
Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Fungal Isolation
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Fermentation and Extraction
2.5. Isolation of Compounds 1–3
2.5.1. Panapophenanthrin (1)
2.5.2. Panepophenanthrin (2)
2.5.3. Dihydrohypnophilin (3)
2.6. Antimicrobial Assay
2.7. Cytotoxicity Assay
2.8. Computational Section
3. Results and Discussion
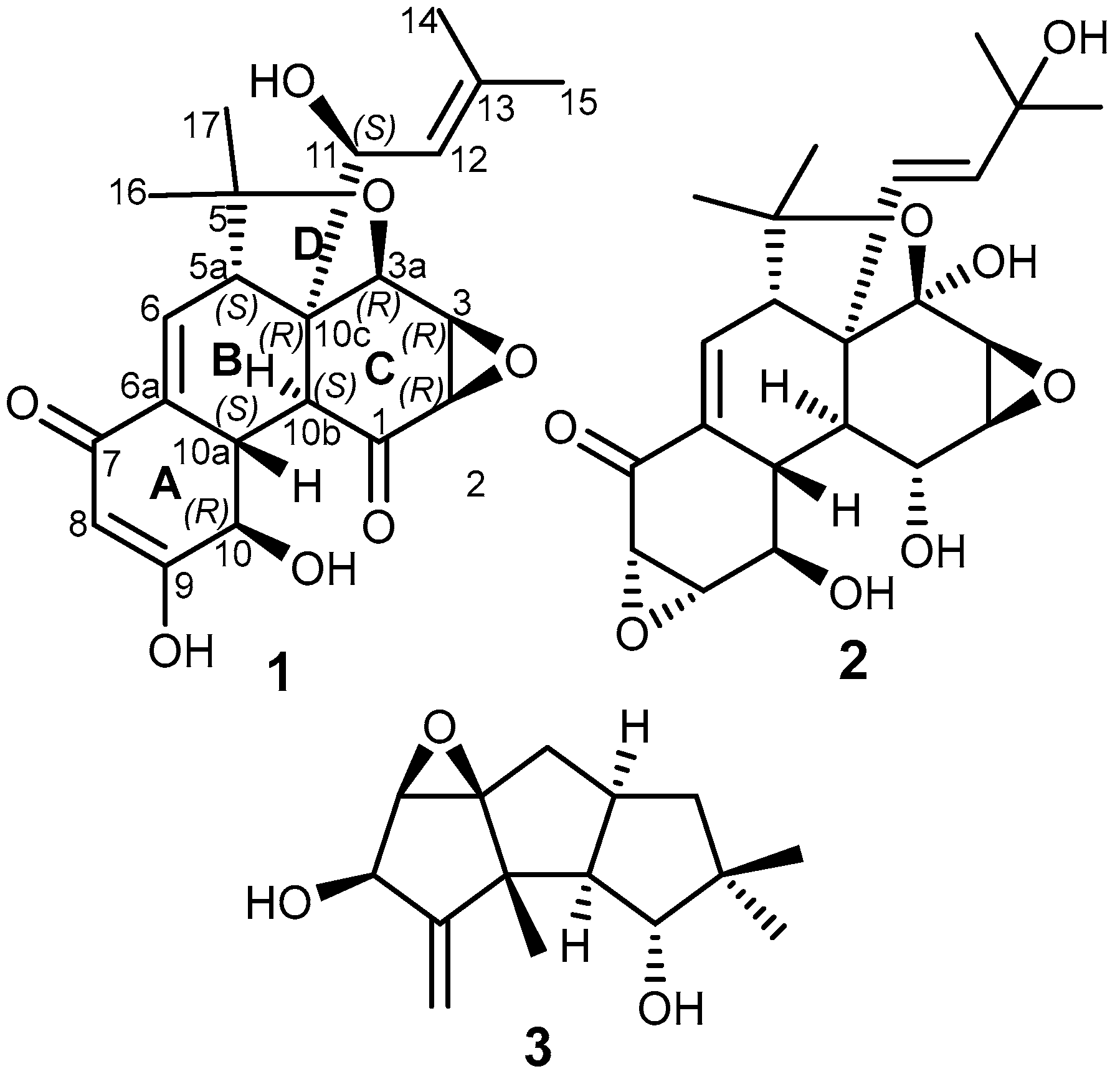
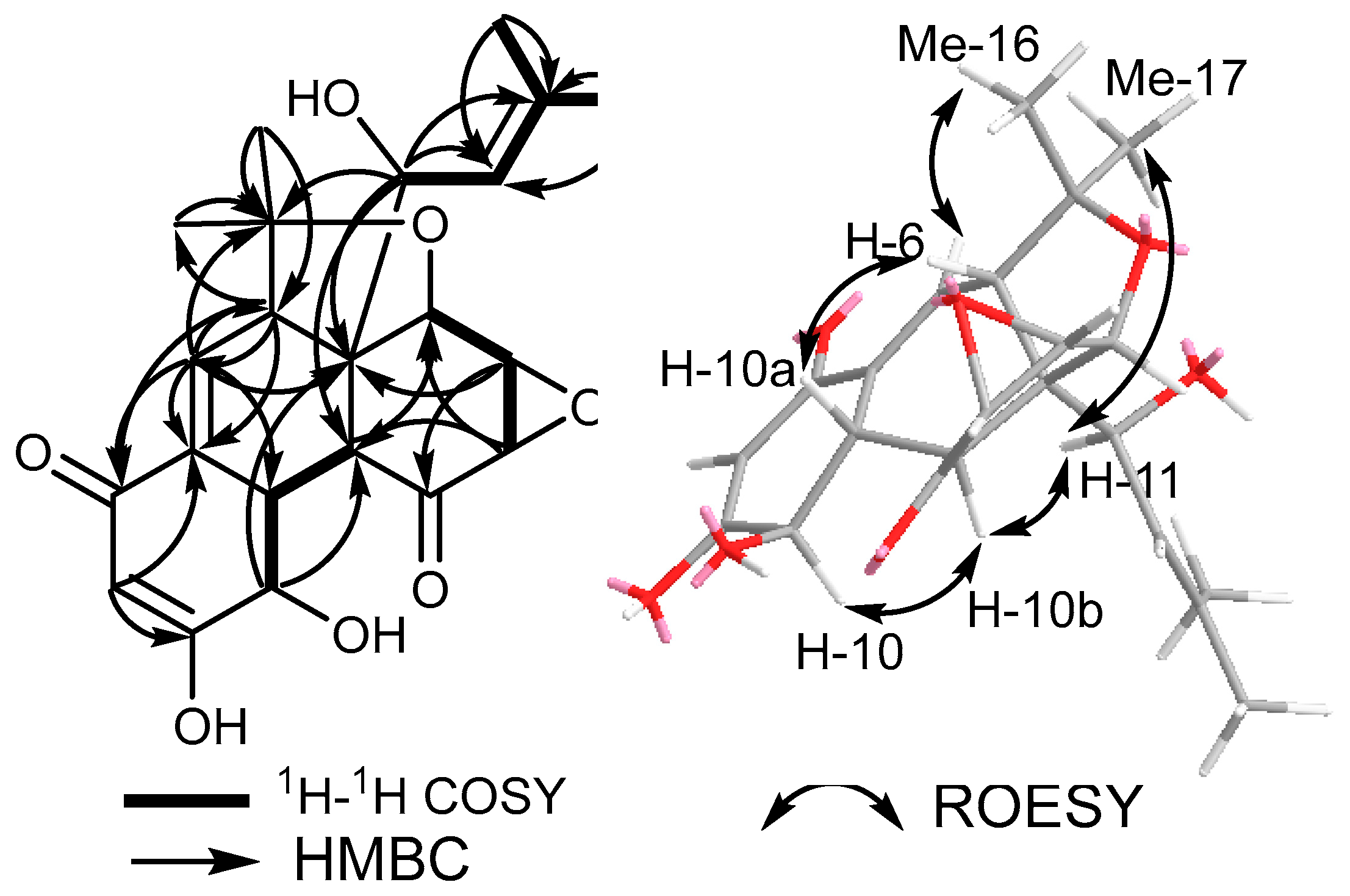
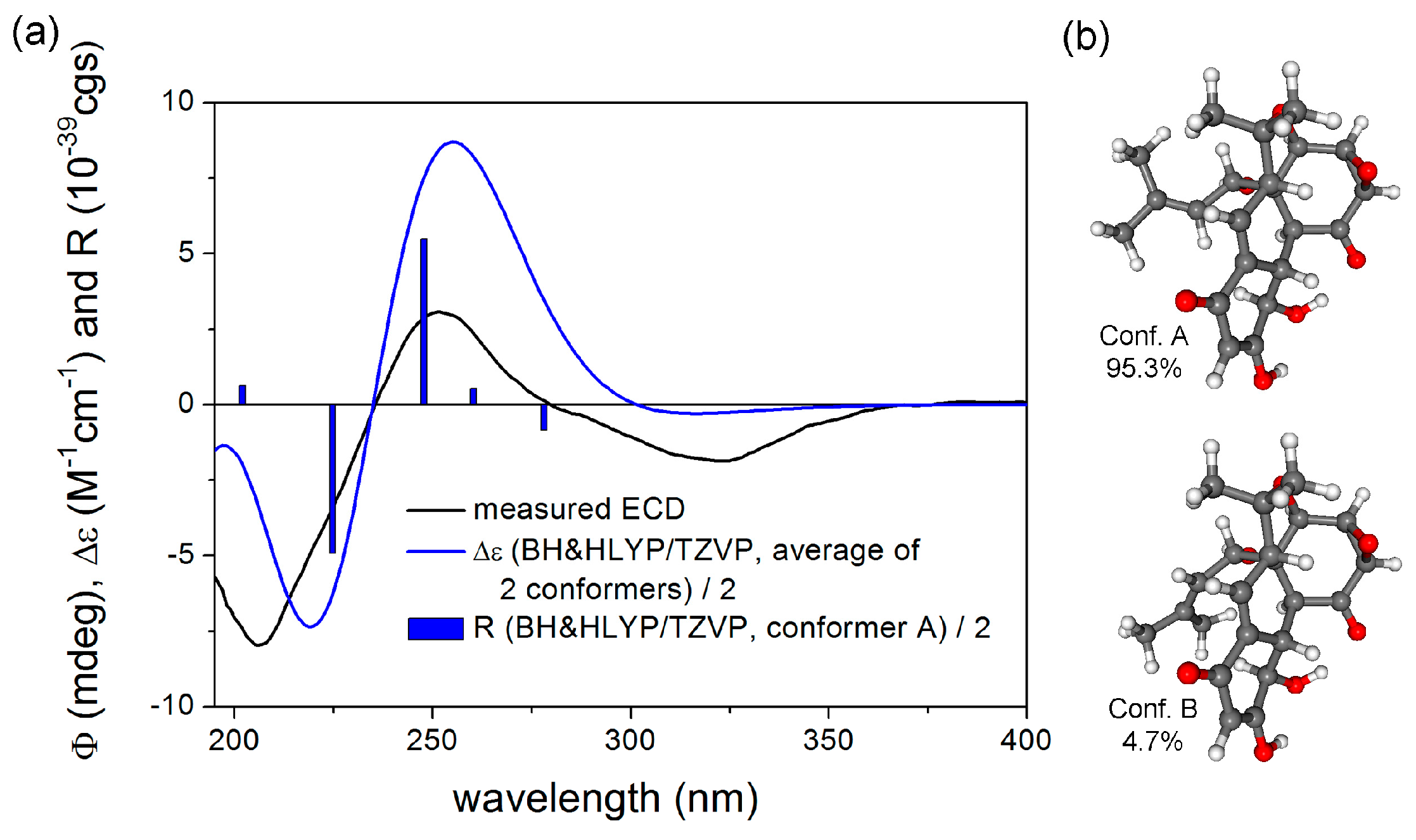
3.1. Isolation and Identification of Compounds (1–3)
3.2. Biological Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Suryadi, H.; Judono, J.J.; Putri, M.R.; Eclessia, A.D.; Ulhaq, J.M.; Agustina, D.N.; Sumiati, T. Biodelignification of lignocellulose using ligninolytic enzymes from white-rot fungi. Heliyon 2022, 8, e08865. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, X.; Zhang, M.; Zhu, Y.; Zhuo, R. Removal of heavy-metal pollutants by white rot fungi: Mechanisms, achievements, and perspectives. J. Clean Prod. 2022, 354, 131681. [Google Scholar] [CrossRef]
- Jaén-Gil, A.; Castellet-Rovira, F.; Llorca, M.; Villagrasa, M.; Sarrà, M.; Rodríguez-Mozaz, S.; Barceló, D. Fungal treatment of metoprolol and its recalcitrant metabolite metoprolol acid in hospital wastewater: Biotransformation, sorption and ecotoxicological impact. Water Res. 2019, 152, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Kristanti, R.A.; Hadibarata, T.; Toyama, T.; Tanaka, Y.; Mori, K. Bioremediation of crude oil by white rot fungi Polyporus sp. S133. J. Microbiol. Biotechnol. 2011, 21, 995–1000. [Google Scholar] [CrossRef]
