CrossRef]
- Colabroy, K.L.; Begley, T.P. Tryptophan catabolism: Identification and characterization of a new degradative pathway. J. Bacteriol. 2005, 187, 7866–7869. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ metabolism: Pathophysiologic mechanisms and therapeutic potential. Signal Transduct. Target. Ther. 2020, 5, 227. [Google Scholar] [CrossRef] [PubMed]
- Sallin, O.; Reymond, L.; Gondrand, C.; Raith, F.; Koch, B.; Johnsson, K. Semisynthetic biosensors for mapping cellular concentrations of nicotinamide adenine dinucleotides. Elife 2018, 7, 32638. [Google Scholar] [CrossRef] [PubMed]
- Heng, B.; Lim, C.K.; Lovejoy, D.B.; Bessede, A.; Gluch, L.; Guillemin, G.J. Understanding the role of the kynurenine pathway in human breast cancer immunobiology. Oncotarget 2016, 7, 6506–6520. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 6, 360438. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.E.; Leopold, M.C.; Huang, X.; Atwood, C.S.; Saunders, A.J.; Hartshorn, M.; Lim, J.T.; Faget, K.Y.; Muffat, J.A.; Scarpa, R.C.; et al. 3-Hydroxykynurenine and 3-hydroxyanthranilic acid generate hydrogen peroxide and promote alpha-crystallin cross-linking by metal ion reduction. Biochemistry 2000, 39, 7266–7275. [Google Scholar] [CrossRef]
- Hiraku, Y.; Inoue, S.; Oikawa, S.; Yamamoto, K.; Tada, S.; Nishino, K.; Kawanishi, S. Metal-mediated oxidative damage to cellular and isolated DNA by certain tryptophan metabolites. Carcinogenesis 1995, 16, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.; Grieco, D. New insights on oxidative stress in cancer. Curr. Opin. Drug Discov. Dev. 2009, 12, 240–245. [Google Scholar]
- Smith, A.J.; Smith, R.A.; Stone, T.W. 5-Hydroxyanthranilic acid, a tryptophan metabolite, generates oxidative stress and neuronal death via p38 activation in cultured cerebellar granule neurones. Neurotox. Res. 2009, 15, 303–310. [Google Scholar] [CrossRef]
- Hiramatsu, R.; Hara, T.; Akimoto, H.; Takikawa, O.; Kawabe, T.; Isobe, K.; Nagase, F. Cinnabarinic acid generated from 3-hydroxyanthranilic acid strongly induces apoptosis in thymocytes through the generation of reactive oxygen species and the induction of caspase. J. Cell Biochem. 2008, 103, 42–53. [Google Scholar] [CrossRef]
- Krause, D.; Suh, H.S.; Tarassishin, L.; Cui, Q.L.; Durafourt, B.A.; Choi, N.; Bauman, A.; Cosenza-Nashat, M.; Antel, J.P.; Zhao, M.L.; et al. The tryptophan metabolite 3-hydroxyanthranilic acid plays anti-inflammatory and neuroprotective roles during inflammation: Role of hemeoxygenase-1. Am. J. Pathol. 2011, 179, 1360–1372. [Google Scholar] [CrossRef]
- Backhaus, C.; Rahman, H.; Scheffler, S.; Laatsch, H.; Hardeland, R. NO scavenging by 3-hydroxyanthranilic acid and 3-hydroxykynurenine: N-nitrosation leads via oxadiazoles to o-quinone diazides. Nitric Oxide 2008, 19, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.I.; Collins, C.A.; Stone, T.W.; Jacob, C. Electrochemical and in vitro evaluation of the redox-properties of kynurenine species. Biochem. Biophys. Res. Commun. 2003, 300, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Latini, A.; Rodriguez, M.; Borba, R.R.; Scussiato, K.; Leipnitz, G.; Reis de Assis, D.; da Costa Ferreira, G.; Funchal, C.; Jacques-Silva, M.C.; Buzin, L.; et al. 3-Hydroxyglutaric acid moderately impairs energy metabolism in brain of young rats. Neuroscience 2005, 135, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Leipnitz, G.; Schumacher, C.; Dalcin, K.B.; Scussiato, K.; Solano, A.; Funchal, C.; Dutra-Filho, C.S.; Wyse, A.T.; Wannmacher, C.M.; Latini, A.; et al. In vitro evidence for an antioxidant role of 3-hydroxykynurenine and 3-hydroxyanthranilic acid in the brain. Neurochem. Int. 2007, 50, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Pláteník, J.; Stopka, P.; Vejrazka, M.; Stípek, S. Quinolinic acid-iron(ii) complexes: Slow autoxidation, but enhanced hydroxyl radical production in the Fenton reaction. Free Radic. Res. 2001, 34, 445–459. [Google Scholar] [CrossRef]
- Kubicova, L.; Hadacek, F.; Chobot, V. Quinolinic Acid: Neurotoxin or Oxidative Stress Modulator? Int. J. Mol. Sci. 2013, 14, 21328–21338. [Google Scholar] [CrossRef]
