
Human Skin Drug Metabolism: Relationships between Methyl Salicylate Metabolism and Esterase Activities in IVPT Skin Membranes
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Membrane Preparation
2.3. In Vitro Skin Permeation of Methyl Salicylate from Metsal™ Cream
2.4. Sample Analysis
2.5. Non-Specific Esterase Staining by α-Naphthyl Acetate
2.6. Data and Statistical Analysis
3. Results
3.1. In Vitro Skin Permeation Testing (IVPT) of Metsal™ Cream
3.2. Non-Specific Esterase Staining by α-Napthyl Acetate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Cheruvu, H.S.; Anissimov, Y.G.; van der Hoek, J.; Tsakalozou, E.; Ni, Z.; Ghosh, P.; Grice, J.E.; Roberts, M.S. Percutaneous absorption of steroids from finite doses: Predicting urinary excretion from in vitro skin permeation testing. Int. J. Pharm. 2022, 625, 122095. [Google Scholar] [CrossRef]
- Liu, X.; Anissimov, Y.G.; Grice, J.E.; Cheruvu, H.S.; Ghosh, P.; Raney, S.G.; Maibach, H.I.; Roberts, M.S. Relating transdermal delivery plasma pharmacokinetics with in vitro permeation test (IVPT) findings using diffusion and compartment-in-series models. J. Control. Release 2021, 334, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.E.; Anderson, C.; Roberts, M.S. Topical penetration of commercial salicylate esters and salts using human isolated skin and clinical microdialysis studies. Br. J. Clin. Pharmacol. 1998, 46, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Luu-The, V.; Duche, D.; Ferraris, C.; Meunier, J.R.; Leclaire, J.; Labrie, F. Expression profiles of phases 1 and 2 metabolizing enzymes in human skin and the reconstructed skin models Episkin and full thickness model from Episkin. J. Steroid Biochem. Mol. Biol. 2009, 116, 178–186. [Google Scholar] [CrossRef]
- Zhang, Q.; Grice, J.E.; Wang, G.; Roberts, M.S. Cutaneous metabolism in transdermal drug delivery. Curr. Drug Metab. 2009, 10, 227–235. [Google Scholar] [CrossRef]
- Roberts, M.S.; Cheruvu, H.S.; Mangion, S.E.; Alinaghi, A.; Benson, H.A.E.; Mohammed, Y.; Holmes, A.; van der Hoek, J.; Pastore, M.; Grice, J.E. Topical drug delivery: History, percutaneous absorption, and product development. Adv. Drug Deliv. Rev. 2021, 177, 113929. [Google Scholar] [CrossRef]
- Ahmad, N.; Mukhtar, H. Cytochrome p450: A target for drug development for skin diseases. J. Investig. Dermatol. 2004, 123, 417–425. [Google Scholar] [CrossRef]
- Bucks, D.A. Skin structure and metabolism: Relevance to the design of cutaneous therapeutics. Pharm. Res. 1984, 1, 148–153. [Google Scholar] [CrossRef]
- Kazem, S.; Linssen, E.C.; Gibbs, S. Skin metabolism phase I and phase II enzymes in native and reconstructed human skin: A short review. Drug Discov. Today 2019, 24, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Pyo, S.M.; Maibach, H.I. Skin Metabolism: Relevance of Skin Enzymes for Rational Drug Design. Skin Pharmacol. Physiol. 2019, 32, 283–294. [Google Scholar] [CrossRef]