- Kumar, A.; Yadav, A.N.; Mondal, R.; Kour, D.; Subrahmanyam, G.; Shabnam, A.A.; Khan, S.A.; Yadav, K.K.; Sharma, G.K.; Cabral-Pinto, M.; et al. Myco-Remediation: A mechanistic understanding of contaminants alleviation from natural environment and future prospect. Chemosphere 2021, 284, 131325. [Google Scholar] [CrossRef]
- Wu, J.; Yu, H.-Q. Biosorption of 2,4-dichlorophenol by immobilized white-rot fungus Phanerochaete chrysosporium from aqueous solutions. Bioresour. Technol. 2007, 98, 253–259. [Google Scholar] [CrossRef]
- Corner, E.J.H. The agaric genera Lentinus, Panus, and Pleurotus, with particular reference to Malaysian species. Nova Hedwig. 1981, 69, 1–169. [Google Scholar]
- Vargas-Isla, R.; Capelari, M.; Menolli, N.; Nagasawa, E.; Tokimoto, K.; Ishikawa, N.K. Relationship between Panus lecomtei and P. strigellus inferred from their morphological, molecular and biological characteristics. Mycoscience 2015, 56, 561–571. [Google Scholar] [CrossRef]
- Pegler, D.N. The Genus Lentinus: A World Monograph; Coode, M., Ed.; Kew Bulletin Additional Series X: London, UK, 1983. [Google Scholar]
- Nuñez, M.; Ryvarden, L. Polyporus (Basidiomycotina) and Related Genera. Synop. Fungorum 1995, 10, 1–85. [Google Scholar]