- Kumar, N.; Rachagani, S.; Natarajan, G.; Crook, A.; Gopal, T.; Rajamanickam, V.; Kaushal, J.B.; Nagabhishek, S.N.; Powers, R.; Batra, S.K.; et al. Histidine Enhances the Anticancer Effect of Gemcitabine against Pancreatic Cancer via Disruption of Amino Acid Homeostasis and Oxidant—Antioxidant Balance. Cancers 2023, 15, 2593. [Google Scholar] [CrossRef]
- Cantoni, O.; Sestili, P.; Guidarelli, A.; Giacomoni, P.U.; Cattabeni, F. Effects of L-histidine on hydrogen peroxide-induced DNA damage and cytotoxicity in cultured mammalian cells. Mol. Pharmacol. 1992, 41, 969–974. [Google Scholar]
- Herman, S.; Niemelä, V.; Emami Khoonsari, P.; Sundblom, J.; Burman, J.; Landtblom, A.M.; Spjuth, O.; Nyholm, D.; Kultima, K. Alterations in the tyrosine and phenylalanine pathways revealed by biochemical profiling in cerebrospinal fluid of Huntington’s disease subjects. Sci. Rep. 2019, 9, 4129. [Google Scholar] [CrossRef]
- Lai, H.S.; Lee, J.C.; Lee, P.H.; Wang, S.T.; Chen, W.J. Plasma free amino acid profile in cancer patients. Semin. Cancer Biol. 2005, 15, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, T.; Kumar, S.; Markar, S.R.; Antonowicz, S.; Hanna, G.B. Tyrosine, phenylalanine, and tryptophan in gastroesophageal malignancy: A systematic review. Cancer Epidemiol. Biomark. Prev. 2015, 24, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Nguyen, H.Q.; Le, D.H. Unveiling prognostics biomarkers of tyrosine metabolism reprogramming in liver cancer by cross-platform gene expression analyses. PLoS ONE 2020, 15, e0229276. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Dong, S.S.; Xie, Y.W.; Tai, L.S.; Chen, L.; Kong, K.L.; Man, K.; Xie, D.; Li, Y.; Cheng, Y.; et al. Down-regulation of tyrosine aminotransferase at a frequently deleted region 16q22 contributes to the pathogenesis of hepatocellular carcinoma. Hepatology 2010, 51, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Lazebnik, Y. Caspase-9 and APAF-1 form an active holoenzyme. Genes Dev. 1999, 13, 3179–3184. [Google Scholar] [CrossRef] [PubMed]
- Santacatterina, F.; Sánchez-Cenizo, L.; Formentini, L.; Mobasher, M.A.; Casas, E.; Rueda, C.B.; Martínez-Reyes, I.; Núñez de Arenas, C.; García-Bermúdez, J.; Zapata, J.M.; et al. Down-regulation of oxidative phosphorylation in the liver by expression of the ATPase inhibitory factor 1 induces a tumor-promoter metabolic state. Oncotarget 2016, 7, 490–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bowers, J.; Liu, L.; Wei, S.; Gowda, G.A.; Hammoud, Z.; Raftery, D. Esophageal cancer metabolite biomarkers detected by LC-MS and NMR methods. PLoS ONE 2012, 7, e30181. [Google Scholar] [CrossRef]
- Walenta, S.; Schroeder, T.; Mueller-Klieser, W. Lactate in solid malignant tumors: Potential basis of a metabolic classification in clinical oncology. Curr. Med. Chem. 2004, 11, 2195–2204. [Google Scholar] [CrossRef]
- Wu, H.; Xue, R.; Lu, C.; Deng, C.; Liu, T.; Zeng, H.; Wang, Q.; Shen, X. Metabolomic study for diagnostic model of oesophageal cancer using gas chromatography/mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 3111–3117. [Google Scholar] [CrossRef]
- Ayxiam, H.; Ma, H.; Ilyar, S.; Zhang, L.W.; Ablizi, A.; Batur, M.; Lu, X.M. Metabonomic variation of esophageal cancer within different ethnic groups in Xinjiang, China. Zhonghua Yu Fang. Yi Xue Za Zhi 2009, 43, 591–596. [Google Scholar]
- Boros, L.G. Metabolic targeted therapy of cancer: Current tracer technologies and future drug design strategies in the old metabolic network. Metabolomics 2005, 1, 11–15. [Google Scholar] [CrossRef]
- Maguire, G.; Lee, P.; Manheim, D.; Boros, L. SiDMAP: A metabolomics approach to assess the effects of drug candidates on the dynamic properties of biochemical pathways. Expert. Opin. Drug Discov. 2006, 1, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Fan, T.W. NMR-based Stable Isotope Resolved Metabolomics in systems biochemistry. Arch. Biochem. Biophys. 2017, 628, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Aledo, J.C.; Cantón, F.R.; Veredas, F.J. Sulphur atoms from methionines interacting with aromatic residues are less prone to oxidation. Sci. Rep. 2015, 5, 16955. [Google Scholar] [CrossRef] [PubMed]