- Lau, W.M.; Ng, K.W.; Sakenyte, K.; Heard, C.M. Distribution of esterase activity in porcine ear skin, and the effects of freezing and heat separation. Int. J. Pharm. 2012, 433, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Stinchcomb, A.L.; Swaan, P.W.; Ekabo, O.; Harris, K.K.; Browe, J.; Hammell, D.C.; Cooperman, T.A.; Pearsall, M. Straight-chain naltrexone ester prodrugs: Diffusion and concurrent esterase biotransformation in human skin. J. Pharm. Sci. 2002, 91, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Sugibayashi, K.; Hayashi, T.; Morimoto, Y. Simultaneous transport and metabolism of ethyl nicotinate in hairless rat skin after its topical application: The effect of enzyme distribution in skin. J. Control. Release 1999, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Tang-Liu, D.D.; Matsumoto, R.M.; Usansky, J.I. Clinical pharmacokinetics and drug metabolism of tazarotene: A novel topical treatment for acne and psoriasis. Clin. Pharmacokinet. 1999, 37, 273–287. [Google Scholar] [CrossRef]
- Batz, F.M.; Klipper, W.; Korting, H.C.; Henkler, F.; Landsiedel, R.; Luch, A.; von Fritschen, U.; Weindl, G.; Schafer-Korting, M. Esterase activity in excised and reconstructed human skin—Biotransformation of prednicarbate and the model dye fluorescein diacetate. Eur. J. Pharm. Biopharm. 2013, 84, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Sadgrove, M.; Marson, L.; Jay, M. Biotransformation Capacity of Carboxylesterase in Skin and Keratinocytes for the Penta-Ethyl Ester Prodrug of DTPA. Drug Metab. Dispos. 2016, 44, 1313–1318. [Google Scholar] [CrossRef]
- Boehnlein, J.; Sakr, A.; Lichtin, J.L.; Bronaugh, R.L. Characterization of esterase and alcohol dehydrogenase activity in skin. Metabolism of retinyl palmitate to retinol (vitamin A) during percutaneous absorption. Pharm. Res. 1994, 11, 1155–1159. [Google Scholar] [CrossRef]
- Amr, S.; Brown, M.B.; Martin, G.P.; Forbes, B. Activation of clindamycin phosphate by human skin. J. Appl. Microbiol. 2001, 90, 550–554. [Google Scholar] [CrossRef]
- Bailly, J.; Crettaz, M.; Schifflers, M.H.; Marty, J.P. In vitro metabolism by human skin and fibroblasts of retinol, retinal and retinoic acid. Exp. Dermatol. 1998, 7, 27–34. [Google Scholar] [CrossRef]
- Morin, A.; Simard, M.; Rioux, G.; Grenier, A.; Morin, S.; Pouliot, R. Application of an In Vitro Psoriatic Skin Model to Study Cutaneous Metabolization of Tazarotene. Processes 2019, 7, 871. [Google Scholar] [CrossRef]
- Roberts, M.S.; Favretto, W.A.; Meyer, A.; Reckmann, M.; Wongseelashote, T. Topical bioavailability of methyl salicylate. Aust. N. Z. J. Med. 1982, 12, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Bernauer, U.; Bodin, L.; Chaudhry, Q.; Coenraad, P.J.; Dusinska, M.; Ezendam, J.; Gaffet, E.; Galli, C.L.; Granum, B.B.; Panteri, E. Opinion on Methyl Salicylate (Methyl 2- Hydroxybenzoate)-Submission I-SCCS/1633/21-Preliminary Opinion. 2021. Available online: https://health.ec.europa.eu/system/files/2022-08/sccs_o_255.pdf (accessed on 16 July 2022).
- Kligman, A.M.; Christophers, E. Preparation of isolated sheets of human stratum corneum. Arch. Dermatol. 1963, 88, 702–705. [Google Scholar] [CrossRef]
- Jynto. Ball-and-Stick Model of the Methyl Salicylate Molecule. Available online: https://commons.wikimedia.org/wiki/File:Methyl_salicylate_3D_ball.png (accessed on 5 July 2023).
- Mills, B. Ball-and-Stick Model of the Salicylic Acid Molecule. Available online: https://commons.wikimedia.org/wiki/File:Salicylic-acid-from-xtal-2006-3D-balls.png (accessed on 5 July 2023).