- Grand, E.A. Systematics and Species Concepts in the Genera Lentinus Fr. and Panus Fr., with Emphasis on the Lentinus tigrinus, L. Crinitus and Panus Lecomtei Complexes; Univ. Tennessee Press: Knoxville, TN, USA, 2004. [Google Scholar]
- Hibbett, D.S.; Vilgalys, R. Phylogenetic relationships of Lentinus (basidiomycotina) inferred from molecular and morphological characters. Syst. Bot. 1993, 18, 409–433. [Google Scholar] [CrossRef]
- Seelan, J.S.S.; Justo, A.; Nagy, L.G.; Grand, E.A.; Redhead, S.A.; Hibbett, D. Phylogenetic relationships and morphological evolution in Lentinus, Polyporellus and Neofavolus, emphasizing southeastern Asian taxa. Mycologia 2015, 107, 460–474. [Google Scholar] [CrossRef]
- Senthilarasu, G. The Lentinoid Fungi (Lentinus and Panus) from Western Ghats, India. IMA Fungus 2015, 6, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Cossu, T.A.; Lücking, R.; Vargas-Estupiñán, N.; Carretero, J.; Vasco-Palacios, A.M.; Moncada, B.; Kirk, P.M.; Almeida, R.F.; Gaya, E.; Coca, L.F.; et al. Annotated checklist of fungi of Colombia. In Catalogue of Fungi of Colombia; Kew Publishing, Royal Botanic Gardens: Key, UK, 2022; pp. 209–424. [Google Scholar]
- Prance, G.T. The mycological diet of the Yanomam Indians. Mycologia 1973, 65, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Vasco-Palacios, A.M.; Suaza, S.C.; Castanõ-Betancur, M.; Franco-Molano, A.E. Conocimiento Etnoecólogico de Los Hongos Entre Los Indígenas Uitoto, Muinane y Andoke de La Amazonía Colombiana. Acta Amaz. 2008, 38, 17–30. [Google Scholar] [CrossRef]