- Aledo, J.C.; Li, Y.; de Magalhães, J.P.; Ruíz-Camacho, M.; Pérez-Claros, J.A. Mitochondrially encoded methionine is inversely related to longevity in mammals. Aging Cell 2011, 10, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Aledo, J.C.; Valverde, H.; De Magalhães, J.P. Mutational bias plays an important role in shaping longevity-related amino acid content in mammalian mtDNA-encoded proteins. J. Mol. Evol. 2012, 74, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Shuvalov, O.; Petukhov, A.; Daks, A.; Fedorova, O.; Vasileva, E.; Barlev, N.A. One-carbon metabolism and nucleotide biosynthesis as attractive targets for anticancer therapy. Oncotarget 2017, 8, 23955–23977. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef]
- Green, C.R.; Wallace, M.; Divakaruni, A.S.; Phillips, S.A.; Murphy, A.N.; Ciaraldi, T.P.; Metallo, C.M. Branched-chain amino acid catabolism fuels adipocyte differentiation and lipogenesis. Nat. Chem. Biol. 2016, 12, 15–21. [Google Scholar] [CrossRef]
- Carew, L.B.; McMurtry, J.P.; Alster, F.A. Efects of methionine defciencies on plasma levels of thyroid hormones, insulin-like growth factors-I and -II, liver and body weights, and feed intake in growing chickens. Poult. Sci. 2003, 82, 1932–1938. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yu, L.; Fang, J.; Hu, C.A.; Yin, J.; Ni, H.; Ren, W.; Duraipandiyan, V.; Chen, S.; Al-Dhabi, N.A.; et al. Methionine restriction on oxidative stress and immune response in dss-induced colitis mice. Oncotarget 2017, 8, 44511–44520. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ji, Y.; Wu, G.; Sun, K.; Dai, Z.; Wu, Z. Dietary L-methionine restriction decreases oxidative stress in porcine liver mitochondria. Exp. Gerontol. 2015, 65, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Y.; Sun, J.; Zhang, J.; Guo, H.; Shi, Y.; Cheng, X.; Tang, X.; Le, G. Dietary methionine restriction reduces hepatic steatosis and oxidative stress in high-fat-fed mice by promoting H2S production. Food Funct. 2019, 10, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Roman, I.; Barja, G. Regulation of longevity and oxidative stress by nutritional interventions: Role of methionine restriction. Exp. Gerontol. 2013, 48, 1030–1042. [Google Scholar] [CrossRef]
- Maddineni, S.; Nichenametla, S.; Sinha, R.; Wilson, R.P.; Richie, J.P., Jr. Methionine restriction affects oxidative stress and glutathione-related redox pathways in the rat. Exp. Biol. Med. 2013, 238, 392–399. [Google Scholar] [CrossRef]
- Pamplona, R.; Barja, G. Mitochondrial oxidative stress, aging and caloric restriction: The protein and methionine connection. Biochim. Biophys. Acta. 2006, 1757, 496–508. [Google Scholar] [CrossRef]
- Caro, P.; Gomez, J.; Lopez-Torres, M.; Sanchez, I.; Naudi, A.; Jove, M.; Pamplona, R.; Barja, G. Forty percent and eighty percent methionine restriction decrease mitochondrial ROS generation and oxidative stress in rat liver. Biogerontology 2008, 9, 183–196. [Google Scholar] [CrossRef]
- Chevallier, V.; Zoller, M.; Kochanowski, N.; Andersen, M.R.; Workman, C.T.; Malphettes, L. Use of novel cystine analogs to decrease oxidative stress and control product quality. J. Biotechnol. 2021, 327, 1–8. [Google Scholar] [CrossRef]
- Paul, B.D.; Sbodio, J.I.; Snyder, S.H. Cysteine metabolism in neuronal redox homeostasis. Trends Pharmacol. Sci. 2018, 39, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.M.; Chandler, J.D.; Jones, D.P. The cysteine proteome. Free Radic. Biol. Med. 2015, 84, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Scozzafava, A.; Supuran, C.T. Cysteine-modifying agents: A possible approach for effective anticancer and antiviral drugs. Environ. Health Perspect. 2002, 110, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Balendiran, G.K.; Dabur, R.; Fraser, D. The role of glutathione in cancer. Cell Biochem. Funct. 2004, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Stepka, P.; Vsiansky, V.; Raudenska, M.; Gumulec, J.; Adam, V.; Masarik, M. Metabolic and amino acid alterations of the tumor microenvironment. Curr. Med. Chem. 2021, 28, 1270–1289. [Google Scholar] [CrossRef]
- Wang, W.; Kryczek, I.; Dostál, L.; Lin, H.; Tan, L.; Zhao, L.; Lu, F.; Wei, S.; Maj, T.; Peng, D.; et al. Effector T cells abrogate stroma-mediated chemoresistance in ovarian cancer. Cell 2016, 165, 1092–1105. [Google Scholar] [CrossRef]