- Benjah-bmm27. Methanol-3D-Balls.png. Available online: https://commons.wikimedia.org/wiki/File:Methanol-3D-balls.png (accessed on 5 July 2023).
- Yousef, S.; Mohammed, Y.; Namjoshi, S.; Grice, J.; Sakran, W.; Roberts, M. Mechanistic Evaluation of Hydration Effects on the Human Epidermal Permeation of Salicylate Esters. AAPS J. 2017, 19, 180–190. [Google Scholar] [CrossRef]
- Bajza, A.; Kocsis, D.; Berezvai, O.; Laki, A.J.; Lukacs, B.; Imre, T.; Ivan, K.; Szabo, P.; Erdo, F. Verification of P-Glycoprotein Function at the Dermal Barrier in Diffusion Cells and Dynamic “Skin-On-A-Chip” Microfluidic Device. Pharmaceutics 2020, 12, 804. [Google Scholar] [CrossRef]
- Schafer-Korting, M.; Bock, U.; Gamer, A.; Haberland, A.; Haltner-Ukomadu, E.; Kaca, M.; Kamp, H.; Kietzmann, M.; Korting, H.C.; Krachter, H.U.; et al. Reconstructed human epidermis for skin absorption testing: Results of the German prevalidation study. Altern. Lab. Anim. 2006, 34, 283–294. [Google Scholar] [CrossRef]
- Neupane, R.; Boddu, S.H.S.; Renukuntla, J.; Babu, R.J.; Tiwari, A.K. Alternatives to Biological Skin in Permeation Studies: Current Trends and Possibilities. Pharmaceutics 2020, 12, 152. [Google Scholar] [CrossRef]
- Risueno, I.; Valencia, L.; Jorcano, J.L.; Velasco, D. Skin-on-a-chip models: General overview and future perspectives. APL Bioeng. 2021, 5, 030901. [Google Scholar] [CrossRef] [PubMed]
- Montagna, W. Histology and cytochemistry of human skin. IX. The distribution of non-specific esterases. J. Biophys. Biochem. Cytol. 1955, 1, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Kasper, M.; Surber, C.; Imanidis, G. Permeation, metabolism and site of action concentration of nicotinic acid derivatives in human skin. Correlation with topical pharmacological effect. Eur. J. Pharm. Sci. 2003, 20, 181–195. [Google Scholar] [CrossRef]
- Tokudome, Y.; Katayanagi, M.; Hashimoto, F. Esterase Activity and Intracellular Localization in Reconstructed Human Epidermal Cultured Skin Models. Ann. Dermatol. 2015, 27, 269–274. [Google Scholar] [CrossRef]
- Kitano, Y.; Okada, N. Separation of the epidermal sheet by dispase. Br. J. Dermatol. 1983, 108, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Sano, K.; Yoshizato, K.; Shioya, N.; Sasaki, K. Comparative studies on methods of isolating rat epidermal cells. Ann. Plast. Surg. 1985, 14, 258–266. [Google Scholar] [CrossRef]
- Wei, J.C.J.; Haridass, I.N.; Crichton, M.L.; Mohammed, Y.H.; Meliga, S.C.; Sanchez, W.Y.; Grice, J.E.; Benson, H.A.E.; Roberts, M.S.; Kendall, M.A.F. Space- and time-resolved investigation on diffusion kinetics of human skin following macromolecule delivery by microneedle arrays. Sci. Rep. 2018, 8, 17759. [Google Scholar] [CrossRef] [PubMed]
- Okano, J.; Kojima, H.; Katagi, M.; Nakae, Y.; Terashima, T.; Nakagawa, T.; Kurakane, T.; Okamoto, N.; Morohashi, K.; Maegawa, H.; et al. Epidermis-dermis junction as a novel location for bone marrow-derived cells to reside in response to ionizing radiation. Biochem. Biophys. Res. Commun. 2015, 461, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Sigma-Aldrich. D4693 Dispase® II. Available online: https://www.sigmaaldrich.com/AU/en/product/sigma/d4693 (accessed on 22 June 2023).