- Nandi, S.; Sikder, R.; Acharya, K. Secondary metabolites of mushrooms: A potential source for anticancer therapeutics with translational opportunities. In Advancing Frontiers in Mycology & Mycotechnology; Springer: Singapore, 2019; pp. 563–598. [Google Scholar]
- Song, J.-G.; Ha, L.S.; Ki, D.-W.; Choi, D.-C.; Lee, I.-K.; Yun, B.-S. Chemical constituents of the culture broth of Panus rudis. Mycobiology 2021, 49, 604–606. [Google Scholar] [CrossRef]
- Ding, J.-H.; Li, Z.-H.; Feng, T.; Liu, J.-K. A new cadinane sesquiterpenoid from cultures of the basidiomycete Panus conchatus. Nat. Prod. Res. 2018, 32, 2333–2337. [Google Scholar] [CrossRef]
- Wang, S.-X.; Zhao, R.-L.; Guo, C.; Chen, B.-S.; Dai, H.-Q.; Liu, G.-Q.; Liu, H.-W. New meroterpenoid compounds from the culture of mushroom Panus lecomtei. Chin. J. Nat. Med. 2020, 18, 268–272. [Google Scholar] [CrossRef]
- Erkel, G.; Anke, T.; Sterner, O. Inhibition of NF-ΚB activation by panepoxydone. Biochem. Biophys. Res. Commun. 1996, 226, 214–221. [Google Scholar] [CrossRef]
- Schlegel, B.; Härtl, A.; Dahse, H.-M.; Gollmick, F.A.; Gräfe, U.; Dörfelt, H.; Kappes, B. Hexacylinol, a new antiproliferative metabolite of Panus rudis HKI 0254. J. Antibiot. 2002, 55, 814–817. [Google Scholar] [CrossRef]