- Lewerenz, J.; Hewett, S.J.; Huang, Y.; Lambros, M.; Gout, P.W.; Kalivas, P.W.; Massie, A.; Smolders, I.; Methner, A.; Pergande, M.; et al. The cystine/glutamate antiporter system xc− in health and disease: From molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 522–555. [Google Scholar] [CrossRef]
- Min, J.Y.; Chun, K.S.; Kim, D.H. The versatile utility of cysteine as a target for cancer treatment. Front. Oncol. 2023, 12, 997919. [Google Scholar] [CrossRef]
- Lim, J.K.M.; Delaidelli, A.; Minaker, S.W.; Zhang, H.F.; Colovic, M.; Yang, H.; Negri, G.L.; von Karstedt, S.; Lockwood, W.W.; Schaffer, P.; et al. Cystine/glutamate antiporter xCT (SLC7A11) facilitates oncogenic RAS transformation by preserving intracellular redox balance. Proc. Natl. Acad. Sci. USA 2019, 116, 9433–9442. [Google Scholar] [CrossRef]
- Nunes, S.C.; Ramos, C.; Santos, I.; Mendes, C.; Silva, F.; Vicente, J.B.; Pereira, S.A.; Félix, A.; Gonçalves, L.G.; Serpa, J. Cysteine boosts fitness under hypoxia-mimicked conditions in ovarian cancer by metabolic reprogramming. Front. Cell Dev. Biol. 2021, 9, 722412. [Google Scholar] [CrossRef]
- Nunes, S.C.; Lopes-Coelho, F.; Gouveia-Fernandes, S.; Ramos, C.; Pereira, S.A.; Serpa, J. Cysteine boosters the evolutionary adaptation to CoCl2 mimicked hypoxia conditions, favouring carboplatin resistance in ovarian cancer. BMC Evol. Biol. 2018, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.C.; Ramos, C.; Lopes-Coelho, F.; Sequeira, C.O.; Silva, F.; Gouveia-Fernandes, S.; Rodrigues, A.; Guimarães, A.; Silveira, M.; Abreu, S.; et al. Cysteine allows ovarian cancer cells to adapt to hypoxia and to escape from carboplatin cytotoxicity. Sci. Rep. 2018, 8, 9513. [Google Scholar] [CrossRef] [PubMed]
- Fruehauf, J.P.; Meyskens, F.L. Reactive oxygen species: A breath of life or death? Clin. Cancer Res. 2007, 13, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Zhang, W.; Wu, L.; Yang, G.; Li, H.; Wang, R. Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production. Proc. Natl. Acad. Sci. USA 2012, 109, 2943–2948. [Google Scholar] [CrossRef] [PubMed]
- Akaike, T.; Ida, T.; Wei, F.Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef] [PubMed]
- Koppula, P.; Zhang, Y.; Zhuang, L.; Gan, B. Amino acid transporter SLC7A11/xCT at the crossroads of regulating redox homeostasis and nutrient dependency of cancer. Cancer Commun. 2018, 381, 12. [Google Scholar] [CrossRef] [PubMed]
- Chaiswing, L.; Zhong, W.; Liang, Y.; Jones, D.P.; Oberley, T.D. Regulation of prostate cancer cell invasion by modulation of extra- and intracellular redox balance. Free Radic. Biol. Med. 2012, 52, 452–461. [Google Scholar] [CrossRef]
- Onesti, C.E.; Boemer, F.; Josse, C.; Leduc, S.; Bours, V.; Jerusalem, G. Tryptophan catabolism increases in breast cancer patients compared to healthy controls without affecting the cancer outcome or response to chemotherapy. J. Transl. Med. 2019, 17, 239. [Google Scholar] [CrossRef]
- D’Amato, N.C.; Rogers, T.J.; Gordon, M.A.; Greene, L.I.; Cochrane, D.R.; Spoelstra, N.S.; Nemkov, T.G.; D’Alessandro, A.; Hansen, K.C.; Richer, J.K. A TDO2-AhR signaling axis facilitates anoikis resistance and metastasis in triple-negative breast cancer. Cancer Res. 2015, 75, 4651–4664. [Google Scholar] [CrossRef]
- Papadimitriou, N.; Gunter, M.J.; Murphy, N.; Gicquiau, A.; Achaintre, D.; Brezina, S.; Gumpenberger, T.; Baierl, A.; Ose, J.; Geijsen, A.J.M.R.; et al. Circulating tryptophan metabolites and risk of colon cancer: Results from case-control and prospective cohort studies. Int. J. Cancer 2021, 149, 1659–1669. [Google Scholar] [CrossRef]
- Lukey, M.J.; Katt, W.P.; Cerione, R.A. Targeting amino acid metabolism for cancer therapy. Drug Discov. Today 2017, 22, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.N.; Buttar, H.S. Evaluation of the antioxidant properties of tryptophan and its metabolites in in vitro assay. J. Complement. Integr. Med. 2016, 13, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Baldea, I.; Mocan, T.; Cosgarea, R. The role of ultraviolet radiation and tyrosine stimulated melanogenesis in the induction of oxidative stress alterations in fair skin melanocytes. Exp. Oncol. 2009, 31, 200–208. [Google Scholar] [PubMed]
- Shinohara, M.; Adachi, Y.; Mitsushita, J.; Kuwabara, M.; Nagasawa, A.; Harada, S.; Furuta, S.; Zhang, Y.; Seheli, K.; Miyazaki, H.; et al. Reactive oxygen generated by NADPH oxidase 1 (Nox1) contributes to cell invasion by regulating matrix metalloprotease-9 production and cell migration. J. Biol. Chem. 2010, 285, 4481–4488. [Google Scholar] [CrossRef] [PubMed]