- Zou, Y.; Maibach, H.I. Dermal-epidermal separation methods: Research implications. Arch. Dermatol. Res. 2018, 310, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Roberts, M.S. Skin permeability and local tissue concentrations of nonsteroidal anti-inflammatory drugs after topical application. J. Pharmacol. Exp. Ther. 1994, 268, 144–151. [Google Scholar]
- Bronaugh, R.L. Methods for in vitro skin metabolism studies. Toxicol. Mech. Methods 1995, 5, 275–281. [Google Scholar] [CrossRef]





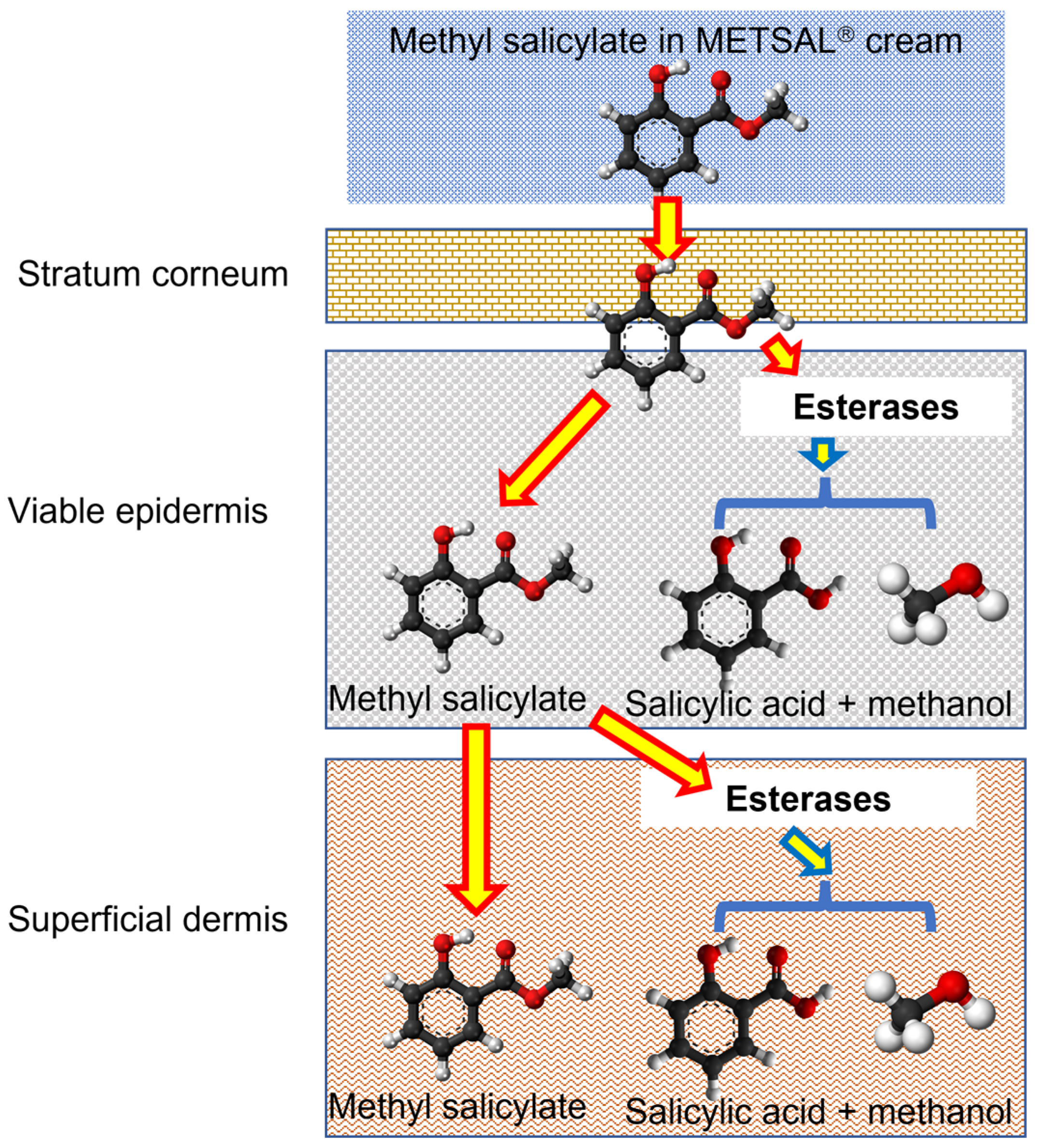{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Type | Applied Methyl Salicylate (mg/cm2) | % As Methyl Salicylate | % as Salicylic Acid | % as Total Actives | Ratio (SA/MeSA) |
|---|---|---|---|---|---|
| Dermis | 25.47 ± 1.36 | 4.44 ± 0.68 | 1.16 ± 0.31 | 5.61 ± 0.74 | 0.26 |
| Dermatomed skin | 26.3 ± 0.75 | 3.59 ± 0.35 a | 0.97 ± 0.25 | 4.56 ± 0.56 b | 0.27 c |
| Dispase-separated epidermis | 27.2 ± 1.28 | 3.88 ± 0.46 a | 0.05 ± 0.004 * | 3.93 ± 0.47 b | 0.013 # |
| Heat-separated epidermis | 28.21 ± 2.23 | 2.88 ± 0.79 a | 0.03 ± 0.01 * | 3.86 ± 0.8 b | 0.010 # |
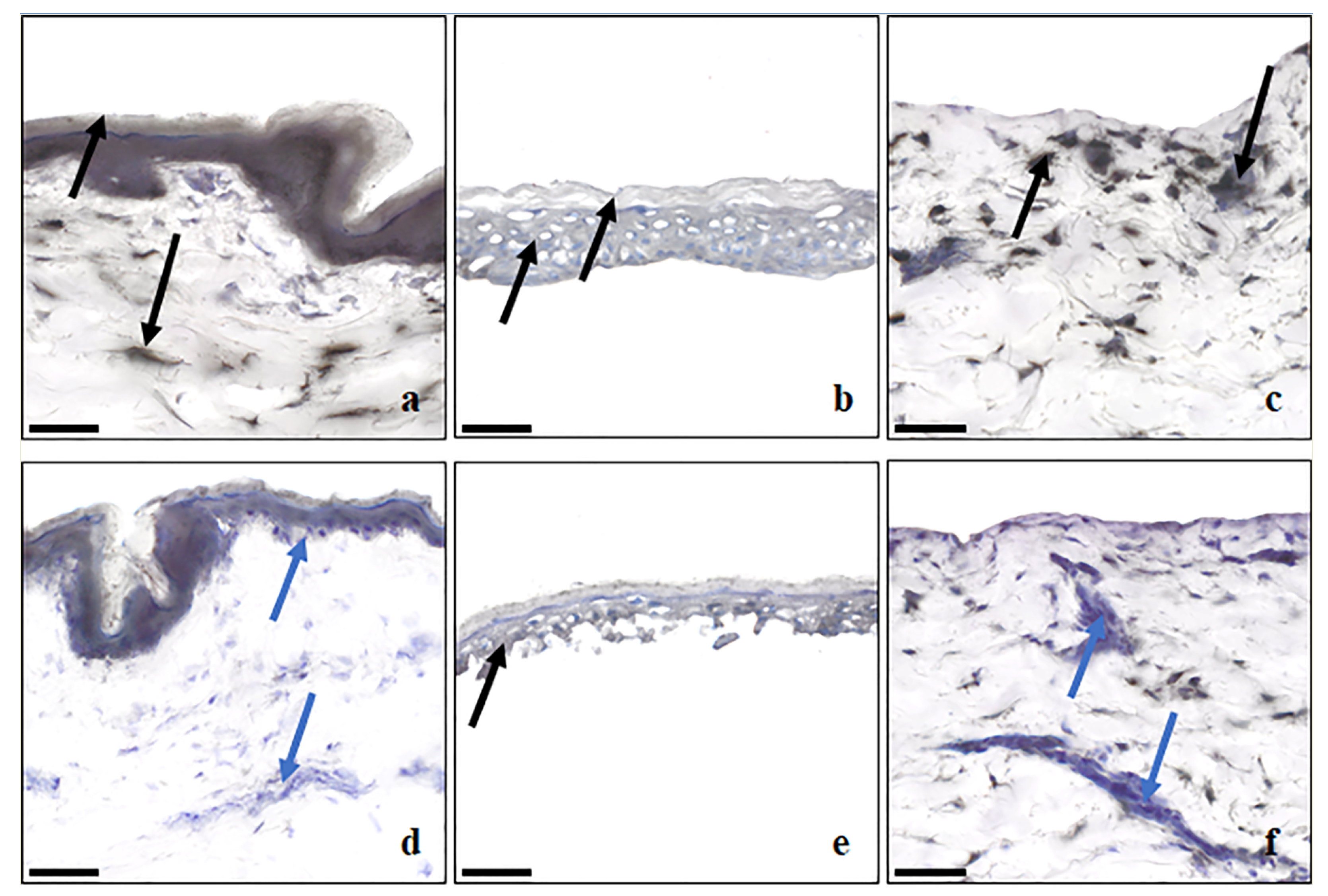
| Membrane Type | Location of Esterase Enzymes |