- Sekizawa, R.; Ikeno, S.; Nakamura, H.; Naganawa, H.; Matsui, S.; Iinuma, H.; Takeuchi, T. Panepophenanthrin, from a mushroom strain, a novel inhibitor of the ubiquitin-activating enzyme. J. Nat. Prod. 2002, 65, 1491–1493. [Google Scholar] [CrossRef]
- Abate, D.; Abraham, W.-R. Antimicrobial metabolites from Lentinus crinitus. J. Antibiot. 1994, 47, 1348–1350. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Tansakul, C.; Saithong, S.; Pakawatchai, C.; Isaka, M.; Suvannakad, R. Hirsutane Sesquiterpenes from the Fungus Lentinus connatus BCC 8996. J. Nat. Prod. 2005, 68, 1674–1676. [Google Scholar] [CrossRef]
- O’Dell, T.E.; Lodge, D.J.; Mueller, G.M. Approaches to Sampling Macrofungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Elsevier Academic Press: Amsterdam, The Netherland, 2004; pp. 163–168. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Don Pavey, Ed.; Methuen & Co., Ltd.: London, UK, 1978. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes-Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Gelfand, M., Sninsky, J.I., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Charria-Girón, E.; Stchigel, A.M.; Čmoková, A.; Kolařík, M.; Surup, F.; Marin-Felix, Y. Amesia hispanica sp. nov., Producer of the Antifungal Class of Antibiotics Dactylfungins. J. Fungi 2023, 9, 463. [Google Scholar] [CrossRef]
- MacroModel. Schrödinger LLC. Available online: http://www.schrodinger.com/MacroModel (accessed on 10 May 2023).