- Vettore, L.A.; Westbrook, R.L.; Tennant, D.A. Proline metabolism and redox; maintaining a balance in health and disease. Amino Acids 2021, 53, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; Alghadir, A.H.; Sherif, Y.E.; Ghfar, A.A. Hydroxyproline as a Biomarker in Liver Disease. In Biomarkers in Liver Disease; Springer: Dordrecht, The Netherlands, 2016; pp. 1–21. [Google Scholar]
- Newton, H.; Wang, Y.F.; Camplese, L.; Mokochinski, J.B.; Kramer, H.B.; Brown, A.E.X.; Fets, L.; Hirabayashi, S. Systemic muscle wasting and coordinated tumour response drive tumourigenesis. Nat. Commun. 2020, 11, 4653. [Google Scholar] [CrossRef]
- Liu, Z.; Jeppesen, P.B.; Gregersen, S.; Bach, L.L.; Hermansen, K. Chronic Exposure to Proline Causes Aminoacidotoxicity and Impaired Beta-Cell Function: Studies In Vitro. Rev. Diabet. Stud. 2016, 13, 66–78. [Google Scholar] [CrossRef]
- Arvanitis, C.D.; Ferraro, G.B.; Jain, R.K. The blood-brain barrier and blood-tumour barrier in brain tumours and metastases. Nat. Rev. Cancer 2020, 20, 26–41. [Google Scholar] [CrossRef]
- Oldendorf, W.H. Brain uptake of radiolabeled amino acids, amines, and hexoses after arterial injection. Am. J. Physiol. 1971, 221, 1629–1639. [Google Scholar] [CrossRef]
- Sershen, H.; Lajtha, A. Capillary transport of amino acids in the developing brain. Exp. Neurol. 1976, 53, 465–474. [Google Scholar] [CrossRef]
- Yudilevich, D.L.; De Rose, N.; Sepúlveda, F.V. Facilitated transport of amino acids through the blood-brain barrier of the dog studied in a single capillary circulation. Brain Res. 1972, 44, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grünewald, T.G.P.; Fendt, S.M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017, 8, 15267. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mao, C.; Wang, M.; Liu, N.; Ouyang, L.; Liu, S.; Tang, H.; Cao, Y.; Liu, S.; Wang, X.; et al. Cancer progression is mediated by proline catabolism in non-small cell lung cancer. Oncogene 2020, 39, 2358–2376. [Google Scholar] [CrossRef] [PubMed]
- Schwörer, S.; Berisa, M.; Violante, S.; Qin, W.; Zhu, J.; Hendrickson, R.C.; Cross, J.R.; Thompson, C.B. Proline biosynthesis is a vent for TGFβ-induced mitochondrial redox stress. EMBO J. 2020, 39, 103334. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.; Guterman, I.; Palacios, G.R.; Britton, R.G.; Burschowsky, D.; Tufarelli, C.; Rufini, A. The Janus-like role of proline metabolism in cancer. Cell Death Discov. 2020, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, R.J. Fibroblasts and myofibroblasts: Their source, function and role in disease. Int. J. Biochem. Cell Biol. 2007, 39, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Sharma, S.S.; Mehta, I.K. Free radical scavenging potential of L-proline: Evidence from in vitro assays. Amino Acids 2008, 34, 315–320. [Google Scholar] [CrossRef]
- Loayza-Puch, F.; Rooijers, K.; Buil, L.C.; Zijlstra, J.; Oude, V.J.F.; Lopes, R.; Ugalde, A.P.; van Breugel, P.; Hofland, I.; Wesseling, J.; et al. Tumour-specific proline vulnerability uncovered by differential ribosome codon reading. Nature 2016, 530, 490–494. [Google Scholar] [CrossRef]
- Diehl, F.F.; Vander, H.M.G. Mitochondrial NADPH is a pro at Pro synthesis. Nat. Metab. 2021, 3, 453–455. [Google Scholar] [CrossRef]
- Zhu, J.; Schwörer, S.; Berisa, M.; Kyung, Y.J.; Ryu, K.W.; Yi, J.; Jiang, X.; Cross, J.R.; Thompson, C.B. Mitochondrial NADP(H) generation is essential for proline biosynthesis. Science 2021, 372, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.R.; Levine, R.L.; Stadtman, E.R. Recent advances in the analysis of oxidized proteins. Amino Acids 2003, 25, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Amici, A.; Levine, R.L.; Tsai, L.; Stadtman, E.R. Conversion of amino acid residues in proteins and amino acid homopolymers to carbonyl derivatives by metal-catalyzed oxidation reactions. J. Biol. Chem. 1989, 264, 3341–3346. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.R.; Chao, C.C.; Levine, R.L.; Stadtman, E.R. Glutamic and aminoadipic semialdehydes are the main carbonyl products of metal-catalyzed oxidation of proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Rabe von Pappenheim, F.; Wensien, M.; Ye, J.; Uranga, J.; Irisarri, I.; de Vries, J.; Funk, L.M.; Ricardo, A.M.; Tittmann, K. Widespread occurrence of covalent lysine–cysteine redox switches in proteins. Nat. Chem. Biol. 2002, 18, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Bravard, A.; Vacher, M.; Gouget, B.; Coutant, A.; de Boisferon, F.H.; Marsin, S.; Chevillard, S.; Radicella, J.P. Redox regulation of human OGG1 activity in response to cellular oxidative stress. Mol. Cell Biol. 2006, 26, 7430–7436. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Maayah, M.; Sweasy, J.B.; Alnajjar, K.S. The role of cysteines in the structure and function of OGG1. J. Biol. Chem. 2021, 296, 100093. [Google Scholar] [CrossRef] [PubMed]
- Schulpis, K.H.; Tsakiris, S.; Traeger-Synodinos, J.; Papassotiriou, I. Low total antioxidant status is implicated with high 8-hydroxy-2-deoxyguanosine serum concentrations in phenylketonuria. Clin. Biochem. 2005, 38, 239–242. [Google Scholar] [CrossRef]
- Sitta, A.; Manfredini, V.; Biasi, L.; Treméa, R.; Schwartz, I.V.; Wajner, M.; Vargas, C.R. Evidence that DNA damage is associated to phenylalanine blood levels in leukocytes from phenylketonuric patients. Mutat. Res. 2009, 679, 13–16. [Google Scholar] [CrossRef]
- Sanayama, Y.; Nagasaka, H.; Takayanagi, M.; Ohura, T.; Sakamoto, O.; Ito, T.; Ishige-Wada, M.; Usui, H.; Yoshino, M.; Ohtake, A.; et al. Experimental evidence that phenylalanine is strongly associated to oxidative stress in adolescents and adults with phenylketonuria. Mol. Genet. Metab. 2011, 103, 220–225. [Google Scholar] [CrossRef]
- Rosa, A.P.; Jacques, C.E.; Moraes, T.B.; Wannmacher, C.M.; Dutra Ade, M.; Dutra-Filho, C.S. Phenylpyruvic acid decreases glucose-6-phosphate dehydrogenase activity in rat brain. Cell Mol. Neurobiol. 2012, 32, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Moraes, T.B.; Jacques, C.E.; Rosa, A.P.; Dalazen, G.R.; Terra, M.; Coelho, J.G.; Dutra-Filho, C.S. Role of catalase and superoxide dismutase activities on oxidative stress in the brain of a phenylketonuria animal model and the effect of lipoic acid. Cell Mol. Neurobiol. 2013, 33, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Balluff, B.; Arts, M.; Dubois, L.J.; van Loon, L.J.C.; Hackeng, T.M.; van Eijk, H.M.H.; Eijkel, G.; Heij, L.R.; Soons, Z.; et al. Mass spectrometry imaging of L-[ring-13C6]-labeled phenylalanine and tyrosine kinetics in non-small cell lung carcinoma. Cancer Metab. 2021, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.H.; Wen, G.M.; Song, C.L.; Ji, L.J.; Xia, P. Amino acid profiles in the tissue and serum of patients with liver cancer. Open Med. 2022, 17, 1797–1802. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.M.; Yu, Z.X.; Ferrans, V.J.; Meadows, G.G. Tyrosine and phenylalanine restriction induces G0/G1 cell cycle arrest in murine melanoma in vitro and in vivo. Nutr. Cancer 1997, 29, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.H.; Cheng, M.L.; Lo, C.J.; Lin, Y.H.; Lai, M.W.; Lin, W.R.; Yeh, C.T. Plasma phenylalanine and glutamine concentrations correlate with subsequent hepatocellular carcinoma occurrence in liver cirrhosis patients: An exploratory study. Sci. Rep. 2020, 10, 10926. [Google Scholar] [CrossRef] [PubMed]
- Vettore, L.; Westbrook, R.L.; Tennant, D.A. New aspects of amino acid metabolism in cancer. Br. J. Cancer 2020, 122, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Buehner, G.; Chang, Y.; Harper, J.M.; Sigler, R.; Smith-Wheelock, M. Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance. Aging Cell 2005, 4, 119–125. [Google Scholar] [CrossRef]
- Choi, B.H.; Coloff, J.L. The diverse functions of non-essential amino acids in cancer. Cancers 2019, 11, 675. [Google Scholar] [CrossRef]
- Sedillo, J.C.; Cryns, V.L. Targeting the methionine addiction of cancer. Am. J. Cancer Res. 2022, 12, 2249–2276. [Google Scholar]