|---|---|
| Dermatomed Skin | Prescence of esterase staining indicated by black and grey color staining uniform across the epidermal regions and pockets of staining areas across dermis. |
| Dispase-Separated Epidermis | Prescence of esterase staining indicated by uniform grey color staining across SC and VE parts of the membrane. |
| Dermis (remained after dispase-separated epidermis) | Prescence of esterase staining indicated by pockets of black color staining distributed throughout dermis and more towards the dermal epidermal junction. |
| Frozen full thickness skin | Decrease in esterase activity because of freezing indicated by appearance of blue to violet color because of hematoxylin staining in the epidermis regions and more regions in dermis. |
| Heat-Separated epidermis | Decrease in esterase activity because of heat application indicated by appearance of blue to violet color because of hematoxylin staining in the epidermis regions and more regions in dermis. |
| Dermis (remained after HSE separation) | Decrease in esterase activity because of heat application indicated by appearance of blue to violet color because of hematoxylin staining throughout dermis and more towards the dermal epidermal junction. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telaprolu, K.C.; Grice, J.E.; Mohammed, Y.H.; Roberts, M.S. Human Skin Drug Metabolism: Relationships between Methyl Salicylate Metabolism and Esterase Activities in IVPT Skin Membranes. Metabolites 2023, 13, 934. https://doi.org/10.3390/metabo13080934
Telaprolu KC, Grice JE, Mohammed YH, Roberts MS. Human Skin Drug Metabolism: Relationships between Methyl Salicylate Metabolism and Esterase Activities in IVPT Skin Membranes. Metabolites. 2023; 13(8):934. https://doi.org/10.3390/metabo13080934
Chicago/Turabian StyleTelaprolu, Krishna C., Jeffrey E. Grice, Yousuf H. Mohammed, and Michael S. Roberts. 2023. "Human Skin Drug Metabolism: Relationships between Methyl Salicylate Metabolism and Esterase Activities in IVPT Skin Membranes" Metabolites 13, no. 8: 934. https://doi.org/10.3390/metabo13080934
APA StyleTelaprolu, K. C., Grice, J. E., Mohammed, Y. H., & Roberts, M. S. (2023). Human Skin Drug Metabolism: Relationships between Methyl Salicylate Metabolism and Esterase Activities in IVPT Skin Membranes. Metabolites, 13(8), 934. https://doi.org/10.3390/metabo13080934