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision E.01; Gaussian Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Stephens, P.J.; Harada, N. ECD Cotton Effect Approximated by the Gaussian Curve and Other Methods. Chirality 2010, 22, 229–233. [Google Scholar] [CrossRef]
- Varetto, U. MOLEKEL, 5.4; Swiss National Supercomputing Centre: Manno, Switzerland, 2009. [Google Scholar]
- Mándi, A.; Kurtán, T. Applications of OR/ECD/VCD to the Structure Elucidation of Natural Products. Nat. Prod. Rep. 2019, 36, 889–918. [Google Scholar] [CrossRef] [PubMed]
- Superchi, S.; Scafato, P.; Gorecki, M.; Pescitelli, G. Absolute Configuration Determination by Quantum Mechanical Calculation of Chiroptical Spectra: Basics and Applications to Fungal Metabolites. Curr. Med. Chem. 2018, 25, 287–320. [Google Scholar] [CrossRef]
- Chai, J.-D.; Head-Gordon, M. Systematic Optimization of Long-Range Corrected Hybrid Density Functionals. J. Chem. Phys. 2008, 128, 084106. [Google Scholar] [CrossRef] [PubMed]
- Moses, J.E.; Commeiras, L.; Baldwin, J.E.; Adlington, R.M. Total Synthesis of Panepophenanthrin. Org. Lett. 2003, 5, 2987–2988. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Johnson, R.P.; Porco Jr, J.A. Total Synthesis of the Ubiquitin-Activating Enzyme Inhibitor (+)-Panepophenanthrin. Angew. Chem. Int. Ed. 2003, 42, 3913–3917. [Google Scholar] [CrossRef]
- Lohr, N.J.; Molleston, J.P.; Strauss, K.A.; Torres-Martinez, W.; Sherman, E.A.; Squires, R.H.; Rider, N.L.; Chikwava, K.R.; Cummings, O.W.; Morton, D.H.; et al. Human ITCH E3 Ubiquitin Ligase Deficiency Causes Syndromic Multisystem Autoimmune Disease. Am. J. Hum. Genet. 2010, 86, 447–453. [Google Scholar] [CrossRef]
- Duan, S.; Pagano, M. Ubiquitin Ligases in Cancer: Functions and Clinical Potentials. Cell Chem. Biol. 2021, 28, 918–933. [Google Scholar] [CrossRef]
- Mehta, G.; Islam, K. Enantioselective Total Synthesis of Epoxyquinone Natural Products (−)-Phyllostine, (+)-Epoxydon, (+)-Epiepoxydon and (−)-Panepophenanthrin: Access to Versatile Chiral Building Blocks through Enzymatic Kinetic Resolution. Tetrahedron Lett. 2004, 45, 7683–7687. [Google Scholar] [CrossRef]
- Comméiras, L.; Moses, J.E.; Adlington, R.M.; Baldwin, J.E.; Cowley, A.R.; Baker, C.M.; Albrecht, B.; Grant, G.H. Total Synthesis of the Epoxyquinol Dimer (+)-Panepophenanthrin: Application of a Diastereospecific Biomimetic Diels–Alder Dimerisation. Tetrahedron 2006, 62, 9892–9901. [Google Scholar] [CrossRef]
- Cota, B.B.; Rosa, L.H.; Fagundes, E.M.S.; Martins-Filho, O.A.; Correa-Oliveira, R.; Romanha, A.J.; Rosa, C.A.; Zani, C.L. A Potent Trypanocidal Component from the Fungus Lentinus strigosus Inhibits Trypanothione Reductase and Modulates PBMC Proliferation. Mem. Inst. Oswaldo Cruz 2008, 103, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-Q.; Chen, Y.-C.; Sun, Z.-H.; Liu, H.-X.; Li, S.-N.; Li, H.-H.; Gao, X.-X.; Zhang, W.-M. Sesquiterpenoids of Endophytic Fungus Cerrena sp. from Pogostemon cablin and Their Cytotoxic Activities. Zhongguo Zhong Yao Za Zhi 2017, 42, 1693–1698. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pos. | δH (Multi, J [Hz]) a | δC, Type b,e | δH (Multi, J [Hz]) c | δC, Type d,e |
|---|---|---|---|---|
| 1 | 201.2, CO | 202.9, CO | ||
| 2 | 5.13 (br s, 1H) | 64.0, CH | 5.17 (br s, 1H) | 64.5, CH |