- Roci, I.; Watrous, J.D.; Lagerborg, K.A.; Lafranchi, L.; Lindqvist, A.; Jain, M.; Nilsson, R. Mapping metabolic events in the cancer cell cycle reveals arginine catabolism in the committed SG2M phase. Cell Rep. 2019, 26, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.W.; Chi, C.W.; Lin, E.C.; Lui, W.Y.; P’Eng, F.K.; Wang, S.R. Serum arginase level in patients with gastric cancer. J. Clin. Gastroenterol. 1994, 18, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Leu, S.Y.; Wang, S.R. Clinical significance of arginase in colorectal cancer. Cancer 1992, 70, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Sovova, V.; Sloncova, E.; Fric, P. Differences of alkaline phosphatase and arginase activities in human colorectal carcinoma cell lines. Folia Biol. 1997, 43, 101–104. [Google Scholar]
- Chrzanowska, A.; Grabon, W.; Mielczarek-Puta, M.; Baranczyk-Kuzma, A. Significance of arginase determination in body fluids of patients with hepatocellular carcinoma and liver cirrhosis before and after surgical treatment. Clin. Biochem. 2014, 47, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.; Yu, Y.; Li, Z.; Ren, Y.; Li, Z.; Ye, Q.; Liu, P.; Ji, C.; Qian, L.; Xiong, Y. Arginase: An emerging and promising therapeutic target for cancer treatment. Biomed. Pharmacother. 2022, 149, 112840. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.D.; Alejo, S.; Jayamohan, S.; Sareddy, G.R. Lysine-specific demethylase 1 as a therapeutic cancer target: Observations from preclinical study. Expert Opin. Ther. Targets 2023, 24, 1–12. [Google Scholar] [CrossRef]
- Yuta, K.; Yukio, N. Design of cysteine-based self-assembling polymer drugs for anticancer chemotherapy. Colloid. Surf. B 2022, 220, 112909. [Google Scholar]
- Lin, W.; Wang, C.; Liu, G.; Bi, C.; Wang, X.; Zhou, Q.; Jin, H. SLC7A11/xCT in cancer: Biological functions and therapeutic implications. Am. J. Cancer Res. 2020, 10, 3106–3126. [Google Scholar]
- Zhong, W.; Weiss, H.L.; Jayswal, R.D.; Hensley, P.J.; Downes, L.M.; St. Clair, D.K.; Chaiswing, L. Extracellular redox state shift: A novel approach to target prostate cancer invasion. Free Radic. Biol. Med. 2018, 117, 99–109. [Google Scholar] [CrossRef]
- Hanigan, M.H.; Ricketts, W.A. Extracellular glutathione is a source of cysteine for cells that express gamma-glutamyl transpeptidase. Biochemistry 1993, 32, 6302–6306. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. Glutathione, ascorbate, and cellular protection. Cancer Res. 1994, 54, 1969–1975. [Google Scholar]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 49, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohe, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Fujii, J.; Homma, T.; Kobayashi, S. Ferroptosis caused by cysteine insufficiency and oxidative insult. Free Radic. Res. 2020, 54, 969–980. [Google Scholar] [CrossRef]
- Hellmich, M.R.; Coletta, C.; Chao, C.; Szabo, C. The therapeutic potential of cystathionine beta synthetase/hydrogen sulfide inhibition in cancer. Antioxid. Redox Signal 2015, 22, 424–448. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, A.L.; et al. Cystathionine beta-synthase (CBS) contributes to advanced ovarian cancer progression and drug resistance. PLoS ONE 2013, 8, e79167. [Google Scholar] [CrossRef]
- Szabo, C.; Coletta, C.; Chao, C.; Modis, K.; Szczesny, B.; Papapetropoulos, A.; Hellmich, M.R. Tumor-derived hydrogen sulfide, produced by cystathionine-β-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 12474–12479. [Google Scholar] [CrossRef]
- Pupo, E.; Pla, A.F.; Avanzato, D.; Moccia, F.; Cruz, J.E.; Tanzi, F.; Merlino, A.; Mancardi, D.; Munaron, L. Hydrogen sulfide promotes calcium signals and migration in tumor-derived endothelial cells. Free Radic. Biol. Med. 2011, 51, 1765–1773. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Jurkowska, H.; Roman, H.B.; Niewiadomski, J.; Hirschberger, L.L. Insights into taurine synthesis and function based on studies with cysteine dioxygenase (CDO1) knockout mice. Adv. Exp. Med. Biol. 2015, 803, 29–39. [Google Scholar] [PubMed]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xia, Y.; Zhang, X.; Liu, L.; Tu, S.; Zhu, W.; Yu, L.; Wan, H.; Yu, B.; Wan, F. Roles of the MST1-JNK signaling pathway in apoptosis of colorectal cancer cells induced by taurine. Libyan J. Med. 2018, 13, 1500346. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.; Zhang, X.-L.; Wan, H.-F.; Xia, Y.-Q.; Liu, Z.-Q.; Yang, X.-H.; Wan, F.-S. Effect of taurine on cell proliferation and apoptosis human lung cancer A549 cells. Oncol. Lett. 2018, 15, 5473–5480. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lee, H.; Park, K.K.; Kim, H.W.; Park, T. Taurine-responsive genes related to signal transduction as identified by cDNA microarray analyses of HepG2 cells. J. Med. Food. 2006, 9, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Kim, A.K. Effect of taurine on antioxidant enzyme system in B16F10 melanoma cells. Adv. Exp. Med. Biol. 2009, 643, 491–499. [Google Scholar] [PubMed]
- Choi, E.J.; Tang, Y.; Lee, C.B.; Cheong, S.H.; Sung, S.H.; Oh, M.R.; Jang, S.Y.; Park, P.J.; Kim, E.K. Effect of taurine on in vitro migration of MCF-7 and MDA-MB-231 human breast carcinoma cells. Adv. Exp. Med. Biol. 2015, 803, 191–201. [Google Scholar]
- Lennicke, C.; Cocheme, H.M. Redox metabolism: ROS as specific molecular regulators of cell signaling and function. Mol. Cell 2021, 81, 3691–3707. [Google Scholar] [CrossRef]
- Turdo, A.; D’Accardo, C.; Glaviano, A.; Porcelli, G.; Colarossi, C.; Colarossi, L.; Mare, M.; Faldetta, N.; Modica, C.; Pistone, G.; et al. Targeting phosphatases and kinases: How to checkmate cancer. Front. Cell Dev. Biol. 2021, 9, 690306. [Google Scholar] [CrossRef]
- Neel, B.G.; Tonks, N.K. Protein tyrosine phosphatases in signal transduction. Curr. Opin. Cell Biol. 1997, 9, 193–204. [Google Scholar] [CrossRef]
- Zhang, Z.Y. Protein tyrosine phosphatases: Structure and function, substrate specificity, and inhibitor development. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.L.; Wu, S.T.; Yin, M.C.; Chen, H.T.; Chen, H.C. Protective effects from carnosine and histidine on acetaminophen-induced liver injury. J. Food Sci. 2009, 74, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Han, Y.; Kim, D.; Cho, S.; Kim, W.; Hwang, H.; Lee, H.W.; Han, D.H.; Kim, K.S.; Yun, M.; et al. Impact of Exogenous Treatment with Histidine on Hepatocellular Carcinoma Cells. Cancers 2022, 14, 1205. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Bin, H.; Yanping, H.; Cenxin, W.; Huandong, L.; Jinxin, L.; Keyu, S.; Xiaoyu, Z.; Yuanyuan, L.; Zhuoran, Z.; et al. Pyrroline-5-carboxylate reductase 1 reprograms proline metabolism to drive breast cancer stemness under psychological stress. Cell Death Dis. 2023, 14, 682. [Google Scholar]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting autophagy in cancer: Recent advances and future directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, S.A.; Davis, G.E. Differential gene expression in p53-mediated apoptosis-resistant vs. apoptosis-sensitive tumor cell lines. Proc. Natl. Acad. Sci. USA 2000, 97, 13009–13014. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, I.; Ciribilli, Y.; Monti, P.; Bisio, A.; Pollegioni, L.; Fronza, G.; Inga, A.; Campomenosi, P. P53 Family Members Modulate the Expression of PRODH, but Not PRODH2, via Intronic p53 Response Elements. PLoS ONE 2013, 8, e69152. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, G.; Chen, Y.; Xu, W.; Liu, Y.; Ji, G.; Xu, H. Can proline dehydrogenase-a key enzyme involved in proline metabolism-be a novel target for cancer therapy? Front. Oncol. 2023, 13, 1254439. [Google Scholar] [CrossRef]
- Kay, E.J.; Zanivan, S.; Rufini, A. Proline metabolism shapes the tumor microenvironment: From collagen deposition to immune evasion. Curr. Opin. Biotechnol. 2023, 84, 103011. [Google Scholar] [CrossRef]
- Chen, Y.; Kim, J.; Yang, S.; Wang, H.; Wu, C.J.; Sugimoto, H.; LeBleu, V.S.; Kalluri, R. Type I collagen deletion in αSMA(+) myofibroblasts augments immune suppression and accelerates progression of pancreatic cancer. Cancer Cell 2021, 39, 548–565. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Knittel, J.G.; Yan, L.; Rueden, C.T.; White, J.G.; Keely, P.J. Collagen density promotes mammary tumor initiation and progression. BMC Med. 2008, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R. The matrix in cancer. Nat. Rev. Cancer 2021, 11, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Tryptophan metabolism and disposition in cancer biology and immunotherapy. Biosci. Rep. 2022, 42, BSR20221682. [Google Scholar] [CrossRef] [PubMed]
- Shyam Sunder, S.; Sharma, U.C.; Pokharel, S. Adverse effects of tyrosine kinase inhibitors in cancer therapy: Pathophysiology, mechanisms and clinical management. Signal Transduct. Target. Ther. 2023, 8, 262. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).