| 3 | 3.58 (m, J = 1.6 Hz, 1H) | 55.7, CH | 3.50 (m, overlapped, 1H) | 57.8, CH |
| 3a | 3.66 (dd, J = 3.6, 1.3 Hz, 1H) | 60.4, CH | 3.61 (dd, J = 3.8, 1.2 Hz, 1H) | 62.5, CH |
| 5 | 81.3, C | 83.0, C | ||
| 5a | 3.52 (t, J = 3.7 Hz, 1H) | 50.3, CH | 3.50 (t, J = 3.2 Hz, 1H) | 50.0, CH |
| 6 | 6.52 (dd, J = 4.1, 2.8 Hz, 1H) | 134.4, CH | 6.31 (dd, J = 4.8, 2.8 Hz, 1H) | 133.2, CH |
| 6a | 133.3, C | 136.6, C | ||
| 7 | 193.9, CO | 198.0, CO | ||
| 8 | 8.08 (d, J = 2.0 Hz, 1H) | 129.5, CH | 8.56 (s, 1H) | 130.9, CH |
| 9 | 165.2, C | 170.3, C | ||
| 10 | 4.29 (t, J = 5.7 Hz, 1H) | 68.6, CH | 4.12 (d, J = 5.7 Hz, 1H) | 70.5, CH |
| 10a | 2.65 (ddd, J = 11.0, 5.7, 2.8 Hz, 1H) | 44.8, CH | 2.59 (ddt, J = 11.2, 5.7, 2.8 Hz, 1H) | 45.9, CH |
| 10b | 2.38 (dd, J = 11.0, 1.6 Hz, 1H) | 47.5, CH | 2.46 (dt, J = 11.2, 1.6 Hz, 1H) | 48.4, CH |
| 10c | 59.6, C | 61.5, C | ||
| 11 | 5.10 (d, J = 10.0 Hz, 1H) | 80.3, CH | 5.56 (d, J = 9.9 Hz, 1H) | 80.8, CH |
| 12 | 5.03 (dt, J = 10.0, 1.4 Hz, 1H) | 120.0, CH | 4.98 (dt, J = 9.9, 1.4 Hz, 1H) | 123.1, CH |
| 13 | 142.8, C | 140.0, C | ||
| 14 | 1.78 (d, J = 1.4 Hz, 3H) | 18.5, CH3 | 1.76 (d, J = 1.4 Hz, 3H) | 19.1, CH3 |
| 15 | 1.76 (d, J = 1.4 Hz, 3H) | 26.4, CH3 | 1.71 (d, J = 1.4 Hz, 3H) | 26.6, CH3 |
| 16 | 1.48 (s, 3H) | 28.3, CH3 | 1.42 (s, 3H) | 30.2, CH3 |
| 17 | 1.09 (s, 3H) | 23.7, CH3 | 1.06 (s, 3H) | 25.9, CH3 |
| MIC (µg/mL) | ||||
|---|---|---|---|---|
| Test Organism | 1 | 2 | 3 | Reference |
| Schizosaccharomyces pombe | n.t | - | - | 4.2 N |
| Pichia anomala | n.t | - | - | 8.3 N |
| Mucor hiemalis | - | - | - | 4.2 N |
| Candida albicans | n.t | - | - | 4.2 N |
| Rhodotorula glutinis | n.t | - | - | 2.1 N |
| Acinetobacter baumanii | n.t | - | - | 0.52 C |
| Escherichia coli | - | - | - | 0.83 G |
| Bacillus subtilis | 33.3 | - | 66.6 | 16.6 O |
| Mycobacterium smegmatis | n.t | - | - | 1.7 K |
| Staphylococcus aureus | 66.6 | - | - | 0.21 G |
| Pseudomonas aeruginosa | - | - | - | 0.21 G |
| Chromobacterium violaceum | n.t | - | - | 0.83 G |
| IC50 (µM) | ||||
|---|---|---|---|---|
| Cell Lines | 1 | 2 | 3 | Epothilone B |
| L929 | 13.2 | * | 103.9 | 6.5 × 10−4 |
| KB3.1 | 17.9 | * | ** | 1.73 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llanos-López, N.A.; Ebada, S.S.; Vasco-Palacios, A.M.; Sánchez-Giraldo, L.M.; López, L.; Rojas, L.F.; Mándi, A.; Kurtán, T.; Marin-Felix, Y. Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus. Metabolites 2023, 13, 848. https://doi.org/10.3390/metabo13070848
Llanos-López NA, Ebada SS, Vasco-Palacios AM, Sánchez-Giraldo LM, López L, Rojas LF, Mándi A, Kurtán T, Marin-Felix Y. Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus. Metabolites. 2023; 13(7):848. https://doi.org/10.3390/metabo13070848
Chicago/Turabian StyleLlanos-López, Natalia A., Sherif Saeed Ebada, Aída M. Vasco-Palacios, Laura M. Sánchez-Giraldo, Lina López, Luisa F. Rojas, Attila Mándi, Tibor Kurtán, and Yasmina Marin-Felix. 2023. "Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus" Metabolites 13, no. 7: 848. https://doi.org/10.3390/metabo13070848
APA StyleLlanos-López, N. A., Ebada, S. S., Vasco-Palacios, A. M., Sánchez-Giraldo, L. M., López, L., Rojas, L. F., Mándi, A., Kurtán, T., & Marin-Felix, Y. (2023). Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus. Metabolites, 13(7), 848. https://doi.org/10.3390/metabo13070848